Embed Size (px)
Citation preview

Development/Plasticity/Repair
Sec24- and ARFGAP1-Dependent Trafficking of GABATransporter-1 Is a Prerequisite for Correct Axonal Targeting
Veronika Reiterer,1 Susanne Maier,1 Harald H. Sitte,1 Alexander Kriz,2 Markus A. Ruegg,2 Hans-Peter Hauri,2
Michael Freissmuth,1 and Hesso Farhan1
1Institute of Pharmacology, Medical University of Vienna, 1090 Vienna, Austria, and 2Biozentrum, University of Basel, 4056 Basel, Switzerland
The GABA transporter-1 (GAT1) is a prototypical protein of the synaptic specialization. Export of GAT1 from the endoplasmic reticulum(ER) is contingent on its interaction with the COPII (coatomer protein-II) coat subunit Sec24D. Here we show that silencing all four Sec24isoforms strongly inhibits transport of GAT1 to the cell surface. In contrast, transport of GAT1-RL/AS, a mutant that is deficient in Sec24Drecruitment, was not inhibited, suggesting a nonconventional, COPII-independent pathway. However, ARFGAP1 bound directly to the Cterminus of both GAT1-RL/AS and wild-type GAT1. Surface expression of GAT1-RL/AS involved ARFGAP1. GAT1-RL/AS appeared tobypass the ER-Golgi-intermediate compartment, but its pathway to the plasma membrane still involved passage through the Golgi. Thus,the GAT1-RL/AS mutant allowed to test whether COPII-dependent ER-export is required for correct sorting of GAT1 to the axon terminalin neuronal cells. In contrast to wild-type GAT1, GAT1-RL/AS failed to be specifically enriched at the tip of neurite extensions of CAD.acells (a neuroblastoma cell line that can be differentiated into a neuron-like phenotype) and in the axon terminals of hippocampalneurons. These findings indicate that correct sorting to the axon is contingent on ER export via the COPII machinery and passage throughthe ER-Golgi-intermediate compartment.
Key words: GABA transporter-1; axonal targeting; ARFGAP1; COPII; Sec24; endoplasmic reticulum export
IntroductionNeuronal differentiation involves compartmentalized sorting ofmembrane constituents to generate at least two distinct surfaces,the somatodendritic and the axonal membrane. These surfacesdiffer substantially in their composition. Nevertheless, expansionof either compartment is thought to be dependent on the secre-tory pathway, which provides for continuous flow of cargo fromthe endoplasmic reticulum to the cell surface. The basic compo-nents of the secretory pathway have been characterized in con-siderable detail. Export of proteins from the endoplasmic reticu-lum (ER) is mediated by COPII-coated vesicles (Lee et al., 2004).Assembly of the COPII coat is initiated by activation of the GT-Pase Sar1, and the sequential recruitment of Sec23-Sec24 dimersand Sec13-Sec31 heterotetramers to form the inner and outerlayer of the COPII coat, respectively. COPII vesicles bud fromspecialized regions of the ER termed ER exit sites (ERES). Inmammalian cells, COPII vesicles fuse with the ER-Golgi interme-diate compartment (ERGIC) (Appenzeller-Herzog and Hauri,2006). In addition, it was shown (in vitro) that COPII vesicles fuse
homotypically and thus may generate the ERGIC (Xu and Hay,2004). Anterograde movement of cargo also requires the COPIcoat, the assembly of which is initiated by the recruitment of theGTPase ARF1 (Lee et al., 2004).
In neuronal cells, the secretory pathway has features indicativeof specialization. Originally, ERGIC-53 [the marker protein ofthe intermediate compartment (Hauri et al., 2000)] was thoughtto be confined to the neuronal soma (Krijnse-Locker et al., 1995).However, its presence was subsequently demonstrated in den-dritic spines (Pierce et al., 2000). Similarly, Golgi outposts wereidentified in dendrites (Horton et al., 2005). Mutations have re-cently been identified, which specifically inhibit dendritic ar-borization. These included loss of function mutations of Sec23and Sar1 (Ye et al., 2007). Together, these findings are consistentwith a specialized role of the secretory pathway in dendritogen-esis. In contrast, the generation of axons is apparently COPIIindependent, because inactivation of genes encoding Sar1, Sec23,and Rab1 or downregulation of Sar1 expression does not affectthe axonal length (Ye et al., 2007).
Neurotransmitter transporters terminate synaptic transmis-sion by retrieving released neurotransmitters from the synapticcleft into the presynaptic specialization. It is evident that trans-porters have to be specifically targeted to the axonal compart-ment. However, it is not clear how GAT1 and other related SLC6family members are enriched in the axonal compartment. ERexport of GAT1 requires the formation of homooligomers(Scholze et al., 2002) and the recruitment of Sec24D to a motifcomprising 566RL 567 in its C terminus (Farhan et al., 2004, 2007).If the Sec24D binding site is mutated, the resulting GAT1-RL/ASfails to undergo COPII-dependent export from the ER, but
Received July 22, 2008; revised Oct. 8, 2008; accepted Oct. 14, 2008.This work was supported by grants from the Austrian National Bank (10507) and from the Austrian Science Fund
(FWF) P18072 and the FWF-sponsored SFB-35. H.F. is a recipient of a Schrodinger Fellowship from the FWF (J2680).We thank Julie Donaldson and Jennifer Lippincott-Schwartz for generous gifts of plasmids. We thank Dan Cassel forARFGAP1 expression vectors, for anti-ARFGAP1 antibody, and for his comments on this manuscript.
Correspondence should be addressed to Michael Freissmuth, Institute of Pharmacology, Medical University ofVienna, Waehringer Strasse 13, A-1090 Vienna, Austria. E-mail: [email protected].
V. Reiterer’s and H. Farhan’s present address: Biozentrum, University of Basel, Klingelbergstrasse 70, 4056 Basel,Switzerland.
DOI:10.1523/JNEUROSCI.3451-08.2008Copyright © 2008 Society for Neuroscience 0270-6474/08/2812453-12$15.00/0
The Journal of Neuroscience, November 19, 2008 • 28(47):12453–12464 • 12453

reaches the cell surface via an uncharacterized pathway (Farhanet al., 2007). This mutant provides an opportunity to test whetheror not targeting to the axonal compartment depends on theCOPII-machinery. Here, we show that trafficking of GAT1-RL/AS to the Golgi is COPII independent but involves ARF-GAP1. This nonconventional ER exit route, however, precludescorrect sorting to the axonal compartment.
Materials and MethodsCell culture and transfection. HEK293 cells and HeLa cells were culturedin DMEM supplemented with 10% fetal bovine serum, L-glutamine, andantibiotics. For microscopy of HEK293 cells, 3 � 10 5 cells were seeded on15 mm poly-D-lysine-coated coverslips. Transient transfections weredone with the CaPO4 precipitation method. HeLa cells were transfectedusing FuGene (Roche). When we wanted to purify 6xHis-tagged con-structs the transfection of HEK293 cells was performed with Lipo-fectamine Plus (Invitrogen). LdLF cells were cultured in Ham’s F-12medium supplemented with 5% fetal bovine serum and antibiotics. Formicroscopy, 4 � 10 5 cells were seeded on 15 mm poly-D-lysine-coatedcoverslips. Cells were transfected with Lipofectamine Plus according tothe manufacturer’s instructions. CAD cells were cultured in an 1:1 mix ofDMEM and Ham’s F-12 medium supplemented with 1% glutamine, 5%FCS, and antibiotics. For microscopy, 2.5 � 10 5 cells were seeded onpoly-D-lysine-coated coverslips. CAD cells were transiently transfectedusing Lipofectamine Plus (Invitrogen). For experiments with siRNA (allfrom Qiagen), 2 � 10 5 HeLa cells were reverse transfected with 5 nM
siRNA using HiPerFect (Qiagen) according to the manufacturer’sinstructions.
Antibodies and plasmids. The following antibodies were used: mousemonoclonal antibody against ERGIC-53 (G1/93) and rabbit polyclonalantibody against giantin were from Alexis Corporation, the anti-HA an-tibody was from Santa Cruz, the anti-His6 antibody was from Qiagen.Plasmids encoding for the following constructs were used: HA-taggedARF1 was a gift from Julie Donaldson (National Institutes of Health,Bethesda, MD), His-tagged ARFGAP1 was a gift from Dan Cassel (Tech-nion, Haifa, Israel), and YFP-tagged ARFGAP1 and �64ARFGAP1 weregifts from Jennifer Lippincott-Schwartz (National Institutes of Health,Bethesda, MD).
Assay for [3H]GABA uptake. Uptake of [ 3H]GABA was determined asdescribed in detail previously (Scholze et al., 2002). In brief, transiently
transfected HEK293 cells (3 � 10 5/well) were seeded into 48 well dishes24 h after transfection; after 24 h, the cells were incubated in the presenceof 20 �M [ 3H]GABA (specific activity �83 cpm/pmol) for 3 min; non-specific uptake was defined as the amount of cell associated radioactivitydetermined in the presence of 100 �M tiagabine.
Confocal fluorescence microscopy and FRAP. Fluorescence images wereacquired with a 40� oil-immersion objective with a Zeiss LSM510 con-focal microscope and digitally captured. The images were converted toTIF files and analyzed using the ImagePro Plus software (Media Cyber-netics). The colored images were converted to 12-bit gray scale images.Under these conditions the software recognizes 4095 potential gray val-ues. The contrast was enhanced by excluding the lower 20% of the grayscale. For quantifying the accumulation of proteins in the ERGIC andtheir colocalization (see Figs. 3, 11), all bright objects that were between1.5 and 10 pixels were defined as peripheral ERGIC structures. The sameprocedure was used to estimate the extent of colocalization in neurons,
Figure 1. Depletion of Sec24-isoforms reduces the amount of wild-type GAT1 but not ofmutant GAT1-RL/AS at the cell surface. A, HeLa cells (2 � 10 5) plated into 6-well plates werereverse transfected with control siRNA (Co) or with a mix of siRNAs against Sec24A, Sec24B,Sec24C, and Sec24D (si). Total amount of oligonucleotides was the same in both transfections(260 ng). Cells were collected and lysed 48 and 72 h after transfection, and immunoblots forindividual Sec24 isoforms were performed. The numbers on the right side indicate the densito-metrically estimated extent (using ImageJ software) of knockdown in percentage of control at72 h; in all instances (n � 3), levels of Sec24A and Sec24B were �� 5%; levels of Sec24C andSec24D were 10 � 8.6 and 15 � 5.7% of control at 72 h (means � SD). B, HeLa cells werepretreated with siRNAs as outlined for A. After 48 h, cells were transfected with a plasmidencoding either YFP-tagged wild-type GAT1 (WT) or YFP-tagged GAT1-RL/AS (RL). Eight hoursthereafter, the content of each well was transferred to seven wells of a 24-well plate. Uptake of[ 3H]GABA (specific activity �15 cpm/pmol) was performed 24 h after plasmid transfection.The total GABA concentration was 5.5 �M. Incubation time was 3 min, and the experiment wasperformed at room temperature (�24°C). Nonspecific uptake was measured in the presence of10 �M tiagabine. Data are means � SEM (n � 3).
Figure 2. Effect of temperature on the subcellular distribution of GAT1 and GAT1-RL/AS. A,HEK293 cells expressing YFP-tagged GAT1 or YFP-tagged GAT1-RL/AS were kept either at 37°Cor incubated at 15°C for 3 h. Cells were fixed with 4% paraformaldehyde and imaged subse-quently. The number of cells containing fluorescence in punctate structures were scored; thesenumbers were 0/20 and 16/26 for YFP-tagged GAT1 and 0/20 and 0/18 for YFP-tagged GAT1-RL/AS at 37° and 15°, respectively (data from 3 independent experiments). B, ldlF cells express-ing YFP-tagged GAT1 (top row) or YFP-tagged GAT1-RL/AS were either kept at 34°C (permissivetemperature) or incubated for 5 h at 40°C (restrictive temperature) and subsequently fixed with4% paraformaldehyde. Images were captured by confocal microscopy. The number of cellscontaining fluorescence in punctate structures were scored: these numbers were 0/20 and20/20 for YFP-tagged GAT1 and 0/20 and 0/20 for YFP-tagged GAT1-RL/AS at 34° and 42°,respectively (data from 3 independent experiments).
12454 • J. Neurosci., November 19, 2008 • 28(47):12453–12464 Reiterer et al. • Trafficking of GAT1 in the Early Secretory Pathway

where larger visual field were examined, except that no upper limit wasimposed, i.e., structures larger than 1.5 pixels were quantified. Fluores-cence recovery after photobleaching (FRAP) was also recorded on a ZeissLSM510 confocal laser scanning microscope. Transfected cells were ex-amined with an oil-immersion objective (40-fold magnification). Circu-lar regions of interest were specified for bleaching and scanning (bleachregions). Before bleaching, a scan was obtained; the area of interest was
then bleached with 70 iterations at maximumlaser power (30 mW, at 514 nm). Subsequently,pictures were captured (40 scans in 40 s) with�4% of laser power. The recorded fluorescenceintensities were digitized and averaged over thebleach region using ScionImage 4.0.2; these val-ues were plotted and subjected to nonlinearcurve fitting to the equation describing themonoexponential rise to a maximum: y � bot-tom � (top � bottom) � (1 � e �kx), where“bottom” refers to the relative fluorescence in-tensity immediately after bleaching (before re-covery) and “top” to the value that is reached atequilibrium (after recovery). Imaging of neu-rons was performed with a LeicaSPE confocallaser scanning microscope using the 40� oil-immersion objective.
Purification of GST-tagged proteins, GST-pulldown and purification, and His6-taggedARFGAP1 and ARF. Purification of GST-taggedproteins and GST pulldown have been de-scribed in detail previously (Farhan et al.,2007). For purification of His6-ARF1 and His6-ARFGAP1, HEK293 cells were transfected withplasmids encoding either His6-ARF1 or His6-ARFGAP1. The transfection was performedwith Lipofectamine Plus (Invitrogen). Twenty-four hours after transfection, cells werescrapped down and lysed in lysis buffer (300mM NaCl, 50 mM NaH2PO4, 10 mM imidazole,pH adjusted to 7.5 with HCl, and 0.05% Tween20) and then sonicated 4 times in a water bathfor 10 s. The reaction was then centrifuged at10,000 � g for 10 min. The supernatant wasincubated with Ni-NTA Magnetic Agarosebeads (Qiagen) for 90 min. Beads were washedtwice in wash buffer (300 mM NaCl, 50 mM
NaH2PO4, 20 mM imidazole, pH adjusted to 7.5with HCl, and 0.05% Tween 20). The proteinwas eluted in elution buffer (300 mM NaCl, 50mM NaH2PO4, 250 mM imidazole, pH adjustedto 7.5 with HCl, and 0.05% Tween 20).
Preparation of rat hippocampal neurons.High-density cultures (�5 � 10 4 cells/cm 2)were established from 18-d-old Wistar rat hip-pocampi. The fetal hippocampi were removedfrom the brain, washed once in HBSS, andtrypsinized for 15 min at 37°C. Thereafter, hip-pocampi were washed again in HBSS followedby additional washes in prewarmed plating me-dium (DMEM/Glutamax, 0.6% glucose, 10%fetal calf serum, and antibiotics). Finally, thehippocampi were dissociated in the plating me-dium on glass coverslips (precoated with poly-L-lysine; 1 mg/ml in borate buffer, pH 8.5;coated overnight and washed twice with water).After 3 h from the initial seeding the mediumwas changed to Neurobasal medium (Invitro-gen) supplemented with 0.5 mM glutamine, an-tibiotics, and B27 supplement (Invitrogen).Neurons were transfected after 7 d in vitro usingLipofectamine 2000 (Invitrogen) according tothe manufacturer’s instructions.
Immunoprecipitation experiments. Brains from adult rats (cerebrum,cerebellum, and medulla oblongata) were homogenized in lysis buffer(50 mM Tris.HCl, pH 7.4, 100 mM NaCl, 1% Triton X-100) supplementedwith protease inhibitors. The homogenate was centrifuged at 10,000 � gfor 10 min at 4°C. The supernatant was recentrifuged at 25,000 � g foradditional 10 min at 4°C. Anti-GAT1 antibody (ab426 from Abcam) was
Figure 3. Effect of temperature on localization of GAT1 in the ERGIC in the presence or absence of Sec24. HeLa cells weretransfected with a control siRNA (A) or siRNAs against Sec24A-D (B). The total amount of siRNA and the amount of transfectionreagent was the same. After 48 h, cells were transfected with plasmids encoding YFP-tagged GAT1. On the next day, cells wereeither fixed directly (top rows in A and B) or after an incubation for 3 h at 15°C (bottom rows in A and B) followed by immuno-staining for ERGIC-53. Images were acquired with a confocal microscope. C, The total number of ERGIC vesicles was quantified bycounting the ERGIC-53 positive structures using ImagePro Plus software as outlined in Materials and Methods. The number ofERGIC53 vesicles that contained YFP-GAT1 was also quantified in the same cells; numbers above the bars indicate the percentageof vesicles containing YFP-GAT1. Data are from three independent transfections.
Reiterer et al. • Trafficking of GAT1 in the Early Secretory Pathway J. Neurosci., November 19, 2008 • 28(47):12453–12464 • 12455

added to the supernatant (1 ml), and the samplewas rotated overnight at 4°C. On the next dayprotein G beads (50 �l of 1:1 slurry) were addedfor additional 3 h. After washing the beads threetimes in lysis buffer, proteins were released byboiling for 5 min in sample buffer. The elutedproteins were subjected to SDS-PAGE.
ResultsCOPII depletion does not affecttrafficking of GAT1-RL/AS to theplasma membrane.Our previous observation that GAT1-RL/AS is unable to bind to Sec24D (Farhanet al., 2007) raises the possibility that itsexocytic trafficking occurs in a COPII-independent manner. Alternatively,GAT1-RL/AS may be incorporated by de-fault into COPII vesicles. To test for COPIIdependence, we performed a quadrupleknockdown of all Sec24 isoforms (Wen-deler et al., 2007) and estimated the pre-sumptive surface expression of GAT1 by[ 3H]GABA uptake. We chose HeLa cellsbecause of their efficient uptake of siRNA(Wendeler et al., 2007). Sec24 isoformswere efficiently knocked down (Fig. 1A).GAT1 was transfected 48 h after knock-down, and [ 3H]GABA uptake was mea-sured 72 h after knockdown. Of note,Sec24 levels were strongly reduced alreadyafter 48 h, indicating that when these cellsstarted to express GAT1, they already hadlow levels of Sec24 which continued to below to the day of the uptake experiment.As expected, the quadruple knockdown ofSec24 isoforms strongly reduced surfaceexpression of wild-type GAT1 down to alevel comparable to GAT1-RL/AS (Fig.1B). In contrast, the level of GAT1-RL/ASsurface expression was not affected by theknockdown.
Given the inability of GAT1-RL/AS tointeract with Sec24 and the strongly re-duced cellular Sec24 levels, it becomes veryunlikely that anterograde trafficking ofGAT1-RL/AS is COPII dependent. It rather appears that the mu-tant exits by another mechanism, which, for the lack of betterunderstanding, we will refer to as a nonconventional, defaultpathway.
GAT1-RL/AS cannot be trapped in the ERGICIt was of interest to understand the route taken by vesicles subjectto this nonconventional ER exit, specifically whether the vesiclesalso reach the ERGIC. Using an in vitro system, it was shown thatCOPII vesicles homotypically fuse (Xu and Hay, 2004). This wasproposed to be a mechanism for de novo formation of the ERGIC(Xu and Hay, 2004). Alternatively, COPII vesicles may fuse het-erotypically with the ERGIC. In any case trafficking of a proteinthrough the ERGIC requires it to be incorporated into COPIIvesicles.
Many secretory proteins can be trapped in the ERGIC by low-ering the temperature to 15°C (Saraste and Kuismanen, 1984,Schweizer et al., 1990). Incubation of HEK293 cells at 15°C for 3 h
caused retention of wild-type GAT1 in punctate structures thatstained for ERGIC-53 (Fig. 2A). In contrast, the distribution pat-tern of GAT1-RL/AS was not affected by low temperature (Fig.2A). An alternative way to arrest protein transport in the ERGICis to inactivate COPI function (Pepperkok et al., 1993). This con-dition can be probed with ldlF cells which express a temperature-sensitive mutant of �-COP that misfolds at the elevated temper-ature of 40°C (restricted temperature) and thereby bocks COPIfunction (Guo et al., 1994). Protein transport at 34°C (permissivetemperature) is normal in ldlF cells. At the permissive temperature,wild-type GAT1 was transported to the plasma membrane, whereasGAT1-RL/AS was predominantly in the ER (Fig. 2B). When cellswere incubated at 40°C for 4 h, wild-type GAT1 accumulated inpunctate structures (Fig. 2B) that stained for ERGIC-53 (data notshown). In contrast, the temperature shift did not affect the distri-bution of GAT1-RL/AS (Fig. 2B, bottom row). Thus, the noncon-ventional route taken by GAT1-RL/AS is most readily explained by atransport pathway that bypasses the ERGIC.
Figure 4. Effect of temperature on delivery of GAT1 and GAT-RL/AS to the Golgi apparatus. A, B, HEK293 cells coexpressingCFP-tagged GAT1-RL/AS (A) or GAT1 (B) together with YFP-Golgi were incubated at 20°C for 5 h, fixed, and imaged subsequently.C, D, HeLa cells were reverse transfected with siRNAs against Sec24A-D (B) or with control siRNA (C). After 48 h, cells weretransfected with a plasmid encoding YFP-tagged GAT1. On the next day, cells were incubated at 20°C for 5 h and fixed subse-quently. In C and D, the Golgi apparatus was detected using immunofluorescence staining against giantin. Colocalization with theGolgi marker was 100% at 20°C (coded printouts of 20 cells from 3 independent experiments scored by an unbiased observer).
12456 • J. Neurosci., November 19, 2008 • 28(47):12453–12464 Reiterer et al. • Trafficking of GAT1 in the Early Secretory Pathway
If GAT1-RL/AS was not concentrated in the ERGIC becauseof its inability to bind Sec24, it should in principle be possible tomimic the situation for wild-type GAT1 by depletion of Sec24.We tested this prediction by silencing all four Sec24 isoforms.Accumulation of GAT1 in the ERGIC was assessed by colocaliza-tion with ERGIC-53. In cells transfected with the control siRNA,wild-type GAT1 was seen at the cell surface (Fig. 3A, top row). Ifcells were incubated at 15°C for 3 h, GAT1 was found in punctateERGIC-53 positive structures (Fig. 3A, bottom row). After Sec24silencing ERGIC-53 showed an ER pattern and only few punctatestructures were seen (Fig. 3B, top row, middle image). AlthoughGAT1 was predominantly retained in the ER in these cells, afraction of it was still transported to the plasma membrane (Fig.3B, top row, leftmost image). The distribution pattern of GAT1did not change, if cells deficient in Sec24 isoforms were placed at15°C for 3 h (Fig. 3B). Most importantly, the occasional ERGIC-53-containing punctate structures were devoid of GAT1 (overlayin bottom right panel of Fig. 3B). These results suggest that afraction of GAT1 can take the alternative, nonconventional routeto the plasma membrane in cells in which Sec24 is silenced.
The pathway of GAT1-RL/AS involves the GolgiDoes GAT1-RL/AS pass through the Golgi on its way to the cellsurface? We addressed this question by imposing a 20°C temper-ature block known to inhibit exocytic traffic from the Golgi (Grif-fiths et al., 1985). HEK293 cells were cotransfected with a plasmidencoding YFP-Golgi together with either CFP-tagged wild-typeGAT1 or CFP-tagged GAT1-RL/AS. On the following day, thecells were incubated at 20°C for 5 h. Both proteins were found tobe retained in the Golgi under these conditions (Fig. 4A,B). Thedata shown in Figure 3 suggested that, in the absence of all Sec24isoforms, a fraction of wild-type GAT1 also used the nonconven-
tional default pathway. We analyzedwhether GAT1 reaches the Golgi in Sec24-silenced cells. Figure 4, C and D, showsthat GAT1 indeed reaches the Golgi underthese conditions at 20°C. We concludethat the nonconventional default pathwayinvolves the Golgi.
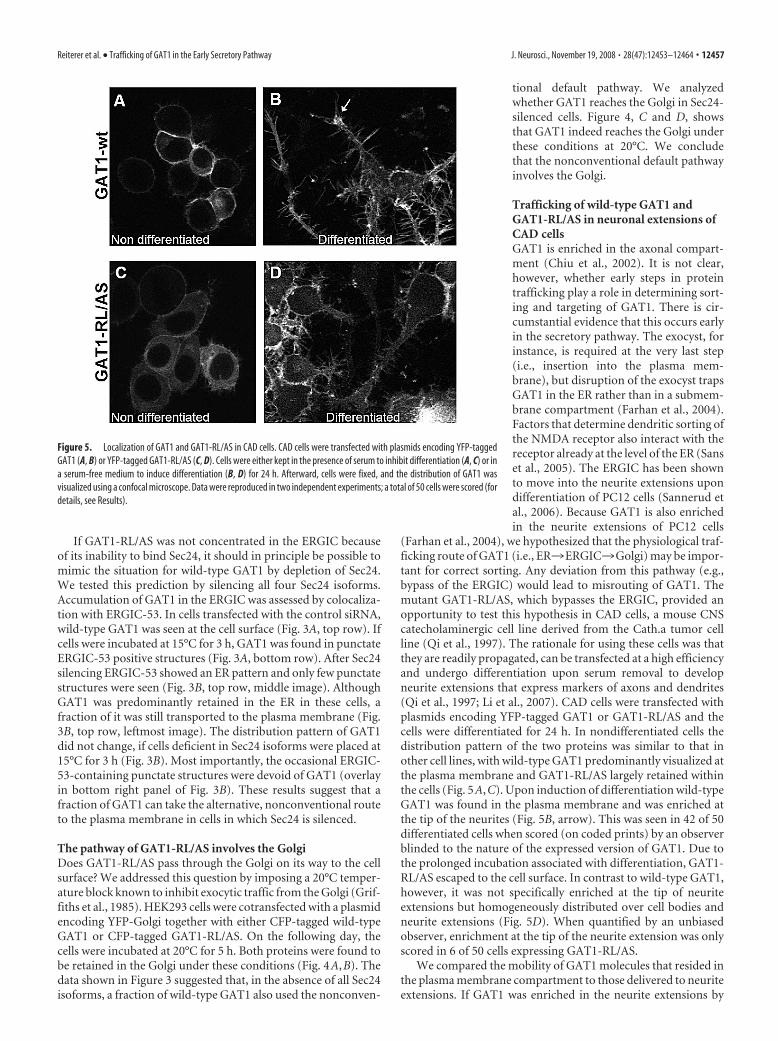
Trafficking of wild-type GAT1 andGAT1-RL/AS in neuronal extensions ofCAD cellsGAT1 is enriched in the axonal compart-ment (Chiu et al., 2002). It is not clear,however, whether early steps in proteintrafficking play a role in determining sort-ing and targeting of GAT1. There is cir-cumstantial evidence that this occurs earlyin the secretory pathway. The exocyst, forinstance, is required at the very last step(i.e., insertion into the plasma mem-brane), but disruption of the exocyst trapsGAT1 in the ER rather than in a submem-brane compartment (Farhan et al., 2004).Factors that determine dendritic sorting ofthe NMDA receptor also interact with thereceptor already at the level of the ER (Sanset al., 2005). The ERGIC has been shownto move into the neurite extensions upondifferentiation of PC12 cells (Sannerud etal., 2006). Because GAT1 is also enrichedin the neurite extensions of PC12 cells
(Farhan et al., 2004), we hypothesized that the physiological traf-ficking route of GAT1 (i.e., ER3ERGIC3Golgi) may be impor-tant for correct sorting. Any deviation from this pathway (e.g.,bypass of the ERGIC) would lead to misrouting of GAT1. Themutant GAT1-RL/AS, which bypasses the ERGIC, provided anopportunity to test this hypothesis in CAD cells, a mouse CNScatecholaminergic cell line derived from the Cath.a tumor cellline (Qi et al., 1997). The rationale for using these cells was thatthey are readily propagated, can be transfected at a high efficiencyand undergo differentiation upon serum removal to developneurite extensions that express markers of axons and dendrites(Qi et al., 1997; Li et al., 2007). CAD cells were transfected withplasmids encoding YFP-tagged GAT1 or GAT1-RL/AS and thecells were differentiated for 24 h. In nondifferentiated cells thedistribution pattern of the two proteins was similar to that inother cell lines, with wild-type GAT1 predominantly visualized atthe plasma membrane and GAT1-RL/AS largely retained withinthe cells (Fig. 5A,C). Upon induction of differentiation wild-typeGAT1 was found in the plasma membrane and was enriched atthe tip of the neurites (Fig. 5B, arrow). This was seen in 42 of 50differentiated cells when scored (on coded prints) by an observerblinded to the nature of the expressed version of GAT1. Due tothe prolonged incubation associated with differentiation, GAT1-RL/AS escaped to the cell surface. In contrast to wild-type GAT1,however, it was not specifically enriched at the tip of neuriteextensions but homogeneously distributed over cell bodies andneurite extensions (Fig. 5D). When quantified by an unbiasedobserver, enrichment at the tip of the neurite extension was onlyscored in 6 of 50 cells expressing GAT1-RL/AS.
We compared the mobility of GAT1 molecules that resided inthe plasma membrane compartment to those delivered to neuriteextensions. If GAT1 was enriched in the neurite extensions by
Figure 5. Localization of GAT1 and GAT1-RL/AS in CAD cells. CAD cells were transfected with plasmids encoding YFP-taggedGAT1 (A, B) or YFP-tagged GAT1-RL/AS (C, D). Cells were either kept in the presence of serum to inhibit differentiation (A, C) or ina serum-free medium to induce differentiation (B, D) for 24 h. Afterward, cells were fixed, and the distribution of GAT1 wasvisualized using a confocal microscope. Data were reproduced in two independent experiments; a total of 50 cells were scored (fordetails, see Results).
Reiterer et al. • Trafficking of GAT1 in the Early Secretory Pathway J. Neurosci., November 19, 2008 • 28(47):12453–12464 • 12457

active targeting/sorting and specific reten-tion, it was predicted to be less mobile. Weverified this conjecture by fluorescence re-covery after photobleaching (FRAP) ex-periments: CAD cells were transfectedwith plasmids encoding YFP-tagged GAT1and cells were differentiated by serumwithdrawal for 24 h. Regions of interestwere selected over the neurite (Fig. 6A)and over the cell body (Fig. 6B) andbleached. The intensely fluorescent accu-mulations in the neurite extensions didnot recover over the time course of the ex-periment (Fig. 6A,B). In addition, the mo-bile fraction was higher in somatic regionsthan in neurites and this difference was sta-tistically significant (Fig. 6C) (21.2 � 3.9%versus 32.2 � 4.2%—mean � SD; n � 3;p � 0.028—for neurite and somatic region,respectively). Based on these findings, weconclude that GAT1 is specifically sorted tothe neurite extensions to accumulate in spe-cific areas such that it cannot exchange rap-idly with the pool of transporter in the othersurface membrane compartments.
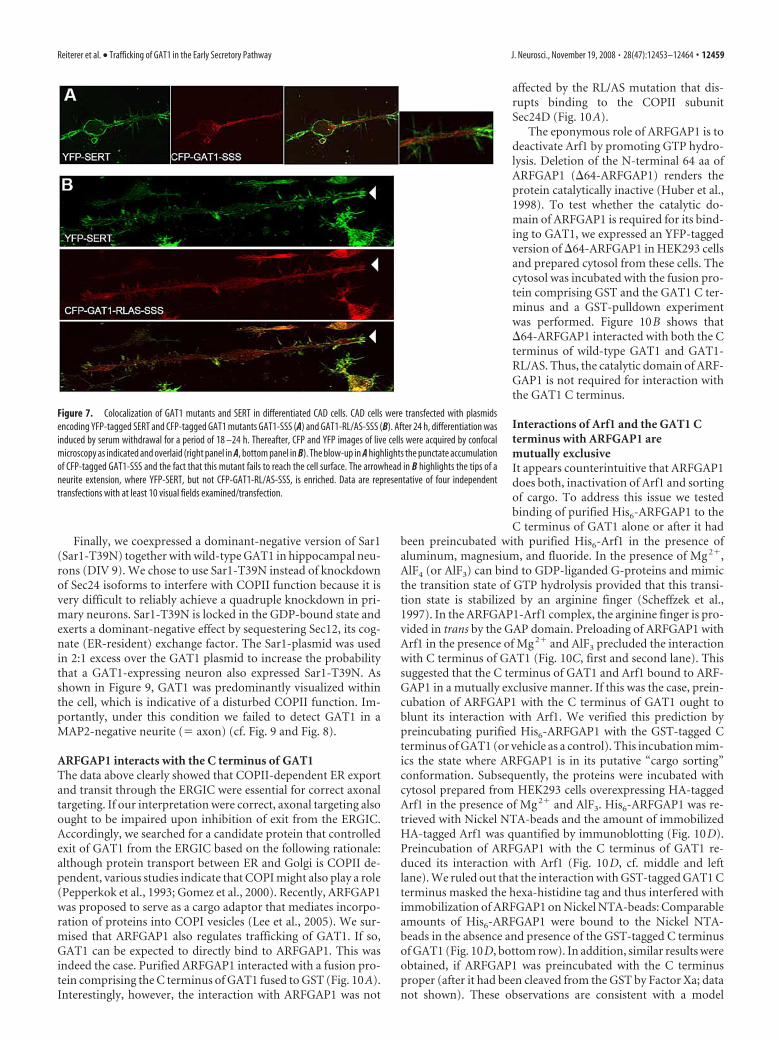
The RL/AS-mutation acts as a secondsite suppressor for an ERGIC-trappedGAT1 mutantIf the tri-hydrophobic motif 569VMI 571 inthe C terminus of GAT1 is replaced bythree serines, the resulting GAT1-SSS un-dergoes COPII-dependent export but be-comes trapped in the ERGIC and does notreach the cell surface (Farhan et al., 2008).This was also recapitulated in CAD cells(Fig. 7A): we coexpressed CFP-taggedGAT1-SSS and YFP-tagged serotonintransporter (SERT). The latter was used to unequivocally visual-ize the cell surface and in particular the sprouting neuronal ex-tensions and spikes. It is evident from Figure 7A that GAT1-SSSremained in punctate structures, while the serotonin transporterresided at the cell surface and was enriched on filopodia-likeprojections arising from the developing neurite extensions. In allcells examined (including neurons), GAT1-SSS is trapped in theERGIC after undergoing (COPII-dependent) ER export (Farhanet al., 2008). Because the RL/AS mutation prevents COPII re-cruitment and thus COPII-dependent export, we surmised that amutation of the RL-motif should act as a second site suppressorwithin the GAT1-SSS background. This was the case: while thebulk of the double mutant GAT1-RL/AS-SSS was diffusely dis-tributed within the cell, a fraction of this transporter did escape to thecell surface (Fig. 7B, middle panel). However, in contrast to the co-expressed SERT (Fig. 7B, top panel), GAT1-RL/AS-SSS was not en-riched at the tips of the growing neurite extensions (marked by ar-rowhead, in Fig. 7B), where the green pseudocolor representingSERT was dominant in the overlay (Fig. 7B, bottom panel).
Lack of axonal enrichment of GAT1-RL/AS inhippocampal neuronsWhile neuroblastoma and related cells can differentiate into aphenotype resembling catecholaminergic neurons, the nature oftheir neurite extensions is difficult to assess; in fact, their neuritic
extensions contain markers of both axons and dendrites (Li et al.,2007). Therefore, we investigated trafficking of wild-type GAT1and GAT1-RL/AS in rat hippocampal neurons prepared fromembryonic rats (E17). After 7 d in culture neurons were trans-fected with the indicated plasmids and fixed after 48 h. Immuno-fluorescent labeling of MAP2 was performed to stain dendrites.Processes that were negative for MAP2 were considered axons.While wild-type GAT1 nicely decorated the plasma membrane(Fig. 8A), GAT1-RL/AS was found predominantly in the ER anda minor fraction of it was found at the plasma membrane (Fig.8B). The axons terminal is the site of most active membraneaddition. When we inspected axons we found that wild-typeGAT1 was strongly enriched in the axon terminal (Fig. 8C), whileGAT1-RL/AS was not (Fig. 8D). To quantify the magnitude ofthis differential enrichment, we compared the fluorescence in-tensity of the axon terminal with the axon which is immediatelyadjacent to it. The measured fluorescence intensity was dividedby the area to extract the density of YFP molecules (i.e., GAT1 orits mutant). Because we observed that neuronal processes runthrough several focus planes, we measured the density of fluores-cent molecules in 4 different confocal planes, each separated 200nm from the next. Using this approach we found that the densityof wild-type GAT1 was �2-fold higher in the axon terminal thanin more proximal parts (Fig. 8E). No such difference was ob-served with GAT1-RL/AS (Fig. 8F).
Figure 6. Mobility of GAT1 in the plasma membrane of differentiated CAD cells. CAD cells were transfected with plasmidsencoding YFP-tagged GAT1. After 24 h, differentiation was induced by serum withdrawal for a period of 18 –24 h. FRAP experi-ments were performed as described in Materials and Methods. A and B show a representative cell in which first the neurite (A) wasbleached and followed for 38 s to detect fluorescence recovery. Afterward, a region on the soma was bleached (B) and followed for38 s to detect fluorescence recovery. C, Time-dependent fluorescence recovery after photobleaching of a membrane strip on thesoma (triangles) and the tip of neurite extensions (squares) expressed as a percentage of prebleach fluorescence intensity. Dataare means � SEM (n � 3).
12458 • J. Neurosci., November 19, 2008 • 28(47):12453–12464 Reiterer et al. • Trafficking of GAT1 in the Early Secretory Pathway
Finally, we coexpressed a dominant-negative version of Sar1(Sar1-T39N) together with wild-type GAT1 in hippocampal neu-rons (DIV 9). We chose to use Sar1-T39N instead of knockdownof Sec24 isoforms to interfere with COPII function because it isvery difficult to reliably achieve a quadruple knockdown in pri-mary neurons. Sar1-T39N is locked in the GDP-bound state andexerts a dominant-negative effect by sequestering Sec12, its cog-nate (ER-resident) exchange factor. The Sar1-plasmid was usedin 2:1 excess over the GAT1 plasmid to increase the probabilitythat a GAT1-expressing neuron also expressed Sar1-T39N. Asshown in Figure 9, GAT1 was predominantly visualized withinthe cell, which is indicative of a disturbed COPII function. Im-portantly, under this condition we failed to detect GAT1 in aMAP2-negative neurite (� axon) (cf. Fig. 9 and Fig. 8).
ARFGAP1 interacts with the C terminus of GAT1The data above clearly showed that COPII-dependent ER exportand transit through the ERGIC were essential for correct axonaltargeting. If our interpretation were correct, axonal targeting alsoought to be impaired upon inhibition of exit from the ERGIC.Accordingly, we searched for a candidate protein that controlledexit of GAT1 from the ERGIC based on the following rationale:although protein transport between ER and Golgi is COPII de-pendent, various studies indicate that COPI might also play a role(Pepperkok et al., 1993; Gomez et al., 2000). Recently, ARFGAP1was proposed to serve as a cargo adaptor that mediates incorpo-ration of proteins into COPI vesicles (Lee et al., 2005). We sur-mised that ARFGAP1 also regulates trafficking of GAT1. If so,GAT1 can be expected to directly bind to ARFGAP1. This wasindeed the case. Purified ARFGAP1 interacted with a fusion pro-tein comprising the C terminus of GAT1 fused to GST (Fig. 10A).Interestingly, however, the interaction with ARFGAP1 was not
affected by the RL/AS mutation that dis-rupts binding to the COPII subunitSec24D (Fig. 10A).
The eponymous role of ARFGAP1 is todeactivate Arf1 by promoting GTP hydro-lysis. Deletion of the N-terminal 64 aa ofARFGAP1 (�64-ARFGAP1) renders theprotein catalytically inactive (Huber et al.,1998). To test whether the catalytic do-main of ARFGAP1 is required for its bind-ing to GAT1, we expressed an YFP-taggedversion of �64-ARFGAP1 in HEK293 cellsand prepared cytosol from these cells. Thecytosol was incubated with the fusion pro-tein comprising GST and the GAT1 C ter-minus and a GST-pulldown experimentwas performed. Figure 10B shows that�64-ARFGAP1 interacted with both the Cterminus of wild-type GAT1 and GAT1-RL/AS. Thus, the catalytic domain of ARF-GAP1 is not required for interaction withthe GAT1 C terminus.
Interactions of Arf1 and the GAT1 Cterminus with ARFGAP1 aremutually exclusiveIt appears counterintuitive that ARFGAP1does both, inactivation of Arf1 and sortingof cargo. To address this issue we testedbinding of purified His6-ARFGAP1 to theC terminus of GAT1 alone or after it had
been preincubated with purified His6-Arf1 in the presence ofaluminum, magnesium, and fluoride. In the presence of Mg 2�,AlF4 (or AlF3) can bind to GDP-liganded G-proteins and mimicthe transition state of GTP hydrolysis provided that this transi-tion state is stabilized by an arginine finger (Scheffzek et al.,1997). In the ARFGAP1-Arf1 complex, the arginine finger is pro-vided in trans by the GAP domain. Preloading of ARFGAP1 withArf1 in the presence of Mg 2� and AlF3 precluded the interactionwith C terminus of GAT1 (Fig. 10C, first and second lane). Thissuggested that the C terminus of GAT1 and Arf1 bound to ARF-GAP1 in a mutually exclusive manner. If this was the case, prein-cubation of ARFGAP1 with the C terminus of GAT1 ought toblunt its interaction with Arf1. We verified this prediction bypreincubating purified His6-ARFGAP1 with the GST-tagged Cterminus of GAT1 (or vehicle as a control). This incubation mim-ics the state where ARFGAP1 is in its putative “cargo sorting”conformation. Subsequently, the proteins were incubated withcytosol prepared from HEK293 cells overexpressing HA-taggedArf1 in the presence of Mg 2� and AlF3. His6-ARFGAP1 was re-trieved with Nickel NTA-beads and the amount of immobilizedHA-tagged Arf1 was quantified by immunoblotting (Fig. 10D).Preincubation of ARFGAP1 with the C terminus of GAT1 re-duced its interaction with Arf1 (Fig. 10D, cf. middle and leftlane). We ruled out that the interaction with GST-tagged GAT1 Cterminus masked the hexa-histidine tag and thus interfered withimmobilization of ARFGAP1 on Nickel NTA-beads: Comparableamounts of His6-ARFGAP1 were bound to the Nickel NTA-beads in the absence and presence of the GST-tagged C terminusof GAT1 (Fig. 10D, bottom row). In addition, similar results wereobtained, if ARFGAP1 was preincubated with the C terminusproper (after it had been cleaved from the GST by Factor Xa; datanot shown). These observations are consistent with a model
Figure 7. Colocalization of GAT1 mutants and SERT in differentiated CAD cells. CAD cells were transfected with plasmidsencoding YFP-tagged SERT and CFP-tagged GAT1 mutants GAT1-SSS (A) and GAT1-RL/AS-SSS (B). After 24 h, differentiation wasinduced by serum withdrawal for a period of 18 –24 h. Thereafter, CFP and YFP images of live cells were acquired by confocalmicroscopy as indicated and overlaid (right panel in A, bottom panel in B). The blow-up in A highlights the punctate accumulationof CFP-tagged GAT1-SSS and the fact that this mutant fails to reach the cell surface. The arrowhead in B highlights the tips of aneurite extension, where YFP-SERT, but not CFP-GAT1-RL/AS-SSS, is enriched. Data are representative of four independenttransfections with at least 10 visual fields examined/transfection.
Reiterer et al. • Trafficking of GAT1 in the Early Secretory Pathway J. Neurosci., November 19, 2008 • 28(47):12453–12464 • 12459

where ARFGAP1 interacts directly withthe cargo GAT1 and where this interactionprecedes the deactivation of Arf1.
Interaction of GAT1 and ARFGAP1 innative tissueThe bulk of GAT1 in the brain resides atthe plasma membrane, but there must be afraction of the protein that, at steady state,is in the secretory pathway to account forthe rate of protein turnover. This portionof GAT1 must be associated with compo-nents of the secretory machinery includingARFGAP1, if this interaction was relevantin vivo. We therefore prepared lysates fromadult rat brain and immunoprecipitatedGAT1 (Fig. 10E, middle row). Sec24D wasfound in a complex with GAT1 (Fig. 10E,top row) as predicted from our previouswork (Farhan et al., 2004, 2007). In addi-tion and importantly, ARFGAP1 was alsocoimmunoprecipitated in a complex withGAT1 (Fig. 10E, bottom row). This showsthat the interaction between GAT1 andARFGAP1 (and Sec24D) does take place inthe brain and therefore is most likely ofphysiological relevance.
ARFGAP1 controls trafficking ofwild-type GAT1Incorporation of cargo into COPI vesiclesis strictly dependent on GTP hydrolysis(Nickel et al., 1998; Pepperkok et al.,2000). The above results suggested a rolefor ARFGAP1 in trafficking of GAT1.Since ERGIC to Golgi transport requiresCOPI we expect �64-ARFGAP1 to arresttrafficking of wild-type GAT1 in the ER-GIC. We first tested whether �64-ARFGAP1 is associated with the ERGIC.Cells were transfected with YFP-tagged�64-ARFGAP1 and stained for ERGIC-53. �64-ARFGAP1 was found in punctatestructures that colocalized with ERGIC-53(Fig. 11A). Coexpression of YFP-tagged�64-ARFGAP1 with the CFP-tagged wild-type GAT1 led to retention of the latter inpunctate structures which contained bothproteins (Fig. 11B). Membrane recruit-ment of ARFGAP1 can be blocked by in-cubating cells in the presence of IC261, acasein kinase inhibitor (Yu and Roth,2002). When HEK293 cells were treatedwith IC261 for 2 h, GAT1 accumulated inpunctate structures that costained forERGIC-53 (Fig. 11C). Together, these re-sults suggest that ARFGAP1 is required forexit of wild-type GAT1 from the ERGIC.
ARFGAP1 controls trafficking of GAT1 in neuronsIt was of interest to document whether ARFGAP1 controls traf-ficking of (endogenous) GAT1 in neurons. We therefore overex-pressed �64-ARFGAP1 and immunostained for GAT1 using the
same antibody used for immunoprecipitations. This antibodyhas been used by others for immunohistochemical stainings ofGAT1 (Manzano et al., 2007). GAT1 was expressed on the plasmamembrane in cells that did not express �64-ARFGAP1 and was
Figure 8. Localization of GAT1 and GAT1-RL/AS in hippocampal neurons. Hippocampal neurons were prepared from ratembryos (E17). After 7 d in culture, neurons were transfected with plasmids encoding YFP-tagged GAT1 (A, C) or YFP-taggedGAT1-RL/AS (B, D). Cells were fixed 48 h after transfection, and images were acquired using a confocal microscope. A and B showthe neuronal soma with dendrites and the axon which is indicated by white arrowheads. C and D show an axon with its terminalcontaining wild-type GAT1 or GAT1-RL/AS, respectively. E and F show an evaluation of fluorescence intensity of the axon (an areaadjacent to the terminal) versus the axon terminal in neurons expressing wild-type GAT1 (E) or GAT1-RL/AS (F ). The fluorescencewas recorded in four confocal planes separated by 200 nm. The fluorescence intensities of these four individual confocal planeswere averaged and treated as a single determination. This analysis was performed for 6 and 5 neurons expressing wild-type GAT1and GAT1-RL/AS, respectively. To account for variability in pixel intensity between individual neurons, fluorescence intensity inthe terminal was expressed as a percentage of the intensity in axon. Shown are the means � SD (n � 5 or 6). Asterisk indicatesstatistically significant difference ( p � 0.01; Student’s t test).
Figure 9. Localization of wild-type YFP-GAT1 in the absence of COPII function in neurons. Hippocampal neurons were preparedfrom rat embryos (E17). After 9 d in culture, neurons were transfected with plasmids encoding for YFP-tagged GAT1 and Sar1-T39N. Cells were fixed after 24 h and immunostained for MAP2. Images were captured by confocal microscopy under settings thatallow for visualization of YFP-tagged GAT1 (left panel) and MAP2 immunoreactivity (middle panel). The right panel was gener-ated by overlaying the pictures. Data are representative of three independent transfections.
12460 • J. Neurosci., November 19, 2008 • 28(47):12453–12464 Reiterer et al. • Trafficking of GAT1 in the Early Secretory Pathway

found there in the soma and in one neurite at high density (Fig.12A). In cells expressing �64-ARFGAP1, GAT1 and �64-ARFGAP1 coaccumulated in punctate structures (Fig. 12B). To-gether, these data show that ARFGAP1 controls neuronal traf-ficking of GAT1.
DiscussionIn the current work, we used GAT1 as a model cargo to study therelevance of the early secretory pathway for sorting decisions ofmembrane proteins. There are several advantages of using GAT1.
One is that GAT1 must be delivered to the rim of the presynapticspecialization to allow for neuronal retrieval of synaptically re-leased GABA. In fact, in mice engineered to express a fluorescentGAT1, presynaptic boutons of GABAergic interneurons in cere-bellum and hippocampus have 800 –1300 GAT1 molecules/�m 2
and the axons that connect boutons have a linear density of 640molecules/�m (Chiu et al., 2002). Another advantage is that traf-ficking of GAT1 from ER to ERGIC (Scholze et al., 2002; Farhanet al., 2004, 2007) and from the ERGIC (Farhan et al., 2008) isreasonably well understood.
Recently, Ye et al. (2007) concluded from a mutational anal-ysis of Drosophila neurons that dendritic growth required COPII-dependent ER export but that axonal growth was not disruptedby mutations of COPII components. In other words, the flow oflipids to support plasma membrane expansion is not dependenton COPII. However, it remains unclear whether protein traffick-ing to the axon requires the COPII machinery. Moreover theabsence of the Golgi from axons (Horton et al., 2005; Ye et al.,2007) and polarization of the Golgi toward the major dendrite(Horton et al., 2005) raises the question whether sorting andtargeting of proteins to the axon is dependent on the conven-tional secretory pathway. Here, we studied both, axonal deliveryof the membrane cargo GAT1 and the relevance of the conven-tional secretory pathway for its correct axonal targeting.
We found that the Sec24-binding deficient mutant GAT1-RL/AS reached the plasma membrane via an alternative noncon-ventional (i.e., COPII independent). This finding was importantbecause it allowed us to compare GAT1-RL/AS with the wild-type transporter. We further characterized this pathway andfound that it bypasses the ERGIC. The dependence of the wild-type GAT1 on COPII for ER export and its passage through theERGIC gave us the opportunity to test whether transit throughthe conventional secretory pathway ER3ERGIC3Golgi) deter-mines correct axonal delivery of membrane cargo. Two alterna-tive scenarios are conceivable: the first model posits that polar-ized sorting of membrane cargo is specified in the trans-Golginetwork regardless of how the protein reaches the Golgi. In thealternative scenario, sorting decisions are contingent on correcttransit through earlier steps, i.e., ER and/or ERGIC. Our data un-equivocally show that wild-type GAT1 is transported along a COPII-dependent pathway from ER to ERGIC and that it must transit theERGIC to be efficiently enriched in axon terminals. The failure ofGAT1-RL/AS to be enriched in axon terminals clearly indicatesthat (1) COPII plays a role in delivering GAT1 to the axon termi-nal and (2) that early stations of the secretory pathway predeter-mine correct axonal delivery of membrane cargo.
As another remarkable finding, we report that ARFGAP1 in-teracts with GAT1 and is required for GAT1 trafficking. Origi-nally, ARFGAP1 was identified by its eponymous action, i.e.,triggering the GTPase of ARF1 (Cukierman et al., 1995) and thusdriving the disassembly of the COPI coat by triggering the ARF1GTPase. However, more recently, it has been appreciated thatARFGAP1 may play a role in earlier steps of vesicle budding: theyeast ARFGAP ortholog Glo3p was shown to be recruited toCOPI-coated vesicles before ARF and to facilitate assembly of theCOPI coat (Rein et al., 2002). In fact, mammalian ARFGAP1apparently functions as a cargo adaptor that supports sorting ofproteins into nascent COPI vesicles (Lee et al., 2005). Our obser-vations are also consistent with a role of ARFGAP1 as a cargoadaptor: it mediates packaging of GAT1 into COPI vesicles thatmediate anterograde transport from the ERGIC to the Golgi.Interference with exit of GAT1 from the ERGIC precluded itsaccumulation in neurites. This finding underscores the notion
Figure 10. Interaction of ARFGAP1 with the C terminus of GAT1. A, Purified GST-tagged Cterminus (10 �g) of wild-type GAT1 (GST-GAT-Ct) or that of GAT1-RL/AS (GST-RLAS-Ct) or GSTwere incubated with His-tagged ARFGAP1 (3 �g) for 2 h on ice. Subsequently, GSH-agarose wasused to recover the proteins. B, Cytosol (100 �g) prepared from HEK293 cells overexpressingYFP-tagged �64ARFGAP1 was incubated with GST-tagged C terminus of wild-type GAT1 (GST-GAT-Ct) or of GAT1-RL/AS (GST-RL/AS-Ct) for 2 h on ice followed by a GST-pulldown. C, GST-tagged C terminus (10 �g) of GAT1 (GST-GAT-Ct) was incubated with His-tagged ARFGAP1 (4�g; His-ARFGAP1) for 2 h on ice (left lane) or His-ARFGAP1 was preincubated with 4 �g ofHis-tagged Arf1 in the presence of AlF3 for 30 min followed by 2 h incubation with GST-GAT1-Cton ice (middle lane). GST-pulldown was performed, and ARFGAP1 was detected using an anti-His antibody. The right lane represents 20% of ARFGAP1 input used for the reaction. D, His6-tagged ARFGAP1 was incubated with cytosol (100 �g) overexpressing HA-tagged Arf1 in thepresence of AlF3 for 30 min (left lane) or His6-tagged ARFGAP1 was preincubated with GST-tagged GAT1 C terminus for 2 h before the reaction. Thereafter, His6-tagged ARFGAP1 wasrecovered using magnetic Ni-NTA beads. The amount of Arf1 that was pulled down was deter-mined using anti-HA antibody (top blot). An antibody against the His6-tag was used to verifythat the amount of ARFGAP1 retrieved by the beads was comparable (bottom blot). E, GAT1 wasimmunoprecipitated form adult rat brain as described in Materials and Methods. The right lanecontains 5% input, the middle lane the eluate from the immunoprecipitation (IP) with ananti-GAT1 antibody (IP GAT1) and the left lane the mock immunoprecipitation with irrelevantIgG (IP IgG). The numbers on the right side indicate the position of the molecular weight marker.Data are representative of three independent experiments.
Reiterer et al. • Trafficking of GAT1 in the Early Secretory Pathway J. Neurosci., November 19, 2008 • 28(47):12453–12464 • 12461

that early trafficking steps play a crucialrole in determining proper targeting ofGAT1.
Our observations imply that GAT1must already recruit proteins required forsorting before reaching the Golgi. Earlierfindings support this conjecture. The lastthree amino acids of GAT1 represent atype II-PDZ domain binding motif(McHugh et al., 2004). This motif putsGAT1 surface expression under the con-trol of the exocyst which interacts with itsC terminus (Farhan et al., 2004) (our un-published observations). The exocyst cat-alyzes the very last stage in membrane pro-tein trafficking, i.e., the fusion of thesecretory vesicle with the cell surface. Nev-ertheless, if the interaction between GAT1and the exocyst is disrupted, GAT1 doesnot accumulate in a submembranouscompartment as would be expected, if onlythe insertion of secretory vesicles was im-peded: in the absence of a functional exo-cyst, or upon mutation of the PDZ-motif,GAT1 is rather retained in the ER (Farhanet al., 2004). Thus, the exocyst must be re-cruited at an early stage. In fact, there isevidence that components of the exocystsreside on the ER membrane (Shin et al.,2000; Lipschutz et al., 2003; Guo and Nov-ick, 2004). Finally, Muniz et al. (2001)showed that different subtypes of COPIIvesicles are generated from the ER, at leastin vitro, and that these vesicles contain dif-ferent types of cargo. This is a good prece-dent for sorting decisions made very earlyin the secretory pathway and importantlybefore the passage through the Golgi. It ishighly likely that a similar distinctive sort-ing also operates at the level of the ER inneurons. Our observations suggest thatmembrane carriers required for axonalsorting of GAT1 are COPII-dependent.This COPII dependence sorts GAT1 intoan anterograde pathway which assures itscorrect targeting. If GAT1 fails to enter thispathway, as in the case of GAT1-RL/AS, itdoes not become enriched in the axonalcompartment.
Trafficking of membrane proteinsthrough the secretory pathway is contin-gent on their sequential interaction withbinding partners which allow for theirrecruitment into transport vesicles andfor their correct delivery into the nextcompartment. Several proteins havebeen identified as binding partners ofGAT1, with the potential to direct theroute of GAT1 along the secretory path-way from the ER to its final destination,the presynaptic specialization. These in-clude syntaxin-1 (Quick et al., 1997),GAT1 itself (Schmid et al., 2001), Pals-1
Figure 11. Effect of ARFGAP1 on subcellular localization of GAT1. A, HeLa cells were transfected with a plasmid encodingYFP-tagged �64ARFGAP1. On the next day, colocalization with ERGIC-53 was determined using immunofluorescence. B, HEK293cells were cotransfected with plasmids encoding CFP-tagged GAT1 and YFP-tagged�64ARFGAP1, and images were acquired 24 hlater. C, HEK293 cells were transfected with a plasmid encoding CFP-tagged GAT1. On the next day, cells were treated with 50 �M
IC261. Cells were fixed with 4% paraformaldehyde, and ERGIC-53 was detected by immunofluorescence. All images were acquiredwith a confocal microscope. The extent of colocalization was quantified in three independent experiments as outlined underMaterials and Methods and in the legend to Figure 3 with 600/640 of ERGIC-53-containing vesicles (A) and 440/440 CFP-GAT1-containing vesicles (B) also containing YFP-tagged �64ARFGAP1 and 160/420 ERGIC-containing vesicles also containing YFP-tagged �64ARFGAP1.
Figure 12. Effect of ARFGAP1 on subcellular localization of GAT1 in neurons. Hippocampal neurons were prepared from ratembryos (E17). After 9 d in culture, neurons were transfected with plasmids encoding for YFP-tagged �64ARFGAP1. Cells werefixed after 48 h and immunostained for GAT1. Images were captured by confocal microscopy. A shows immunostaining for GAT1(left panel) in a neuron that underwent the transfection procedure but did not express YFP-tagged �64ARFGAP1 (right panel). Bshows staining for GAT1 (left panel) in a neuron expressing YFP-tagged �64ARFGAP1 (middle panel) and the resulting overlay(right panel). The extent of colocalization was quantified in three independent experiments as outlined under Materials andMethods and in the legend to Figure 3 with 111/136 of YFP-tagged �64ARFGAP1 vesicles also containing GAT1.
12462 • J. Neurosci., November 19, 2008 • 28(47):12453–12464 Reiterer et al. • Trafficking of GAT1 in the Early Secretory Pathway

(McHugh et al., 2004), and Sec24D (Farhan et al., 2004, 2007).While syntaxin-1 and Pals-1 may be important in the finalstage, i.e., the accumulation of GAT1 at the synapse, self-assembly into GAT1 oligomers (Farhan et al., 2006) and inter-action with Sec24D are required for the very first step ofCOPII-dependent export of GAT1 from the ER (Farhan et al.,2007). Here, we identified ARFGAP1 as an additional interac-tion partner of GAT1. Our experiments showed that ARF-GAP1 was required to support trafficking of GAT1 from theERGIC. There are many arguments derived from live cell im-aging in support of the notion that the ERGIC is a stationarycompartment (Ben-Tekaya et al., 2005). Moreover, a muta-tion can be introduced into the GAT1 C terminus that resultsin selective retention of the resulting GAT1-SSS in the ERGIC(Farhan et al., 2008). Thus, anterograde carriers must existthat transfer cargo, such as GAT1, from ERGIC to Golgi. Ourfinding that ARFGAP1 can act as a cargo adaptor for GAT1 istherefore consistent with a process of regulated sorting of pro-teins at the level of the ERGIC and a role of COPI in thisprocess (Shima et al., 1999).
In summary, the different mutants of GAT1 enabled us tocharacterize its trafficking in the secretory pathway. In additionthey allowed us to test the dependence of axonal sorting on earlysecretory events. Our results clearly show that COPII-mediatedER export is crucial for correct targeting of GAT1 to the axonterminal.
ReferencesAppenzeller-Herzog C, Hauri HP (2006) The ER-Golgi intermediate com-
partment (ERGIC): in search of its identity and function. J Cell Sci119:2173–2183.
Ben-Tekaya H, Miura K, Pepperkok R, Hauri HP (2005) Live imaging ofbidirectional traffic from the ERGIC. J Cell Sci 118:357–367.
Chiu CS, Jensen K, Sokolova I, Wang D, Li M, Deshpande P, Davidson N,Mody I, Quick MW, Quake SR, Lester HA (2002) Number, density, andsurface/cytoplasmic distribution of GABA transporters at presynapticstructures of knock-in mice carrying GABA transporter subtype 1-greenfluorescent protein fusions. J Neurosci 22:10251–10266.
Cukierman E, Huber I, Rotman M, Cassel D (1995) The ARF1 GTPase-activating protein: zinc finger motif and Golgi complex localization. Sci-ence 270:1999 –2002.
Farhan H, Korkhov VM, Paulitschke V, Dorostkar MM, Scholze P, KudlacekO, Freissmuth M, Sitte HH (2004) Two discontinuous segments in thecarboxyl terminus are required for membrane targeting of the rat gamma-aminobutyric acid transporter-1 (GAT1). J Biol Chem 279:28553–28563.
Farhan H, Freissmuth M, Sitte HH (2006) Oligomerization of neurotrans-mitter transporters: a ticket from the endoplasmic reticulum to theplasma membrane. Handb Exp Pharmacol 175:233–249.
Farhan H, Reiterer V, Korkhov VM, Schmid JA, Freissmuth M, Sitte HH(2007) Concentrative export from the endoplasmic reticulum of thegamma-aminobutyric acid transporter 1 requires binding to SEC24D.J Biol Chem 282:7679 –7689.
Farhan H, Reiterer V, Kriz A, Hauri H-P, Pavelka M, Sitte HH, Freissmuth M(2008) Signal-dependent export of the GABA transporter-1 from theintermediate compartment is specified by a carboxyl terminal motif. J CellSci 121:753–761.
Gomez M, Scales SJ, Kreis TE, Perez F (2000) Membrane recruitment ofcoatomer and binding to dilysine signals are separate events. J Biol Chem275:29162–29169.
Griffiths G, Pfeiffer S, Simons K, Matlin K (1985) Exit of newly synthesizedmembrane proteins from the trans cisterna of the Golgi complex to theplasma membrane. J Cell Biol 101:949 –964.
Guo Q, Vasile E, Krieger M (1994) Disruptions in Golgi structure and mem-brane traffic in a conditional lethal mammalian cell mutant are correctedby epsilon-COP. J Cell Biol 125:1213–1224.
Guo W, Novick P (2004) The exocyst meets the translocon: a regulatorycircuit for secretion and protein synthesis? Trends Cell Biol 14:61– 63.
Hauri HP, Kappeler F, Andersson H, Appenzeller C (2000) ERGIC-53 andtraffic in the secretory pathway. J Cell Sci 113:587–596.
Horton AC, Racz B, Monson EE, Lin AL, Weinberg RJ, Ehlers MD (2005)Polarized secretory trafficking directs cargo for asymmetric dendritegrowth and morphogenesis. Neuron 48:757–771.
Huber I, Cukierman E, Rotman M, Aoe T, Hsu VW, Cassel D (1998) Re-quirement for both the amino-terminal catalytic domain and a noncata-lytic domain for in vivo activity of ADP-ribosylation factor GTPase-activating protein. J Biol Chem 273:24786 –24791.
Krijnse-Locker J, Parton RG, Fuller SD, Griffiths G, Dotti CG (1995) Theorganization of the endoplasmic reticulum and the intermediate com-partment in cultured rat hippocampal neurons. Mol Biol Cell6:1315–1332.
Lee MC, Miller EA, Goldberg J, Orci L, Schekman R (2004) Bi-directionalprotein transport between the ER and Golgi. Annu Rev Cell Dev Biol20:87–123.
Lee SY, Yang JS, Hong W, Premont RT, Hsu VW (2005) ARFGAP1 plays acentral role in coupling COPI cargo sorting with vesicle formation. J CellBiol 168:281–290.
Li Y, Hou LX, Aktiv A, Dahlstrom A (2007) Studies of the central nervoussystem-derived CAD cell line, a suitable model for intraneuronal trans-port studies? J Neurosci Res 85:2601–2609.
Lipschutz JH, Lingappa VR, Mostov KE (2003) The exocyst affects proteinsynthesis by acting on the translocation machinery of the endoplasmicreticulum. J Biol Chem 278:20954 –20960.
Manzano J, Cuadrado M, Morte B, Bernal J (2007) Influence of thyroidhormone and thyroid hormone receptors in the generation of cerebellar�-aminobutyric acid-ergic interneurons from precursor cells. Endocri-nology 148:5746 –5751.
McHugh EM, Zhu W, Milgram S, Mager S (2004) The GABA transporterGAT1 and the MAGUK protein Pals1: interaction, uptake modulation,and coexpression in the brain. Mol Cell Neurosci 26:406 – 417.
Muniz M, Morsomme P, Riezman H (2001) Protein sorting upon exit fromthe endoplasmic reticulum. Cell 104:313–320.
Nickel W, Malsam J, Gorgas K, Ravazzola M, Jenne N, Helms JB, WielandFT (1998) Uptake by COPI-coated vesicles of both anterograde andretrograde cargo is inhibited by GTP�S in vitro. J Cell Sci111:3081–3090.
Pepperkok R, Scheel J, Horstmann H, Hauri HP, Griffiths G, Kreis TE (1993)Beta-COP is essential for biosynthetic membrane transport from the en-doplasmic reticulum to the Golgi complex in vivo. Cell 74:71– 82.
Pepperkok R, Whitney JA, Gomez M, Kreis TE (2000) COPI vesicles accu-mulating in the presence of a GTP restricted arf1 mutant are depleted ofanterograde and retrograde cargo. J Cell Sci 113:134 –144.
Pierce JP, van Leyen K, McCarthy JB (2000) Translocation machinery forsynthesis of integral membrane and secretory proteins in dendritic spines.Nat Neurosci 3:311–313.
Qi Y, Wang JK, McMillian M, Chikaraishi DM (1997) Characterization of aCNS cell line, CAD, in which morphological differentiation is initiated byserum deprivation. J Neurosci 17:1217–1225.
Quick MW, Corey JL, Davidson N, Lester HA (1997) Second messengers,trafficking-related proteins, and amino acid residues that contribute tothe functional regulation of the rat brain GABA transporter GAT1. J Neu-rosci 17:2967–2979.
Rein U, Andag U, Duden R, Schmitt HD, Spang A (2002) ARF-GAP-mediated interaction between the ER-Golgi v-SNAREs and the COPIcoat. J Cell Biol 157:395– 404.
Sannerud R, Marie M, Nizak C, Dale HA, Pernet-Gallay K, Perez F, Goud B,Saraste J (2006) Rab1 defines a novel pathway connecting the pre-Golgiintermediate compartment with the cell periphery. Mol Biol Cell17:1514 –1526.
Sans N, Wang PY, Du Q, Petralia RS, Wang YX, Nakka S, Blumer JB, MacaraIG, Wenthold RJ (2005) mPins modulates PSD-95 and SAP102 traffick-ing and influences NMDA receptor surface expression. Nat Cell Biol7:1179 –1190.
Saraste J, Kuismanen E (1984) Pre- and post-Golgi vacuoles operate in thetransport of Semliki forest virus membrane glycoproteins to the cell sur-face. Cell 38:535–549.
Scheffzek K, Ahmadian MR, Kabsch W, Wiesmuller L, Lautwein A, SchmitzF, Wittinghofer A (1997) The Ras-RasGAP complex: structural basis forGTPase activation and its loss in oncogenic Ras mutants. Science277:333–338.
Schmid JA, Scholze P, Kudlacek O, Freissmuth M, Singer EA, Sitte HH(2001) Oligomerization of the human serotonin transporter and of the
Reiterer et al. • Trafficking of GAT1 in the Early Secretory Pathway J. Neurosci., November 19, 2008 • 28(47):12453–12464 • 12463

rat GABA transporter 1 visualized by fluorescence resonance energytransfer microscopy in living cells. J Biol Chem 276:3805–3810.
Scholze P, Freissmuth M, Sitte HH (2002) Mutations within an intramem-brane leucine heptad repeat disrupt oligomer formation of the rat GABAtransporter 1. J Biol Chem 277:43682– 43690.
Schweizer A, Fransen JA, Matter K, Kreis TE, Ginsel L, Hauri HP (1990) Iden-tification of an intermediate compartment involved in protein transportfrom endoplasmic reticulum to Golgi apparatus. Eur J Cell Biol 53:185–196.
Shima DT, Scales SJ, Kreis TE, Pepperkok R (1999) Segregation of COPI-rich and anterograde-cargo-rich domains in endoplasmic-reticulum-to-Golgi transport complexes. Curr Biol 9:821– 824.
Shin DM, Zhao XS, Zeng W, Mozhayeva M, Muallem S (2000) The mam-
malian Sec6/8 complex interacts with Ca(2�) signaling complexes andregulates their activity. J Cell Biol 150:1101–1112.
Wendeler MW, Paccaud JP, Hauri HP (2007) Role of Sec24 isoforms inselective export of membrane proteins from the endoplasmic reticulum.EMBO Rep 8:258 –264.
Xu D, Hay JC (2004) Reconstitution of COPII vesicle fusion to generate apre-Golgi intermediate compartment. J Cell Biol 167:997–1003.
Ye B, Zhang Y, Song W, Younger SH, Jan LY, Jan YN (2007) Growing den-drites and axons differ in their reliance on the secretory pathway. Cell130:717–729.
Yu S, Roth MG (2002) Casein kinase I regulates membrane binding by ARFGAP1. Mol Biol Cell 13:2559 –2570.
12464 • J. Neurosci., November 19, 2008 • 28(47):12453–12464 Reiterer et al. • Trafficking of GAT1 in the Early Secretory Pathway
























![B¨okstedtperiodicityandquotientsofDVRs · Lurie, P. Scholze and B.Bhatt. It appeared in work of Bhatt-Morrow-Scholze [BMS19] as well as in [AMN18]. But the maneuver of working relative](https://img.pdfslide.us/doc/110x75/605aa78f8ed29f5c5d69c014/bokstedtperiodicityandquotientsofdvrs-lurie-p-scholze-and-bbhatt-it-appeared.jpg)




