Embed Size (px)
Citation preview

Developmentally distinct in vivo effects of FSH on proliferation andapoptosis during testis maturation
Sarah J Meachem1, Saleela M Ruwanpura1, Jessica Ziolkowski2,Jacquelyn M Ague3, Michael K Skinner3 and Kate L Loveland2,4
1Prince Henry’s Institute of Medical Research, Monash Medical Centre, 246 Clayton Road, Clayton, Victoria, 3168, Australia2Monash Institute for Medical Research, Monash University, Clayton, Victoria, 3168, Australia3Center for Reproductive Biology, School of Molecular Biosciences, Washington State University, Pullman, Washington, 99164, USA4The Australian Research Council Centre of Excellence in Biotechnology and Development
(Requests for offprints should be addressed to S J Meachem; Email: [email protected])
Abstract
The critical influence of follicle stimulating hormone(FSH) on male fertility relates both to its impact on Sertolicell proliferation in perinatal life and to its influence on thesynthesis of Sertoli cell-derived products essential for germcell survival and function in the developing adult testis.The nature and timing of this shift of germ cells to theirreliance on specific Sertoli cell-derived products are notdefined. Based on existing data, it is apparent that thedominant function of FSH shifts between 9 and 18 daypostpartum (dpp) during the first wave of spermatogenesisfrom driving Sertoli cell proliferation to support germ cells.To enable comprehensive analysis of the impact of acutein vivo FSH suppression on Sertoli and germ cell develop-ment, FSH was selectively suppressed in Sprague–Dawleyrats by passive immunisation for 2 days and/or 4 days priorto testis collection at 3, 9 and 18 dpp. The 3 dpp samplesdisplayed no measurable changes, while 4 days of FSHsuppression decreased Sertoli cell proliferation andnumbers in 9 dpp, but not 18 dpp, animals. In contrast,
germ cell numbers were unaffected at 9 dpp but decreasedat 18 dpp following FSH suppression, with a correspond-ing increase in germ cell apoptosis measured at 18 dpp.Sixty transcripts were measured as changed at 18 dpp inresponse to 4 days of FSH suppression, as assessed usingAffymetrix microarrays. Some of these are known asSertoli cell-derived FSH-responsive genes (e.g. StAR,cathepsin L, insulin-like growth factor binding protein-3),while others encode proteins involved in cell cycle andsurvival regulation (e.g. cyclin D1, scavenger receptor classB 1). These data demonstrate that FSH differentiallyaffects Sertoli and germ cells in an age-dependent mannerin vivo, promoting Sertoli cell mitosis at day 9, andsupporting germ cell viability at day 18. This model hasenabled identification of candidate genes that contribute tothe FSH-mediated pathway by which Sertoli cells supportgerm cells.Journal of Endocrinology (2005) 186, 429–446
Introduction
Sperm production in the adult male requires establishmentof the Sertoli cell population and initiation of the firstspermatogenic wave, events that occur during foetal andearly postnatal life. In the rat, Sertoli cells proliferate untilaround 15 days after birth, setting the full complement ofSertoli cells. Surrounded by Sertoli cells, the gonocytesmultiply in the foetal testis and then enter a period ofquiescence lasting a week. They resume mitosis at 3 daysafter birth and migrate to the basement membrane at theseminiferous cord perimeter. Once they have contactedthe basement membrane they are called spermatogonia.Spermatogonia subsequently pass through three develop-mental phases involving mitosis, meiosis and spermiogen-esis to form spermatozoa, with the germ cells completing
the first wave of spermatogenesis at 43 days after birth inthe rat (de Rooij 1998).
The mechanisms by which the Sertoli and germ cellpopulations are regulated during the first spermatogenicwave are under investigation and appear to be influencedby a complex network of interacting signals. It is knownthat pituitary-derived follicle stimulating hormone (FSH)exerts its effects on the developing germ cells indirectly, asonly the Sertoli cells bear FSH receptors. A key role ofFSH in setting the size of the Sertoli cell population inearly postnatal life has been attributed to its stimulatoryeffect on Sertoli cell division (Orth 1984, Orth et al. 1988,Boitani et al. 1995). The response of the Sertoli cell toFSH changes during the first wave of spermatogenesisin rodents, and this influences the proliferation ofSertoli cells (Boitani et al. 1995, Meehan et al. 2000,
429
Journal of Endocrinology (2005) 186, 429–4460022–0795/05/0186–429 � 2005 Society for Endocrinology Printed in Great Britain
DOI: 10.1677/joe.1.06121Online version via http://www.endocrinology-journals.org
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access

Buzzard et al. 2003). Synthesis of several Sertoli cellproducts also changes during development, howeverwhether some of these are regulated by FSH in vivoremains to be determined (Munsie et al. 1997, Yan et al.2001, Fragale et al. 2001, Migrenne et al. 2003, Pellegriniet al. 2003). FSH-regulated apoptosis during the firstspermatogenic wave has not been examined, however inadult rats, germ cells are lost through apoptosis (Meachemet al. 1999), rather than through reduced proliferationwhen FSH is manipulated (McLachlan et al. 1995).However the regulation of adult spermatogenesis cannotbe assumed to be mediated in the same manner as is thefirst spermatogenic wave, as shifts in the expressionpatterns of key regulatory genes (i.e., Bcl-2 family mem-bers and stem cell factor (SCF) have been observed asSertoli cells mature and eventually mature germ cellpopulations emerge (Munsie et al. 1997, Huang et al.1992, Meehan et al. 2001).
A large number of FSH responsive genes have beenidentified, however in most cases, their specific contribu-tion to Sertoli cell and germ cell viability and function isundefined. The interaction of SCF with its receptor, c-kit,is essential for germ cell progression and survival in firstwave and adult spermatogenesis (Yoshinaga et al. 1991,Packer et al. 1995), and expression of SCF in Sertoli cellshas been shown to be regulated by FSH in vitro and in vivo(Rossi et al. 1993, McLean et al. 2002). Expression ofthe transcription factor DMRT1 in Sertoli cells can beelevated by FSH in vivo (Chen & Heckert 2001) and a rolefor DMRT1 in early postnatal testicular developmentrelating to the termination of Sertoli cell proliferation hasbeen deduced from the phenotype of DMRT -/- mice(Raymond et al. 2000). Some genes encoding apoptoticregulators in the Bcl-2 family of proteins have also beenshown to be responsive to FSH, including Bcl-w and Bok(Yan et al. 2000, Suominen et al. 2001). A recent studyusing microarray analyses has defined 100–300 knowntranscripts that are regulated by FSH in cultures of Sertolicells from 20 days post partum (dpp) rats (McLean et al.2002). It is now timely to identify the FSH regulated genesinvolved in testicular development and spermatogenesisusing an in vivo approach.
To more precisely understand the dynamic mechanismsby which FSH acts in immature rats, we set out to identifythe cells that respond to in vivo changes in FSH levelsduring the first wave of spermatogenesis and to examinethe functional changes in these cells. Administration of anantibody to FSH was performed to selectively suppressFSH for two or four days by immunoneutralisation. Thisacute FSH suppression differentially affected Sertoli andgerm cell numbers during development as assessed by the,unbiased, optical disector stereological technique, corre-sponding with selective changes in proliferation andapoptosis in these cell types. Rat genomic microarraysenabled identification of candidate genes regulated byFSH from 14 to 18 dpp. The value of this model for
interrogating the changing roles of FSH in the first wave ofspermatogenesis is discussed.
Material and Methods
Animals
Male outbred Sprague–Dawley rat pups with motherswere obtained from the Monash University CentralAnimal House (Clayton, Australia). They were maintainedat 20 �C in a fixed 12 h light:12 h darkness cycle with freeaccess to food and water in accordance with the AustralianCode of Practice for Care and Use of Animals for ScientificPurposes (1997, National Health and Medical ResearchCouncil, Australia). This study was approved by theMonash Medical Centre Animal Ethics Committee.
Passive immunisation against FSH
The ability of the polyclonal ovine antisera raised againstrat FSH (FSHAb) to neutralise rat FSH in vitro and in vivohas been previously described (Meachem et al. 1998). Thelevel of neutralisation of serum FSH achieved in adultrats was greater than 90% (reaching the limit of assaydetection). Rat pups were immunised with the FSHAb orwith normal sheep immunoglobulin (ConAb) at a dailydose of 10 mg/kg, a 5-fold higher dose than that pre-viously administered to adult rats. Each animal receiveds.c. injections 2 days prior to death at 3 days dpp, and 2 and4 days prior to death at 9 and 18 dpp. Ten rats wereinjected for each data point.
Tissue collection and preparation
One h prior to death, each rat received 5-bromodeoxyurdine(BrdU; 50 mg/kg, s.c. Sigma) to enable proliferationanalysis. Rats were killed 24 h after the final injection ofthe antibody, at 3, 9 or 18 dpp, by decapitation. Trunkblood was collected and allowed to clot overnight at 4 �Cprior to serum collection for hormone assays. Due to thesmall amounts of serum collected for day 3 and 9 dpp rats,these samples were pooled in groups of three and twoanimals, respectively. The testes were then excised andweighed. The right testis of each animal was snap frozenon dry ice and stored at �75 �C for RNA preparation,while left testis was immersion-fixed with Bouin’s solutionfor less than 5 h and sliced into 2-mm thick slabs ortho-gonal to the long axis of the testis. The testes from 9 and18 dpp rats were divided into two, or four slices, respect-ively, and half of each processed into hydroxyethylmeth-acrylate resin (Technovit 7100; Kulzer and Co. GmBH,Friedrichsdorf, Germany) according to the manufacturer’sinstructions, while the other half was used for routineembedding into paraffin. The whole left testis from 3 dpprats was embedded in resin and paraffin in every second
S J MEACHEM and others · FSH effects on the immature testis430
www.endocrinology-journals.orgJournal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access

animal. Thick (25 µm) resin sections were serially cut(Supercut Microtome, Reichert Jung 2050, Nossloch,Germany), stained with the Periodic acid-Schiff’s reactionreagents and counterstained with Mayer’s Haematoxylinfor the determination of cell number. Thin (5µm) paraffinsections were placed on Superfrost Plus slides for analysis ofproliferation and apoptosis.
Cell number estimates using the optical disector
The optical disector stereological method (Wreford 1995)was used to determine the total number of cell nuclei pertestis. All measurements were performed using a 100�objective on an Olympus BX-50 microscope (Olympus,Tokyo, Japan). A microcator (D 8225; Heideinhain,Traunreut, Germany) that monitored scanned depth wasattached to the microscope stage. The images werecaptured by a JVC TK-C1381 video camera coupled to aPentium PC computer using a Screen Machine II fastmultimedia video adapter (FAST, Hamburg, Germany).The software package, CASTGRID V1·60 (Olympus,Denmark, Germany), was used to generate an unbiasedcounting frame superimposed on video image. Fields wereselected by a systematic uniform random samplingscheme as previously described (McLachlan et al. 1994,Wreford 1995) with the use of a motorized stage (Multi-control 2000; ITK, Lahnau, Germany). The final screenmagnification was 2708-fold.
Sertoli cells were identified by their irregularly shapednuclei, which were often positioned towards to thebasement membrane and contained multiple nucleoli.Gonocytes were relatively large circular to ovoid cellswith large circular nuclei, centrally located within theepithelium. Type A and B spermatogonia/preleptotenespermatocytes, leptotene/zygotene spermatocytes andpachytene spermatocytes were identified according to thecharacteristics described (Russell et al. 1990). Type Bspermatogonia and preleptotene spermatocytes were notreadily distinguishable at these stages of development andwere counted as one. At least 300 Sertoli cell nuclei in totalper testis were counted using the unbiased counting frame(175–430 µm2) and at least 80 gonocytes, spermatogonia(type A, B spermatogonia and preletotene spermatocytes)or spermatocytes (leptotene, zygotene and pachytene sper-matocytes) were counted employing a larger frame (430–1923 µm2). The frame size was selected based on cellfrequency at different time points; less abundant cell typeswere counted in a larger counting frame. No correction forshrinkage was required (Meachem et al. 1996). Slides weremasked prior to each type of quantitation (cell number,proliferation and apoptosis) to facilitate unbiased counting.
Proliferation analysis
BrdU incorporation into testicular cells at 3, 9 and 18 dppwas detected by immunohistochemistry as previously
described (Schlatt et al. 1999), with minor modifications asdescribed below (see Fig 1A). In brief, slides bearingparaffin-embedded tissue sections were deparaffinized,rehydrated, and subjected to antigen retrieval in citratebuffer (0·1 M, pH 6; 90–95 �C for 10 min then roomtemperature for 20 min). Sections were treated succes-sively with trypsin (0·25%, 1·5 min) and 3% hydrogenperoxide in methanol (20 min) and washed in PBS. Allsubsequent procedures were performed at room tempera-ture, washes were in PBS, and incubations were in ahumid chamber. Sections were treated with first withSuperblock (40 min; DAKO, Carpenteria, CA, USA) andthen with the monoclonal antibody to BrdU (clone BU-33, 4µl/ml in PBS, 1 h; Sigma). Following washes,biotinylated rabbit anti-mouse IgG was added (1:300 inPBS, 30 min; Zymed, California, USA). Sections werewashed and incubated first in ABC complex (40 min;Vectastain Elite, Vector Laboratories, Burlingame, CA)and then with Tyramide Signal Amplification Biotinreagent (30 min; PerkinElmer Life Sciences, Inc, Boston,MA, USA) according to the manufacturers’ instructions.After washing, diaminobenzidine (DAB) was added toreveal sites of antibody binding with a dark brown reactionproduct (2–3 min; DAKO Liquid DAB Substrate Chro-mogen System), and sections were counterstained withMayer’s Haematoxylin (3 min; Sigma), blued in Scott’s tapwater (1 min), and finally dehydrated and mounted inDepex (BDH Laboratory Suppliers, Poole, Dorset, UK)under glass coverslips. Cell types with anti-BrDU nuclearstaining were identified on the basis of their locationwithin the cord and the size and shape of cell nuclei, asdescribed above.
The percentage of BrdU labelled cells was assessed usingan unbiased counting frame of 430 µm2. To determine theproliferation index for each cell type, the total number ofBrdU-labelled cells was divided by the total number oflabelled and unlabelled cells.
Apoptosis analysis
Tissue sections (5 µm) were deparaffinized and rehydratedprior to the detection of DNA fragmentation (see Fig 1B).Apoptotic cells were detected using the terminal deoxy-nucleotidyl transferase (TdT)-mediated dUTP-biotinnick-end labelling (TUNEL) method (Meachem et al.1999). Apoptotic cells were visualised using the chro-mogen DAB and processed as described for BrDU detec-tion. On control sections, the TdT enzyme was omitted.TUNEL-positive cell types were identified based on theirlocation within the cord, their size, and the shape of thecell nucleus.
The percentage of cells with TUNEL labelling wasassessed using an unbiased counting frame of 430 µm2 andthe apoptotic index calculated by dividing the number ofTUNEL labelled cells by the total number of labelled andunlabelled cells.
FSH effects on the immature testis · S J MEACHEM and others 431
www.endocrinology-journals.org Journal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access
Serum androgen levels
Serum androgen levels were measured by RIA usingiodinated histamine-testosterone in combination with anacidic buffer (pH 5·1) to disrupt binding between testos-terone and binding proteins in unextracted serum samples(O’Donnell et al. 1994). Serum samples (5–25µl) wereassayed in duplicate across two assays. Within assay vari-ation for both assays was 3%, and between assay variationwas 9%. Assay sensitivity was 0·6 ng/ml.
Serum inhibin levels
Immunoreactive inhibin was measured by heterologousRIA as described previously (Robertson et al. 1988).Results are expressed in terms of an in-house rat ovarianextract calibrated against human recombinant (hr)-inhibin. Iodinated hr-inhibin was used as tracer. Theantiserum used was rabbit antiserum (#1989) which isdirected towards the �-subunit thereby measuring bothinhibin A and B, and cross-reacts 288% with pro-⋅C, theprosequence of the inhibin alpha subunit (Robertson et al.1989). Goat anti-rabbit IgG (GAR#12; Monash Instituteof Reproduction & Development, Monash University,Melbourne, Australia) was used as second antibody. Theassay buffer used was 0·01 M PBS containing 0·5% BSA(Sigma). Mouse serum pools diluted in a dose-dependentmanner and were parallel to the standard curve (data notshown). The samples were run in a single assay in 20µlduplicates. The within-assay variation was 7·2%. Assaysensitivity was 0·17 ng/ml.
Oligonucleotide microarray hybridisation
To identify genes regulated by FSH at the 18 day timepoint, total RNA was isolated from total testes using acidphenol extraction (Chomczynski & Sacchi 1987). RNAfrom two individual animals treated from 14 dpp-18 dppwith either FSHAb or ConAb (4 samples in total) wasused. The gene expression profile of FSHAb- andConAb-treated testis were individually determined usingAffymetrix RG_U34A rat chips (Affymetrix Inc., SantaClara, CA, USA) as previously described (McLeanet al. 2002). The RNA from the FSHAb- and the
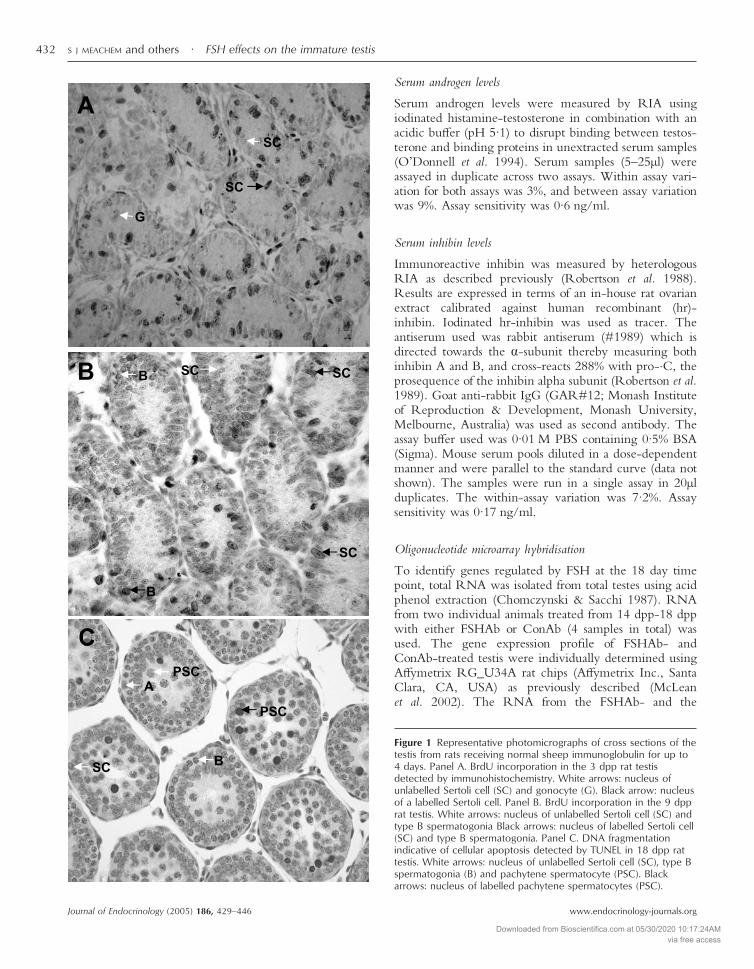
Figure 1 Representative photomicrographs of cross sections of thetestis from rats receiving normal sheep immunoglobulin for up to4 days. Panel A. BrdU incorporation in the 3 dpp rat testisdetected by immunohistochemistry. White arrows: nucleus ofunlabelled Sertoli cell (SC) and gonocyte (G). Black arrow: nucleusof a labelled Sertoli cell. Panel B. BrdU incorporation in the 9 dpprat testis. White arrows: nucleus of unlabelled Sertoli cell (SC) andtype B spermatogonia Black arrows: nucleus of labelled Sertoli cell(SC) and type B spermatogonia. Panel C. DNA fragmentationindicative of cellular apoptosis detected by TUNEL in 18 dpp rattestis. White arrows: nucleus of unlabelled Sertoli cell (SC), type Bspermatogonia (B) and pachytene spermatocyte (PSC). Blackarrows: nucleus of labelled pachytene spermatocytes (PSC).
S J MEACHEM and others · FSH effects on the immature testis432
www.endocrinology-journals.orgJournal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access

ConAb-treated testis was extracted and purified beforebiotin labelling and hybridisation to the microarray genechip. Two replicates of each sample were hybridised toseparate chips giving a total of four chips in the analysis.Each gene set on the Affymetrix chip is composed of 16pairs of 24-mer oligonucleotides, with 8799 genes on eachRG_U34A chip. Included in each set is one anti-sensestrand specific for the gene and one anti-sense strandwith single point mutations is used as a comparativenegative control. The labelled RNA was visualized on aHewlett-Packard Gene Array Scanner (Hewlett-PackardCo., Palo Alto, CA, USA). An initial two way comparisonwas performed using the Affymetrix Suite software. Inboth experiments the FSHAb- and ConAb-treated chipswere compared against each other.
Bioinformatics and microarray statistics
The bioinformatics and statistical analysis performed is aspreviously described (Eisen et al. 1998, Chaudhary et al.2005, Kezele et al. 2002). Microarray output was exam-ined visually for excessive background noise and physicalanomalies. The default Microarray Suite (MAS, SiliconGenetics, Redwood City, CA, USA) statistical valueswere used for all analyses. An absolute analysis using MASwas performed to assess the relative abundance of thetranscripts based on signal and detection (present, absent,or marginal) for the 16 different oligonucleotides per geneand comparison for analysis. The absolute analysis fromMAS was imported into GeneSpring 5·1 software (SiliconGenetics, Redwood City, CA, USA). The data werenormalized within GeneSpring using the default/recommended normalisation methods. These include set-ting of signal values below 0·01 to 0·01, total chipnormalisation to the 50th percentile, and normalisation ofeach gene to the median. These normalisations allowed forthe comparison of data based on relative abundance in anysample set rather than compared with a specific controlvalue. Transcripts expressed differentially at a statisticallysignificant level were determined using a one-wayANOVA parametric test with variances not assumedequal, and a P-value cutoff of 0·05. This was applied to allsamples and considered all transcripts represented on thearrays. Two independent samples for each treatment groupwere analyzed was performed and allowed a 2�2 factorialcomparison in the experiment. Subsequently, expressionrestrictions were applied to identify the transcriptsexpressed in a significant manner. These restrictions weredesigned so that the remaining transcripts met the follow-ing requirements in addition to being expressed in asignificant manner: 1) each transcript have a signal value ofat least 100 in the average of both samples, from at leastone of the treatments tested and 2) had an average foldchange of 1·5 or greater in signal intensity betweentreatments. Transcripts that passed these restrictions wereconsidered for further analysis. Previous studies have
shown that microarray data correlates well with real-timequantitative PCR and Northern analysis (Eisen et al.1998, Chaudhary et al. 2005, Kezele et al. 2002, Sadate-Ngatchou et al. 2004) . Therefore, microarray data doesnot need to be confirmed as previously suggested (Shimaet al. 2004). However, two selected genes were used in areal-time quantitative PCR procedure as previouslydescribed (McChlery & Clarke 2003) to help confirm themicroarray procedure. The microarray chip data can beaccessed at www.skinner.wsu.edu.
Reverse transcription (RT) and real-time PCR analysis
Real time PCR was used to measure the relative levels oftwo candidate FSH-regulated genes. Total RNA collectedfrom two individual day 18 rats treated with either ConAbor with FSH Ab for 4 days was treated to remove residualgenomic DNA (Ambion DNA-free Treatment Kit;Ambion, Austin TX, USA). RNA (500 ng) was convertedto cDNA in a final volume of 20 µl using Superscript IIaccording to the manufacturer’s protocol (Invitrogen). Foreach sample, the absence of contaminating genomic DNAin cDNA samples was confirmed using reactions in whichthe RT enzyme was omitted. Quantitative RT-PCRanalysis was performed using the Roche LightCycler(Roche, Mannheim, Germany) and the FastStart DNAMaster SYBR-green 1 system (Roche). Oligonucleotideprimer sequences for IGFBP-3, Smad3, and beta-actinwere obtained either from published sources or Frodo.wi.mit.edu/cgi-bin/primer3/primer3_www.cgi;version 3·0(Table 1). For PCR analysis, sample cDNA was diluted1:10- to 1:50-fold, and PCR reaction conditions, includ-ing Mg2+ concentration, primer concentration, annealtime and extension time were optimized for each primerpair as summarized in Table 1. For all PCR analyses,standard curves were generated using dilutions of animmature rat testicular cDNA preparation of arbitraryunits (i.e. 17 dpp). PCR of all standards and samples wereperformed using duplicate reactions for approximately40–45 cycles, after which a melting curve analysis wasperformed to monitor PCR product purity (see Table 1).In initial experiments, PCR product identities were veri-fied by agarose gel electrophoresis and DNA sequencing(data not shown).
Statistics
A two-sample t-test was used to determine differencesbetween FSHAb and ConAb treated samples with theassumption that data were normally distributed for allhistological and hormone data. If data did not show normaldistribution, then a Mann–Whitney test was carriedout using Sigmastat for Windows version 2·0 (JandelCorporation, CA, USA). Data are expressed as mean�S.E.M., n=7 rats per group. Statistical analysis of micro-array data is described in Bioinformatics and microarraystatistics.
FSH effects on the immature testis · S J MEACHEM and others 433
www.endocrinology-journals.org Journal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access

Results
Testicular weights and serum hormone measurements
Testicular weights were not significantly different follow-ing 2 days of FSH suppression in 3 dpp rats compared withtheir corresponding controls (Table 2), however testicularweights were reduced in 9 and 18 dpp animals after 2 (to84% of control) and 4 days (to 78% of control) of FSHsuppression. FSH suppression did not affect serum andro-gen levels at any time point compared with controls(Table 2). Serum inhibin levels following 2 days of FSHsuppression in 3, 9 and 18 dpp rats were significantlyreduced to 74%, 71% and 69%, and after 4 days 73% and53% in 9 and 18 dpp rats compared with control values,respectively (Table 2). This reduction in circulatinginhibin levels is presumed to reflect the successful neu-tralisation of FSH bioactivity through administration of theantibody raised against FSH, as FSH normally stimulatesproduction of inhibin from Sertoli cells.
Cell populations decreased in FSH withdrawn rats
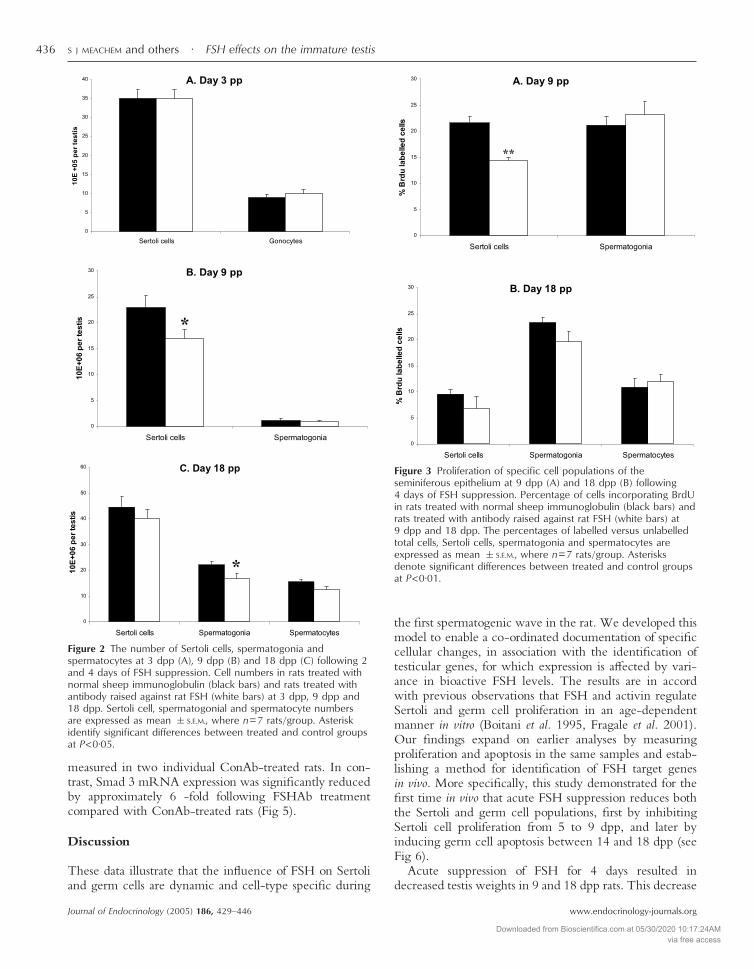
At 3 dpp there were no differences in gonocyte (0·098�0·010 vs 0·091�0·054 million/testis) or Sertoli cell num-bers (3·50�2·4 vs 3·50�2·3 million/testis) following2 days of FSHAb treatment compared with ConAb treat-ment (Fig 2A), correlating no change in testis weight. At9 dpp, there was a significant reduction (to 63% of control;P<0·05) in the number of Sertoli cells following 4 days ofFSH suppression (16·8�1·7 vs 23·0�1·7 million/testis),while spermatogonial numbers were unchanged (0·9�0·08 vs 1·1�0·4 million/testis) (Fig 2B). At 18 dpp nodifference in the number of Sertoli cells were observed inresponse to 4 days of FSHAb treatment (40·2�3·20 vs44·2�4·18 million/testis). A reduction in spermatogoniato 75% (16·9�1·8 vs 22·2�1·0 million/testis, P<0·05) ofcontrol group values was observed following 4 days of FSHsuppression. Spermatocyte number tended to be reducedto 80% (12·4�1·1 vs 15·5�1·0 million/testis, P=0·061)of control group values was observed following 4 days ofFSH suppression, although this did not achieve significance(Fig 2C).
FSH suppression decreases Sertoli cell proliferation inday 9 rats
At 3 dpp, no differences in Sertoli cell proliferation wereobserved between FSHAb- and ConAb-treated rats(23·4%�3·2 vs 27·2%�1·8); no BrdU labelled gonocyteswere observed in these samples (Refer Fig 1). At9 dpp, there was a significant reduction (to 66% of control,P<0·01) in the proliferation rate of Sertoli cells followingFSH suppression (14·3%�0·6 vs 21·6%�1·2) (Fig 2A),while no change was observed for spermatogonia(23·2%�2·4 vs 21·1%�1·6, P=0·11) (Fig 3A). AtTa
ble
1Pr
imer
-spe
cific
Ligh
tCyc
ler
cond
ition
sus
edfo
rPC
Ram
plifi
catio
nof
rat
IGF-
BP3,
Smad
3an
dbe
ta-a
ctin
Prim
erse
quen
cePr
oduc
tsi
ze(b
p)
Prim
erco
ncen
trat
ion
(pm
ol)
Mg2
+
conc
entr
atio
n(m
M)
Ann
ealin
gte
mpe
ratu
re(�
C)
Exte
nsio
nti
me
(Sec
)
Dat
ate
mpe
ratu
re(�
C)*
PCR
prod
uct
Tm (�C
)**
Nam
e[A
cces
sion
No.
]Ra
tIG
F-BP
3[N
M_0
1258
8]se
nse
5�-A
ATG
CTG
GG
AG
TGTG
GA
AA
G-3
�an
tisen
se5�
-GC
GG
TATC
TAC
TGG
CTC
TGC
-3�
145
303·
558
1890
90
Rat
Smad
3[U
6647
9]se
nse
5�-A
AG
GG
CG
AG
CA
GA
AC
GG
G-3
�an
tisen
se5�
-GG
GA
TGG
AA
TGG
CTG
TAG
TC-3
�42
535
3·0
5719
9292
Rat
�-ac
tin[N
M_0
3114
4]se
nse
5�-C
CG
TAA
AG
AC
CTC
TATG
CC
AA
CA
-3�
antis
ense
5�-G
CTA
GG
AG
CC
AG
GG
CA
GTA
ATC
-3�
103
502·
567
580
86
*Tem
pera
ture
atw
hich
the
fluor
esce
nce
ofth
ePC
Rpr
oduc
tw
asqu
antifi
eddu
ring
Ligh
tCyc
ler
anal
ysis
;**P
CR
prod
ucts
have
char
acte
ristic
mel
ting
poin
ts(T
m).
The
Tmfo
ra
DN
Apr
oduc
tis
defin
edas
the
tem
pera
ture
atw
hich
half
ofth
eD
NA
helic
alst
ruct
ure
islo
st,a
ndis
dete
rmin
edby
mel
ting
tem
pera
ture
anal
ysis
onth
eLi
ghtC
ycle
r.
S J MEACHEM and others · FSH effects on the immature testis434
www.endocrinology-journals.orgJournal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access

18 dpp, in response to FSH suppression, there was nochange in Sertoli cell (6·8%�2·3 vs 9·5%�1·0, P=0·30),spermatogonial (19·6%�1·9 vs 23·3%�1·0, P=0·11) orspermatocyte proliferation (11·9%�1·4 vs 10·8%�1·7,P=0·64) (Fig 3B) compared with their correspondingcontrols. These data indicate that the reduction in testisweight due to FSH suppression at 9 dpp, but not at 18dpp, reflects the lower number of Sertoli cells in thesesamples. The incidence of apoptosis was next investigatedas an alternative basis for the reduced testis weights in the18 dpp samples (Fig 3B).
Germ cell apoptosis is affected by FSH suppression in18 dpp rats
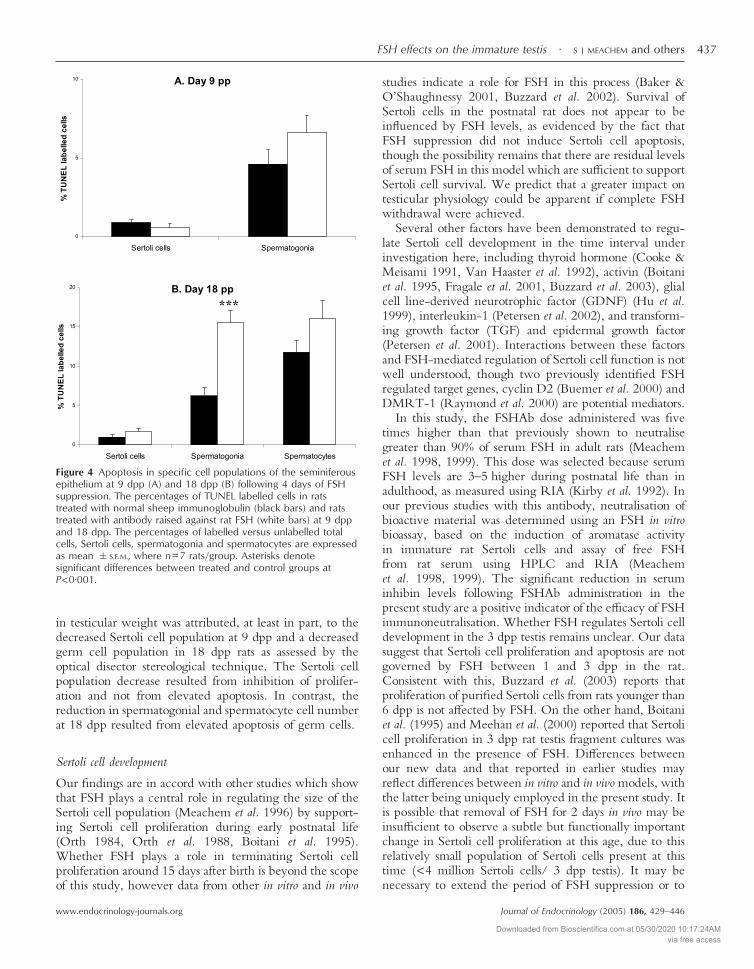
No apoptosis was observed in Sertoli cells and gonocytes at3 dpp (Refer Fig 1). No differences in Sertoli cell(0·6%�0·2 vs 0·9%�0·2, P=0·36) or spermatogonialapoptosis (6·6%�1·1 vs 4·6%�0·9, P=0·20) wereobserved following 4 days of FSH suppression at 9 dppcompared with controls (Fig 4A). However, at 18 dpp,there was a significant elevation of spermatogonial apop-tosis (250% of control) in response to FSH suppression(15·5%�1·6 vs 6·2%�1·0, P<0·001); spermatocyteapoptosis increased to 136% of control levels (16·1%�2·2vs 11·8%�1·4, P=0·13), although this did not achievesignificance (Fig 4B). No change in Sertoli cell apoptosiswas observed at any time point (Fig 4). These data indicatethat a shift occurs between 9 and 18 dpp within the germcell population towards reliance for support cues onFSH-derived products. The nature of this support system
and the effects of its removal were examined using agenomic microarray to identify genes regulated by alteredlevels of FSH at 18 dpp.
Microarray analysis
Thirty genes were identified in the 18 dpp samples asup-regulated and 30 as down-regulated (Table 3) follow-ing FSH suppression by treatment with FSHAb for 4 days.These 60 genes of interest met the selection criteriaoutlined previously; all give at least one signal intensitygreater than 100, are flagged as changing, and have foldchanges of 1·5 or greater.
Amongst these genes, 12 out of 60 (20%) have pre-viously been reported to be regulated by FSH, predomi-nantly through in vitro studies. Many of the genes on thislist have not yet been identified as being expressed in thetestis (26/60), and some of those known to be in the testishave not previously been assessed for regulation by FSH(14/34). This latter category of candidate in vivo FSHtarget genes have been reported to be expressed in Sertolicells (4/14), germ cells (8/14) and/or non-Sertoli somaticcells (6/14); clearly the non-Sertoli cells expressedgenes that would be regulated indirectly, through FSHregulation of Sertoli cell biosynthesis or other functions.
RT-PCR analysis
IGFBP-3 mRNA expression levels (measured in twoindividual animals) in FSHAb-treated rats were signifi-cantly elevated approximately 2·5-fold above the levels
Table 2 The testicular weights (mg), serum testosterone (ng/ml) and inhibin (ng/ml) levelsat 3 dpp, 9 dpp and 18 dpp following 2 and 4 days of FSH suppression. Rats were treatedwith normal sheep immunoglobulin (ConAb) and with antibody raised against rat FSH(FSHAb) at 3 dpp, 9 dpp and 18 dpp following 2 and 4 days of treatment. The data areexpressed as mean�S.E.M., with n=10 rats/group.
Testicular weight(mg)
Serum testosterone(ng/ml)
Serum inhibin(ng/ml)
Treatment3 days post partum
+ConAb 2d 5·1�0·2 0·64�0·08 3·57�0·21+FSHAb 2d 5·1�0·1 0·63�0·11 2·65�0·06**
9 days post partum+ConAb 2d 17·9�0·9 1·36�0·3 4·53�0·3+FSHAb 2d 15·2�0·5* 1·18�0·1 3·23�0·2*+ConAb 4d 18·7�0·4 0·89�0·2 4·17�0·24+FSHAb 4d 14·6�0·5** 0·70�0·1 3·03�0·19*
18 days post partum+ConAb 2d 75·4�3·0 0·92�0·3 3·51�0·4+FSHAb 2d 63·2�3·4* 0·92�0·2 2·43�0·2*+ConAb 4d 74·1�1·2 1·80�0·5 5·75�0·31+FSHAb 4d 57·9�2·1** 1·36�0·2 3·09�0·20**
Asterisks denote significant differences between rats receiving FSHAb and their corresponding controls.*P<0·01 and **P<0·001.
FSH effects on the immature testis · S J MEACHEM and others 435
www.endocrinology-journals.org Journal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access
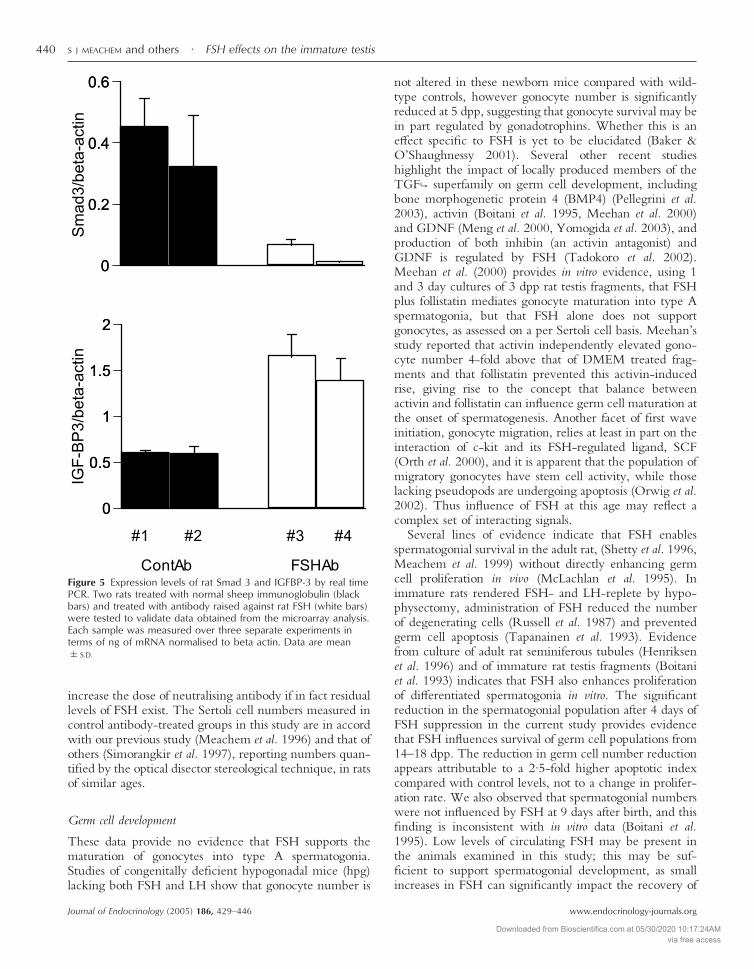
measured in two individual ConAb-treated rats. In con-trast, Smad 3 mRNA expression was significantly reducedby approximately 6 -fold following FSHAb treatmentcompared with ConAb-treated rats (Fig 5).
Discussion
These data illustrate that the influence of FSH on Sertoliand germ cells are dynamic and cell-type specific during
the first spermatogenic wave in the rat. We developed thismodel to enable a co-ordinated documentation of specificcellular changes, in association with the identification oftesticular genes, for which expression is affected by vari-ance in bioactive FSH levels. The results are in accordwith previous observations that FSH and activin regulateSertoli and germ cell proliferation in an age-dependentmanner in vitro (Boitani et al. 1995, Fragale et al. 2001).Our findings expand on earlier analyses by measuringproliferation and apoptosis in the same samples and estab-lishing a method for identification of FSH target genesin vivo. More specifically, this study demonstrated for thefirst time in vivo that acute FSH suppression reduces boththe Sertoli and germ cell populations, first by inhibitingSertoli cell proliferation from 5 to 9 dpp, and later byinducing germ cell apoptosis between 14 and 18 dpp (seeFig 6).
Acute suppression of FSH for 4 days resulted indecreased testis weights in 9 and 18 dpp rats. This decrease
Figure 2 The number of Sertoli cells, spermatogonia andspermatocytes at 3 dpp (A), 9 dpp (B) and 18 dpp (C) following 2and 4 days of FSH suppression. Cell numbers in rats treated withnormal sheep immunoglobulin (black bars) and rats treated withantibody raised against rat FSH (white bars) at 3 dpp, 9 dpp and18 dpp. Sertoli cell, spermatogonial and spermatocyte numbersare expressed as mean �S.E.M., where n=7 rats/group. Asteriskidentify significant differences between treated and control groupsat P<0·05.
Figure 3 Proliferation of specific cell populations of theseminiferous epithelium at 9 dpp (A) and 18 dpp (B) following4 days of FSH suppression. Percentage of cells incorporating BrdUin rats treated with normal sheep immunoglobulin (black bars) andrats treated with antibody raised against rat FSH (white bars) at9 dpp and 18 dpp. The percentages of labelled versus unlabelledtotal cells, Sertoli cells, spermatogonia and spermatocytes areexpressed as mean �S.E.M., where n=7 rats/group. Asterisksdenote significant differences between treated and control groupsat P<0·01.
S J MEACHEM and others · FSH effects on the immature testis436
www.endocrinology-journals.orgJournal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access
in testicular weight was attributed, at least in part, to thedecreased Sertoli cell population at 9 dpp and a decreasedgerm cell population in 18 dpp rats as assessed by theoptical disector stereological technique. The Sertoli cellpopulation decrease resulted from inhibition of prolifer-ation and not from elevated apoptosis. In contrast, thereduction in spermatogonial and spermatocyte cell numberat 18 dpp resulted from elevated apoptosis of germ cells.
Sertoli cell development
Our findings are in accord with other studies which showthat FSH plays a central role in regulating the size of theSertoli cell population (Meachem et al. 1996) by support-ing Sertoli cell proliferation during early postnatal life(Orth 1984, Orth et al. 1988, Boitani et al. 1995).Whether FSH plays a role in terminating Sertoli cellproliferation around 15 days after birth is beyond the scopeof this study, however data from other in vitro and in vivo
studies indicate a role for FSH in this process (Baker &O’Shaughnessy 2001, Buzzard et al. 2002). Survival ofSertoli cells in the postnatal rat does not appear to beinfluenced by FSH levels, as evidenced by the fact thatFSH suppression did not induce Sertoli cell apoptosis,though the possibility remains that there are residual levelsof serum FSH in this model which are sufficient to supportSertoli cell survival. We predict that a greater impact ontesticular physiology could be apparent if complete FSHwithdrawal were achieved.
Several other factors have been demonstrated to regu-late Sertoli cell development in the time interval underinvestigation here, including thyroid hormone (Cooke &Meisami 1991, Van Haaster et al. 1992), activin (Boitaniet al. 1995, Fragale et al. 2001, Buzzard et al. 2003), glialcell line-derived neurotrophic factor (GDNF) (Hu et al.1999), interleukin-1 (Petersen et al. 2002), and transform-ing growth factor (TGF) and epidermal growth factor(Petersen et al. 2001). Interactions between these factorsand FSH-mediated regulation of Sertoli cell function is notwell understood, though two previously identified FSHregulated target genes, cyclin D2 (Buemer et al. 2000) andDMRT-1 (Raymond et al. 2000) are potential mediators.
In this study, the FSHAb dose administered was fivetimes higher than that previously shown to neutralisegreater than 90% of serum FSH in adult rats (Meachemet al. 1998, 1999). This dose was selected because serumFSH levels are 3–5 higher during postnatal life than inadulthood, as measured using RIA (Kirby et al. 1992). Inour previous studies with this antibody, neutralisation ofbioactive material was determined using an FSH in vitrobioassay, based on the induction of aromatase activityin immature rat Sertoli cells and assay of free FSHfrom rat serum using HPLC and RIA (Meachemet al. 1998, 1999). The significant reduction in seruminhibin levels following FSHAb administration in thepresent study are a positive indicator of the efficacy of FSHimmunoneutralisation. Whether FSH regulates Sertoli celldevelopment in the 3 dpp testis remains unclear. Our datasuggest that Sertoli cell proliferation and apoptosis are notgoverned by FSH between 1 and 3 dpp in the rat.Consistent with this, Buzzard et al. (2003) reports thatproliferation of purified Sertoli cells from rats younger than6 dpp is not affected by FSH. On the other hand, Boitaniet al. (1995) and Meehan et al. (2000) reported that Sertolicell proliferation in 3 dpp rat testis fragment cultures wasenhanced in the presence of FSH. Differences betweenour new data and that reported in earlier studies mayreflect differences between in vitro and in vivo models, withthe latter being uniquely employed in the present study. Itis possible that removal of FSH for 2 days in vivo may beinsufficient to observe a subtle but functionally importantchange in Sertoli cell proliferation at this age, due to thisrelatively small population of Sertoli cells present at thistime (<4 million Sertoli cells/ 3 dpp testis). It may benecessary to extend the period of FSH suppression or to
Figure 4 Apoptosis in specific cell populations of the seminiferousepithelium at 9 dpp (A) and 18 dpp (B) following 4 days of FSHsuppression. The percentages of TUNEL labelled cells in ratstreated with normal sheep immunoglobulin (black bars) and ratstreated with antibody raised against rat FSH (white bars) at 9 dppand 18 dpp. The percentages of labelled versus unlabelled totalcells, Sertoli cells, spermatogonia and spermatocytes are expressedas mean �S.E.M., where n=7 rats/group. Asterisks denotesignificant differences between treated and control groups atP<0·001.
FSH effects on the immature testis · S J MEACHEM and others 437
www.endocrinology-journals.org Journal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access

Tabl
e3
Gen
esup
and
dow
nre
gula
ted
byFS
Hin
day
18po
stpa
rtum
rat
test
es.D
ay15
rats
wer
etr
eate
dw
ithno
rmal
shee
pim
mun
oglo
bulin
orw
ithan
antib
ody
rais
edag
ains
tra
tFS
Hfo
r4
days
.Res
ults
are
base
don
two
mic
roar
ray
hybr
idiz
atio
nsof
targ
etsy
nthe
size
dfro
mRN
Afro
mtw
oin
depe
nden
tra
tte
stes
from
each
trea
tmen
tgr
oup.
The
Affy
met
rixm
icro
arra
yda
tase
tw
asan
alyz
edby
pairw
ise
com
paris
onus
ing
Gen
eSi
fter.
Sam
ple
sets
with
asi
gnal
at�
100
dete
cted
inat
leas
ton
epo
sitio
non
the
arra
yan
da
fold
chan
geof
1·5
orgr
eate
rar
ere
port
ed.R
efer
ence
sfo
rth
ose
gene
spr
evio
usly
repo
rted
tobe
inth
era
tte
stis
and/
orre
gula
ted
byFS
Har
ein
pare
nthe
ses.
Acc
essi
onnu
mbe
rFo
ldch
ange
Ave
rage
sign
alFS
HA
b+
/-S.
E.M
.Te
stis
expr
essi
onpr
evio
usly
repo
rted
FSH
regu
lati
onpr
evio
usly
repo
rted
Gen
esup
-reg
ulat
edby
FSH
supp
ress
ion
Insu
lin-li
kegr
owth
fact
orbi
ndin
gpr
otei
n3
AI0
0940
5;M
3183
76·
47;3
·12
17·4
55·
85;1
·99
Yes;
Sert
olic
ells
(Sm
ithet
al.1
990,
Rapp
apor
t&
Smith
1995
,Kh
anet
al.2
002)
Yes
Test
inU
1685
83·
9730
·65
0·90
Yes;
Sert
olic
ells
(Grim
aet
al.1
998,
Luie
tal
.200
3)Ye
s
HN
F-3/
fork
head
hom
olog
-1L1
3201
3·14
36·6
50·
39Ye
s;(K
aest
ner
etal
.199
8,M
igre
nne
etal
.200
3,W
olfru
met
al.2
003)
No
2·4
kbre
peat
DN
Arig
htte
rmin
alre
gion
X054
722·
6253
·01·
99N
oN
oEn
doth
elin
1M
6471
12·
4636
·15
0·46
Yes;
Sert
olic
ells
(Fan
toni
etal
.199
3,Tr
ipic
iano
etal
.199
9)Ye
s
cDN
Acl
one
rx01
264
3�A
I639
097
2·44
121·
550·
25N
oN
oA
TP-b
indi
ngca
sset
te,s
ub-fa
mily
Csu
lfony
lure
are
cept
or2B
mRN
A(S
UR2
B);
AF0
1962
82·
3320
·75
1·01
Yes;
Leyd
igce
lls(S
chin
kel&
Jonk
er20
03,O
’Sha
ughn
essy
etal
.20
02)
No
Cyc
linD
1A
I231
257
2·04
8·50
1·60
Yes;
gono
cyte
s&
sper
mat
ogon
ia(W
olge
mut
het
al.1
998,
Buem
eret
al.2
000)
No
cDN
A,3
�en
d/c
lone
=RK
IBA
73A
A89
2986
1·98
6·05
0·90
No
No
Lipa
seA
,lys
osom
alac
idS8
1497
1·98
;1·8
455
·95
0·63
;0·6
3Ye
s;G
uine
api
g(K
abba
jet
al.2
001)
No
SH3
dom
ain
bind
ing
prot
ein
CR1
6U
3115
91·
9310
·80·
63N
oN
oG
-pro
tein
-cou
pled
rece
ptor
105
U76
206
1·81
8·85
0·56
No
No
Fibr
inog
en,b
eta
poly
pept
ide
M35
602
1·80
7·85
0·52
No
No
Cyc
licPr
otei
n-2=
cath
epsi
nL
proe
nzym
eS8
5184
1·74
;1·6
758
·15
0·12
;0·1
2Ye
s;Se
rtol
icel
ls(P
entti
laet
al.1
995,
Wrig
htet
al.2
003)
Yes
Mas
tce
llpr
otea
se7
U67
910
1·74
20·6
0·85
No
No
UD
Pgl
ycos
yltr
ansf
eras
e1
fam
ily,p
olyp
eptid
eA
7A
F039
212
1·71
38·5
0·18
No
No
N-m
ycdo
wns
trea
m-r
egul
ated
gene
2A
A79
9560
1·70
185·
550·
10N
oN
oRa
tre
tinol
-bin
ding
prot
ein
(RBP
)M
1093
41·
6938
·50
0·37
Yes;
Sert
olic
ells
(Esk
ildet
al.2
000)
Yes
Cry
stal
lin,a
lpha
BM
5553
41·
6743
·90
0·84
Yes;
sper
mat
ocyt
es(K
ato
etal
.199
1)N
o
Sim
ilar
tom
icro
som
algl
utat
hion
eS-
tran
sfer
ase
3(L
OC
2891
97)
AA
8922
341·
6635
8·95
0·25
No
No
Ribo
nucl
ease
7pr
ecur
sor;
eosi
noph
ilca
tioni
cpr
otei
nD
8858
61·
6330
·70·
27N
oN
oM
itoge
nac
tivat
edpr
otei
nki
nase
kina
seki
nase
1U
4859
61·
5832
1·3
0·55
No
No
Cad
herin
2;N
-Cad
herin
AF0
9759
31·
5765
·15
0·30
Yes;
Sert
olic
ells
(Lam
paet
al.1
999)
Yes
Rat
nucl
ear
rece
ptor
co-r
egul
ator
1M
8176
61·
5675
·85
0·09
Yes;
Sert
olic
ells
(Duf
our
&Ki
m19
99,K
astn
eret
al.1
996)
No
Reel
inA
A89
3471
1·55
39·2
50·
44Ye
s;(Ik
eda
&Te
rash
ima
1999
,And
erse
net
al.
2003
)N
o
Vdu
p1;u
preg
ulat
edby
1,25
-dih
ydro
xyvi
tam
inD
-3A
I014
169
1·55
490·
70·
20N
oN
oPC
4;in
terfe
ron-
rela
ted
deve
lopm
enta
lreg
ulat
or1
AI0
1416
31·
5478
·05
0·37
Yes;
germ
cells
(Mbi
kay
etal
.199
7)N
o
N-a
cety
lglu
cosa
min
yltr
ansf
eras
eI
D16
302
1·51
132·
450·
09Ye
s;(N
ozak
iet
al.2
003)
No
mRN
Afo
rM
HC
clas
sII
antig
enRT
1.B-
1be
ta-c
hain
M36
151
1·51
50·9
02·
25Ye
s;ge
rmce
lls(M
orie
tal
.200
0)N
o
Rat
tus
norv
egic
ustr
ansc
ribed
sequ
ence
sA
A89
1962
1·51
11·6
0·16
No
No
S J MEACHEM and others · FSH effects on the immature testis438
www.endocrinology-journals.orgJournal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access

Tabl
e3
Con
tinue
d
Acc
essi
onnu
mbe
rFo
ldch
ange
Ave
rage
sign
alFS
HA
b+
/-S.
E.M
.Te
stis
expr
essi
onpr
evio
usly
repo
rted
FSH
regu
lati
onpr
evio
usly
repo
rted
Gen
esup
-reg
ulat
edby
FSH
supp
ress
ion
Ster
oido
geni
cac
ute
regu
lato
rypr
otei
nA
B001
349;
AB0
0600
74·
34;4
·21
59·8
5;81
·05
6·81
;1·3
6Ye
s;Se
rtol
icel
ls(G
rego
ry&
De
Phill
ip19
98,M
cLea
net
al.2
002,
Man
naet
al.2
003)
Yes
Am
inol
evul
inic
acid
synt
hase
2D
8629
73·
0018
8·20
0·64
No
No
Phos
phod
iest
eras
e4B
AA
7997
292·
55;1
·80
14·4
50·
51;0
·42
Yes;
Sert
olic
ells
(Sw
inne
net
al.1
991)
Yes
MA
Dho
mol
og3
(Dro
soph
ila)
U66
479
2·39
40·6
50·
12Ye
s;Se
rtol
i,Le
ydig
cells
&sp
erm
atoc
ytes
(Xu
etal
.200
3)N
o
Mel
anom
ain
hibi
tory
activ
ity;c
artil
age
deriv
edre
tinoi
cac
idse
nsiti
vepr
otei
nU
6788
42·
2735
7·45
1·12
No
No
Synd
ecan
1S6
1865
2·24
82·1
00·
48Ye
s;Se
rtol
icel
ls(B
ruca
toet
al.2
002)
Not
test
ed
75kD
agl
ucos
ere
gula
ted
prot
ein
S785
562·
0853
4·40
0·23
No
No
Gap
junc
tion
mem
bran
ech
anne
lpro
tein
alph
a1
(gja
-1);
conn
exin
43A
I029
183
2·06
50·1
00·
38Ye
s;Se
rtol
i,Le
ydig
&ge
rmce
lls(S
omm
ersb
erg
etal
.200
0,Br
avo-
Mor
eno
etal
.200
2)Ye
s
Neu
rona
lact
ivity
-reg
ulat
edpe
ntra
xin
S826
492·
0525
3·4
1·51
No
No
Prot
ein
Cin
hibi
tor
AB0
1312
82·
0367
6·65
0·28
Yes;
Leyd
ig&
germ
cells
(Uhr
inet
al.2
000,
Ode
tet
al.2
003)
No
Org
anic
anio
ntr
ansp
orte
rpr
otei
n3
(OA
T3);
solu
teca
rrie
rfa
mily
21(fa
ttyac
idtr
ansp
orte
r),m
embe
r7
AF0
4110
51·
9227
·30
0·45
Yes
No
Am
yloi
dbe
ta(A
4)pr
ecur
sor-
like
prot
ein
2X7
7934
1·87
431·
550·
20Ye
s;(v
onde
rKa
mm
eret
al.1
994)
No
R.n
orve
gicu
scD
NA
clon
erx
0502
43�
AI6
3911
51·
8252
·90
0·44
No
No
R.n
orve
gicu
ssi
mila
rto
Esop
hagu
sca
ncer
-rel
ated
gene
-2pr
otei
npr
ecur
sor
(EC
RG-2
)(L
OC
3010
16),
mRN
AA
A79
9734
1·82
336·
750·
23N
oN
o
Add
ucin
3,ga
mm
aU
3577
51·
7912
7·8
0·00
No
No
Mon
ocar
boxy
late
tran
spor
ter
(MC
T1);
solu
teC
arrie
rfa
mily
16,m
embe
r1
D63
834
1·77
45·8
0·20
Yes;
germ
cells
(Jack
son
etal
.199
7,Bo
usso
uar
etal
.200
3,G
odda
rdet
al.2
003)
No
Scav
enge
rre
cept
orcl
ass
B,m
embe
r1
AA
8748
431·
7734
5·00
0·35
Yes;
Sert
oli&
Leyd
igce
lls(R
eave
net
al.2
000,
Kaw
asak
iet
al.2
002)
No
Lysy
loxi
dase
AI2
3406
01·
7644
·30·
16Ye
s;(A
sunc
ion
etal
.200
1,M
akie
tal
.200
1)N
oTr
ansc
ribed
sequ
ence
with
mod
erat
esi
mila
rity
topr
otei
nre
f:N
P_06
1921
·1(H
omo
sapi
ens)
hypo
thet
ical
prot
ein
FLJ2
0752
(H.s
apie
ns)
AA
7995
311·
6717
4·65
0·42
Yes;
(Eric
cson
etal
.199
3)N
o
Prot
eolip
idpr
otei
nA
I070
277
1·67
169·
750·
20Ye
s;(L
iet
al.2
002)
No
IP63
prot
ein
X993
301·
6680
·35
0·08
No
No
Fum
arat
ehy
drat
ase
1A
I171
734
1·63
61·4
50·
18N
o,bu
tit
isub
iqui
tous
No
cAM
Pre
spon
sive
elem
ent
mod
ulat
orU
0483
51·
6336
·85
0·74
Yes;
(Wal
ker
etal
.199
4,W
est
etal
.199
4,M
onac
oet
al.1
995)
Yes
Lam
inB1
U72
353
1·63
41·5
50·
59Ye
s;Se
rtol
i,Le
ydig
,per
itubu
lar
&ge
rmce
lls(M
achi
els
etal
.199
7)N
o
Cyc
linD
2A
A89
9106
1·60
279·
550·
45Ye
s;Se
rtol
icel
ls&
sper
mat
ogon
ia(W
olge
mut
het
al.1
998,
Buem
eret
al.2
000,
Stro
thm
ann
etal
.200
4)
Yes
And
roge
nBi
ndin
gPr
otei
n(A
BP);
sex
horm
one
bind
ing
glob
ulin
M31
179
1·59
873·
950·
11Ye
s;Se
rtol
icel
ls(S
kinn
eret
al.1
989,
Dan
zo19
95,L
arrib
aet
al.
1995
,Jos
eph
etal
.199
7)
Yes
Supe
roxi
dedi
smut
ase
3X6
8041
1·57
147·
150·
21N
oN
oC
hond
roiti
nsu
lfate
prot
eogl
ycan
AA
8188
941·
5617
·85
0·34
Yes;
Leyd
igce
lls(U
ngef
rore
net
al.1
995)
No
Nic
otin
amid
enu
cleo
tide
tran
shyd
roge
nase
AA
8918
721·
5340
5·45
0·30
Yes;
(Ark
blad
etal
.200
2)N
oTr
ansc
ribed
sequ
ence
with
mod
erat
esi
mila
rity
topr
otei
nsp
:P0
0722
(E.c
oli)
BGA
L_EC
OLI
Beta
-gal
acto
sida
se(L
acta
se)
AA
8925
221·
5260
0·8
0·39
No
No
FSH effects on the immature testis · S J MEACHEM and others 439
www.endocrinology-journals.org Journal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access
increase the dose of neutralising antibody if in fact residuallevels of FSH exist. The Sertoli cell numbers measured incontrol antibody-treated groups in this study are in accordwith our previous study (Meachem et al. 1996) and that ofothers (Simorangkir et al. 1997), reporting numbers quan-tified by the optical disector stereological technique, in ratsof similar ages.
Germ cell development
These data provide no evidence that FSH supports thematuration of gonocytes into type A spermatogonia.Studies of congenitally deficient hypogonadal mice (hpg)lacking both FSH and LH show that gonocyte number is
not altered in these newborn mice compared with wild-type controls, however gonocyte number is significantlyreduced at 5 dpp, suggesting that gonocyte survival may bein part regulated by gonadotrophins. Whether this is aneffect specific to FSH is yet to be elucidated (Baker &O’Shaughnessy 2001). Several other recent studieshighlight the impact of locally produced members of theTGF® superfamily on germ cell development, includingbone morphogenetic protein 4 (BMP4) (Pellegrini et al.2003), activin (Boitani et al. 1995, Meehan et al. 2000)and GDNF (Meng et al. 2000, Yomogida et al. 2003), andproduction of both inhibin (an activin antagonist) andGDNF is regulated by FSH (Tadokoro et al. 2002).Meehan et al. (2000) provides in vitro evidence, using 1and 3 day cultures of 3 dpp rat testis fragments, that FSHplus follistatin mediates gonocyte maturation into type Aspermatogonia, but that FSH alone does not supportgonocytes, as assessed on a per Sertoli cell basis. Meehan’sstudy reported that activin independently elevated gono-cyte number 4-fold above that of DMEM treated frag-ments and that follistatin prevented this activin-inducedrise, giving rise to the concept that balance betweenactivin and follistatin can influence germ cell maturation atthe onset of spermatogenesis. Another facet of first waveinitiation, gonocyte migration, relies at least in part on theinteraction of c-kit and its FSH-regulated ligand, SCF(Orth et al. 2000), and it is apparent that the population ofmigratory gonocytes have stem cell activity, while thoselacking pseudopods are undergoing apoptosis (Orwig et al.2002). Thus influence of FSH at this age may reflect acomplex set of interacting signals.
Several lines of evidence indicate that FSH enablesspermatogonial survival in the adult rat, (Shetty et al. 1996,Meachem et al. 1999) without directly enhancing germcell proliferation in vivo (McLachlan et al. 1995). Inimmature rats rendered FSH- and LH-replete by hypo-physectomy, administration of FSH reduced the numberof degenerating cells (Russell et al. 1987) and preventedgerm cell apoptosis (Tapanainen et al. 1993). Evidencefrom culture of adult rat seminiferous tubules (Henriksenet al. 1996) and of immature rat testis fragments (Boitaniet al. 1993) indicates that FSH also enhances proliferationof differentiated spermatogonia in vitro. The significantreduction in the spermatogonial population after 4 days ofFSH suppression in the current study provides evidencethat FSH influences survival of germ cell populations from14–18 dpp. The reduction in germ cell number reductionappears attributable to a 2·5-fold higher apoptotic indexcompared with control levels, not to a change in prolifer-ation rate. We also observed that spermatogonial numberswere not influenced by FSH at 9 days after birth, and thisfinding is inconsistent with in vitro data (Boitani et al.1995). Low levels of circulating FSH may be present inthe animals examined in this study; this may be suf-ficient to support spermatogonial development, as smallincreases in FSH can significantly impact the recovery of
Figure 5 Expression levels of rat Smad 3 and IGFBP-3 by real timePCR. Two rats treated with normal sheep immunoglobulin (blackbars) and treated with antibody raised against rat FSH (white bars)were tested to validate data obtained from the microarray analysis.Each sample was measured over three separate experiments interms of ng of mRNA normalised to beta actin. Data are mean�S.D.
S J MEACHEM and others · FSH effects on the immature testis440
www.endocrinology-journals.orgJournal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access

spermatogonial number in adult rats (Meachem et al.1998). In the present study, spermatogonial number wasassessed using a sensitive, unbiased optical disector methodappropriate for counting these cells. Effects of loweredFSH levels may have been masked due to factors associatedwith counting a small population of cells. Spermatogonialand spermatocyte numbers in control antibody-treated ratsin this study were in accord with our (Meachem et al.1996) and that of others (Simorangkir et al. 1997) reportingdata that were assessed in rats of similar ages by the usingthe optical disector technique.
Age dependent changes in cell survival resulting fromFSH suppression may be due to developmentally regulatedchanges in the cellular synthesis of apoptotic regulators,such that documented for members of the Bcl-2 familyduring the first wave of mouse spermatogenesis (Meehanet al. 2001). The Bcl-w (Yan et al. 2000) and Bok(Suominen et al. 2001) mRNAs were shown to be up- anddown-regulated respectively by FSH in vitro, and the bcl-wknockout mouse has elevated germ cell apoptosis following
14 dpp, indicating the potential influence of Bcl-2 familymembers on the cellular responses in this model.
Identification of genes regulated by FSH in vivo
Interrogation of a rat gene microarray with RNA from18 dpp rats in this study identified 60 genes that wereregulated by 4 days of FSH suppression in vivo. Twentypercent of these differentially expressed genes have beenpreviously identified as FSH-regulated, predominantlythrough in vitro methods, a finding which indicates thevalidity of this model and experimental approach. Forexample, in an analysis of 20 dpp rat Sertoli cells using thesame microarray platform, FSH-responsive mRNAs wereidentified at 2, 4, 8 and 24 h of culture (McLean et al.2002). In accord with the present analysis, that in vitroanalysis identified the genes encoding steroidogenic acuteregulatory (StAR) protein and endothelin as down- andup- regulated, respectively, in the absence of FSH.FSH has now been identified through both in vitro
Figure 6 Dynamic effects of FSH on Sertoli and germ cell development during first wave in rats assessed through in vivo and in vitroanalyses. Time points listed illustrate progressive maturation of germ cells and Sertoli cells in the first wave of spermatogenesis in rat. At3 dpp, Sertoli cells are proliferating and gonocytes are re-entering the cell cycle and migrating to the basement membrane of theseminiferous cord to become undifferentiated and stem cell spermatogonia. At 9 dpp, proliferating Sertoli cells and differentiatingspermatogonia are observed. By 18 dpp, Sertoli cell proliferation has ceased, and the cords have transformed into seminiferous tubules astight junctions form between Sertoli cells and vectorial secretion from this epithelium commences. Germ cells that have progressed to thepachytene stage of meiosis are predominant in the seminiferous tubules. In this study, suppression of FSH by passive immunisation for2 and 4 days revealed the age-dependent differential in vivo affects of FSH on Sertoli and germ cells: promotion of Sertoli cell mitosis at9 dpp and increasing germ cell viability at 18 dpp. These data are in agreement with previous in vitro observations that FSH regulatesSertoli cell proliferation in an age-dependent manner, interacting with activin to promote Sertoli proliferation and germ cell developmentat discrete ages. This comparison also highlights differences that may specifically reflect the use of in vitro versus in vivo models.
FSH effects on the immature testis · S J MEACHEM and others 441
www.endocrinology-journals.org Journal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access

(McLean et al. 2002) and in vivo studies to be one of thewide variety of transcriptional regulators of StAR mRNAsynthesis (Gregory & De Phillip 1998, Manna et al. 2003).Cyclin D2 and androgen binding protein (ABP) areadditional Sertoli cell proteins up-regulated by FSH thatare required for normal testicular function (Skinner et al.1989, Danzo 1995, Larriba et al. 1995, Sicinski et al. 1996,Joseph et al. 1997), while protein C inhibitor, also requiredfor spermatogenesis (Uhrin et al. 2000, Odet et al. 2003),appears to be synthesized only in germ cells and Leydigcells (Odet et al. 2003). In vivo confirmation of the negativeregulation by FSH of genes encoding insulin-like growthfactor binding protein-3 (IGFBP-3) (Smith et al. 1990,Rappaport & Smith 1995, Khan et al. 2002), HNF-3/forkhead (Wolfrum et al. 2003), transferring (Kaestneret al. 1998), endothelin-1 (Fantoni et al. 1993, Tripicianoet al. 1999) and cathepsin-L (Penttila et al. 1995, Wrightet al. 2003) is presented. Members of the HNF-3/forkhead transcription factor family are part of a signaltransduction cascade from Akt/protein kinase B thatregulates transcription of apoptosis-related genes(Wolfrum et al. 2003). HNF-gamma is known to regulatetransferrin (Kaestner et al. 1998), a gene that is itselfregulated by FSH (Migrenne et al. 2003); we hypothesizethat HNF-3 represents an intermediate for FSH regulationof several genes.
The high concordance of the present data with previousreports provides validation of the methodological approachused to identify in vivo targets of FSH regulation, thoughsome differences may be noted. In this study, FSHsuppression led to elevated testicular N-cadherin mRNA,but in a previous 2 day in vitro analysis, FSH alone did notaffect N-cadherin expression; FSH in combination withtestosterone stimulated it (Lampa et al. 1999). This mostlikely reflects a specific difference between in vitro andin vivo models regarding hormone responses; alternatively,the duration of suppression or addition may underpin thedifference between these two results.
In addition, novel candidates for FSH regulation havebeen uncovered, some of which may be indirectly regu-lated by altered levels of available FSH or represent bonafide novel FSH-regulated genes. Targets not previouslyidentified in the testis include aminolevulinic acid synthase2 and melanoma inhibitory activity.
Several categories of encoded gene products within thearray of candidate FSH target genes may aid identificationof the pathways and processes that underpin the reductionin germ cell survival that was measured after 4 days oftreatment with the FSHAb. Most striking are the genesrelating to cell cycle regulation and apoptosis (e.g. lamin,cyclin D1, cyclin D2, Scavenger Receptor B1). Productsof other FSH target genes comprise or influence thearchitecture of the seminiferous epithelium (e.g. cadherin,cathepsin L), while others are known to function in signaltransduction (e.g. HNF3, MAPKKK, Smad 3, rat retinol-binding protein) and to participate in regulation of hor-
monal inputs (StAR and androgen binding protein).However, because the microarray analysis presented in thisstudy addresses a single time point, early and acutelyaffected FSH gene targets may be undetected. The cellularresponses identified in this study (i.e. germ cell apoptosis)would have been initiated before the time point of analysis,so future studies will address this in order to more fullycomprehend the pathways through which FSH levelsinfluence Sertoli cell function and germ cell survival.
The limit for microarray signal changes considered to besignificant was a 1·5-fold difference between the controland treatment groups. The highest difference detected was6·57-fold, with several others showing around a 4-folddifference. While this level of change is low in comparisonto systems where a single cell type is tested, these data dodemonstrate that this approach can identify FSH-regulatedgenes in a complex cell mixture (i.e. total testis). This is, infact, comparable to the situation that might be required foranalysis of clinical samples, where the small sample sizewould preclude the use of cell separation methods prior toanalysis. The fact that up- and down-regulation of genes ismeasurable following 4 days of treatment indicates that inconditions where FSH suppression has been long term,FSH target gene expression interrogation may be a usefultool for identifying the basis of pathological change. It isimportant to note that some alterations in gene expressionmay be in part attributed to the reduction in testicularweight as a consequence of cell loss. However, we believethe disproportionate change in gene expression (1·5–6fold) compared with the 22% reduction in testis weightfollowing FSH suppression indicates that the changes ingene expression are primarily due to specific FSH effects.
In conclusion, the model used in this study has success-fully identified distinct developmental responses to FSH inSertoli and germ cells in the postnatal rat and identifiedcandidate genes that may underpin these responses at 18dpp. These data illustrate the switch in Sertoli cell functionduring the first spermatogenic wave, as they cease pro-liferation and establish a niche for support of germ celldevelopment. In addition, the emerging dependence ofgerm cells on Sertoli cell-derived products can be exploredthrough identification of FSH-target genes and thepathways in which they function.
Acknowledgements
We thank Liana Nagley for the preparation of RNA priorto microarray analysis, Sue Hayward for measurement ofinhibin levels and Dr Ingrid Sadler-Riggleman for hertechnical assistance.
Funding
Supported by the Wellcome Trust Fellow Scheme(#058479 to S M), and The National Health and Medical
S J MEACHEM and others · FSH effects on the immature testis442
www.endocrinology-journals.orgJournal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access

Research Council of Australia, Program Grants (#050387to S J M; #1147386 to K L L) and Research Fellowship(#143792 to K L L). The authors declare that there is noconflict of interest that would prejudice the impartiality ofthis scientific work.
References
Andersen OM, Yeung CH, Vorum H, Wellner M, Andreassen TK,Erdmann B, Mueller EC, Herz J, Otto A, Cooper TG & WillnowTE 2003 Essential role of the apolipoprotein E receptor-2 in spermdevelopment. Journal of Biological Chemistry 278 23989–23995.
Arkblad EL, Egorov M, Shakhparonov M, Romanova L, Polzikov M& Rydstrom J 2002 Expression of proton-pumping nicotinamidenucleotide transhydrogenase in mouse, human brain and C elegans.Comparative Biochemistry and Physiology. Part B, Biochemistry andMolecular Biology 133 13–21.
Asuncion L, Fogelgren B, Fong KS, Fong SF, Kim Y & Csiszar K2001 A novel human lysyl oxidase-like gene (LOXL4) onchromosome 10q24 has an altered scavenger receptor cysteine richdomain. Matrix Biology 20 487–491.
Baker PJ & O’Shaughnessy PJ 2001 Role of gonadotrophins inregulating numbers of Leydig and Sertoli cells during fetal andpostnatal development in mice. Reproduction 122 227–234.
Beumer TL, Roepers-Gajadien HL, Gademan IS, Kal HB & de RooijDG 2000 Involvement of the D-type cyclins in germ cellproliferation and differentiation in the mouse. Biology of Reproduction63 1893–1898.
Boitani C, Politi MG & Menna T 1993 Spermatogonial cellproliferation in organ culture of immature rat testis. Biology ofReproduction 48 761–767.
Boitani C, Stefanini M, Fragale A & Morena AR 1995 Activinstimulates Sertoli cell proliferation in a defined period of rat testisdevelopment. Endocrinology 136 5438–5444.
Boussouar F, Mauduit C, Tabone E, Pellerin L, Magistretti PJ &Benahmed M 2003 Developmental and hormonal regulation of themonocarboxylate transporter 2 (MCT2) expression in the mousegerm cells. Biology of Reproduction 69 1069–1078.
Bravo-Moreno JF, Diaz-Sanchez V, Montoya-Flores JG, Lamoyi E,Saez JC & Perez-Armendariz EM 2001 Expression of connexin43in mouse Leydig, Sertoli and germinal cells at different stages ofpostnatal development. Anatomical Record 264 13–24.
Brucato S, Bocquet J & Villers C 2002 Regulation of glypican-1,syndecan-1 and syndecan-4 mRNAs expression byfollicle-stimulating hormone, cAMP increase and calcium influxduring rat Sertoli cell development. European Journal of Biochemistry269 3461–3469.
Buemer TL, Roepers-Gajadien HL, Gademan IS, Kal HB & de RooijDG 2000 Involvement of the D-type cyclins in germ cellproliferation and differentiation in the mouse. Biology of Reproduction63 1893–1898.
Buzzard JJ, Wreford NG & Morrison JR 2002 Marked extension ofproliferation of rat Sertoli cells in culture using recombinant humanFSH. Reproduction 124 633–634.
Buzzard JJ, Farnworth PG, De Kretser DM, O’Connor AE &Wreford NG 2003 Proliferative phase Sertoli cells display adevelopmentally regulated response to activin in vitro. Endocrinology144 474–483.
Chaudhary J, Sadler-Riggleman I, Ague JM & Skinner MK 2005 TheHelix-Loop-Helix inhibitor of differentiation proteins inducepost-mitotic terminally differentiated Sertoli cells to re-enter thecell cycle and proliferate. Biology of Reproduction 72 1205–1217.
Chen JK & Heckert LL 2001 Dmrt1 expression is regulated byfollicle-stimulating hormone and phorbol esters in postnatal Sertolicells. Endocrinology 142 1167–1178.
Chomczynski P & Sacchi N 1987 Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction. Analytical Biochemistry 162 156–159.
Cooke PS & Meisami E 1991 Early hypothyroidism in rats causesincreased adult testis and reproductive organ size but does notchange testosterone levels. Endocrinology 129 237–243.
Danzo BJ 1995 The effects of a gonadotropin-releasing hormoneantagonist on androgen-binding protein distribution and otherparameters in the adult male rat. Endocrinology 136 4004–4011.
Dufour JM & Kim KH 1999 Cellular and subcellular localisation of sixretinoid receptors in rat testis during postnatal development:identification of potential heterodimeric receptors. Biology ofReproduction 61:1300–1308.
Eisen MB, Spellman PT, Brown PO & Botstein D 1998 Clusteranalysis and display of genome-wide expression patterns PNAS 9514863–11488.
Ericsson J, Runquist M, Thelin A, Andersson M, Chojnacki T &Dallner G 1993 Distribution of prenyltransferases in rat tissues.Evidence for a cytosolic all-trans- geranylgeranyl diphosphatesynthase. Journal of Biological Chemistry 268 832–838.
Eskild W, Troen G, Blaner WS, Nilsson A & Hansson V 2000Evidence for independent control at the mRNA and protein levelsof cellular retinol binding protein 1 in rat Sertoli cells. Journal ofReproduction and Fertility 119 101–109.
Fantoni G, Morris PL, Forti G, Vannelli GB, Orlando C, Barni T,Sestini R, Danza G & Maggi M 1993 Endothelin-1: a newautocrine/paracrine factor in rat testis. American Journal of Physiology– Endocrinology and Metabolism 265 E267–E274.
Fragale A, Puglisi R, Morena AR, Stefanini M & Boitani C 2001Age-dependent activin receptor expression pinpoints activin A as aphysiological regulator of rat Sertoli cell proliferation. MolecularHuman Reproduction 71 107–114.
Goddard I, Florin A, Mauduit C, Tabone E, Contard P, Bars R,Chuzel F & Benahmed M 2003 Alteration of lactate productionand transport in the adult rat testis exposed in utero to flutamide.Molecular Cell Endocrinology 206 137–146.
Gregory CW & De Phillip RM 1998 Detection of steroidogenic acuteregulatory protein (StAR) in mitochondria of cultured rat Sertolicells incubated with follicle-stimulating hormone. Biology ofReproduction 58 470–474.
Grima J, Wong CC, Zhu LJ, Zong SD & Cheng CY 1998 Testinsecreted by Sertoli cells is associated with the cell surface, and itsexpression correlates with the disruption of Sertoli-germ celljunctions but not the inter-Sertoli tight junction. Journal of BiologicalChemistry 273 21040–21053.
Henriksen K, Kulmala J, Toppari J, Mehrotra K & Parvinen M 1996Stage-specific apoptosis in the rat seminiferous epithelium:quantification of irradiation effects. Journal of Andrology 17 394–402.
Hu J, Shima H & Nakagawa H 1999 Glial cell line-derivedneurotropic factor stimulates Sertoli cell proliferation in the earlypostnatal period of rat testis development. Endocrinology 1403416–3421.
Huang HF, Pogach L, Giglio W, Nathan E & Seebode J 1992GnRH-A induced arrest of spermiogenesis in rats is associated withaltered androgen binding protein distribution in the testis andepididymis. Journal of Andrology 13 153–159.
Ikeda Y & Terashima T 1999 Expression of reelin, the generesponsible for the reeler mutation, in embryonic development andadulthood in the mouse. Developmental Dynamics 210 157–172.
Jackson VN, Price NT, Carpenter L & Halestrap AP 1997 Cloning ofthe monocarboxylate transporter isoform MCT2 from rat testisprovides evidence that expression in tissues is species-specific andmay involve post-transcriptional regulation. Biochemical Journal 324447–453.
Joseph DR, O’Brien DA, Sullivan PM, Becchis M, Tsuruta JK &Petrusz P 1997 Overexpression of androgen-binding protein/sex
FSH effects on the immature testis · S J MEACHEM and others 443
www.endocrinology-journals.org Journal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access

hormone-binding globulin in male transgenic mice: tissuedistribution and phenotypic disorders. Biology of Reproduction 5621–32.
Kabbaj O, Holm C, Vitale C & Pelletier RM 2001 Expression,activity and subcellular localization of testicular hormone-sensitivelipase during postnatal development in the guinea pig. Biology ofReproduction 65 601–612.
Kaestner KH, Hiemisch H & Schutz G 1998 Targeted disruption ofthe gene encoding hepatocyte nuclear factor 3 gamma results inreduced transcription of hepatocyte-specific genes. Molecular CellBiology 18 4245–4251.
von der Kammer H, Hanes J, Klaudiny J & Scheit KH 1994 Ahuman amyloid precursor-like protein is highly homologous to amouse sequence-specific DNA-binding protein. DNA Cell Biology13 1137–1143.
Kastner P, Mark M, Leid M, Gansmuller A, Chin W, Grondona JM,Decimo D, Krezel W, Dierich A & Chambon P 1996 Abnormalspermatogenesis in RXR beta mutant mice. Genes and Development10 80–92.
Kato K, Shinohara H, Kurobe N, Inaguma Y, Shimizu K & OhshimaK 1991 Tissue distribution and developmental profiles ofimmunoreactive alpha B crystallin in the rat determined with asensitive immunoassay system. Biochimica et Biophysica Acta 1074201–208.
Kawasaki Y, Nakagawa A, Nagaosa K, Shiratsuchi A & Nakanishi Y2002 Phosphatidylserine binding of class B scavenger receptor typeI, a phagocytosis receptor of testicular Sertoli cells. Journal ofBiological Chemistry 277 27559–27566.
Kezele PR, Ague JM, Nilsson E & Skinner MK 2002 Alterations inthe ovarian transcriptome during primordial follicle assembly anddevelopment. Biology of Reproduction 72 241–255.
Khan SA, Ndjountche L, Pratchard L, Spicer LJ & Davis JS 2002Follicle-stimulating hormone amplifies insulin-like growth factorI-mediated activation of AKT/protein kinase B signaling inimmature rat Sertoli cells. Endocrinology 143 2259–2267.
Kirby JD, Jetton AE, Cooke PS, Hess RA, Bunick D, Ackland JF,Turek FW & Schwartz NB 1992 Developmental hormonal profilesaccompanying the neonatal hypothyroidism-induced increase inadult testicular size and sperm production in the rat. Endocrinology131 559–565.
Lampa J, Hoogerbrugge JW, Baarends WM, Stanton PG, PerrymanKJ, Grootegoed JA & Robertson DM 1999 Follicle-stimulatinghormone and testosterone stimulation of immature and matureSertoli cells in vitro: inhibin and N-cadherin levels and roundspermatid binding. Journal of Andrology 20 399–406.
Larriba S, Esteban C, Toran N, Gerard A, Audi L, Gerard H &Reventos J 1995 Androgen binding protein is tissue-specificallyexpressed and biologically active in transgenic mice. Journal ofSteroid Biochemistry and Molecular Biology 53 573–578.
Li S, Moore CL, Dobretsova A & Wight PA 2002 Myelin proteolipidprotein (Plp) intron 1 DNA is required to temporally regulatePlp gene expression in the brain. Journal of Neurochemistry 83193–201.
Lui WY, Mruk D, Lee WM & Cheng CY 2003 Sertoli cell tightjunction dynamics: their regulation during spermatogenesis. Biologyof Reproduction 68 1087–1097.
Machiels BM, Ramaekers FC, Kuijpers HJ, Groenewoud JS,Oosterhuis JW & Looijenga LH 1997 Nuclear lamin expression innormal testis and testicular germ cell tumours of adolescents andadults. Journal of Pathology 182 197–204.
Maki JM, Tikkanen H & Kivirikko KI 2001 Cloning andcharacterization of a fifth human lysyl oxidase isoenzyme: the thirdmember of the lysyl oxidase-related subfamily with four scavengerreceptor cysteine-rich domains. Matrix Biology 20 493–496.
Manna P, Wang X-J & Stocco DM 2003 Involvement of multipletranscription factors in the regulation of steroidogenic acuteregulatory protein gene expression. Steroids 68 1125–1134.
Mbikay M, Tadros H, Ishida N, Lerner CP, De Lamirande E, ChenA, El-Alfy M, Clermont Y, Seidah NG, Chretien M, Gagnon C &Simpson EM 1997 Impaired fertility in mice deficient for thetesticular germ-cell protease PC4. PNAS 94 6842–6846.
McChlery SM & Clarke SC 2003 The use of hydrolysis and hairpinprobes in real-time PCR Molecular Biotechnology 25 267–274.
McLachlan RI, Wreford NG, De Kretser DM & Robertson DM 1995The effects of recombinant follicle-stimulating hormone on therestoration of spermatogenesis in the gonadotropin-releasinghormone-immunized adult rat. Endocrinology 136 4035–4043.
McLachlan RI, Wreford NG, Meachem SJ, De Kretser DM &Robertson DM 1994 Effects of testosterone on spermatogenic cellpopulations in the adult rat. Biology of Reproduction 51 945–955.
McLean DJ, Friel PJ, Pouchnik D & Griswold MD 2002Oligonucleotide microarray analysis of gene expression infollicle-stimulating hormone-treated rat Sertoli cells. MolecularEndocrinology 16 2780–2792.
Meachem SJ, McLachlan RI, de Kretser DM, Robertson DM &Wreford NG 1996 Neonatal exposure of rats to recombinanthuman follicle-stimulating hormone increases adult Sertoli andspermatogenic cell numbers. Biology of Reproduction 54 36–44.
Meachem SJ, Wreford NG, Stanton PG, Robertson DM &McLachlan RI 1998 Follicle-stimulating hormone is required forthe initial phase of spermatogenic restoration in adult rats followinggonadotrophin suppression. Journal of Andrology 19 725–735.
Meachem SJ, McLachlan RI, Stanton PG, Robertson DM & WrefordNG 1999 FSH immunoneutralization acutely impairsspermatogonial development in normal adult rats. Journal ofAndrology 20 756–762.
Meehan T, Schlatt S, O’Bryan MK, de Kretser DM & Loveland KL2000 Regulation of germ cell and Sertoli cell development byactivin, follistatin and FSH. Developmental Biology 220 225–237.
Meehan T, Loveland KL, de Kretser D, Cory S & Print CG 2001Developmental regulation of the bcl-2 family duringspermatogenesis: Insights into the sterility of bcl-w-/- male mice.Cell Death and Differentiation 8 225–233.
Meng X, Lindahl M, Hyvonen ME, Parvinen M, de Rooij DG, HessMW, Raatikainen-Ahokas A, Sainio K, Rauvala H, Lakso M,Pichel JG, Westphal H, Saarma M & Sariola H 2000 Regulation ofcell fate decision of undifferentiated spermatogonia by GDNF.Science 287 1489–1493.
Migrenne S, Racine C, Guillou F & Habert R 2003 Pituitaryhormones inhibit the function and differentiation of fetal Sertolicells. Endocrinology 144 2617–2622.
Monaco L, Foulkes NS & Sassone-Corsi P 1995 Pituitaryfollicle-stimulating hormone (FSH) induces CREM gene expressionin Sertoli cells: involvement in long-term desensitization of the FSHreceptor. PNAS 92 10673–10677.
Mori T, Guo MW, Sato E, Baba T, Takasaki S & Mori E2000 Molecular and immunological approaches to mammalianfertilization. Journal of Reproductive Immunology 47 139–158.
Munsie M, Schlatt S, deKretser DM & Loveland KL 1997 Expressionof stem cell factor in the postnatal rat testis. Molecular Reproductionand Development 47 19–25.
Nozaki H, Matsuzawa T, Nakamura T, Arai I & Urashima T 2003Are there two forms of beta 2-N-acetylglucosaminyltransferase I inrat testicular and epididymal fluids? Biochimica et Biophysica Acta1649 140–145.
O’Donnell L, McLachlan RI, Wreford NG & Robertson DM 1994Testosterone promotes the conversion of round spermatids betweenstages VII and VIII of the rat spermatogenic cycle. Endocrinology135 2608–2614.
Odet F, Guyot R, Leduque P & Le Magueresse-Battistoni B 2003Evidence for Similar Expression of Protein C Inhibitor and theUrokinase-Type Plasminogen Activator System during Mouse TestisDevelopment. Endocrinology 145 1481–1489.
S J MEACHEM and others · FSH effects on the immature testis444
www.endocrinology-journals.orgJournal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access

Orth JM 1984 The role of follicle-stimulating hormone in controllingSertoli cell proliferation in testes of foetal rats. Endocrinology 1151248–1255.
Orth JM, Gunsalus GL & Lamperti AA 1988 Evidence from Sertolicell-depleted rats indicates that spermatid number in adults dependson numbers of Sertoli cells produced during perinatal development.Endocrinology 122 787–794.
Orth JM, Jester WF, Li LH & Laslett AL 2000 Gonocyte-Sertoli cellinteractions during development of the neonatal rodent testis.Current Topics in Developmental Biology 50 103–124.
Orwig KE, Ryu BY, Avarbock MR & Brinster RL 2002 Malegerm-line stem cell potential is predicted by morphology of cells inneonatal rat testes. PNAS 99 11706–11711.
O’Shaughnessy PJ, Willerton L & Baker PJ 2002 Changes in Leydigcell gene expression during development in the mouse. Biology ofReproduction 66 966–975.
Packer AI, Besmer P & Bachvarova RF 1995 Kit ligand mediatessurvival of type A spermatogonia and dividing spermatocytes inpostnatal mouse testes. Molecular Reproduction and Development 42303–310.
Pellegrini M, Grimaldi P, Rossi P, Geremia R & Dolci S 2003Developmental expression of BMP4/ALK3/SMAD5 signalingpathway in the mouse testis: a potential role of BMP4 inspermatogonia differentiation. Journal of Cell Science 116 3363–3372.
Penttila TL, Hakovirta H, Mali P, Wright WW & Parvinen M 1995Follicle-stimulating hormone regulates the expression of cyclicprotein-2/cathepsin L messenger ribonucleic acid in rat Sertoli cellsin a stage-specific manner. Molecular and Cellular Endocrinology 113175–181.
Petersen C, Boitani C, Froysa B & Soder O 2001 Transforminggrowth factor-alpha stimulates proliferation of rat Sertoli cells.Molecular and Cellular Endocrinology 181 221–227.
Petersen C, Boitani C, Froysa B & Soder O 2002 Interleukin-1 is apotent growth factor for immature rat sertoli cells. Molecular andCellular Endocrinology 186 37–47.
Rappaport MS & Smith EP 1995 Insulin-like growth factor (IGF)binding protein 3 in the rat testis: follicle-stimulating hormonedependence of mRNA expression and inhibition of IGF-I action oncultured Sertoli cells. Biology of Reproduction 52 419–425.
Raymond CS, Murphy MW, O’Sullivan MG, Bardwell VJ &Zarkower D 2000 Dmrt1, a gene related to worm and fly sexualregulators is required for mammalian testis differentiation. Genes andDevelopment 14 2587–2595.
Reaven E, Zhan L, Nomoto A, Leers-Sucheta S & Azhar S 2000Expression and microvillar localization of scavenger receptor class B,type I (SR-BI) and selective cholesteryl ester uptake in Leydig cellsfrom rat testis. Journal of Lipid Research 41 343–356.
Robertson DM, Hayward S, Irby D, Jacobsen J, Clarke L, McLachlanRI & de Kretser DM 1988 Radioimmunoassay of rat seruminhibin: changes after PMSG stimulation and gonadectomy.Molecular Cellular Endocrinology 58 1–8.
Robertson DM, Giacometti, M, Foulds L, Lahnstein J, Goss NH,Hearn MTW & de Kretser DM 1989 Isolation of inhibinalpha-subunit precursor proteins from bovine follicular fluid.Endocrinology 125 2141–2149.
de Rooij DG 1998 Stem cells in testis. International Journal ofExperimental Pathology 79 67–80.
Rossi P, Dolci S, Albanesi C, Grimaldi P, Ricca R & Geremia R1993 Follicle-stimulating hormone induction of steel factor (SLF)mRNA in mouse Sertoli cells and stimulation of DNA synthesis inspermatogonia by soluble SLF. Developmental Biology 155 68–74.
Russell LD, Alger LE & Nequin LG 1987 Hormonal control ofpubertal spermatogenesis. Endocrinology 120 1615–1632.
Russell LD, Ettlin RA, Sinha Hikim AP & Clegg ED 1990Histological and histopathological evaluation of the testis. Ed. LD Russell.Clearwater, Florida: Cache River Press.
Sadate-Ngatchou PI, Pouchnik DJ & Griswold MD 2004Identification of testosterone-regulated genes in testes ofhypogonadal mice using oligonucleotide microarray MolecularEndocrinology 18 422–433.
Schinkel AH & Jonker JW 2003 Mammalian drug efflux transportersof the ATP binding cassette (ABC) family: an overview. AdvancedDrug Delivery Reviews 55 3–29.
Schlatt S, Zhengwei Y, Meehan T, de Kretser DM & Loveland KL1999 Application of morphometric techniques to postnatal rat testesin organ culture: insights into testis growth. Cell and Tissue Research298 335–343.
Shetty J, Marathe GK & Dighe RR 1996 Specificimmunoneutralization of FSH leads to apoptotic cell death of thepachytene spermatocytes and spermatogonial cells in the rat.Endocrinology 137 2179–2182.
Shima JE, McLean DJ, McCarrey JR & Griswold MD 2004 Themurine testicular transcriptome: Characterizing gene expression inthe testis during the Progression of spermatogenesis. Biology ofReproduction 71 319–330.
Sicinski P, Donaher JL, Geng Y, Parker SB, Gardner H, Park MY,Robker RL, Richards JS, McGinnis LK, Biggers JD, Eppig JJ,Bronson RT, Elledge SJ & Weinberg RA 1996 Cyclin D2 is anFSH-responsive gene involved in gonadal cell proliferation andoncogenesis. Nature 384 470–474.
Simorangkir DR, Wreford NG & de Kretser DM 1997 Impairedgerm cell development in the testes of immature rat with neonatalhypothyroidism. Journal of Andrology 18 186–193.
Skinner MK, Schlitz SM & Anthony CT 1989 Regulation of Sertolicell differentiated function: testicular transferrin andandrogen-binding protein expression. Endocrinology 124 3015–3024.
Smith EP, Dickson BA & Chernausek SD 1990 Insulin-like growthfactor binding protein-3 secretion from cultured rat Sertoli cells:dual regulation by follicle stimulating hormone and insulin-likegrowth factor-I. Endocrinology 127 2744–2751.
Sommersberg B, Bulling A, Salzer U, Frohlich U, Garfield RE,Amsterdam A & Mayerhofer A 2000 Gap junction communicationand connexin 43 gene expression in a rat granulose cell line:regulation by follicle-stimulating hormone. Biology of Reproduction 631661–1668.
Strothmann K, Simoni M, Mathur P, Siakhamary S, Nieschlag E &Gromoll J 2004 Gene expression profiling of mouse Sertoli celllines. Cell Tissue Research 315 249–257.
Suominen JS, Yan W, Toppari J & Kaipia A 2001 The expression andregulation of Bcl-2-related ovarian killer (Bok) mRNA in thedeveloping and adult rat testis. European Journal of Endocrinology 145771–778.
Swinnen JV, Tsikalas KE & Conti M 1991 Properties and hormonalregulation of two structurally related cAMP phosphodiesterases fromthe rat Sertoli cell. The Journal of Biological Chemistry 26618370–18377.
Tadokoro Y, Yomogida K, Ohta H, Tohda A & Nishimune Y 2002Homeostatic regulation of germinal stem cell proliferation by theGDNF/FSH pathway. Mechanisms of Development 113 29–39.
Tapanainen JS, Tilly JL, Vihko KK & Hsueh AJ 1993 Hormonalcontrol of apoptotic cell death in the testis: gonadotropins andandrogens as testicular cell survival factors. Molecular Endocrinology76 43–50.
Tripiciano A, Peluso C, Morena AR, Palombi F, Stefanini M, ZiparoE, Yanagisawa M & Filippini A 1999 Cyclic expression ofendothelin-converting enzyme-1 mediates the functional regulationof seminiferous tubule contraction. Journal of Cell Biology 1451027–1038.
Uhrin P, Dewerchin M, Hilpert M, Chrenek P, Schofer C,Zechmeister-Machhart M, Kronke G, Vales A, Carmeliet P, BinderBR & Geiger M 2000 Disruption of the protein C inhibitor generesults in impaired spermatogenesis and male infertility. Journal ofClinical Investigation 106 1531–1539.
FSH effects on the immature testis · S J MEACHEM and others 445
www.endocrinology-journals.org Journal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access

Ungefroren H, Ergun S, Krull NB & Holstein AF 1995 Expression ofthe small proteoglycans biglycan and decorin in the adult humantestis. Biology of Reproduction 52 1095–1105.
Van Haaster LH, De Jong FH, Docter R & de Rooij DG 1992 Theeffect of hypothyroidism on Sertoli cell proliferation anddifferentiation and hormone levels during testicular development inthe rat. Endocrinology 131 1574–1576.
Walker WH, Sanborn BM & Habener JF 1994 An isoform oftranscription factor CREM expressed during spermatogenesis lacksthe phosphorylation domain and represses cAMP-inducedtranscription. PNAS 91 12423–12427.
West AP, Sharpe RM & Saunders PT 1994 Differential regulation ofcyclic adenosine 3�,5�-monophosphate (cAMP) responseelement-binding protein and cAMP response element modulatormessenger ribonucleic acid transcripts by follicle-stimulatinghormone and androgen in the adult rat testis. Biology of Reproduction50 869–881.
Wolfrum C, Besser D, Luca E & Stoffel M 2003 Insulin regulates theactivity of forkhead transcription factor Hnf-3 beta/Foxa-2 byAkt-mediated phosphorylation and nuclear/cytosolic localization.PNAS 100 11624–11629.
Wolgemuth DJ, Besset V, Liu D, Zhang Q & Rhee K 1998 Role ofcell-cycle genes in the regulation of mammalian meiosis. In GermCell Development, Division, Disruption and Death, pp 49–60. EdZirkin B. New York: Springer-Verlag.
Wreford NG 1995 Theory and practice of stereological techniquesapplied to the estimation of cell number and nuclear volume in thetestis. Microscopy Research and Technique 32 423–436.
Wright WW, Smith L, Kerr C & Charron M 2003 Mice that expressenzymatically inactive cathepsin L exhibit abnormalspermatogenesis. Biology of Reproduction 68 680–687.
Xu J, Beyer AR, Walker WH & McGee EA 2003 Developmentaland stage-specific expression of Smad2 and Smad3 in rat testis.Journal of Andrology 24 192–200.
Yan W, Kero J, Suominen J & Toppari J 2001 Differential expressionand regulation of the retinoblastoma family of proteins duringtesticular development and spermatogenesis: roles in the control ofgerm cell proliferation, differentiation and apoptosis. Oncogenetics 201343–1356.
Yan W, Suominen J, Samson M, Jegou B & Toppari J 2000Involvement of Bcl-2 family proteins in germ cell apoptosis duringtesticular development in the rat and pro-survival effect of stem cellfactor on germ cells in vitro. Molecular and Cellular Endocrinology 165115–129.
Yomogida K, Yagura Y, Tadokoro Y & Nishimune Y 2003 DramaticExpansion of Germinal Stem Cells by Ectopically- ExpressedHuman Glial Cell Line-Derived Neurotrophic Factor in MouseSertoli Cells. Biology of Reproduction 69 1303–1307.
Yoshinaga K, Nishikawa S, Ogawa M, Hayashi S, Kunisada T,Fujimoto T & Nishikawa S 1991 Role of c-kit in mousespermatogenesis: identification of spermatogonia as a specific site ofc-kit expression and function. Development 113 (Suppl) 689–699.
Received 16 May 2005Accepted 27 June 2005
S J MEACHEM and others · FSH effects on the immature testis446
www.endocrinology-journals.orgJournal of Endocrinology (2005) 186, 429–446
Downloaded from Bioscientifica.com at 05/30/2020 10:17:24AMvia free access





























