Embed Size (px)
Citation preview

TERATOLOGY 53:361373 (1996)
Developing Allantois Is a Primary Site of 2'-Deoxycoformycin Toxicity MARK J. AIRHART, CHARLES M. ROBBINS, THOMAS B. KNUDSEN, JOYCE K. CHURCH, AND RICHARD G. SKALKO Department of Anatomy and Cell Biology, James H . Quillen College of Medicine, East Tennessee State University, Johnson City, Tennessee 37614 (M.J.A., C.M.R., JX.C., R.G.S.); Department of Pathology, Anatomy, and Cell Biology, Jefferson Medical College, Philadelphia, Pennsylvania 191 07 (TJ3.K.)
ABSTRACT In this study, an assessment of normal mouse allantoic development and its sen- sitivity to 2'-(R)-deoxycoformycin (dCF; Pentosta- tin) exposure were examined. Both dissecting mi- croscopy and scanning electron microscopy were used to describe the normal growth and morpho- genesis of the mouse allantois over gestational days 7-10 as a preliminary step in evaluating po- tential abnormal allantoic ontogeny and its effect on umbilical cord and placental development. Two abnormal allantoic/umbilical cord phenotypes were observed subsequent to injecting pregnant mice with 5 mg dCF/kg, i.p., on gestational day 7 (GD 7) and evaluating litters on GD 10, 1 1, and 12. Abnormal phenotypes included: ( 1) an allantois which extended approximately halfway across the exocoelom but failed to establish a functional con- tact with the chorion; and (2) a phenotype charac- terized by reduced expansion of the allantois across the chorionic surface, a very thin umbilical cord, and aberrant vascularization throughout the structure. Both abnormal phenotypes exhibited ei- ther an agenesis or hypogenesis of the umbilical cord and chorioallantoic plate, respectively. Nei- ther abnormal phenotype, however, exhibited er- rors in the directionality of allantoic growth toward the chorion nor in the formation of aberrant con- tacts between allantois and adjacent yolk sac or amnionic mesenchyme. Statistical interpretation of the experimental data strongly suggested that ab- normalities in allantoic/umbilical cord development were directly associated with embryolethality as evidenced by a decline in the frequency of abnor- mal allantoic/umbilical cord phenotypes over GD 10-12 (73, 36, and 4%; respectively) and a con- comitant increase in the frequency of implantation site resorptions over the same time period (7, 47, and 78%). These results strongly suggest that the developing allantois is very sensitive to the effects of dCF exposure, and that interference with its de- velopment leads to embryolethality by GD 12. 0 1996 Wiley-Liss, Inc.
During most of mammalian pregnancy, the physio- logical exchange of nutrients and gases between mother and embryo is mediated by the umbilical cord and chori- oallantoic placenta. These transitory organs exhibit a wide range of anatomical variations across species but, for the most part, consist of cellular contributions from the allantois, chorion, ectoplacental cone, and mesome- trial decidua. The allantois is an extraembryonic mem- brane that is common to reptiles, birds, and mammals. In general, it is a hollow sac-like structure of endoderm with an outer covering of vascularized splanchnopleuric mesoderm (Nelson, '53). In primates and rodents, this basic anatomical pattern has been modified so that the endodermal component is either small (primates) or non-existent (rodents) while the mesodermal compo- nent is much more prominent (Mossman, '37; Jollie, '90). In primate species, the endoderm of the allantoic diverticulum and the extraembryonic mesoderm make up the body stalk, the developmental precursor of the umbilical cord and chorioallantoic plate. In rodents, the allantois is exclusively comprised of mesodermal cells which originate from the posterior amniotic fold and grow across the exocoelom to contact and fuse with the chorion. Following contact, vascular channels appear within the allantois which are the precursors of the umbilical artery and vein. The chorioallantoic plate, derived from the fusion between the distal end of the allantois and the chorion, contains the radiating branches of both the umbilical artery and vein; vessels that will eventually provide the fetal circulatory com- ponent of the placental labyrinth. This entire sequence of events (formation, growth, contact) has been shown to occur over a 36 hour time period: from GD 9.0 to 10.5 in the rat (Ellington, '85; Vickers, '85) and from GD 7.50 to 9.0 in the mouse (Theiler, '72; Van Maele-Fabry et al., '90, '92).
In spite of its central role in placentation, the rodent
Received August 23, 1995; accepted April 29, 1996. Address reprint requests to Mark J. Airhart, Ph.D., Department of Anatomy and Cell Biology, James H. Quillen College of Medicine, East Tennessee State University, P.O. Box 70582, Johneon City, TN 37614-0582.
0 1996 WILEY-LISS, INC.

362 M.J. AIRHART ET AL.
allantois has received scant attention with respect to the consequences of its potential for abnormal morpho- genesis. Its role in embryolethality was dramatically demonstrated in the brachyury mouse. Gluecksohn- Schoenheimer (1944) showed that a major characteris- tic of mouse embryos carrying the brachyury gene in the homozygous state (T/T) was an abnormal allantois which failed to grow across the exocoelom. As a result, the umbilical vessels failed to develop connections be- tween the embryonic and maternal circulations and the embryos died by GD 12. Additionally, it has been shown that all-trans-retinoic acid (RA) is highly em- bryolethal when administered by gavage to pregnant rats on GD 9. and that this effect is the result of dam-
embryolethal. The absence of a clear-cut embryolethal abnormality led us to speculate that an extraembry- onic target tissue might be involved.
Preliminary studies have shown that dCF (5 mgkg) treatment on GD 7 causes abnormal morphogenesis of the allantois and umbilical cord (Airhart et al., '92). This abnormality is clearly embryolethal and might account for the high resorption rate noted. The present study will extend these preliminary observations and detail dCFs adverse effect on allantois and umbilical cord formation.
MATERIALS AND METHODS Care and breeding of animals
Y
age to the allantois leading to agenesis or hypogenesis Crl:CD-l (ICR)BR mice from Charles R~~~~ ~ ~ b o ~ ~ - of the umbilical cord and chorioallantoic placenta tories (wilmington, MA) were allowed 2-4 weeks to (Love and Vickers, '76; Vickers, '85). More recent stud- acclimate in A A A L A C - ~ ~ ~ ~ ~ ~ ~ ~ facility under con- ies using recombinant DNA technology to "knock out" trolled environmental conditions of light (0200-2200 the Of adhesion molecule h) temperature, and humidity. Animals were provided
of these two molecules in normal umbilical cord and ison WI) and tap water ad libitum. Four nulliparous placental development. The absence of either molecule females (aged 3-5 months) were in a breeding frequently interfered with the initial contact and ad- cage overnight with one male of the Same strain. The hesion of the growing allantois to the chorion and re- presence of a plug was designated gestational s u h d in embryolethality by GD 12 (Gurtner et al., '95; day 0 (GD 0). Plug-positive dams were removed from
studying the effects of genetic and epigenetic factors on experiment number by earpunch, and placed in a sep- the normal morphogenesis of the umbilical cord and arate cage. placenta is further emphasized in the clinical litera-
Preparation and administration of ture by the existence of a strong association between abnormal umbilical cord and placental development 2'-(R)-deoxycoformycin (pentostatin) with fetal malformations and prenatal death (Ber- nischke and B ~ ~ ~ ~ , 955; ciausen, '89; ~ i ~ ~ ~ i ~ , 992; vis- Under sterile conditions, a stock solution was pre- carello et al., '92). pared by dissolving 7.5 mg of (R)-2'-deoxycoformycin,
Earlier studies from our laboratory have shown that dCF (Parke-Davis, Ann Arbor, MI) in 10 ml deionized a transient inhibition of adenosine deaminase (ADA) water. The precise concentration of the stock solution in the GD 7 embryo/decidual unit of the mouse leads to W a s determined spectrophotometrically using a molar a high percentage of embryolethality. Adenosine deam- extinction Of 8.2 lo3 mol-' at 282 nm inase is a critical enzyme in the purine catabolic path- (Schramm and Baker, '85). The solution was drawn way, t,, &,aminate the natural purines, adeno- into a sterile plastic syringe, and aliquots were deliv- sine, and 2'-deoxyadenosine to either inosine or 2'- ered through a MillexGV 0.22 pm filter unit into au- deoxyinosine (Arch and Newsholme, '78). Utilizing the toclaved 2 ml serum bottles. The serum bottles contain- therapeutic drug 2~~deoxycofomycin (dCF; Penbsta- ing the stock solution were stored at 0-5°C in tin), a tight binding non-competitive inhibitor of ADA accordance with the supplier's recommendation that used in the treatment of various types of leukemia, we 100% of dCF's activity will be maintained in solution found that a single i.p. injetion of dCF (5 mg/kg) into for up to 13 weeks. Five milligrams dCF/kg was admin- pregnant mice on either GD 7 or 8 resulted in a 61- istered at 0900 to GD 7 dams via a single intraperito- 81% resorption rate at GD 12 or 17 (Knudsen et al., '89, neal injection of 0.2 CC for every 30 g total mouse body '92; Airhart et 993). A recent study detailing the weight (Knudsen et al., '89; Airhart et al., '93). At this temporal course of dCF's embryotoxicity showed that there was a POS-
GD 7 exposure caused maximum resorption as early as terior amniotic fold and occasionally the very begin- GD 12 preceded by an equally large percentage of ab- nings of an allantoic bud (Theiler, '72; independent ob- normal embryos at GD 10 (Airhart et al., '93). Statis- servation)- tical evaluation strongly suggested that the abnormal embryos at GD 10 died and were resorbed by GD 12. The embryos displayed a wide range of abnormalities a t GD 10 but analysis of the frequency of specific mal- formations did not identify any one as being clearly
(VCAM-l) O r cx4-intWin demonstrated the T&ad Mouse Breeder Diet (W) (Harlan "&lad, Mad-
Kwee et al.9 '95; Yang et al.3 '95). The significance of the breeding cage, individually weighed, assigned an
appmximate13'
Experimental procedures At 0900 on each of GD 10, 11, and 12, dCF-treated
(n = 10 litters for each day examined) and untreated control dams (n = 10 litters for each day examined)

ALLANTOIS IS A PRIMARY SITE OF dCF TOXICITY 363
were euthanized via C02 asphyxiation. Each uterus was removed and immediately placed in 0.1M phos- phate-buffered saline (PBS). Under a dissecting micro- scope at a magnification of 7 to 30 x , individual im- plantation sites were separated from the uterus and transferred to a Petri dish containing fresh PBS for evaluation of embryo viability (+ heartbeat), embryo growth (crown-rump length), morphogenesis of the em- bryo proper (with respect to the stages outlined in Theiler, 19721, allantoidumbilical cord development (with respect to the stages outlined in this study), and dead or resorbed embryos. Crown-rump lengths as well as the length and largest width of both normal and abnormal allantoides were determined using a dissect- ing microscope equipped with an ocular micrometer calibrated to a stage graticule. Embryos together with their extraembryonic membranes and ectoplacental cones (or placentae) were then fixed for 15 minutes in fresh cold absolute Et0H:Acetic acid (3:1), rinsed in 95% EtOH, and stored in appropriately labeled vials containing 70% EtOH for potential histological exam- ination. In addition, several representative dCF- treated and untreated control specimens from GD 7-10 were reserved and processed for scanning electron mi- croscopy.
Scanning electron microscopy Specimens were fixed for 2 to 4 hours with 3.0% glu-
taraldehyde in 0.1M cacodylate-HCl buffer (pH7.3, 25°C). Following three 30 minute rinses in 0.1M caco- dylate-HC1 buffer, microdissection was performed in an effort to optimize the visualization of the embryo and its umbilical cord and chorioallantoic plate; whereafter the specimens were rinsed in MilliQ reagent grade wa- ter (2 x 30 minutes), dehydrated through a graded ethanol series, and critical point dried from liquid COB (Samdri PVT-3B Critical Point Dryer, Tousimis Re- search Corp., Rockville, MD). Dried specimens were attached to aluminum stubs using a thin layer of tele- vision tube coat (Ted Pella, Inc., Redding, CA) and sputter-coated with gold-palladium for 2 minutes at 40 mA and 75 mTorr (Desk-1 Sputter Coater, Denton Vac- uum, Inc., Cherry Hill, NJ). All specimens were viewed and photographed with a scanning electron microscope at accelerating voltages of 5 to 20 kV (DSM 940, Carl Zeiss, Inc., Thornwood, NY).
Data analysis Individual implantation sites from each control and
dCF-exposed dam were assessed on the basis of four experimental endpoints: (1) Normal, normal develop- ment of both the embryo proper and allantois/umbilical cord (2) Resorption, degenerative embryonic and de- cidual tissues subsequent to intrauterine death; (3) Ab- normal embryo proper, abnormal development exclu- sive to the embryo proper; and (4) Abnormal embryo proper with abnormal allantoidumbilical cord, abnor- mal development of both the embryo proper and allan-
toidumbilical cord. The only remaining outcome, i.e., normal embryo proper with abnormal allantoidumbil- ical cord, was never observed. A "dCF" prefix for each of the endpoints was used to distinguished data from dCF-treated and control litters. The various types of malformations of the embryo proper and their relative frequencies were not included in the current study but have been described in an earlier report (Airhart et al., '93). Briefly, the morphological criteria used to evalu- ate the embryo proper included: neural tube closure, craniofacial and limb development, turning of the em- bryo, and growth retardation.
The percent frequencies for each endpoint on gesta- tional days 10, 11, and 12 were used in the analysis of control and dCF-treated litters. The mean frequencies of each endpoint were subjected to statistical evalua- tion using the one-way analysis of variance procedure across GD 10, 11, and 12. Simple linear regression and correlation analysis was used to evaluate the relation- ship between the frequencies of abnormal embryo proper with abnormal allantois/umbilical cord and re- sorptions in an effort to determine if a significant causal relationship existed between the two variables across GD 10, 11, and 12. Student's t-test for indepen- dent samples was applied to evaluate the mean fre- quencies of each dCF-treated group to its respective untreated control group for each gestational day exam- ined. A probability level of 0.05 was used to determine statistical significance for all tests performed in this study.
RESULTS Normal allantoic/umbilical cord development Allantois and umbilical cord development in the
mouse has been found to occur between gestational times GD 7.5-10.0 (60 h) beginning with the formation of the allantoic bud and ending with the fully devel- oped umbilical cord and chorioallantoic plate (Fig. la,f). Previous investigations have divided the process into two basic stages: stage I, growth of the allantois across the exocoelom and fusion with the chorion (ap- proximately GD 7.5-8.5); and stage 11, subsequent growth of the fused allantois into the fully formed um- bilical cord and chorioallantoic plate (GD 8.5-10.0) (Downs and Davies, '93; Theiler, '72; Van Maele-Fabry, '90, '92). The latter stage, in particular, has received
Abbreviations: A allantois AMD antimesometrial decidua C chorion E exomelom E X edoplacental cone L labyrinth M mesencephalic fold MD mesometrial decidua PAL proximal allantoic layer VYS visceral yolk sac * umbilical cord
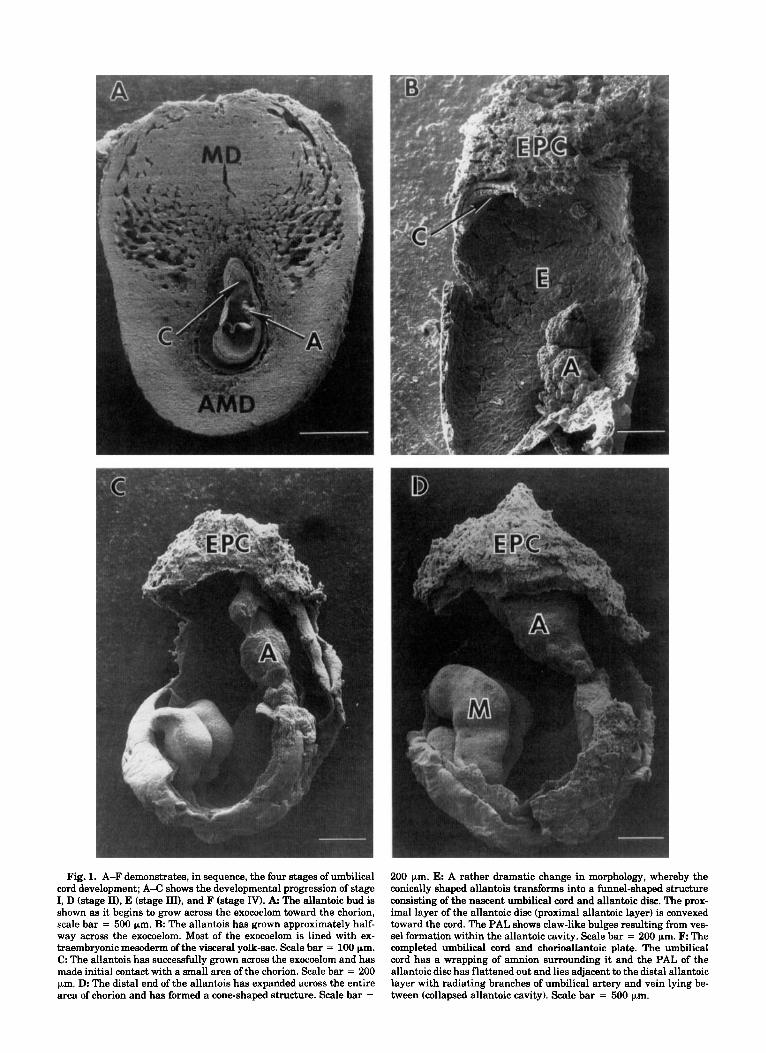
Fig. 1. A-F demonstrates, in sequence, the four stages of umbilical cord development; A-C shows the developmental progression of stage I, D (stage II), E (stage III), and F (stage IV). A The allantoic bud is shown as it begins to grow across the exocoelom toward the chorion, scale bar = 500 pm. B: The allantois has grown approximately half- way across the exocoelom. Most of the exocoelom is lined with ex- traembryonic mesoderm of the visceral yolk-sac. Scale bar = 100 pm. C. The allantois has successfully grown across the exocoelom and has made initial contact with a small area of the chorion. Scale bar = 200 pm. D The distal end of the allantois has expanded across the entire area of chorion and has formed a cone-shaped structure. Scale bar =
200 pm. E: A rather dramatic change in morphology, whereby the conically shaped allantois transforms into a funnel-shaped structure consisting of the nascent umbilical cord and allantoic disc. The prox- imal layer of the allantoic disc (proximal allantoic layer) is convexed toward the cord. The PAL shows claw-like bulges resulting from ves- sel formation within the allantoic cavity. Scale bar = 200 pm. F The completed umbilical cord and chorioallantoic plate. The umbilical cord has a wrapping of amnion surrounding it and the PAL of the allantoic disc has flattened out and lies adjacent to the distal allantoic layer with radiating branches of umbilical artery and vein lying be- tween (collapsed allantoic cavity). Scale bar = 500 pm.

ALLANTOIS IS A PRIMARY SITE OF dCF TOXICITY 365
Fig. 1JC-F.

366 M. J. AIRHART ET AL.
TABLE 1. Incidence of allantoic/umbilical cord abnormalities in response to dCF treatment (5 mgkg) on GD 7
No. of litters examined 10 10 10 10 10 10 Implantsflitter 13.1 f 0.53 12.2 f 0.53 12.6 f 0.76 12.0 f 1.7 13.1 f 0.53 13.0 f 2.2 Survivors' 94.4 f 1.69% 92.5 f 4.25% 98.5 f 1.01% 52.5 f 2.73%* 94.5 f 1.68% 21.7 f 4.62%* Resorptions 5.6 f 1.69% 7.5 f 4.25% 1.5 f 1.01% 47.5 f 2.74%* 6.2 f 1.57% 78.3 f 4.62%* Normal embryos 92.7 f 1.83% 17.9 f 5.72%* 96.9 f 1.31% 16.3 f 2.95%* 92.8 f 2.08% 16.4 f 4.72%* Embryos with 1.7 f 0.53% 1.7 f 1.16% 1.6 f 1.12% 0.0 f 0.00% 1.0 f 1.00% 1.5 f 0.99%
abnormalities exclusive to the embryo proper (abnormal embryo proper)
abnormalities both in the embryo proper and in the allantoisl umbilical cord
Embryos with 0.0 2 0.00% 72.9 f 6.41%* 0.0 f 0.00% 36.2 f 3.57%* 0.0 f 0.00% 3.8 t 3.02%
'Survivors are defined as the total number of embryos that have not been resorbed. Data reported on a per litter basis as means f SEM or mean percentages f SEM. Embryos with abnormalities exclusive to the allantoishmbilical cord were never observed. *P < 0.05 vs. untreated (control) group, t-test for independent samples.
only cursory examination with few details describing the continuing morphogenesis of the allantois.
The current study required a more thorough morpho- logical description of allantoic development than was available in the literature. Our electronmicroscopic ex- amination has confirmed and extended previous find- ings and includes the following description: stage I, growth of the club-shaped allantois across the exocoe- lom and its subsequent contact with the chorion, Theiler stages 11,12 (Fig. la-c); stage 11, the expansion of the distal end of the allantois across the total area of chorion resulting in its transformation from a club- shaped process to a truncated cone with the apex clos- est to the embryo, Theiler stage 13 (Fig. Id); stage 111, progressive morphogenesis beginning in the region proximal to the embryo forming a nascent umbilical cord containing the developing umbilical artery and vein which branch radially within the allantoic cavity, bounded by the remaining conical sheet of allantoic cells just distal to the cord, proximal allantoic layer, and the contiguous sheet lying in contact with the chorion, distal allantoic layer, Theiler stages 14, 15 (Fig. le); stage IV, establishment of the completed um- bilical cord and chorioallantoic plate with a prominent cord carrying the umbilical artery and vein from the embryo to the center of the chorioallantoic plate (col- lapsed walls of allantoic cavity plus chorion) where each vessel forms a radiating branching pattern within the plate, Theiler stage 16 (Fig. If). It is noteworthy that allantoic morphogenesis described in stages I and I1 is identical to comparable stages described in rats using scanning electronmicroscopy (Ellington, '85).
Characterization of abnormal allantoic/umbilical cord development
Administration of 5 mg dCF/kg intraperitoneally to female mice on gestational day 7 resulted in specific,
reproducible defects in allantois and umbilical cord morphology (GD 10 = 73%) and subsequent fetal re- sorption (GD 12 = 78%) (Table 1). Treated embryos exhibited two basic types of allantoic/umbilical cord abnormalities which were arbitrarily designated with a capital letter followed by a subscript indicating the normal stage that each most closely resembled (A, and BIII). The A, allantois/cord extended approximately halfway across the exocoelom, and, unlike normal al- lantoides, contained randomly placed blood vessels con- taining fetal red blood cells (Fig. 2a,c,). In addition, the A, allantois/cord was nearly spherical except in the im- mediate proximal region (long axis = 0.62 mm f 0.02 SEM, short axis = 0.56 mm f 0.03 SEM, n = 27), a morphology very different from the normal, club- shaped allantois a t stage I that has grown halfway across the exocoelom (long axis = 0.66 mm f 0.03 SEM, short axis = 0.21 mm f 0.03 SEM, n = 20). The B,, allantois/cord consisted of an umbilical cord and chorioallantoic plate but both were much smaller in diameter than normally found on GD 10 (Fig. 2b). Vas- cularization was also abnormal. The umbilical cord contained several, small diameter vessels, whose con- tinuity with vessels of the plate was obscure. Vessels observed in the plate were small in diameter and were not arranged in any radiating, symmetrical pattern (Fig. 2d). Of the two abnormal phenotypes observed on GD 10, the A, was more frequent (70 out of a total 89
Ab breuiutions: AC allantoic cavity BV blood vessel DAL distal allantoic layer FL forelimb H heart uc umbilical cord

C.
A,
Fig. 2. A A scanning electron micrograph of a type A, unfused allantois associated with an abnormal GD 10 embryo treated with 5 mg dCFkg on GD 7. Note the abnormal spherical shape of the allan- tois. Scale bar = 500 pm. B A scanning electron micrograph of an abnormal embryo with a type B,,, umbilical cord/placenta subsequent to dCF-treatment on GD 7. The diameters of both the umbilical cord and chorioallantoic plate are much smaller than observed in the un- treated, control GD 10 embryo. The allantoic component of the chori- oallantoic plate does not completely expand across the chorion. The two small arrows indicate the perimeter of the proximal allantoic layer while the perimeter of the chorion is defined by the attached fragments of the visceral yolk sac (large arrow). Scale bar = 500 em. C: A schematic representation of a type A, unfused allantois empha- sizing the nearly spherical shape of the “static” allantois and its po-
D.
B 111
sition approximately halfway across the exocoelom. The attenuated elongation of the allantois results in the absence of allantoic tissue adhering to the chorion, and hence the absence of a chorioallantoic plate. Note several short, blood-filled vessels depicted on the surface of the abnormal allantois. D A schematic representation of a type B,,, umbilical cordlplacenta emphasizing the relatively small diameter of both the umbilical cord, and chorioallantoic plate. The allantois has not expanded completely across the chorion and a large cavity, the allantoic cavity, remains between the proximal and distal allantoic layers of the allantoic disc. The vasculature depicted is abnormal, short, discontinuous blood-filled vessels are shown in both the umbil- ical cord and just below the proximal allantoic layer within the al- lantoic cavity.

368 M.J. AIRHART ET AL.
TABLE 2. Absolute rate of decline in abnormal aUantois/umbilical cord phenotypes over gestational
days 10, 11,12 subsequent to GD 7 dCF exposure (5 mgkg)
Frequency of phenotype
70 20 0 19 24 5 89 44 5 No. embryos
Total no. of implants 122 120 130
Allantois/umbilical cord phenotype GD 10 GD 11 GD 12 A1 BIII
with abnormal allantoid umbilical cords
abnormal allantoideskords) while the second abnormal allantois/cord phenotype, BnI, made up the remaining 19 (Table 2). Both abnormal allantoickord types ap- peared to have resulted from premature developmental arrest and/or miniaturization of a normal stage of de- velopment.
At GD 11,96 hours after dCF exposure, there was a marked increase in the number of resorbed implanta- tion sites (7 to 48%) and a corresponding decrease in the number of embryos with an abnormal allantoid umbilical cord (73 to 36%) (Table 1). The relative pro- portion of abnormal allantoidcord types changed abruptly from GD 10 to 11. By GD 11, the absolute number of embryos with an AI allantoiskord decreased from 70 to 20 over the 24 hour interval, over-shooting slightly the number of resorbed embryos, 50 vs. 48 re- sorbed, while the number of embryos with BIII allan- toideskords remained essentially the same. Therefore, the change in the relative proportion of embryos with abnormal allantoidcord types was due to the high per- centage of AI embryos that died and were resorbed by GD 11 (Table 2).
When litters were examined 120 hours (GD 12) after dCF treatment, 78% of all implantation sites were re- sorbed, while only 4% represented embryos with abnor- mal allantoides/cords (Table 1). The high percentage of resorptions on GD12 was found to be equivalent to the maximum resorption percentages reported on either GD 12 or GD 17 in previous studies in which GD 7 dams were injected with 5 mg dCF/kg (78,61, and 82%) (Airhart et al., ’93; Knudsen et al., ’89, ’92, respec- tively). Of the 5 embryos exhibiting abnormal allan- toidumbilical cord development, all were BIIIs (Table 21, a profile of abnormal allantoidumbilical cord types very different from that recorded for either GD 10 or 11 (Table 2). The slower decline of embryos with BIII al- lantoideskords over GD 10-12, as compared to em- bryos with AI allantoides/cords, suggests that the BIn phenotype is slightly better a t maintaining embryo viability. This may be due to the fact that the BIII al- lantoiskord makes some degree of contact with the chorion and therefore has some potential for placental function.
Relationship between allantoichmbilical cord morphogenesis and abnormalities of the
embryo proper Nearly all treated embryos showing abnormal allan-
toichmbilical cord development also exhibited one or more abnormalities of the embryo proper (Table 1). Ex- ceptions to this included 1 or 2 embryos per dCF treat- ment group (10 litters) that exhibited, exclusively, an abnormality of the embryo proper, usually opened cra- nial neural tube. Since control groups exhibited a fre- quency identical to those noted above, these embryos probably reflect a low frequency of “spontaneous” ab- normal morphogenesis unrelated to allantoiclumbilical cord function.
Abnormal allantoic/umbilical cord morphogenesis was discovered to be tightly correlated with intrauter- ine growth retardation as determined by crown-rump length (CRL). Embryos with either an AI or BIII abnor- mal allantoishmbilical cord were significantly smaller in CRL than dCF normals or control embryos (Table 3). The small difference in CRL over GD 10 and 11 be- tween abnormal embryo with abnormal allantoides1 cords suggests that neither phenotype is functionally capable of supporting anything other than minimal growth (Table 3). In addition, embryos designated dCF normals demonstrated a small but significant reduc- tion in CRL when compared with control embryos (Ta- ble 3). This trend of growth retardation persisted through GD 11; however, by GD12, the CRL for both control and dCF normal embryos were found to be sta- tistically indistinguishable. The small discrepancy be- tween control and dCF normal embryos may indicate some limited effect of dCF on all embryos of an injected dam up through GD 11.
Relationship between embryos with abnormal allantoic/umbilical cord development and
resorbed embryos Because of the close temporal association between
abnormal embryos with abnormal allantoideshmbili- cal cords and resorptions, we set out to determine if they were quantitatively related; that is, does the re- lationship imply a high correlation between abnormal allantoichmbilical cord development and death/resorp- tion of the embryo? An inverse relationship between the frequency of embryos with abnormal allantoided cords and resorbed embryos over GD 10-12 appeared to exist (Fig. 3). Quantitative data showed that the max- imal occurrence of abnormal embryos with abnormal allantoideskords on GD 10 (73%) was followed by a maximal resorption rate on GD 12 of 78% (Fig. 3). The accuracy of this data was further supported by the fact that the percentage of dCF normals did not change over the experimental period and that dCF normals over all gestational times exhibited exclusively normal umbil- ical cords/placentae (Fig. 3). Linear regression analysis revealed a significant decrease in the frequency of ab-

ALLANTOIS IS A PRIMARY SITE OF dCF TOXICITY 369
TABLE 3. Relationship between allantoidumbilical cord morphogenesis and crown-rump lengths (CRL)'
GD 10 GD 11 GD 12 Umbilical cord phenotype n CRL (mm) n CRL (mm) n CRL (mm) A1 70 2.70 t 0.09* 20 3.04 f 0.11* 0 - BIII 19 3.27 t 0.14* 24 3.39 f 0.17* 5 4.01 f 0.09* dCF normal 22 4.10 2 0.09* 19 6.12 f 0.10* 22 8.26 f 0.24 Untreated (control) 121 5.32 & 0.07 122 7.14 f 0.06 122 8.31 * 0.12 'n = number of embryos in sample. Values for untreated groups do not include CRLs from abnormal embryos. CRL data reported as means 2 SEM. *P < 0.05 vs. Untreated (control) group, t-test for independent samples.
normal embryos with abnormal allantoidedcords and a concurrent increase in resorbed embryos over GD 10- 12. When the frequencies of resorbed embryos and ab- normal embryos with abnormal allantoidedcords were plotted for all gestational days, linear regression and correlation analysis demonstrated that these two out- comes were indeed highly associated (Y = 77.3- 0.892X, r = -0.891; P < 0.001) (Fig. 4).
DISCUSSION This study demonstrates that dCF interferes with
multiple stages of umbilical cord and placental devel- opment by inhibiting the normal morphogenesis of the allantois. dCF toxicity interferes with two critical events in allantoic development: (1) growth across the exocoelom (AI); and (2) expansion of the allantois across the chorion and over-all growth of the allantois into the umbilical cord and chorioallantoic plate (Bm). The temporal aspects of allantoic development corre- late well with the stage-dependent nature of dCF tox- icity, i.e., GD 7, GD 8 (Downs and Davies, '93; Kaufman, '90; Van Maele-Fabry, '90, '92; Theiler, '90). The allantois begins to grow across the exocoelom at GD 7.50 and makes contact with the chorion by GD 8.5 to 8.75, a period nearly completely overlapping the crit- ical period of dCF toxicity. Further, the stage of um- bilical cord development requiring the longest period of time, allantoic growth across the exocoelom, is the stage most frequently affected.
This study has also provided evidence that the allan- tois is a major site of action for dCF embryolethality. A recent study demonstrated that GD 7 embryos exposed to dCF exhibited a wide spectrum of abnormalities by GD 10; these included: absence of neural tube closure, abnormal craniofacial and limb development, absence of embryo turning, and growth retardation. None of these individual abnormalities, however, could quanti- tatively account for the high percentage of dead and resorbed embryos by GD 12 (Airhart et al., '93). The present data shows a nearly 1:l relationship between the frequency of GD 10 treated embryos with allantoid umbilical cord defects and the frequency of resorbed embryos at GD 12. Further, the increased resorption frequency between GD 10 and 11 is accounted for by an equal but opposite decreased frequency of embryos with an abnormal allantoishmbilical cord. Finally, the
lethal potential of allantoic defect is clearly supported in the literature (Gluecksohn-Schoenheimer, '44; Love and Vickers, '76; Vickers, '85; Gurtner et al., '95; Kwee et al., '95; Yang et al., '95).
Teratogens affecting allantoic development The importance of normal allantois ontogeny and
umbilical cord function in sustaining normal embry- onic development is undeniable and emphasizes the po- tential embryolethal effect of any xenobiotic that af- fects either structure. Retinoic acid has been shown to interfere with normal allantoichmbilical cord develop- ment by causing allantoic agenesis (Vickers, '85). The inhibitory effect of retinoic acid on allantoic develop- ment was found to be stage specific, acting exclusively over gestational days 9 and 10 in rat, i.e., the period of allantoic growth across the exocoelom. Two basic types of abnormal allantoidedumbilical cords were observed and their morphology was similar to the AI and BIII allantoideshmbilical cords described in the present study. The mechanism of retinoic acid toxicity was pos- tulated to be inhibition of allantoic cell proliferation resulting in allantoic agenesis or a miniaturized um- bilical cord and chorioallantoic plate with immature vascularization (Vickers, '85).
In addition, several other teratogens have been shown to cause allantoic dysmorphogenesis and a high percentage of embryolethality subsequent to early ges- tational exposure. Examples of such compounds in- clude: myleran (Alexandrov, '66), cadmium chloride (Khera, '9U, ethylene glycol (Khera, '91), and stauro- sporine (Fujinaga et al., '94). The diverse mechanisms of toxicity exhibited by these compounds suggest that the developing rodent allantois is very sensitive to chemical insult. Furthermore, the impact of abnormal allantoidumbilical cord formation on dysmorphogene- sis of the embryo proper has broad developmental im- plications. The present study shows that dCF treated embryos always exhibit both abnormalities of the em- bryo proper along with abnormal allantoichmbilical cord development. These results raise the following possibilities: (1) dCF treatment on GD 7 interferes with normal allantoichmbilical cord development, which in turn interferes with normal embryo morphogenesis; or (2) dCF treatment affects multiple target sites within the embryo and developing allantoishmbilical cord.

370 M.J. AIRHART ET AL.
loo e T I T 80
70
30
20
10
0 10 11 12
Gestational Day Examined
Fig. 3. Frequency histogram showing the relationship among em- bryos characterized as dCF normals, embryos having abnormalities of the embryo proper with abnormal allantois/umbilical cord, and re- sorptions on GD 10-12 subsequent to 5 mg dCFkg treatment on GD 7. One-way ANOVA revealed significant differences for embryos hav-
The present data does not allow us to distinguish be- tween these choices.
Potential homology between rodent allantois and human body stalk
The pertinence of abnormal allantoidumbilical cord development in rodents to abnormal human cord and placental development depends upon whether there is a homologous structure to the allantois in humans. The body stalk (connecting stalk) appears to be such a structure; it develops during early gestation exclu- sively from extraembryonic mesoderm and is critical for the formation of the umbilical vein and arteries and their functional connections with the placental villi (Enders and King, '88; Hertig and Rock, '41; Luckett, '78; O'Rahilly, '73; Streeter, '42). The allantois in hu- mans is not homologous to the rodent allantois since it is an endodermal evagination of the hindgut which eventually becomes entrapped in the growing body stalk (Luckett, '78). A detailed comparison between the ontogeny of rodent allantois and human body stalk suf- fers from the lack of details regarding body stalk de- velopment. There is controversy regarding the lineage of extraembryonic mesoderm cells composing the body stalk and the details of body stalk morphogenesis. The most recent findings indicate that the body stalk con- sists of caudal epiblast cells that migrate through a
0 rn
dCF Normal Abnormal embryo proper with abnormal allantoidumbilical cord* Resorption*
ing abnormalities of the embryo proper with abnormal allantoidum- bilical cord and resorptions. Note there was no statistical difference in the frequency of treated normals over GD 10-12. *P < 0.05 over GD 10-12, one-way ANOVA. All points are means ? SEM.
precociously formed primitive streak. A population of these cells migrates from the caudal end of the embryo to a location between the developing amnion and adja- cent cytotrophoblast region (chorion) forming the rudi- mentary body stalk (Luckett, '71, '78). Even if the spe- cifics of this developmental scenario are found to be incorrect, it is clear the body stalk consists of a popu- lation of extraembryonic mesodermal cells and any dis- ruption of cell accumulation within the body stalk would result in varying degrees of umbilical cord and placental abnormalities. Because of the importance of umbilical cord and placental function in sustaining the growth and the development of the embryo, any abnor- mality in either structure may be life threatening. In fact, the high percentages of spontaneous abortions and miscarriages may be partly due to abnormal body stalk development (Hertig et al., '59; Hertig, '67). Therefore, further insights into potential chemical sensitivities of body stalk (allantois) development are an important avenue for additional research.
Proposed mechanism of dCF toxicity In spite of the significance of umbilical cord and pla-
cental defects in terms of their incidence and embryo- lethal effects, relatively little data exist concerning the cellular basis for their genesis (Coulam et al., '90; Thom et al., '92). Analysis of abnormal umbilical cord

ALLANTOIS IS A PRIMARY SITE OF dCF TOXICITY 371
100 r 1 I I I I I I I I I I - 0 -
90 t- 0
80 W
W
10
0 0‘ .
1
0 GD 1OLitters V GD 11 Litters 0 GD12Litters
0 10 20 30 40 50 60 70 80 90 100 Resorption (%)
Fig. 4. Scatter diagram of resorbed embryos vs. embryos having abnormalities of the embryo proper with abnormal allantoishmbili- cal cord in each litter examined on GD 10, 11, and 12 subsequent to 5 mg dCF/kg treatment on GD 7. Linear regression and correlation analysis revealed a significant straight-line trend and a high corre-
lation in the relationship between the frequency of resorbed embryos and embryos having abnormalities of the embryo proper with abnor- mal allantoidumbilical cord over the gestational days examined (Y = 77.3-0.892X; r = -0.891; P < 0.001). Dashed lines indicate 95% confidence intervals.
and chorioallantoic plate development in terms of the normal stages of their ontogeny affords some opportu- nity for speculating on the mechanism of dCF toxicity. This approach, unfortunately, does not narrow the fo- cus of toxicity since three stages of normal develop- ment (I, 11, III) may be involved. The vast majority of dCF treated GD 10 embryos with abnormal allantoic and umbilical cord formation (73%) exhibited what ap- pears to be inhibition of normal allantoic growth across the extraembryonic coelom (exocoelom), a defect asso- ciated with stage I of development. In the second type of abnormal allantoidumbilical cord (B,) the allantois apparently grew successfully across the exocoelom but its expansion across the chorion was diminished, the diameter of the umbilical cord was much smaller than normal and vascular development was decidedly im- paired. These defects incorporate events occurring over stages I1 and 111. There is, however, a common biolog- ical mechanism, the regulation of cell division amongst allantoic cells, that when altered, could account for both abnormal types. If dCF toxicity inhibits cell pro- liferation in the allantois, this alone could account for the two types of abnormal allantoideshmbilical cords observed. Allantoic cells undergoing a smaller number
of mitotic divisions could clearly result in a “short” allantois, one that never grows completely across the exocoelom and results in the type AI allantoichmbili- cal cord. The same toxic mechanism can also account for the Type BIII allantoishmbilical cord. It can be hy- pothesized that dCF decreases the rate of allantoic cell proliferation at a slightly later stage of allantoic elon- gation, for example when the allantois has grown about half of the distance across the exocoelom. Such variability of allantoic elongation between embryos of the same litter has been observed by several investi- gators (Downs and Gardner, ’95; Theiler, ’89). Cell ac- cumulation continues but at an abnormally slow rate such that a slender allantois just makes contact with the chorion and undergoes a minimal amount of expan- sion across the chorion with minimal vascularization.
Thus far we have no direct evidence to support this hypothesis but there is some correlative data. The large size of the rodent allantois and its rapid ontogeny speak to its dependence on rapid cell division. Over a period of 36 hours the rodent allantois grows more rap- idly than any other tissue in the embryo (Snell and Stevens, ’66). In addition, earlier studies using the de- veloping rat embryo have demonstrated a rapid rate of

372 M.J. AIRHART ET AL.
cell division in the allantois by comparing mitotic in- dexes observed between allantoic cells and cells of other embryonic tissues (Ellington, ’85). One study, us- ing retinoic acid as a teratogen, demonstrated a de- crease in the mitotic index of treated vs. normal allan- toic cells (Vickers, ’85). The observations support the notion that rapid cell proliferation is critical for allan- toic ontogeny. In addition, a study by Gao et al. (‘94) demonstrated that dCF (5 mgkg) injection into GD 7 pregnant mice caused a large increase in physiological cell death (apoptosis) in nearly all tissues/organs (head-fold, spinal cord, body wall, and somites) of the E8 embryo except the amnion, heart, and allantois. These findings seem to rule out the enhancement of physiological cell death as the mechanism of allantoic agenesis and adds focus to the possibility that interfer- ence with cell proliferation is the mechanism of dCF toxicity. We are presently devising an in vitro culture system involving allantoic explants to further explore the hypothesis that dCFs action on purine metabolism inhibits cell proliferation in the allantois.
ACKNOWLEDGMENTS We thank the Parke-Davis Division of Warner-Lam-
bert (Ann Arbor, MI) for their generous gift of 2’-deox- ycoformycin. We are grateful to Dr. Fred E. Hossler for his technical assistance in use of the scanning electron microscope, Dr. Judith A. Airhart for her helpful com- ments and discussions, and Mrs. Tonya Ward for her editorial assistance in the preparation of the manu- script.
LITERATURE CITED Airhart,M.J., C.M. Robbins, J.K. Church, andR.G. Skalko (1992)The
allantois is a primary site of dCF toxicity. Teratology, 45:503 (ab- stract).
Airhart, M.J., C.M. Robbins, T.B. Knudsen, J.K. Church, and R.G. Skalko (1993) Occurrence of embryotoxicity in mouse embryos fol- lowing in utero exposure to 2’-deoxycoformycin (pentostatin). Ter- atology, 47:17-27.
Alexandrov, V.A. (1966) Analysis of the lethal effect of “myleran” on rat embryos. Nature (Lond.), 209:1215-1218.
Arch, J.R.S., and E.A. Newsholme (1978) The control of the metabo- lism and the hormonal role of adenosine. Essays Biochem., 14182- 123.
Benirschke, K., and W.H. Brown (1955) A vascular anomaly of the umbilical cord. Obstet. Gynecol., 6:399-404.
Clausen, I. (1989) Umbilical cord anomalies and antenatal fetal deaths. Obstet. Gynecol. Survey, 44:841-845.
Coulam, C.B., W.P. Faulk, and J.A. McIntyre (1990) Spontaneous and recurrent abortions. In: Current Therapy in Obstetrics and Gyne- cology, vol. 3. E.J. Quilligan, and F.P. Zuspan, eds. W.B. Saunders, Philadelphia.
Downs, K.M., and T. Davies (1993) Staging of gastrulating mouse embryos by morphological landmarks in the dissecting microscope. Development, 118:1255-1266.
Downs, K.M., and R.L. Gardner (1995) An investigation into early placental ontogeny: allantoic attachment to the chorion is selective and developmentally regulated. Development, 121 :407-416.
Ellington, S.K.L. (1985) A morphological study of the development of the allantois of rat embryos in vivo. J. Anat., 142:l-11.
Enders. A.C.. and B. Kine: (1988) Formation and differentiation of
extraembryonic mesoderm in rhesus monkey. Am. J. Anat., 181:
Fujinaga, M., H.W. Park, T.H. Shepard, P.E. Mirkes, and J.F. Baden (1994) Staurosporine does not prevent adrenergic-induced situs in- versus, but causes a unique syndrome of defects in rat embryos grown in culture. Teratology, 50.261-274.
Gao, X., M.R. Blackburn, and T.B. Knudsen (1994) Activation of apop- tosis in early m o w embryos by Z‘deoxyadenosine exposure. Ter- atology, 49:l-12.
Giacoia, G.P. (1992) Body stalk anomaly: congenital absence of the umbilical cord. Obstet. Gynecol., 80:527-529.
Gluecksohn-Schoenheimer, S. (1944) The development of normal and homozygous brachy (T/T) mouse embryos in the extraembryonic coelom of the chick. Proc. Natl. Acad. Sci. U.S.A., 30:134-140.
Gurtner, G.C., V. Davis, H. Li, M.J. McCoy, A. Sharpe, and M.I. Cy- bulsky (1995) Targeted disruption of the murine VCAMl gene: es- sential role of VCAM-1 in chorioallantoic fusion and placentation. Genes Dev., 9:l-14.
Hertig, A.T. (1967) The overall problem in man. In: Comparative Aspects of Reproductive Failure. K. Bernischke, ed. Springer Ver- lag, New York.
Hertig, A.T., and J. Rock (1941) Two human ova of the previllous stage, having a developmental age of about eleven and twelve days respectively. Contrib. Embryol. Carnegie Inst., 29:127-156.
Hertig, A.T., J. Rock, E.C. Adams, and M.C. Menkin (1959) Thirty- four fertilized human ova, good, bad and indifferent, recovered from 210 women of known fertility. Pediatrics, 23:202-213.
Jollie, W.P. (1990) Development, morphology and function of the yolk- sac placenta of laboratory rodents. Teratology, 41:361-381.
Khera, K.S. (1991) Chemically induced alterations in maternal ho- meostasis and histology of conceptus: their etiologic significance in rat fetal anomalies. Teratology, 44:259-297.
Knudsen, T.B., M.K. Gray, J.K. Church, M.R. Blackburn, M.J. Airhart, R.E. Kellems, and R.G. Skalko (1989) Early postimplan- tation embryolethality in mice following in utero inhibition of adenosine deaminase with 2’deoxycoformycin. Teratology, 40:615- 626.
Knudsen, T.B., R.S. Winters, S.K. Otey, M.R. Blackburn, MJ. Airhart, J.K. Church, and R.G. Skalko (1992) Effects of (R) deoxy- coformycin (pentostatin) on intrauterine nucleoside catabolism and embryo viability in the pregnant mouse. Teratology, 45:91-103.
Kwee, L., H.S. Baldwin, H.M. Shen, C.L. Stewart, C. Buck, C.A. Buck, and M.A. Labow (1995) Defective development of the embryonic and extraembryonic circulatory systems in vascular cell adhesion molecule (VCAM-1) deficient mice. Development, 121 :489-503.
Love, A.M., and T.H. Vickers (1976) Placental agenesis, embryonal hydraemia, embryolethality and acute hypervitaminosis A in rats. J. Exp. Pathol., 57:525-541.
Luckett, W.P. (1971) The origin of the extraembryonic mesoderm in the early human and rhesus monkey embryos. Anat. Rec., 169:369- 370.
Luckett, W.P. (1978) Origin and differentiation of the yolk sac and extraembryonic mesoderm in premmite human and rhesus monkey embryos. Am. J. Anat., 152:59-98.
Mossman, H.W. (1937) Comparative morphogenesis of the fetal mem- branes and accessory uterine structures. Contrib. Embryol., 26: 130-301.
Nelson, O.E. (1953) Comparative Embryology of the Vertebrates. New York McGraw-Hill.
O’Rahilly, R. (1973) Developmental Stages in Human Embryos. Part A Embryos of the First Three Weeks (Stages 1 to 9). Carnegie Inst. Wash. Pub. 631, 167 pp.
Schramm, V.L., and D.C. Baker (1985) Spontaneous epimerization of (S)-deoxycoformycin and interaction of (R)-deoxycoformycin, (S)- deoxycoformycin and 8-ketodeoxycoformycin with adenosine deam- inase. Biochemistry, 24:641-646.
Snell, G.D., and L.C. Stevens (1966) Early embryology. In: The Biol- ogy of the Laboratory Mouse, 2nd ed. E.L. Green, ed. McGraw-Hill, New York, pp. 205-245.
327-338.
I . Streeter, G.L.-(1942) Developmental horizons in human embryos. De-

ALLANTOIS IS A PRIMARY SITE OF dCF TOXICITY 373
scription of age groups XI, 13 to 20 somites, and age group XII, 21 to 29 somitea. Contrib. Embryol. Carnegie Inst., 30:211-223.
Theiler, K. (1972) The House Mouse. Development and Normal Stages from Fertilization to 4 Weeks of Age. New York Springer- Verlag.
Thom, D.H., L.M. Nelson, and T.L. Vaughan (1992) Spontaneous abortion and subsequent adverse birth outcomes. Am. J. Obstet. Gynecol., l66:111-132.
Van MaeleFabry, G., F. Delhaise, and J.J. Picard (1990) Morphogen- esis and quantification of the development of post-implantation mouse embryos. Toxicol. In Vitro, 4r149-156.
Van Maele-Fabry, G., F. Delhaise, and J.J. Picard (1992) Evolution of
the developmental scores of sixteen morphological features in mouse embryos displaying 0 to 30 somites. Int. J. Dev. Biol., 36:
Vickers, T.H. (1985) Embryolethality in rats caused by retinoic acid. Teratology, 31r19-33.
Viscarello, R.R., D.D. Ferguson, J. Nores, and J.C. Hobbins (1992) Limb-body wall complex associated with cocaine abuse: Further evidence of cocaine’s teratogenicity. Obstet. Gynecol., 80523-526.
Yang, J.T., H. Rayburn, and R.O. Hynes (1995) Cell adhesion events mediated by a4 integrins are essential in placental and cardiac development. Development, 121.549-560.
161-167.





























