Embed Size (px)
Citation preview

The ultimate goal of cancer research is to learn how to make cancer cells selectively die. Over the past two decades it has become clear that cell death, in both malignant and non-malignant cells, can be an active, regulated programme. Many different types of cell death have been identified. Although it is not yet clear to what extent each describes a distinct process, among the terms used to describe different mechanisms of death are apoptosis (which has both an extrinsic and an intrinsic, or mitochondrial, pathway), necrosis, mitotic catastrophe and autophagy. Of these, apoptosis is the type of death about whose genetically controlled pathways we know the most.
It is likely that a significant portion of the cancer cell death following chemotherapy involves necrosis or mitotic catastrophe. Death following mitotic catastrophe, however, might be simply another means of entry into the apoptotic pathway1. Often when I discuss the apop-totic pathway in the context of individual cancers I am asked, “But surely other death pathways are ‘involved’ in that type of cancer. Don’t you need to consider those?” To some extent, the answer is “of course” — more informa-tion cannot help but be interesting, perhaps even useful. But our goal need not be a complete understanding of cancer biology to learn how to kill cancer cells. We need to remember that for a cancer cell to survive, it must ensure that none of the death pathways are activated. To put it another way, those of us in the business of killing cancer cells need ensure only that one of the death path-ways gets activated. The pathway I have chosen to study most carefully is the intrinsic, or mitochondrial, pathway of apoptosis. The control of that pathway in cancer will be the subject of this Review.
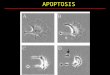
Defining apoptosisInterest in apoptosis was greatly enhanced when it was recognized that a genetically controlled programme governed commitment to and execution of apoptosis in a wide range of multicellular organisms spanning worms, flies and humans2–4. Over the past two decades an impressive amount of research has resulted in a bet-ter understanding of the genes and biochemical events responsible for the process of apoptosis, including the morphological changes that first alerted researchers to this form of cell death (FIG. 1).
Cells destined for apoptosis are at best useless, and at worst harmful to the organism. For the benefit of the organism as a whole, it is important that once a cell is no longer useful the commitment and execution of its death be rapid and irreversible. Otherwise, the organism might have to endure ailing, useless and deleterious cells using resources that could be more efficiently expended. The combined processes of nucleosomal fragmentation by endonucleases, widespread proteolysis by caspases and cell surface tagging to induce phagocytosis are the key elements of apoptosis. Thus a cell that is commit-ted to undergo programmed cell death (PCD) by the intrinsic apoptotic pathway is rapidly converted to a metabolically inactive remnant that cannot replicate its DNA. It is engulfed by phagocytes to prevent poten-tially harmful exposure of intracellular antigens to the immune system.
The crucial event that commits a cell to death by the intrinsic apoptotic pathway is permeabilization of the outer mitochondrial membrane with the subsequent release of multiple pro-apoptotic factors that direct the physiological changes described above5. Although
Dana–Farber Cancer Institute, Harvard Medical School, Dana 530B, 44 Binney Street, Boston, Massachusetts 02052, USA. e-mail: [email protected]:10.1038/nrc2297Published online 18 January 2008
Extrinsic apoptotic pathwayThe apoptotic pathway initiated by ligation of the tumour necrosis factor family of receptors. Following ligation, a death-inducing signalling complex is constructed, which induces activation of the initiator caspase 8, which in turn activates effector caspases such as caspase 3. This pathway can connect with the intrinsic pathway through caspase 8 cleavage and activation of the pro-death BCL2 family protein BID.
Intrinsic or mitochondrial pathwayThe apoptotic pathway controlled by the BCL2 family that results in mitochondrial permeabilization by activated BAX and BAK. Multiple pro-apoptotic factors are released from the permeabilized mitochondria, including cytochrome c and SMAC, which induce caspase activation.
Diagnosing and exploiting cancer’s addiction to blocks in apoptosisAnthony G. Letai
Abstract | Cancer cells survive despite violating rules of normal cellular behaviour that ordinarily provoke apoptosis. The blocks in apoptosis that keep cancer cells alive are therefore attractive candidates for targeted therapies. Recent studies have significantly increased our understanding of how interactions among proteins in the BCL2 family determine cell survival or death. It is now possible to systematically determine how individual cancers escape apoptosis. Such a determination can help predict not only whether cells are likely to be killed by antagonism of BCL2, but also whether they are likely to be sensitive to chemotherapy that kills by the intrinsic apoptotic pathway.
R E V I E W S
NATuRe RevIeWs | cancer vOluMe 8 | FeBRuARy 2008 | 121
© 2008 Nature Publishing Group

NecrosisA type of cell death distinct from apoptosis, characterized by breakdown in the plasma membrane and release of intracellular contents. Though previously thought to be a passive process, it might also be regulated by a genetically encoded programme.
Mitotic catastropheA death resulting from failure of a cell to arrest before mitosis following DNA damage, resulting in severe aberrancies in chromosomal structure and segregation. It might share downstream events with apoptosis.
AutophagyLiterally, a cellular response in which the cell metabolizes its own contents and organelles to maintain energy production. Although such a process can eventually result in cell death, it can also be used to maintain cell survival under conditions of limiting nutrients.
important mechanisms exist for the modulation of specific events following mitochondrial outer mem-brane permeabilization (MOMP), MOMP for most pur-poses may be considered the point at which the cell is irreversibly committed to death by apoptosis. Although some have found the ability to rescue cells following MOMP ex vivo in the absence of phagocytosis, this rescue relies on manipulations, such as caspase inhibi-tion, that are not normally available to cells in vivo6. As the major thrust of cancer therapy is to get the cancer cell to commit to cell death, the events surrounding this decision are of paramount importance. Therefore, I will consider in this discussion primarily the molecules and interactions that control MOMP. MOMP in apoptosis is controlled primarily by the BCl2 family of proteins, which contains the key adjudicators of the cell’s decision to commit suicide by apoptosis, as well as the crucial effectors of MOMP itself.
The BCL2 family of proteinsThe BCL2 gene was initially cloned from the breakpoint of the t(14;18) chromosomal translocation found in the vast majority of follicular lymphomas, an indolent B-cell non-Hodgkin lymphoma7–9. expression of BCl2 was shown to block cell death following numerous cel-lular insults10. As a test of its ability to function as an oncogene, a minigene bearing the Bcl2–immunoglobulin gene fusion was shown, after a period of follicular hyperplasia10, to induce lymphoma in transgenic mice11. Although previously characterized oncogenes had shared the ability to increase cellular proliferation, BCL2 founded a new class of oncogene — inhibitors of PCD.
In the decade following the cloning of BCL2, an entire family of proteins that are related by sequence homol-ogy and participation in the control of apoptosis was identified. some of these proteins share with BCl2 the ability to oppose PCD: BCl-Xl (also known as BCl2-like 1 (BCl2l1))12, BCl-w (also known as BCl2l2)13, myeloid cell leukemia sequence 1 (MCl1)14 and BFl1 (also known as BCl2A1)15. These proteins have sequence homology in four α-helical BCl2 homology (BH) regions, BH1–4. BCl2-associated X protein (BAX)16, BCl2-antagonist/killer (BAK)17 and BCl2-related ovar-ian killer (BOK)18, sharing only the BH1–3 domains, promote cell death. A third class of proteins was later established19. These proteins, including BH3-interacting domain death agonist (BID), BCl2 antagonist of cell death (BAD), BCl2-interacting killer (BIK), PuMA (also known as BCl2-binding component 3 (BBC3)), NOXA (also known as phorbol-12-myristate-13-acetate-induced protein 1 (PMAIP1)), Bcl2-modifying factor (BMF) and HRK, demonstrate homology only in the BH3 region, and are often referred to as BH3-only proteins20,21. like BAX and BAK, the pro-apoptotic BH3-only proteins require an intact BH3 domain to promote apoptosis17,20 (FIG. 2).
Death at the mitochondria. The BCl2 family of proteins governs the commitment to PCD at the mitochondrion — the intrinsic apoptotic pathway22,23. Myriad death sig-nals, including DNA damage, growth-factor deprivation, oncogene activation, cell signalling pathway perturba-tion and microtubule disruption, are communicated through the intrinsic apoptotic pathway. In response to the initial insult, incompletely understood signalling events are eventually converted into active BH3-only family members, which may be considered the cellular sentinels of PCD24. These BH3-only family members become active through many mechanisms, including increased transcription, protein stabilization and post-translational modification25–28. Once active, certain BH3-only proteins, including BIM (also known as BCl2l11) and BID, induce the activation of BAX and BAK29–34,51. BAX and BAK are absolutely essential for death induced by BH3-proteins. BOK expression is restricted to repro-ductive tissues, so deletion of BAX and BAK provides a profound inhibition of apoptosis in most tissues18,35,36.
We term BH3-only proteins that have the ability to activate BAX and BAK activators34 (FIG. 3). Notably, there is no reason to think that BID and BIM are the only activators of BAX and BAK. experiments addressing the properties of the isolated BH3 domain of PuMA have indicated that this domain cannot function as an activa-tor32,34. However, studies using the whole protein suggest that PuMA may indeed act in this way29,37, and in one study PuMA has been shown to directly bind BAX in a manner dependent on its BH3 domain38. Therefore, PuMA might be an example of a molecule in which the BH3 domain gains additional properties in the context of the entire protein.
As only three amino acids out of roughly twenty in the BH3 domain are highly conserved across the family, it is not possible, even with a complete knowledge of the
At a glance
• Cancer cells exhibit many phenotypes, such as genomic instability or oncogene activation, that ought to induce apoptosis, but they nonetheless survive. A block in apoptosis is a likely requirement for cancer maintenance.
• In cancer cells that overexpress BCL2 the protein itself is often largely bound to pro-apoptotic BH3-only proteins like BIM. In such circumstances we describe BCL2 and the cell as being ‘primed’.
• BH3 profiling is a novel tool that exploits selective interaction between BH3 domains and anti-apoptotic BCL2 proteins to reveal the different ways cancer cells escape apoptosis. Certain cancer cells escape apoptosis by expression of BCL2; BH3 profiling can specifically identify these primed, BCL2-dependent cells.
• BCL2 expression does not necessarily confer a chemoresistant phenotype to cancer cells when selected for in previously untreated cells. If the expressed BCL2 is primed, sequestering large amounts of pro-apoptotic proteins such as BIM, it may actually relate to increased chemosensitivity.
• Primed cells are selectively sensitive to antagonists of anti-apoptotic BCL2 proteins like ABT-737. Primed cells might also be selectively more sensitive to conventional chemotherapy agents compared with cancer cells that use a different apoptotic block.
• It has long been suspected that cancer cells are more susceptible to cell death than normal cells. That some cancer cells appear to be primed for death in comparison with normal cells offers a possible biochemical explanation for this clinical observation.
• Small-molecule drugs that target BCL2 and related anti-apoptotic proteins are currently in early-phase clinical trials.
R E V I E W S
122 | FeBRuARy 2008 | vOluMe 8 www.nature.com/reviews/cancer
© 2008 Nature Publishing Group

Nature Reviews | Cancer
Macrophage
Cytochrome cSMAC
Apoptotic cell
human genome, to identify by informatic means proteins that possess functional BH3 domains. Indeed, new BH3 domain-containing proteins continue to be discovered. Furthermore, it is possible that molecules that do not contain a BH3 domain at all can activate BAX and BAK. For instance, the tumour suppressor p53, which does not contain a BH3 domain, has been shown to activate BAX and BAK39–41. Interestingly, BAX and BAK might be sensitive to cues that are not encoded by proteins. For instance, in cell lines in vitro heat has been found to activate BAX and BAK42.
Before its activation, BAX exists in two main pools43,44. One is in the cytosol, where it may be complexed with other proteins that regulate its function. The second is at the mitochondrion, where BAX maintains a loose, alkali-labile attachment. On activation, cytosolic BAX translocates to the mitochondrion, joining BAK and that proportion of BAX that was already loosely associ-ated with the mitochondrion. Activated BAX and BAK undergo an allosteric change and expose their amino ter-minus, a change that can be recognized by conformation-specific antibodies43. This same change can be induced artefactually by interaction with certain commonly used detergents, including Triton X-100 and NP-40. This conformational change also induces interaction of BAX or BAK with anti-apoptotic proteins such as BCl2. Therefore, one must interpret the finding of complexes
of BCl2 with BAX or BAK in buffers containing NP-40 or Triton X-100 with great caution. A preferred method to solubilize BCl2 family proteins from cellular lysates is to use a detergent such as 3-[(3-cholamidopropyl)di-methylammonio]-1-propanesulphonate (CHAPs), which does not artefactually induce BAX or BAK confor-mational change. However, complexes of BAX and BAK with anti-apoptotic proteins have been observed even in lysates made with CHAPs45,46, but generally only a small portion of the total cellular BAX or BAK is involved. How readily complexes of BAX and BAK with anti-apoptotic proteins can be observed seems to depend greatly on the cell being studied.
For BAX, the conformational change accompanies alkali-stable insertion into the mitochondrial membrane. BAX and BAK then undergo homo-oligomerization. There are reports that oligomerization of either can coop-erate to facilitate oligomerization of the other47,48. BAX and BAK oligomers then participate in the formation of pores in the outer mitochondrial membrane inducing MOMP and the release of cytochrome c, sMAC (also known as DIABlO), endonuclease G and other pro-apoptotic proteins from the mitochondrion5,29,31–34,49–51. either BAX or BAK alone is sufficient to perform this function — both are not necessary36. This pore has not been completely characterized, and might include other proteins in addition to BAX and BAK52–54. It has been shown, however, that BAX oligomers by themselves can form channels in liposomes that permit the release of cytochrome c and other macromolecules54. MOMP may be considered the point at which the cell is committed to PCD, and is associated with the activation of caspases5,55 (BOX 1). loss of BAX and BAK completely blocks MOMP through this pathway36,58,59.
Although execution and modulation of the death sig-nals downstream of the commitment that is conferred by MOMP is important and interesting, I will not consider it further due to space constraints. Rather, I will focus on the upstream decision, controlled by the BCl2 family, of whether to commit to PCD.
Modulating death at the mitochondria. Anti-apoptotic BCl2 and related proteins BCl-Xl, MCl1, BCl-w and BFl1 interrupt the PCD pathway upstream of BAX and BAK oligomerization and the commitment to PCD34,58,60. Although the anti-apoptotic proteins have the ability to bind BAX and BAK, particularly in their activated forms, it seems that the most important mechanism of their anti-apoptotic function might be the binding and sequestration of activator BH3-only molecules before they can interact with and activate BAX or BAK30,34,58,60 (FIG. 3). The anti-apoptotic family members bind the hydrophobic face of the amphipathic α-helical BH3 domain of pro-apoptotic proteins in their hydrophobic groove, which is formed by the α-helical BH1, BH2 and BH3 domains60–64.
A subset of BH3-only proteins, known as sensitiz-ers, lack the ability to induce BAX and BAK activation directly34. Instead, they compete for binding in the hydrophobic BH3-binding cleft of anti-apoptotic pro-teins30,32,34,37. In so doing, they behave as antagonists of the anti-apoptotic proteins, displacing activators that
Figure 1 |Howapoptosiskillsandclearscells.Apoptosis can be described morphologically as causing cell shrinkage, convolution and blebbing of the cell surface, and marked condensation of the nucleus93. Inversion of the plasma membrane occurs so that the phosphatidyl serine that is usually restricted to the cytoplasmic face of the inner leaflet is exposed to the extracellular environment. In vivo, the externalized phosphatidyl serine and other induced signals allow recognition by phagocytic cells, which phagocytose apoptotic cells56,57,94. Phagocytosis can rapidly remove apoptotic cells from their environment, which can contribute to their underestimation when investigated in vivo. The entire programme, once initiated, can be completed on a timescale of minutes to a few hours.
R E V I E W S
NATuRe RevIeWs | cancer vOluMe 8 | FeBRuARy 2008 | 123
© 2008 Nature Publishing Group

BH4 BH3 BH1 BH2 TM
BH3
BH3 BH1 BH2
Multidomain anti-apoptotic
Multidomain pro-apoptotic
BH3-only pro-apoptoticActivators:BID, BIM and PUMA(?)
Sensitizers:BAD, BIK, NOXA,BMF and HRK
Nature Reviews | Cancer
BCL2, BCL-XL, MCL1, BCL-w and BFL1
BAX, BAK and BOK TM
might be already bound or binding free anti-apoptotic proteins so that their binding sites are no longer avail-able to sequester activator proteins. A key property of the sensitizers is that they do not interact equivalently with all of the anti-apoptotic proteins. Instead, each anti-apoptotic protein interacts with a subset of the sensitizer BH3-only proteins30,32,65. Below, we show how this selectivity pattern can be exploited to develop an important tool, a tool that can provide a summary interpretation of the participation of all of the BCl2 family members to reveal how ‘primed’ a cell is for death, and what anti-apoptotic protein(s) are required for the ongoing survival of the cell in question. We call this strategy BH3 profiling (BOX 2).
An alternative model of activation of BAX and BAK. An alternative, ‘indirect’ model holds that direct activation of BAX and BAK does not take place65–68. Rather, BAX and BAK are continually kept at bay by anti-apoptotic proteins such as BCl2 and MCl1. Inhibition of all of the anti-apoptotic proteins is necessary and sufficient to cause commitment to PCD. yet even proponents of this model acknowledge that in order to execute MOMP, some change in BAX and BAK must take place68. The changed BAX or BAK is referred to as ‘primed’ rather than activated, but the concept is the same. left unan-swered in this model is the question of what events are responsible for priming or activating BAX and BAK.
Proponents of the indirect model cite as support the interesting observation that activation of BAX and death consistent with apoptosis can occur even in the absence of BID and BIM68. However, as explained above, there is no reason to believe these are the only activators. The
same proponents also point out that if BIM and BID truly interact with BAX and BAK, it should be easier to observe complexes of these proteins by immunoblot. such com-plexes are indeed difficult to observe, but they have been reported20,29,38,50,69. The difficulty in observing them might be due to the transient quality of their interaction: it is hypothesized that the interaction of activators with BAX or BAK is a ‘hit-and-run’ catalytic interaction in which the transient complex comes apart following the conforma-tional change induced in BAX or BAK. It should also be remembered that BID, the first protein to be identified as an activator, was originally cloned through its interaction with BAX as well as BCl2 (ReF. 20).
Proponents, like me, of the activator–sensitizer model point to several key findings. In a simplified liposomal system, it has been clearly shown that recombinant BAX is unable to induce liposomal permeabilization without the presence of an activator like BID or BIM32,33. Definite biochemical measurements can be made of a stable inter-action between BAX and a stabilized BID or BIM BH3 peptide70. In certain systems, at least, inhibition of death by BCl-Xl does not depend on binding BAX or BAK, but it does depend on binding BH3-only proteins58,60.
A position that can accommodate much of this data is that BID and BIM can and do activate BAX and BAK under many circumstances71. In other circumstances, other activators, proteins or otherwise may fulfil this role. BAX and BAK might also be able to undergo relatively slow spontaneous activation independent of interaction with any activator molecule. In this instance, activators may be seen as acting as catalysts to greatly accelerate a slow spontaneous conversion of BAX and BAK to activated forms. This model conforms to the apparently general agreement that an activated form of BAX and BAK must be attained somehow to accomplish apoptosis30,33,51,58,68. Anti-apoptotic proteins prevent death by sequestering activator proteins, but also in some circumstances monomeric activated BAX or BAK.
BCL2 dependence and ‘primed for death’Cancer cells exhibit many abnormalities, such as genomic instability or oncogene activation, which should normally induce death signalling through the intrinsic apoptotic pathway. For example, cell death can be induced by the deregulated expression of an oncogene, such as MyC, which triggers the tumour suppressor ARF (encoded by cyclin-dependent kinase inhibitor 2A (CDKN2A)) that activates p53 (ReF. 73). In turn, p53 can increase the transcription of pro-death BH3-only proteins PuMA28 and NOXA27, resulting in apoptosis. It seems likely that this pathway is used as an important form of cell-autonomous tumour surveillance at the single-cell stage. Of course, cancer cells find a way to survive such signalling, suggesting that a block in apoptosis might be a requirement of oncogenesis. Given our outline of the intrinsic, or mitochondrial, apoptotic pathway, one can envision three distinct ways this block could be effected72. One way they might escape apoptosis is by reducing or eliminating the activation of pro-death BH3-only proteins that otherwise would be activated by the aberrant phenotype (FIG. 4). This might theoretically
Figure 2 |SummaryfigureofthestructuresofBcL2familyproteinsbygroup.There are three main classes of proteins in the BCL2 family. The anti-apoptotic proteins contain four α-helical BCL2 homology (BH) domains. The BH1, BH2, and BH3 domains form a hydrophobic cleft that binds the hydrophobic face of pro-apoptotic BH3 domains. Multidomain pro-apoptotic proteins have the BH1, BH2 and BH3 domains. The pro-apoptotic BH3-only proteins share only a BH3 domain in common with the BCL2 family. Flanking regions are unrelated, and probably relate to non-apoptotic cellular functions. Whether PUMA is a member of the sensitizer or activator class of BH3 proteins still remains to be clarified, as shown by the question mark. BAD, BCL2 antagonist of cell death; BAK, BCL2-antagonist/killer; BAX, BCL2-associated X protein; BID, BH3-interacting domain death agonist; BIK, BCL2-interacting killer; BMF, Bcl2-modifying factor; BOK, BCL2-related ovarian killer; MCL1, myeloid cell leukaemia sequence 1; TM, transmembrane domain.
R E V I E W S
124 | FeBRuARy 2008 | vOluMe 8 www.nature.com/reviews/cancer
© 2008 Nature Publishing Group

Nature Reviews | Cancer
Activator BH3-only
Effector multidomainpro-apoptotic
Multidomainanti-apoptotic
Sensitizer BH3-only
BAX
BAK
BAD
BIK
NOXA
HRK
BMF
BID
BIM
BCL2
BCL-XL
BCL1
BCL-w
BFL1
Cytochrome c release Death
Damage or deregulation signals• Checkpoint violation• Genomic instability• Oncogene activation
PUMA
?
be accomplished by deletion of key BH3-only genes or by genetic modulation of any of the poorly understood interactions functioning upstream of BH3-only upregu-lation. An example would be inactivation of p53, which controls expression of BH3-only proteins PuMA and NOXA. We refer to this as a class A block.
Alternatively, the cancer cell might eliminate the effector arm of this pathway by reducing or eliminat-ing BAX and BAK. It has been shown that loss of BAX and BAK confers a profound apoptotic block, and that BH3-only proteins depend absolutely on BAX or BAK for their pro-death function35,36. We have found tumour cells that use this route of escape, and refer to this as a class B block72.
Finally, the apoptotic fate may be foiled by the increased expression of an inhibitor such as BCl2, MCl1 or a related anti-apoptotic protein. We refer to this as a class C block. In cells employing this block, pro-apoptotic activator BH3-only proteins that would otherwise activate BAX or BAK and cause MOMP are bound and sequestered by the anti-apoptotic proteins. Thus the anti-apoptotic proteins are largely bound to, or primed with death signals in the form of, pro-apoptotic activator proteins. We refer to cells in this state as primed, or primed for death30. Primed cells depend on the continuous function of anti-apoptotic proteins to maintain survival. If anti-apoptotic proteins are lost, or their functions antagonized, the pro-death BH3-only proteins are released to activate BAX and BAK and commit the cell to PCD. The discussion that fol-lows describes priming in terms of activator BH3-only proteins, as that is what we have consistently observed. However, adherents to the indirect activation model may mentally substitute priming with activated BAX or BAK monomers and maintain much of the substance of the discussion.
BH3 profiling — detecting escape from apoptosisAs discussed above, in theory, cancer cells can be clas-sified into three distinct groups on the basis of how apoptosis has been suppressed. To move beyond a merely theoretical treatment of this construct, it was necessary to develop a technique for determining what type of block a given cell employs. To this end, we have developed a strategy we call BH3 profiling30,72 (BOX 2). BH3 profiling is based on the selective interactions that take place between the anti-apoptotic proteins and the BH3 domains of sensitizer BH3-only proteins. In TABLe 1 we summarize the interaction pattern of anti-apoptotic proteins with oligopeptides that are derived from the BH3 domains of BH3-only proteins. It can be seen that, although all of the anti-apoptotic proteins interact with the activators BID and BIM, each of the anti-apoptotic proteins may be distinguished from the others based on its pattern of interaction with the BH3 domains of sensitizer BH3 proteins.
We used these observations to generate BH3 profil-ing, a method to classify types of apoptotic block. We have now validated this strategy in model systems, cell lines and primary cancer cells and have found it to be a useful assessment of the apoptotic blocks employed
by cancer cells. This functional assay, which can be performed in an afternoon, can be thought of as inte-grating the function of the myriad BCl2 family proteins that are simultaneously expressed in most cells to derive a comprehensive picture of the arrangement of the cell death pathway.
Figure 3 |amodelofBcL2familycontrolovermitochondrialapoptosis.In response to cellular damage or deregulation, activator BH3-only proteins such as BH3-interacting domain death agonist (BID) or BIM are activated by transcription or post-translational modification. Activator BH3-only proteins activate effectors such as BCL2-associated X protein (BAX) and BCL2-antagonist/killer (BAK), inducing homo-oligomerization, resulting in mitochondrial permeabilization and commitment to death. Anti-apoptotic proteins sequester activators to prevent them contacting effectors, and also might sequester activated monomeric BAX or BAK. Sensitizers act as selective antagonists of anti-apoptotic proteins. Whether PUMA is a member of the sensitizer or activator class of BH3 proteins remains to be shown clearly, so it is placed in between the two classes in this figure. BAD, BCL2 antagonist of cell death; BID, BH3-interacting domain death agonist; BIK, BCL2-interacting killer; BMF, Bcl2-modifying factor; BOK, BCL2-related ovarian killer. Modified, with permission, from ReF. 72 Elsevier Science (2007).
R E V I E W S
NATuRe RevIeWs | cancer vOluMe 8 | FeBRuARy 2008 | 125
© 2008 Nature Publishing Group

Initiator caspasesThat subset of the caspase family, including caspases 8 and 9, that are activated early in apoptotic signalling. Their cleavage targets are restricted, and include the effector caspases.
Effector caspasesThat subset of the caspase family, including caspases 3 and 7, that are activated by initiator caspases in apoptotic signalling. Their cleavage targets are broadly distributed throughout the cell; activation of effector caspases is generally a terminal event for a cell.
Drugs targeting anti-apoptotic proteinsThe understanding and identification of cancer cells that depend on BCl2 or related anti-apoptotic proteins is of great practical significance owing to the recent emergence of drugs targeting BCl2 from preclinical studies into clinical trials (TABLe 2). The first attempt to introduce an agent that specifically targeted BCl2 was made by Genta with their antisense DNA agent, oblim-ersen (Genasense). On the basis of preclinical studies that found that antisense inhibition of BCL2 levels could induce death in cancer cell lines, they designed a phos-phorothioate DNA molecule complementary to BCL2 (ReF. 74). Preclinical studies with this agent supported its ability to reduce BCl2 in certain cells. However, numerous clinical studies failed to reach their designated clinical endpoints. More recently, in a study of chronic lymphocytic leukaemia (Cll), evidence emerged indi-cating that an improvement in response rate could be observed in relapsed Cll patients when oblimersen was added to fludarabine and cyclophosphamide75. However, it is not clear that this will be enough to prompt approval by the united states Food and Drug Administration.
If dependence on BCl2 is found selectively in cancer cells, as I have argued above, why was this drug not more successful? It may have to do with its mechanism of action. It did not inhibit BCl2 function — rather it attempted to reduce BCl2 levels. There is evidence that this down-regulation can induce other anti-apoptotic proteins in response, which would mitigate the drug’s effectiveness76. Furthermore, it is not clear what level of reduction of
BCl2 could be obtained in vivo as little data to address this question is publicly available. It is possible that the cytotoxic effects that are seen are due to off-target activity. For instance, CpG dinucleotides, which are found in the oblimersen molecule, have been shown to stimulate an immune response independent of interaction with BCL2 (ReF. 77). Off-target mechanisms independent of immune system stimulation have also been proposed78. Finally, with the exception of Cll79, it is not clear which cancers tested in clinical trials required BCl2 for survival. In short, though certainly somewhat disappointing, oblimersen does not represent a sufficient test of BCl2 antagonism to induce general pessimism about the approach.
subsequent approaches have focused on inhibiting BCl2 function with small molecules rather than on decreasing BCl2 protein levels. Prominent among these is the Abbott drug ABT-263, which is now in clinical trials for small cell lung cancer, lymphoma and Cll. ABT-263 is an orally available modification of the ABT-737 com-pound that has been studied extensively in preclinical studies30,80–84. A strategy employed by Abbott laboratories that was nicknamed ‘sAR by NMR’ (structure–activity relationships by NMR) ultimately yielded ABT-737, a molecule that binds the BCl2, BCl-Xl and BCl-w binding pockets with sub-nanomolar affinity. By con-trast, it shows low affinity for MCl1 or BFl1, two other anti-apoptotic proteins. It can be seen that ABT-737 in fact mimics the binding pattern of the BH3 domain of BAD, suggesting that both the compound and the peptide are recognizing features common to the hydrophobic cleft in all three molecules.
ABT-737 is toxic to a subset of cancer cell lines. There are numerous studies that support the contention that the mechanism of action is indeed inhibition of BCl2 (ReFS 30,80–84). It is important to understand what makes certain cells sensitive to ABT-737 and others resistant. One hypothesis is that response to ABT-737 depends primarily on the amount of MCl1 present81,84. As ABT-737 does not efficiently target MCl1, it seems reasonable to suggest that presence of excess MCl1 would maintain survival even if BCl2 were inhibited. Indeed, we also have found that systems defined as MCl1-dependent by BH3 profiling or by genetic design are resistant to ABT-737 (ReFS 30,79). Others have shown that, in
Box 1 | Caspase activation and phagocytosis
Activation of the cysteine proteases from the family known as caspases is an important hallmark of apoptotic death. Mitochondrial outer membrane permeabilization allows the release of cytochrome c from the mitochondrial intermembrane space. Once in the cytosol, cytochrome c interacts with apoptotic peptidase activating factor 1 (APAF1) and the initiator procaspase 9 to form, with dATP, a holoenzyme known as the apoptosome. Activated caspase 9 in the apoptosome then cleaves procaspase 3 into the effector caspase caspase 3 (ReFS 5,55). The activated caspases cause cell-wide specific proteolysis and dysfunction. Resulting effects include a decrease in ATP production and a labelling of the cell with ‘eat me’ signals, allowing the apoptotic cell to be recognized and consumed by phagocytic cells56,57. This results in a minimization of resources wasted by damaged cells, and a reduction in counter-productive immunological stimulation by dead cells.
Box 2 | BH3 profiling
The basic strategy is to incubate mitochondria isolated from the cell of interest with the panel of BH3 peptides from TABLe 1, and observe which peptides cause permeabilization of the mitochondrial outer membrane. We have used release of cytochrome c as a measurement of mitochondrial outer membrane permeabilization. Based on the pattern of response to the panel of peptides, we can then assign the cell to one of the classes defined in FIG. 4. For instance, if a cell has eliminated the upregulation of activator BH3-only proteins, but maintains adequate BCL2-associated X protein (BAX) and BCL2-antagonist/killer (BAK), its mitochondria will respond to BH3-interacting domain death agonist (BID) and BIM, but not to any of the sensitizer BH3 peptides. Note that this is also the pattern that might be expected of normal cells that are not enduring active death signalling. This gives a pattern (FIG. 4) that defines a class A block. If, however, the effector arm is eliminated by loss of BAX and BAK, none of the peptides will cause mitochondrial outer membrane permeabilization, resulting in a flat pattern characteristic of a class B block. Finally, if the cell is primed, and dependent upon anti-apoptotic BCL2 proteins to sequester activator BH3-only proteins, we would see response to both activator and sensitizer peptides. Moreover, by reference to the interaction patterns in TABLe 1, the specific pattern of activity of the sensitizer peptides would reveal the identity of the anti-apoptotic protein(s) that are most responsible for keeping the cell alive. Note that, regardless of whether the priming is mainly by activators, as the direct activation model would suggest, or BAX and BAK, as the indirect model would suggest, BH3 profiling will accurately identify primed cells.
R E V I E W S
126 | FeBRuARy 2008 | vOluMe 8 www.nature.com/reviews/cancer
© 2008 Nature Publishing Group

Nature Reviews | Cancer
Class A
Class B Class C
Activator BH3, such as BIM
Activator BH3, such as BIMI
Activator BH3, such as BIM
Activator BH3, such as BIM
BAX/BAK
Death
BAX/BAK
Survival and oncogenesis
BAX/BAK BCL2MCL1BFL1BCL-wBCL-XL
Abnormal cell behaviour, genomic instability,oncogene activation,check-point violation.
BAX/BAK
small groups of (leukaemia) cell or model systems, high levels of MCl1 expression likewise confer resist-ance to ABT-737 (ReFS 81,84). On the basis of both mechanisms and observed experimental results, there is little reason to dispute the suggestion that high levels of MCl1 contribute to resistance to ABT-737 and other selective antagonists of BCl2.
However, I think it oversimplifies the issue to suggest that low MCl1 levels alone dictate sensitivity to ABT-737. In order to be susceptible to a BCl2 antagonist like ABT-737, the cell has to be dependent on BCl2 in the first place. Above we have described a mechanism for depend-ence on BCl2. When BCl2 is required for the tonic sequestration of pro-death proteins, the cell dies when BCl2 function is attenuated. We can detect such cells through BH3 profiling. We have demonstrated an excel-lent correlation between cells with a BCl2-dependent pattern on BH3 profiling and cells that are sensitive to BCl2 inhibition30,72,79. We have likewise found that dependence on BCl2 corresponds to BCl2’s sequestra-tion of pro-death molecules like BIM. We cannot rule out that in some instances BCl2 is instead primed by mono-mers of activated BAX or BAK, but in our observations so far BCl2 dependence is more closely linked to priming of BCl2 by activator BH3-only molecules. Of course, even primed cells can be rendered more resistant to ABT-737 by the overexpression of MCl1. Thus, MCl1 can be considered a factor promoting resistance to ABT-737, but primed BCl2 is the foundation for the mechanism governing sensitivity to BCl2 inhibition (FIG. 5).
Clinical useA question that often arises is whether antagonists of BCl2 and related anti-apoptotic proteins are likely to find a clinical niche as single-agent therapy or as components of a multiagent chemotherapy regimen. I would suggest that majority opinion would predict that BCl2 antagonists will have little effect as single agents. This opinion comes from the idea that, in cancer, overexpressed BCl2 serves to provide resistance to therapeutic drugs rather than endogenous death signalling. The idea is that BCl2 antag-onists will serve mainly to counteract BCl2-mediated resistance, allowing killing by conventional agents. In this view, BCl2 antagonism by itself is unlikely to cause much cancer cell death. Indeed, many, if not most, successful anti-cancer agents eventually find their primary use in combination with other effective agents, so it is quite likely that anti-BCl2 agents will be used similarly.
yet the ability of BCl2 antagonists like ABT-737 to kill cells as monotherapy in preclinical models has been impressive. Primary Cll cells are killed at concentrations <20 nM within 4 h79, and primary acute lymphoblastic leukaemia cells are killed in the 10–400 nM range85. In addition, many sensitive cancer cell lines are killed in the nanomolar range in vitro30,72,80–84. Of course it remains to be seen whether such impressive anti-cancer activ-ity will be replicated in vivo. However, these in vitro observations demonstrate that single-agent therapy is at least theoretically feasible, an observation that might have surprised some. Of course, before successful results from clinical trials of these agents are obtained, discuss-ing final clinical use is somewhat premature. Moreover, the use will depend on complicated balancing of toxicity to the tumour versus toxicity to normal tissues, a balance that cannot be estimated outside of clinical trials. suffice it to say that current knowledge is insufficient to rule out the optimal use of BCl2 inhibitors as monotherapy or combination therapy.
Another question that often arises is whether the ideal drug would be a selective inhibitor of anti-apoptotic proteins (that is, inhibiting only a subset), or one that inhib-ited all of the anti-apoptotic proteins (a ‘pan-inhibitor’). The feasibility of a synthesizing a pan-inhibitor drug is not far-fetched. Indeed, the PuMA BH3 peptide serves as proof that a single molecule can antagonize at least five cellular anti-apoptotic proteins, though in a peptide form that is pharmacologically useless30. Certain small molecules in pre-clinical and clinical development show the ability to bind all anti-apoptotic family members, but the affinity appears to be relatively low in these cases86. If priming is indeed entirely specific to cancer cells, a pan-inhibitor may be preferable. It would also prevent what seems to be an obvious route of escape from a selective inhibitor, overexpression of an anti-apoptotic protein not targeted by the selective inhibitor. However, it would be no benefit if it was also toxic to non-malignant cells. Already some toxicity is seen in BCl2-specific inhibitors such as ABT-737 in the form of thrombocytopenia and lymphopenia82,87. Only a clini-cal trial can evaluate the complicated balance between tumour and normal cell toxicity adequately enough to answer this question.
Figure 4 |Threeclassesofapoptoticblocksusedtomaintaincancersurvival.Apoptosis is a tumour-suppressive mechanism that cancer cells must overcome to endure. Selective pressure during tumorigenesis can reduce or eliminate the activation of pro-death BH3-only proteins, perhaps through the deletion of key BH3-only genes or by genetic modulation of any of the poorly understood interactions functioning upstream to regulate these proteins. We refer to this as a class A block. Alternatively, the effector arm of the mitochondrial apoptotic pathway could be mutated, resulting in the reduction or elimination of BCL2-associated X protein (BAX) and BCL2-antagonist/killer (BAK). This we refer to as a class B block72. Finally, cancer cells may have increased expression of an inhibitor such as BCL2, myeloid cell leukaemia sequence 1 (MCL1) or a related anti-apoptotic protein. We refer to this as a class C block. Modified, with permission, from ReF. 72 Elsevier Science (2007).
R E V I E W S
NATuRe RevIeWs | cancer vOluMe 8 | FeBRuARy 2008 | 127
© 2008 Nature Publishing Group

BH3 profiling might have a useful role in the clinic as well. By identifying cells that are dependent on anti-apoptotic proteins, it has the potential to select patients that are most likely to respond to targeted inhibitors of anti-apoptotic proteins. Treating tumour cells isolated from tumour biopsies with a number of drugs to ascertain drug efficacy might well be unsatisfactory, owing to the long observed problem of ex vivo culture altering tumour cell biology and, hence, sensitivity to anti-tumour agents. One could consider simply treating mitochondria isolated from the patient’s tumour cells with a drug directly and observ-ing effect. However, we have found this to be less reliable than assessing anti-apoptotic protein dependence using peptides. This might be because proteins and membranes in the concentrated mitochondrial preparations inter-act non-specifically with the drugs. In addition, direct mitochondrial drug treatment works only for the subset of drugs with a mitochondrial target.
As with any summary assay based on an aggregate of many cells, it is quite possible that heterogeneity in tumours could influence results. For instance, if there were a mixture of primed and unprimed cells in a tumour population, BH3 profiling might yield results showing intermediate priming that really did not apply to either subset alone. We are pursuing two ways to address this potential issue. One is to prospec-tively identify such populations and to separate them by cell sorting before mitochondrial purification. The other is to modify BH3 profiling to a single cell-based assay so that individual cells may be evaluated by fluorescence-activated cell sorting. In this way, indi-vidual subsets could be prospectively identified in the case of heterogeneity.
BCL2 as a marker of chemosensitivity?Above we described how cancer cells can be segregated into three classes based on the type of apoptotic block they harbour. We also discussed how membership of a subgroup of one of these classes, class C, confers sensitiv-ity to antagonists of BCl2. However, many other cancer therapeutics make use of the intrinsic apoptotic pathway to kill cancer cells. It is worth considering whether sensi-tivity to these conventional agents might be predicted on the basis of membership of one of these classes.
A priori, one might expect that cells in class C might be most sensitive to agents killing through the intrinsic pathway. These cells possess all the required elements for apoptosis to occur — activator BH3-only proteins and effectors BAX and BAK. They are kept alive in a tenu-ous dynamic equilibrium only by a balancing amount of anti-apoptotic proteins. After treatment with drug, the BH3-only proteins that are generated, whether sensitizer or activator, can either activate BAX and BAK or displace pre-bound activators to do the same. By contrast, class A or B cells lack key components of the pro-death signal-ling pathway: activators or BAX and BAK, respectively. Of course, these blocks might be reversible, and the treatment in question might theoretically contribute to reversing them. However, it would seem that this proc-ess would probably nonetheless result in a less robust response than that expected by class C cells (FIG. 5).
We tested this hypothesis in a limited set of lym-phoma cell lines72. We found that the primed cells of class C were indeed more sensitive than unprimed class A or B cells to treatment with several drugs that operate in the intrinsic apoptotic pathway, including vincristine, doxorubicin and etoposide. As a control, we also tested sensitivity to N-methyl-N′-nitro-N-nitrosoguanidine, an alkylating agent that has been shown to kill efficiently by a non-apoptotic pathway in cells that have profound blocks in apoptosis. As expected, there was no distinc-tion found between the primed and unprimed cells in this case. Thus, membership in an unprimed class (A or B) predicted an inferior in vitro response to one group of drugs, but not to another. Of course, these results need to be confirmed in a larger cohort of cells and drugs. The clinical implications are worth considering, however. One could potentially evaluate the suitability of drugs for individual cancers on the basis of knowing how apop-tosis is blocked. As BH3 profiling can be performed on fresh tissue, the problem of ex vivo cell culture, which has plagued former attempts to predict sensitivity, would be avoided.
It should be noted that our observation has been made on a fairly limited number of cell lines. The con-sistency of the finding surprised us, and it is possible that when we compare BH3 profiling and drug response on a larger number of cell lines we will discover cell lines that
Table 1 | Selective interactions among anti-apoptotic proteins and BH3 domains
anti-apoptoticprotein
activator Sensitizer
BID BIM BaD BIK nOXaa nOXaB HrK PUMa BMF
BCL2 66 (6) <10 11 (3) 151 (2) – – – 18 (1) 24 (1)
BCL-XL 12 (9) <10 <10 10 (2) – – 92 (11) <10 <10
BCL-w <10 38 (7) 60 (19) 17 (12) – – – 25 (12) 11 (3)
MCL1 <10 <10 – 109 (33) 19 (2) 28 (3) – <10 23 (2)
BFL1 53 (3) 73 (3) – – – – – 59 (11) –Dissociation constants for interactions between anti-apoptotic BCL2 family proteins (left) and BCL2 homology 3 (BH3) domains from BH3-only proteins (top) are shown in nM. Standard deviations of at least three independent measurements are in parentheses. Empty cells signify no observed binding (Kd > 2,500 nM). BAD, BCL2 antagonist of cell death; BID, BH3-interacting domain death agonist; BIK, BCL2-interacting killer; BMF, Bcl2-modifying factor; MCL1, myeloid cell leukaemia sequence 1. Modified, with permission, from ReF 30 Elsevier Science (2006).
R E V I E W S
128 | FeBRuARy 2008 | vOluMe 8 www.nature.com/reviews/cancer
© 2008 Nature Publishing Group

have resistance mechanisms outside the mitochondrion, so that the correlation with class C and chemosensitivity may be less tight for certain cells. However, in our exten-sion of BH3 profiling to several dozen cell lines, we find multiple examples of all three classes of apoptotic block, at least raising our confidence that all three are indeed commonly exploited in cancer (Ryan, J. and A.G.l., unpublished data).
so it appears that membership of the primed class C might augur greater sensitivity to numerous chemother-apy agents. However, when one considers what it means to be primed, an apparent paradox is confronted. High expression of BCl2 or a related anti-apoptotic protein is necessary for a cell to be primed. Can we really provide support for the heretical notion that overexpression of BCl2 can imply chemosensitivity? One of the reasons why this idea may seem surprising is that most of our knowledge of BCl2 function in chemoresistance comes from overexpression models in tissue culture cells. In these models, additional BCl2 expression is added to a cell that has already come to terms with its environment, so that the additional BCl2 provides extra anti-apoptotic reserve. such a cell is consistently found to be less sen-sitive than the parental cell to agents that induce the intrinsic apoptotic pathway. The appropriate interpre-tation is that BCl2 confers chemoresistance in these models. However, expression of BCl2 in cancer cells is driven by selection rather than exogenously derived over-expression. High BCl2 expression is selected for during oncogenesis to buffer the death signals generated by the aberrant phenotypes required for illegitimate growth. Here this extra BCl2 does not necessarily provide an additional anti-apoptotic reserve to ensure chemoresist-ance. Rather, the BCl2 selected for is already occupied by pro-death proteins. In other words, in overexpression tissue culture models, the BCl2 is largely ‘empty’; the BCl2 selected for in cancer is already ‘full’ or ‘primed’.
In fact, there are some fairly obvious clinical mod-els that support this interpretation. Cll and follicular lymphoma are both well-established as consistently over-expressing BCl2. Both cancers consist of relatively slowly
dividing cells. By conventional estimation, therefore, one would expect these cancers to be quite resistant to chemotherapy. The consistent clinical observation, however, is that both cancers are extremely sensitive to chemotherapy. When treated with conventional multi-agent regimens, the majority of patients with either dis-ease will enter a complete remission. In support of our biochemical interpretation of this behaviour, it has been shown that cells from both cancers are sensitive in vitro to BCl2 antagonism by ABT-737, and hence likely to be primed. Indeed, we have shown explicitly that the BCl2 in Cll is consistently primed with pro-apoptotic BIM79. Moreover, several studies have shown that expression of BCl2 confers a superior prognosis to patients with breast cancer88–91.
It is true that, despite a robust initial response, nei-ther Cll or follicular lymphoma are generally cured by chemotherapy, or any other modality, but this may be because of either the existence of a minority stem-like cell with different properties to the bulk population or the evolution of cells within the original tumour. It is worth noting that on treatment with chemotherapy, can-cer cells are exposed to selection pressures that can select for expression of anti-apoptotic proteins at levels greater than before. These levels may then provide excess anti-apoptotic reserve to cancer cells between treatments, and treatment resistance when the treatment cycle resumes.
If the primed cells that are best targeted by antagonists of anti-apoptotic proteins like ABT-263 are also killed more readily by conventional chemotherapy, is it pos-sible that such strategies will only treat cancers that are already adequately treated? I think that there exists ample opportunity for the anti-apoptotic antagonists to provide significant clinical benefit. All too many cancers that respond to conventional agents have an unsatisfactory clinical response. In this case, addition of a tolerable agent that efficiently kills the cancer cell might be expected to improve clinical outcome. In addition, the testing I describe attempts to predict sensitivity to conventional or targeted drugs only as single agents. It seems likely that use of conventional agents in combination with the
Table 2 | Clinical development of drugs targeting anti-apoptotic proteins
Drug company clinicalphase Function additionalcomments
ABT-263 Abbott Laboratories Phase I and I/II a clinical trials in NHL, CLL and SCLC
BH3 mimetic, targets BCL2, BCL-XL, BCL-w
ABT-263 is an orally available compound closely related to ABT-737
Obatoclax (GX15-070)
Gemin X Multiple phase I and phase II clincial trials in haematological malignancies and non-small cell lung cancer
BH3 mimetic Might be a pan-inhibitor of anti-apoptotic proteins
Oblimersen (G3139)
Genta Many clinical trials including phase III in melanoma and CLL
Antisense DNA targeting BCL2
None
AT-101 Ascenta Therapeutics Phase II clinical trials in a variety of cancers
BH3 mimetic AT-101 is the (–) enantiomer of gossypol
BH, BCL2 homology; CLL, chronic lymphocytic leukaemia; NHL, non-Hodgkin lymphoma; SCLC, small-cell lung cancer.
R E V I E W S
NATuRe RevIeWs | cancer vOluMe 8 | FeBRuARy 2008 | 129
© 2008 Nature Publishing Group

Class A
Nature Reviews | Cancer
Class B
Class C
Survival
Cytotoxic chemotherapy
Sensitizer BH3 mimetic
Cytotoxic chemotherapy
Sensitizer BH3 mimetic
Cytotoxic chemotherapy
Sensitizer BH3 mimetic
Survival
Survival
Survival
Death
MOMP
MOMP
Death
Activator BH3 Sensitizer BH3 BCL2 BAX or BAK
a
b
c
anti-apoptotic agents might expand the spectrum of sensitive cells into the class A or even class B group, as the conven-tional agents may provide the extra priming necessary for anti-apoptotic antagonists to work in cancers in these classes. Furthermore, ABT-263 has a fairly restricted pattern of toxicity, without widespread genotoxicity
Figure 5 |WhyprimedcancercellsmaybeselectivelysensitivetoBcL2antagonistsandconventionalchemotherapy.Cytotoxic chemotherapy is shown generating a mixture of pro-apoptotic sensitizer and activator BCL2 homology 3 (BH3)-only molecules. Class A cells lack pre-existing activator bound to anti-apoptotic proteins like BCL2, and thus are relatively resistant to new burdens of BH3-only molecules or BH3 mimietics. Class B cells lack BCL2-associated X protein (BAX) and BCL2-antagonist/killer (BAK), and so are less prone to respond to excess activator molecules with mitochondrial outer membrane permeabilization (MOMP). Class C cells are primed, and therefore are more prone to displacing excess pre-existing activators that can then activate BAX and BAK at the mitochondrion, causing MOMP.
or neurotoxicity, so a greater therapeutic window might be observed than for most conventional agents. Finally, I do not mean to suggest that the mitochondrion is the sole determinant of response, just one that has been somewhat overlooked. There are doubtlessly many mechanisms of resistance to conventional agents that lie upstream of mito-chondria. However, cells resistant to chemotherapy on the basis of these upstream blocks might still be sensitive to drugs like ABT-263.
An explanation for chemotherapy efficacy?In the decades of chemotherapy that modern oncol-ogy has witnessed, it has been observed that cancer cells that are resistant to one agent are often resistant to many agents, even those targeting sites distal to that targeted by the initial agent. On the other hand, many cancers that are sensitive to single agents are sensitive to many agents working by different mechanisms. This can be clearly seen in the fact that most, if not all, cura-tive chemotherapy regimens use a combination of drugs working on different targets. These observations have led to investigation of whether there is not some common mechanism defining resistance in pan-resistant cells. At the very start of the pathway of drug-induced death is the necessity that the drug gain cell entry. One obvious mechanism that has been explored is that of entry of drug into the cancer cell. Indeed, p-glycoprotein-type multi-drug resistance pumps might account for a subset of pan-resistance observed92. However, the proportion is probably less than initially thought and, further-more, intervention by drug pump inhibitors has been disappointing.
I would like to suggest that the specific class of block in apoptosis found in a cancer cell might be a heretofore unappreciated and crucial factor in determining the sensitivity of cancer cells to multiple diverse chemo-therapy agents simultaneously. The death signalling pathways of a wide variety of different cytotoxic agents converge at the intrinsic cell death pathway. Therefore, a particular class of block, such as a class C block, might broadly confer sensitivity to a wide variety of agents. Another block, such as a class B block, might on the other hand confer resistance to a wide variety of agents. Different blocks in apoptosis might be an explana-tion of why certain cancers, such as leukaemias, are intrinsically more sensitive to almost all conventional agents than other cancers, such as pancreatic cancer or melanoma. Moreover, the fact that being primed for death (class C block) is apparently much more common in tumours than in non-malignant tissues might explain why chemotherapy is often more toxic to cancers than normal tissues. The idea that cancer cells might be more prone than normal cells to die is not a new one, but here we present a biochemical explanation for that clinical suspicion. With newly discovered tools we can system-atically explore how often the mechanism of apoptotic escape determines chemosensitivity. Moreover, with novel therapeutic agents, we already have promising evidence that we will be able to selectively target some of the barriers constructed by cancer cells to evade their rightful death sentence.
R E V I E W S
130 | FeBRuARy 2008 | vOluMe 8 www.nature.com/reviews/cancer
© 2008 Nature Publishing Group

1. Castedo, M. et al. Cell death by mitotic catastrophe: a molecular definition. Oncogene 23, 2825–2837 (2004).
2. Ellis, H. M. & Horvitz, H. R. Genetic control of programmed cell death in the nematode C. elegans. Cell 44, 817–829 (1986).
3. Hengartner, M. O. & Horvitz, H. R. C. elegans cell survival gene ced-9 encodes a functional homolog of the mammalian proto-oncogene bcl-2. Cell 76, 665–676 (1994).
4. Vaux, D. L., Weissman, I. L. & Kim, S. K. Prevention of programmed cell death in Caenorhabditis elegans by human bcl-2. Science 258, 1955–1957 (1992).
5. Wang, X. The expanding role of mitochondria in apoptosis. Genes Dev. 15, 2922–2933 (2001).
6. Colell, A. et al. GAPDH and autophagy preserve survival after apoptotic cytochrome c release in the absence of caspase activation. Cell 129, 983–997 (2007).
7. Tsujimoto, Y., Gorham, J., Cossman, J., Jaffe, E. & Croce, C. M. The t(14;18) chromosome translocations involved in B-cell neoplasms result from mistakes in VDJ joining. Science 229, 1390–1393 (1985).
8. Cleary, M. L. & Sklar, J. Nucleotide sequence of a t(14;18) chromosomal breakpoint in follicular lymphoma and demonstration of a breakpoint-cluster region near a transcriptionally active locus on chromosome 18. Proc. Natl Acad. Sci. USA 82, 7439–7443 (1985).
9. Bakhshi, A. et al. Cloning the chromosomal breakpoint of t(14;18) human lymphomas: clustering around JH on chromosome 14 and near a transcriptional unit on 18. Cell 41, 899–906 (1985).
10. McDonnell, T. J. et al. bcl-2-immunoglobulin transgenic mice demonstrate extended B cell survival and follicular lymphoproliferation. Cell 57, 79–88 (1989).
11. McDonnell, T. J. & Korsmeyer, S. J. Progression from lymphoid hyperplasia to high-grade malignant lymphoma in mice transgenic for the t(14; 18). Nature 349, 254–256 (1991).
12. Boise, L. H. et al. bcl-x, a bcl-2-related gene that functions as a dominant regulator of apoptotic cell death. Cell 74, 597–608 (1993).In this report, pro- and anti-death proteins transcribed by the bcl-x locus were first described.
13. Gibson, L. et al. bcl-w, a novel member of the bcl-2 family, promotes cell survival. Oncogene 13, 665–675 (1996).
14. Kozopas, K. M., Yang, T., Buchan, H. L., Zhou, P. & Craig, R. W. MCL1, a gene expressed in programmed myeloid cell differentiation, has sequence similarity to BCL2. Proc. Natl Acad. Sci. USA 90, 3516–3520 (1993).
15. Choi, S. S. et al. A novel Bcl-2 related gene, Bfl-1, is overexpressed in stomach cancer and preferentially expressed in bone marrow. Oncogene 11, 1693–1698 (1995).
16. Oltvai, Z. N., Milliman, C. L. & Korsmeyer, S. J. Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that accelerates programmed cell death. Cell 74, 609–619 (1993).This seminal initial report showed that BAX is a pro-death protein related to BCL2 that exerts a pro-death function. Along with reference 12, this paper clearly indicated that BCL2 was only the first member of what turned out to be a large family of related proteins that control apoptosis.
17. Chittenden, T. et al. A conserved domain in Bak, distinct from BH1 and BH2, mediates cell death and protein binding functions. EMBO J. 14, 5589–5596 (1995).This paper first identified the importance of the BH3 domain in facilitating the heterodimeric interactions that are crucial to BCL2 family function.
18. Hsu, S. Y., Kaipia, A., McGee, E., Lomeli, M. & Hsueh, A. J. Bok is a pro-apoptotic Bcl-2 protein with restricted expression in reproductive tissues and heterodimerizes with selective anti-apoptotic Bcl-2 family members. Proc. Natl Acad. Sci. USA 94, 12401–12406 (1997).
19. Huang, D. C. & Strasser, A. BH3-only proteins — essential initiators of apoptotic cell death. Cell 103, 839–842 (2000).
20. Wang, K., Yin, X. M., Chao, D. T., Milliman, C. L. & Korsmeyer, S. J. BID: a novel BH3 domain-only death agonist. Genes Dev. 10, 2859–2869 (1996).
21. Boyd, J. M. et al. Bik, a novel death-inducing protein shares a distinct sequence motif with Bcl-2 family proteins and interacts with viral and cellular survival-promoting proteins. Oncogene 11, 1921–1928 (1995).
22. Leber, B., Lin, J. & Andrews, D. W. Embedded together: the life and death consequences of interaction of the Bcl-2 family with membranes. Apoptosis 12, 897–911 (2007).
23. Danial, N. N. & Korsmeyer, S. J. Cell death: critical control points. Cell 116, 205–219 (2004).
24. Puthalakath, H. & Strasser, A. Keeping killers on a tight leash: transcriptional and post-translational control of the pro-apoptotic activity of BH3-only proteins. Cell Death Differ. 9, 505–512 (2002).
25. Puthalakath, H., Huang, D. C., O’Reilly, L. A., King, S. M. & Strasser, A. The proapoptotic activity of the Bcl-2 family member Bim is regulated by interaction with the dynein motor complex. Mol. Cell 3, 287–296 (1999).
26. Zha, J., Harada, H., Yang, E., Jockel, J. & Korsmeyer, S. J. Serine phosphorylation of death agonist BAD in response to survival factor results in binding to 14-3-3 not BCL-XL. Cell 87, 619–628 (1996).
27. Oda, E. et al. Noxa, a BH3-only member of the Bcl-2 family and candidate mediator of p53-induced apoptosis. Science 288, 1053–1058 (2000).
28. Nakano, K. & Vousden, K. H. PUMA, a novel proapoptotic gene, is induced by p53. Mol. Cell 7, 683–694 (2001).
29. Cartron, P. F. et al. The first α helix of Bax plays a necessary role in its ligand-induced activation by the BH3-only proteins Bid and PUMA. Mol. Cell 16, 807–818 (2004).
30. Certo, M. et al. Mitochondria primed by death signals determine cellular addiction to antiapoptotic BCL-2 family members. Cancer Cell 9, 351–365 (2006).The concept of priming of BCL2 proteins and mitochondria with pro-death proteins is demonstrated. BH3 profiling is presented and validated in several test systems.
31. Desagher, S. et al. Bid-induced conformational change of Bax is responsible for mitochondrial cytochrome c release during apoptosis. J. Cell Biol. 144, 891–901 (1999).
32. Kuwana, T. et al. BH3 domains of BH3-only proteins differentially regulate Bax-mediated mitochondrial membrane permeabilization both directly and indirectly. Mol. Cell 17, 525–535 (2005).This report uses a completely defined system of liposomes, recombinant proteins, and synthetic liposomes to demonstrate that BAX requires activation by BH3 domains to exert its permeabilizing function.
33. Kuwana, T. et al. Bid, Bax, and lipids cooperate to form supramolecular openings in the outer mitochondrial membrane. Cell 111, 331–342 (2002).
34. Letai, A. et al. Distinct BH3 domains either sensitize or activate mitochondrial apoptosis, serving as prototype cancer therapeutics. Cancer Cell 2, 183–192 (2002).This paper first presents the activator versus sensitizer dichotomy in the function of BH3 domains. Certain BH3 domains are shown to act as antagonists of BCL2 function.
35. Lindsten, T. et al. The combined functions of proapoptotic Bcl-2 family members Bak and Bax are essential for normal development of multiple tissues. Mol. Cell 6, 1389–1399 (2000).
36. Wei, M. C. et al. Proapoptotic BAX and BAK: a requisite gateway to mitochondrial dysfunction and death. Science 292, 727–730 (2001).
37. Kim, H. et al. Hierarchical regulation of mitochondrion-dependent apoptosis by BCL-2 subfamilies. Nature Cell Biol. 8, 1348–1358 (2006).
38. Yee, K. S. & Vousden, K. H. Contribution of membrane localization to the apoptotic activity of PUMA. Apoptosis 30 Oct 2007 (doi:10.1007/s10495-007-0140-2).
39. Chipuk, J. E. et al. Direct activation of Bax by p53 mediates mitochondrial membrane permeabilization and apoptosis. Science 303, 1010–1014 (2004).
40. Moll, U. M., Wolff, S., Speidel, D. & Deppert, W. Transcription-independent pro-apoptotic functions of p53. Curr. Opin. Cell Biol. 17, 631–636 (2005).
41. Leu, J. I., Dumont, P., Hafey, M., Murphy, M. E. & George, D. L. Mitochondrial p53 activates Bak and causes disruption of a Bak–Mcl1 complex. Nature Cell Biol. 6, 443–450 (2004).
42. Pagliari, L. J. et al. The multidomain proapoptotic molecules Bax and Bak are directly activated by heat. Proc. Natl Acad. Sci. USA 102, 17975–17980 (2005).
43. Hsu, Y. T. & Youle, R. J. Nonionic detergents induce dimerization among members of the Bcl-2 family. J. Biol. Chem. 272, 13829–13834 (1997).
44. Wolter, K. G. et al. Movement of Bax from the cytosol to mitochondria during apoptosis. J. Cell Biol. 139, 1281–1292 (1997).
45. Deng, X., Gao, F., Flagg, T., Anderson, J. & May, W. S. Bcl2’s flexible loop domain regulates p53 binding and survival. Mol. Cell Biol. 26, 4421–4434 (2006).
46. Zhou, H., Hou, Q., Chai, Y. & Hsu, Y. T. Distinct domains of Bcl-XL are involved in Bax and Bad antagonism and in apoptosis inhibition. Exp. Cell Res. 309, 316–328 (2005).
47. Sundararajan, R., Cuconati, A., Nelson, D. & White, E. Tumor necrosis factor-α induces Bax–Bak interaction and apoptosis, which is inhibited by adenovirus E1B 19K. J. Biol. Chem. 276, 45120–45127 (2001).
48. Mikhailov, V. et al. Association of Bax and Bak homo-oligomers in mitochondria. Bax requirement for Bak reorganization and cytochrome c release. J. Biol. Chem. 278, 5367–5376 (2003).
49. Luo, X., Budihardjo, I., Zou, H., Slaughter, C. & Wang, X. Bid, a Bcl2-interacting protein, mediates cytochrome c release from mitochondria in response to activation of cell surface death receptors. Cell 94, 481–490 (1998).
50. Marani, M., Tenev, T., Hancock, D., Downward, J. & Lemoine, N. R. Identification of novel isoforms of the BH3 domain protein Bim which directly activate Bax to trigger apoptosis. Mol. Cell Biol. 22, 3577–3589 (2002).
51. Wei, M. C. et al. tBID, a membrane-targeted death ligand, oligomerizes BAK to release cytochrome c. Genes Dev. 14, 2060–2071 (2000).This paper presents the important finding that a BH3-only protein interacts with a multidomain pro-apoptotic protein to induce apoptosis. These findings demonstrate the importance of activation and oligomerization in BAK function.
52. Pavlov, E. V. et al. A novel, high-conductance channel of mitochondria linked to apoptosis in mammalian cells and Bax expression in yeast. J. Cell Biol. 155, 725–731 (2001).
53. Dejean, L. M. et al. Oligomeric Bax is a component of the putative cytochrome c release channel MAC, mitochondrial apoptosis-induced channel. Mol. Biol. Cell 16, 2424–2432 (2005).
54. Saito, M., Korsmeyer, S. J. & Schlesinger, P. H. BAX-dependent transport of cytochrome c reconstituted in pure liposomes. Nature Cell Biol. 2, 553–555 (2000).
55. Li, P. et al. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 91, 479–489 (1997).
56. Savill, J. & Fadok, V. Corpse clearance defines the meaning of cell death. Nature 407, 784–788 (2000).
57. Fadok, V. A. et al. A receptor for phosphatidylserine-specific clearance of apoptotic cells. Nature 405, 85–90 (2000).
58. Cheng, E. H. et al. BCL-2, BCL-XL sequester BH3 domain-only molecules preventing BAX- and BAK-mediated mitochondrial apoptosis. Mol. Cell 8, 705–711 (2001).This paper develops the important concept, first presented in reference 60, that a key way that antiapoptotic proteins such as BCL-2 oppose death is to sequester BH3-only proteins.
59. Zong, W. X., Lindsten, T., Ross, A. J., MacGregor, G. R. & Thompson, C. B. BH3-only proteins that bind pro-survival Bcl-2 family members fail to induce apoptosis in the absence of Bax and Bak. Genes Dev. 15, 1481–1486 (2001).
60. Cheng, E. H., Levine, B., Boise, L. H., Thompson, C. B. & Hardwick, J. M. Bax-independent inhibition of apoptosis by Bcl-XL. Nature 379, 554–556 (1996).
61. Kelekar, A., Chang, B. S., Harlan, J. E., Fesik, S. W. & Thompson, C. B. Bad is a BH3 domain-containing protein that forms an inactivating dimer with Bcl-XL. Mol. Cell Biol. 17, 7040–7046 (1997).
62. Kelekar, A. & Thompson, C. B. Bcl-2-family proteins: the role of the BH3 domain in apoptosis. Trends Cell Biol. 8, 324–330 (1998).
63. Muchmore, S. W. et al. X-ray and NMR structure of human Bcl-XL, an inhibitor of programmed cell death. Nature 381, 335–341 (1996).
64. Sattler, M. et al. Structure of Bcl-XL–Bak peptide complex: recognition between regulators of apoptosis. Science 275, 983–986 (1997).
65. Chen, L. et al. Differential targeting of prosurvival Bcl-2 proteins by their BH3-only ligands allows complementary apoptotic function. Mol. Cell 17, 393–403 (2005).Here selective interactions among BH3-only and anti-apoptotic proteins are characterized.
R E V I E W S
NATuRe RevIeWs | cancer vOluMe 8 | FeBRuARy 2008 | 131
© 2008 Nature Publishing Group

66. Uren, R. T. et al. Mitochondrial permeabilization relies on BH3 ligands engaging multiple prosurvival Bcl-2 relatives, not Bak. J. Cell Biol. 177, 277–287 (2007).
67. Willis, S. N. et al. Proapoptotic Bak is sequestered by Mcl-1 and Bcl-XL, but not Bcl-2, until displaced by BH3-only proteins. Genes Dev. 19, 1294–1305 (2005).
68. Willis, S. N. et al. Apoptosis initiated when BH3 ligands engage multiple Bcl-2 homologs, not Bax or Bak. Science 315, 856–859 (2007).This report is a presentation of the mature indirect model of apoptosis induction.
69. Harada, H., Quearry, B., Ruiz-Vela, A. & Korsmeyer, S. J. Survival factor-induced extracellular signal-regulated kinase phosphorylates BIM, inhibiting its association with BAX and proapoptotic activity. Proc. Natl Acad. Sci. USA 101, 15313–15317 (2004).
70. Walensky, L. D. et al. A stapled BID BH3 helix directly binds and activates BAX. Mol. Cell 24, 199–210 (2006).
71. Green, D. R. At the gates of death. Cancer Cell 9, 328–330 (2006).
72. Deng, J. et al. BH3 profiling identifies three distinct classes of apoptotic blocks to predict response to ABT-737 and conventional chemotherapeutic agents. Cancer Cell 12, 171–185 (2007).
73. Zindy, F. et al. Myc signaling via the ARF tumor suppressor regulates p53-dependent apoptosis and immortalization. Genes Dev. 12, 2424–2433 (1998).
74. Jansen, B. et al. bcl-2 antisense therapy chemosensitizes human melanoma in SCID mice. Nature Med. 4, 232–234 (1998).
75. O’Brien, S. et al. Randomized phase III trial of fludarabine plus cyclophosphamide with or without oblimersen sodium (Bcl-2 antisense) in patients with relapsed or refractory chronic lymphocytic leukemia. J. Clin. Oncol. 25, 1114–1120 (2007).
76. Konopleva, M. et al. Liposomal Bcl-2 antisense oligonucleotides enhance proliferation, sensitize acute myeloid leukemia to cytosine-arabinoside, and induce apoptosis independent of other antiapoptotic proteins. Blood 95, 3929–3938 (2000).
77. Gekeler, V. et al. G3139 and other CpG-containing immunostimulatory phosphorothioate oligodeoxynucleotides are potent suppressors of the growth of human tumor xenografts in nude mice. Oligonucleotides 16, 83–93 (2006).
78. Castro, J. E. et al. Thymidine-phosphorothioate oligonucleotides induce activation and apoptosis of CLL cells independently of CpG motifs or BCL-2 gene interference. Leukemia 20, 680–688 (2006).
79. Del Gaizo Moore, V. et al. Chronic lymphocytic leukemia requires BCL2 to sequester prodeath BIM, explaining sensitivity to BCL2 antagonist ABT-737. J. Clin. Invest. 117, 112–121 (2007).
80. Chen, S., Dai, Y., Harada, H., Dent, P. & Grant, S. Mcl-1 down-regulation potentiates ABT-737 lethality by cooperatively inducing Bak activation and Bax translocation. Cancer Res. 67, 782–791 (2007).
81. Konopleva, M. et al. Mechanisms of apoptosis sensitivity and resistance to the BH3 mimetic ABT-737 in acute myeloid leukemia. Cancer Cell 10, 375–388 (2006).
82. Oltersdorf, T. et al. An inhibitor of Bcl-2 family proteins induces regression of solid tumours. Nature 435, 677–681 (2005).This paper introduces ABT-737, a high affinity small-molecule antagonist of BCL2, BCL-XL and BCL-w.
83. Tahir, S. K. et al. Influence of Bcl-2 family members on the cellular response of small-cell lung cancer cell lines to ABT-737. Cancer Res. 67, 1176–1183 (2007).
84. van Delft, M. F. et al. The BH3 mimetic ABT-737 targets selective Bcl-2 proteins and efficiently induces apoptosis via Bak/Bax if Mcl-1 is neutralized. Cancer Cell 10, 389–399 (2006).
85. Del Gaizo Moore, V., Schlis, K., Sallan, S., Armstrong, S. & Letai, A. BCL-2 dependence and ABT-737 sensitivity in acute lymphblastic leukemia. Blood 4 Dec 2007 (doi 10.1182/blood-2007-06-098012).
86. Zhai, D., Jin, C., Satterthwait, A. C. & Reed, J. C. Comparison of chemical inhibitors of antiapoptotic Bcl-2-family proteins. Cell Death Differ. 13, 1419–1421 (2006).
87. Zhang, H. et al. Bcl-2 family proteins are essential for platelet survival. Cell Death Differ. 14, 943–951 (2007).
88. Gasparini, G. et al. Expression of bcl-2 protein predicts efficacy of adjuvant treatments in operable node-positive breast cancer. Clin. Cancer Res. 1, 189–198 (1995).
89. Lee, K. H. et al. Prognostic significance of bcl-2 expression in stage III breast cancer patients who had
received doxorubicin and cyclophosphamide followed by paclitaxel as adjuvant chemotherapy. BMC Cancer 7, 63 (2007).
90. Sjostrom, J. et al. The predictive value of bcl-2, bax, bcl-xL, bag-1, fas, and fasL for chemotherapy response in advanced breast cancer. Clin. Cancer Res. 8, 811–816 (2002).
91. Yang, Q. et al. Prognostic value of Bcl-2 in invasive breast cancer receiving chemotherapy and endocrine therapy. Oncol. Rep. 10, 121–125 (2003).
92. Ambudkar, S. V., Kimchi-Sarfaty, C., Sauna, Z. E. & Gottesman, M. M. P-glycoprotein: from genomics to mechanism. Oncogene 22, 7468–7485 (2003).
93. Kerr, J. F., Wyllie, A. H. & Currie, A. R. Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 26, 239–257 (1972).
94. Hanayama, R. et al. Identification of a factor that links apoptotic cells to phagocytes. Nature 417, 182–187 (2002).
AcknowledgementsI apologize to those many authors whose important work could not be included in this review owing to space constraints.
Competing interests statementThe author declares competing financial interests: see web version for details.
DATABASESEntrez Gene: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=geneAPAF1 | BAD | BAK | BAX | BBC3 | BCL2 | BCL2A1 | BCL2L1 | BCL2L11 | BCL2L2 | BID | BIK | BMF | BOK | caspase 3 | caspase 9 | CDKN2A | DIABLO | HRK | MCL1 | MYC | p53 | PMAIP1National Cancer Institute: http://www.cancer.gov/breast cancer | leukaemia | melanoma | pancreatic cancerNational Cancer Institute Drug Dictionary: http://www.cancer.gov/drugdictionary/cyclophosphamide | doxorubicin | etoposide | fludarabine | oblimersen | vincristine
FURTHER INFORMATIONAnthony G. Letai’s homepage: http://research.dfci.harvard.edu/letai/
aLLLInKSareacTIveInTHeOnLInePDF
R E V I E W S
132 | FeBRuARy 2008 | vOluMe 8 www.nature.com/reviews/cancer
© 2008 Nature Publishing Group