Embed Size (px)
Citation preview

Developmental C ell, Vol. 8, 689–701, May, 2005, C opyright ©2005 by Elsevier Inc. D OI 10.1016/j.devcel.2005.02.011
FGF Signal Interpretation Is Directedby Sprouty and Spred Proteinsduring Mesoderm FormationJeremy M. Sivak,1,3 Lars F. Petersen,1,2,3 F G FR during this process is the ras/MAPK pathway
(Schlessinger, 2000). Inhibition of any component ofand Enrique Amaya1,2,*this pathway subsequently blocks the expression of1The Wellcome Trust/ C ancer Research UK Gurdonmesodermal markers (Gupta and Mayer, 1998; Mac-InstituteNicol et al., 1993; Tang et al., 1995; Whitman and Mel-University of C ambridgeton, 1992). F G F signaling has also been shown to di-Tennis C ourt Road C ambridgerectly affect morphogenetic cell movements duringC B2 1Q Ngastrulation (Ciruna et al., 1997; Yang et al., 2002). WeUnited Kingdomhave shown that this effect occurs through the F G FR2 Department of Zoologyvia a mechanism that is distinct from the ras/MAPKUniversity of C ambridgepathway (Nutt et al., 2001). Therefore, F G FR signalingDowning Site, C ambridgeis used both for maintaining mesoderm fate and for reg-United Kingdomulating morphogenesis. However, how does the embryointerpret the signals correctly so that mesoderm main-tenance and morphogenesis occur in a coordinatedSummaryfashion?
We have previously reported that the receptor tyro-Vertebrate gastrulation requires coordination of meso-sine kinase (RTK) inhibitor protein, Xsprouty2, inhibitsderm specification with morphogenetic movements.F G FR- mediated morphogenesis but leaves mesodermWhile both of these processes require FGF signaling,formation intact (Nutt et al., 2001). The Sprouty familyit is not known how mesoderm specification and cellwas first discovered from a Drosophila mutation thatmovements are coordinated during gastrulation. Themimicked the effects of overactive F G F signaling (Ha-related Sprouty and Spred protein families are recentlycohen et al., 1998). A family of conserved Sproutydiscovered regulators of receptor tyrosine kinase sig-genes have since been identified that act as intracellu-naling. We identified two genes for each family in Xeno-lar inhibitors of RTK signaling, with homologs found inpus tropicalis: Xtsprouty1, Xtsprouty2, Xtspred1, andmice, humans, zebrafish, and Xenopus (Christofori,Xtspred2. In gain- and loss-of-function experiments we2003; Guy et al., 2003). Sprouty family members shareshow that XtSprouty and XtSpred proteins modulatea conserved cysteine-rich carboxy-terminal domaindifferent signaling pathways downstream of the FGF(Spry) and divergent amino termini (Guy et al., 2003).receptor (FGFR), and consequently different develop-Recently, a Sprouty-related family, the Spreds, has alsomental processes. Notably, XtSproutys inhibit mor-been described; they have been shown to inhibit RTKphogenesis and Ca2+ and PKC! signaling, leaving MAPKsignaling as well. Spreds share the Spry domain butactivation and mesoderm specification intact. In con-contain an N-terminal Enabled/VASP homology 1 do-trast, XtSpreds inhibit MAPK activation and meso-main (EVH1), and most have a binding sequence forderm specification, with little effect on Ca2+ or PKC!the oncogenic RTK c-kit (Kato et al., 2003; Wakioka etsignaling. These differences, combined with the timingal., 2001).of their developmental expression, suggest a mecha-
The biological roles of Sprouty and Spred proteinsnism to switch FGFR signal interpretation to coordi-remain unclear due to results suggesting they have anate mesoderm formation and cell movements dur- variety of effects on RTK signaling. The majority of at-ing gastrulation. tention has focused on inhibition of MAPK activationby Sprouty proteins downstream of RTK signalingIntroduction (C asci et al., 1999; Hanafusa et al., 2002). However,Sprouty family members have been shown to have vari-
The formation and patterning of mesoderm represents able effects on MAPK activation and even enhancea critical stage of vertebrate development. During gas- MAPK signaling (Sasaki et al., 2001; Wong et al., 2002;trulation, mesodermal tissue must be coordinately in- Yusoff et al., 2002). We have shown that Xsprouty2 ef-duced and directed through a series of morphogenetic fectively inhibits C a2+ signaling but has little effect onmovements inside the embryo to form derivatives, such MAPK activation in vivo (Nutt et al., 2001). The Spredsas muscle, notochord, and blood. C ompletion of these also inhibit MAPK activity and have been reported toevents is dependent on appropriate cellular responses block activation of Raf (Kato et al., 2003; Wakioka etto signals from a relatively small number of growth al., 2001). Notably, mammalian Spreds have been ob-factors, including members of the fibroblast growth served to be more potent inhibitors of MAPK activityfactor family (F G Fs) (Sivak and Amaya, 2004). than Sproutys in vitro (Wakioka et al., 2001).
We have shown previously that disrupting F G F re- Here we have identified two members of each theceptor (F G FR) signaling inhibits mesoderm induction Sprouty and Spred families in the diploid frog Xenopusand maintenance (Amaya et al., 1991; Kroll and Amaya, tropicalis: Xtsprouty1 and Xtsprouty2 and Xtspred1 and1996). The prominent pathway downstream of the Xtspred2. Using both gain- and loss-of-function experi-
ments, we show that these two families of proteins helpmodulate F G F signal interpretation in the early embryo*C orrespondence: [email protected]
3 These authors contributed equally to this work. by inhibiting distinct downstream signal transduction
Developmental C ell690
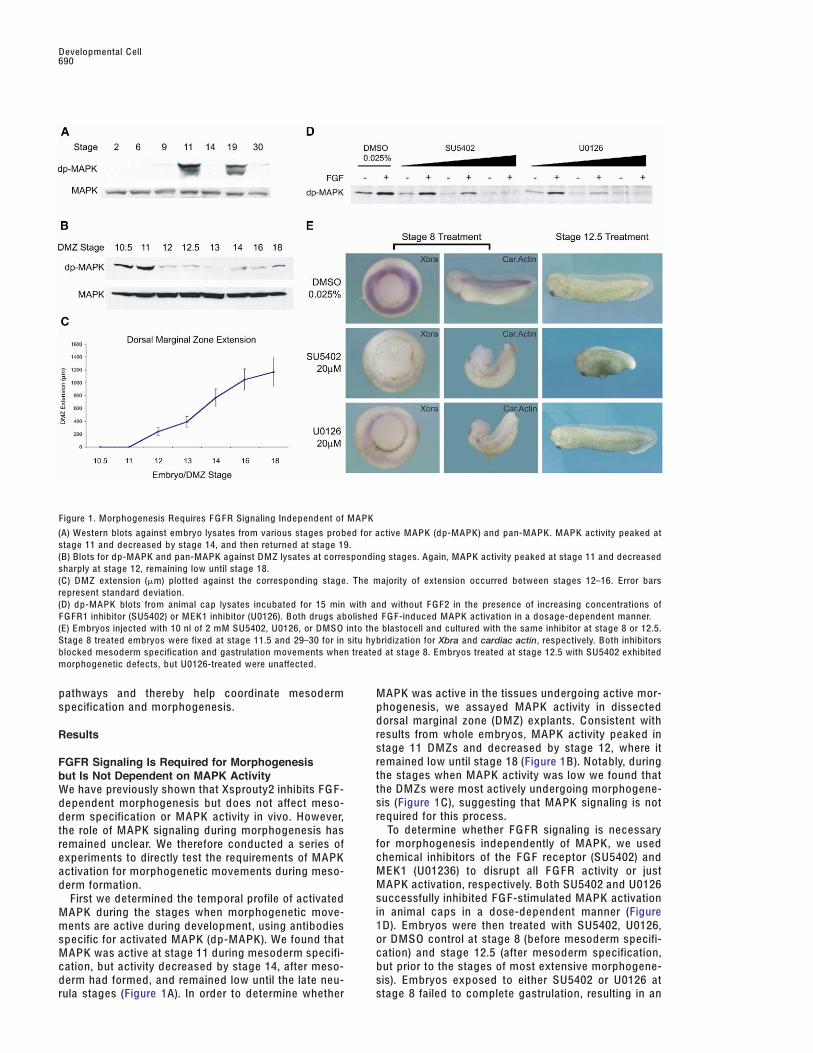
Figure 1. Morphogenesis Requires F G FR Signaling Independent of MAPK
(A) Western blots against embryo lysates from various stages probed for active MAPK (dp-MAPK) and pan-MAPK. MAPK activity peaked atstage 11 and decreased by stage 14, and then returned at stage 19.(B) Blots for dp-MAPK and pan-MAPK against DMZ lysates at corresponding stages. Again, MAPK activity peaked at stage 11 and decreasedsharply at stage 12, remaining low until stage 18.(C) DMZ extension (!m) plotted against the corresponding stage. The majority of extension occurred between stages 12–16. Error barsrepresent standard deviation.(D) dp-MAPK blots from animal cap lysates incubated for 15 min with and without F G F2 in the presence of increasing concentrations ofF G FR1 inhibitor (SU5402) or MEK1 inhibitor (U0126). Both drugs abolished F G F-induced MAPK activation in a dosage-dependent manner.(E) Embryos injected with 10 nl of 2 mM SU5402, U0126, or DMSO into the blastocell and cultured with the same inhibitor at stage 8 or 12.5.Stage 8 treated embryos were fixed at stage 11.5 and 29–30 for in situ hybridization for Xbra and cardiac actin, respectively. Both inhibitorsblocked mesoderm specification and gastrulation movements when treated at stage 8. Embryos treated at stage 12.5 with SU5402 exhibitedmorphogenetic defects, but U0126-treated were unaffected.
pathways and thereby help coordinate mesoderm MAPK was active in the tissues undergoing active mor-phogenesis, we assayed MAPK activity in dissectedspecification and morphogenesis.dorsal marginal zone (DMZ) explants. C onsistent withresults from whole embryos, MAPK activity peaked inResultsstage 11 DMZs and decreased by stage 12, where itremained low until stage 18 (Figure 1B). Notably, duringFGFR Signaling Is Required for Morphogenesisthe stages when MAPK activity was low we found thatbut Is Not Dependent on MAPK Activitythe DMZs were most actively undergoing morphogene-We have previously shown that Xsprouty2 inhibits F G F-sis (Figure 1C), suggesting that MAPK signaling is notdependent morphogenesis but does not affect meso-required for this process.derm specification or MAPK activity in vivo. However,
To determine whether F G FR signaling is necessarythe role of MAPK signaling during morphogenesis hasfor morphogenesis independently of MAPK, we usedremained unclear. We therefore conducted a series ofchemical inhibitors of the F G F receptor (SU5402) andexperiments to directly test the requirements of MAPKMEK1 (U01236) to disrupt all F G FR activity or justactivation for morphogenetic movements during meso-MAPK activation, respectively. Both SU5402 and U0126derm formation.successfully inhibited F G F-stimulated MAPK activationFirst we determined the temporal profile of activatedin animal caps in a dose-dependent manner (FigureMAPK during the stages when morphogenetic move-1D). Embryos were then treated with SU5402, U0126,ments are active during development, using antibodiesor DMSO control at stage 8 (before mesoderm specifi-specific for activated MAPK (dp-MAPK). We found thatcation) and stage 12.5 (after mesoderm specification,MAPK was active at stage 11 during mesoderm specifi-but prior to the stages of most extensive morphogene-cation, but activity decreased by stage 14, after meso-
derm had formed, and remained low until the late neu- sis). Embryos exposed to either SU5402 or U0126 atstage 8 failed to complete gastrulation, resulting in anrula stages (Figure 1A). In order to determine whether

F G F Signal Interpretation by Sprys/Sprds691
open blastopore phenotype with decreased expression weak or absent in the midbrain/hindbrain isthmus andforebrain and did not extend as far posteriorly in theof mesodermal markers Xbrachyury (Xbra) and cardiac
actin (Figure 1E) (percentage of open blastopore phe- tail bud.We next decided to determine whether the Xtsproutynotypes: 0% [n = 48] in DMSO; 76.6% [n = 47] in
SU5402; 84.4% [n = 45] in U0126)]. These results are and Xtspred genes are expressed at the same relativetimes. Since in situ hybridization data are not quantita-consistent with the effects of disrupting the F G FR or
ras/MAPK pathway during early stages of develop- tive, we decided to answer this question using quanti-tative real-time RT-P C R. We found that, although thement. In comparison, embryos treated with the F G FR
inhibitor (SU5402) at stage 12.5 showed morphogenetic Xtsprouty and Xtspred genes share similar spatial pat-terns, they have important temporal differences. Thedefects resulting in a shortened anterior-posterior (A-P)
axis, whereas embryos treated with the MAPK inhibitor relative expression levels by real-time RT-P C R ofXtspry1 and Xtspry2 was higher during early gastrula(U0126) developed normally (Figure 1E) (percentage
shorter than 4.5 mm at stage 29–30: 0% [n = 40] in stages and then decreased, while Xtsprd1 and Xtsprd2remained low during the early gastrula stages but in-DMSO; 92.3% [n = 29] in SU5402; 2.5% [n = 40] in
U0126). Taken together, these results show that F G FR creased significantly from the end of gastrulationthrough neurula stages (Figure 2C).signaling is required for both mesoderm specification
and morphogenesis, but only mesoderm specificationdepends on MAPK activity, suggesting that morpho- XtSprouty and XtSpred Gain-of-Functiongenesis involves an alternate downstream pathway. Experiments Cause Distinct Phenotypes
To begin to address the function of these genes, weinjected in vitro transcribed mRNAs corresponding toXtsproutys and Xtspreds Are Related Families ofthe Xtsprouty and Xtspred genes into the dorsal mar-FGFR Regulatory Genes that Share Overlappingginal zones of X. laevis embryos at the two-cell stage.Expression Patterns, but at Slightly Different TimesEmbryos injected with Xtsproutys developed a short-B ased on our previous work, we hypothesized thatened A-P axis, compared to controls injected with aSprouty family members may be involved in regulatingnonfunctional F G F receptor (HAV\) or uninjected em-F G FR signal interpretation during development. Webryos (Figures 3A and 3B). Interestingly, this phenotypeisolated two X. tropicalis sprouty genes, Xtsprouty1is similar to that caused by treatment with an F G FR(Xtspry1) and Xtsprouty2 (Xtspry2), and two related spredinhibitor at stage 12.5 (Figure 1E). In comparison, em-genes, Xtspred1 (Xtsprd1) and Xtspred2 (Xtsprd2). Asbryos injected with Xtspreds developed an open blas-with their mammalian counterparts, all the identifiedtopore phenotype reminiscent of the effects of earlyXtSprouty and XtSpred proteins share a conserved cys-F G FR inhibition (Figure 1E). These results promptedteine-rich C -terminal Spry domain. The two Sproutyfurther investigations to determine if the differences be-proteins contain an uncharacterized N-terminal domaintween the Xtsproutys and Xtspreds were variations inwhose function remains obscure (Guy et al., 2003),severity of the same phenotype or caused by distinctwhile the two Spreds contain an N-terminal EVH1 do-underlying molecular mechanisms.main and c-Kit binding domain (K B D) (Harmer et al.,
2005; Wakioka et al., 2001) (Figure 2A). All proteins con-tain a high degree of identity to their human counter- XtSpreds Inhibit Mesoderm Specification
but XtSproutys Do Notparts: 72.9% for Sprouty1, 72.7% for Sprouty2, 60.1%for Spred1, and 69.9% for Spred2. Early disruption of F G FR function blocks specification
of mesoderm as well as failure to complete gastrula-The Xtsprouty and Xtspred genes are all expressedin broadly similar patterns to X. laevis sprouty2 and a tion. This effect is due to interruption of the MAPK sig-
naling cascade controlling transcriptional activation ofnumber of F G Fs, including XF G F8, Xe F G F, and F G F-9(Christen and Slack, 1997; Isaacs et al., 1994; Nutt et al., mesodermal genes, such as the T-box transcription
factor Xbrachyury (Xbra) (Sivak and Amaya, 2004). We2001; Song and Slack, 1996). During gastrula stages, ex-pression was first localized to the dorsal marginal zone, tested the effects of Xtsprouty and Xtspred overexpres-
sion on mesoderm specification and MAPK activity. Abut later expanded laterally and ventrally. Although allfour genes displayed this pattern, Xtspry1 and Xtspry2 series of in situ hybridizations was carried out for Xbra
on mid-gastrula embryos that had been coinjected withwere detected earlier and expanded their expressionpattern more quickly than Xtsprd1 and Xtsprd2 (Figure either Xtsprouty or Xtspred, and β-gal RNAs into one
blastomere at the two-cell stage. C ontrol-injected em-2B). During neurula stages, expression became con-fined anteriorly and posteriorly and expanded to vary- bryos showed a characteristic ring of Xbra expression
in the marginal zone around the blastopore. Injectionsing degrees along the neural tube. G enerally, stainingfor Xtsproutys became weaker and more localized, of either Xtsprouty had little effect on Xbra; however,
both Xtspreds completely blocked Xbra expression onwhile Xtspreds became stronger and more broad. Attail bud stages, staining became localized to the de- the injected side (Figure 3C).
We then assayed the effects of XtSprouty andveloping branchial arches, forebrain, otic vesicle, mid-brain/hindbrain isthmus, and tail bud. Xtspry2 staining XtSpred proteins on MAPK activity, using the dp-MAPK
antibody. As shown previously, during gastrulationwas absent from the third branchial arch; however, thisfeature was stained for Xtspry1, Xtsprd1, and Xtsprd2 there is a sharp F G F-dependent increase in the amount
of activated MAPK, concomitant with mesoderm speci-mRNAs. C omparatively, Xtspred gene expression over-lapped considerably with the Xtsprouty genes, with the fication (Figure 1A). When 1.0 ng RNAs was injected
into both blastomeres, neither XtSprouty had an effectexception that staining for the Xtspred genes was either

Developmental C ell692
Figure 2. Xtsproutys and Xtspreds Are Expressed in Similar Patterns, but at Slightly Different Times
(A) C artoon depiction of domain structures for each protein. Spry, conserved Sprouty domain; EVH-1, VASP homology domain; K B D, c-Kitbinding domain.(B) In situ hybridizations for each gene. At gastrula stages (stage 10.5), Xtsproutys showed more extensive staining around the blastoporethan Xtspreds (arrow). Neurula stages showed increased staining of Xtspreds and decreased Xtsproutys. Tail bud stages showed similarstaining in branchial arches (br), otic vesicle (ov), and tail bud (tb), but Xtspred staining was absent in mid-brain/hind-brain isthmus (mhb)and forebrain (fb).(C) Quantitative real-time RT-P C R analysis of each gene’s expression over time. Relative expression levels showed Xtsproutys peak beforestage 12.5 and then diminish, while Xtspreds levels increased after stage 12.5.
on MAPK activity. However, both XtSpred proteins This suggests that the XtSpred proteins are much moreeffective inhibitors of MAPK signaling than the XtSproutyblocked MAPK signaling as efficiently as the dominant-
negative F G F receptor (XF D) (Figure 3D). Since Sprouty proteins in vivo. Others have similarly reported thatmammalian Spred proteins are more effective MAPK in-proteins have been studied as inhibitors of the MAPK
signaling pathway in vitro, we tested the possibility that hibitors than the Sprouty proteins (Wakioka et al.,2001).the XtSproutys might be more effective at inhibiting
MAPK activation at higher concentrations. A dosage We also tested the effect of F G F-induced MAPK acti-vation in animal caps. C ompared to the results in Figureseries with the most effective family member, Xtspry2,
was able to slightly reduce MAPK activation at the 3D, which tested steady state MAPK activity in wholeembryos, this experiment directly tested the F G F-highest amount tested (2 ng), but Xtsprd1 completely
abolished MAPK activation even at 1 ng (Figure 3E). dependent activation of MAPK in animal caps after 15
F G F Signal Interpretation by Sprys/Sprds693
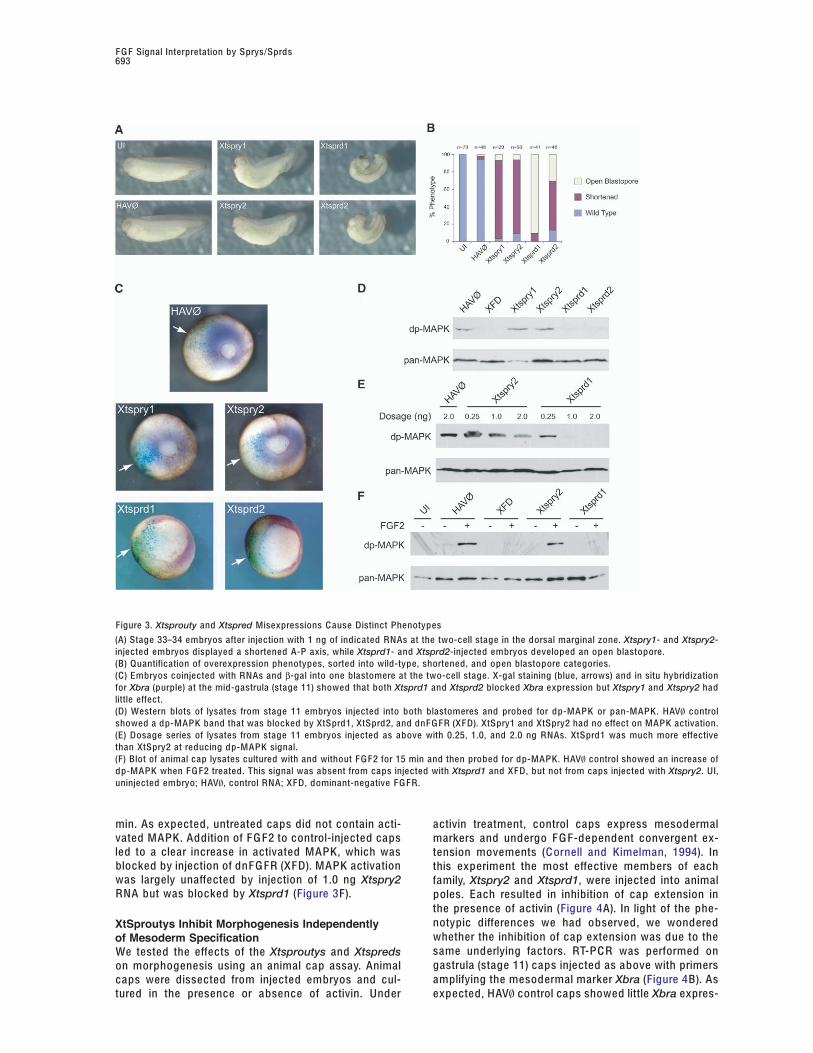
Figure 3. Xtsprouty and Xtspred Misexpressions C ause Distinct Phenotypes
(A) Stage 33–34 embryos after injection with 1 ng of indicated RNAs at the two-cell stage in the dorsal marginal zone. Xtspry1- and Xtspry2-injected embryos displayed a shortened A-P axis, while Xtsprd1- and Xtsprd2-injected embryos developed an open blastopore.(B) Quantification of overexpression phenotypes, sorted into wild-type, shortened, and open blastopore categories.(C) Embryos coinjected with RNAs and β-gal into one blastomere at the two-cell stage. X-gal staining (blue, arrows) and in situ hybridizationfor Xbra (purple) at the mid-gastrula (stage 11) showed that both Xtsprd1 and Xtsprd2 blocked Xbra expression but Xtspry1 and Xtspry2 hadlittle effect.(D) Western blots of lysates from stage 11 embryos injected into both blastomeres and probed for dp-MAPK or pan-MAPK. HAV\ controlshowed a dp-MAPK band that was blocked by XtSprd1, XtSprd2, and dnF G FR (XF D). XtSpry1 and XtSpry2 had no effect on MAPK activation.(E) Dosage series of lysates from stage 11 embryos injected as above with 0.25, 1.0, and 2.0 ng RNAs. XtSprd1 was much more effectivethan XtSpry2 at reducing dp-MAPK signal.(F) Blot of animal cap lysates cultured with and without F G F2 for 15 min and then probed for dp-MAPK. HAV\ control showed an increase ofdp-MAPK when F G F2 treated. This signal was absent from caps injected with Xtsprd1 and XF D, but not from caps injected with Xtspry2. UI,uninjected embryo; HAV\, control RNA; XF D, dominant-negative F G FR.
min. As expected, untreated caps did not contain acti- activin treatment, control caps express mesodermalvated MAPK. Addition of F G F2 to control-injected caps markers and undergo F G F-dependent convergent ex-led to a clear increase in activated MAPK, which was tension movements (C ornell and Kimelman, 1994). Inblocked by injection of dnF G FR (XF D). MAPK activation this experiment the most effective members of eachwas largely unaffected by injection of 1.0 ng Xtspry2 family, Xtspry2 and Xtsprd1, were injected into animalRNA but was blocked by Xtsprd1 (Figure 3F). poles. Each resulted in inhibition of cap extension in
the presence of activin (Figure 4A). In light of the phe-notypic differences we had observed, we wonderedXtSproutys Inhibit Morphogenesis Independentlywhether the inhibition of cap extension was due to theof Mesoderm Specificationsame underlying factors. RT-P C R was performed onWe tested the effects of the Xtsproutys and Xtspredsgastrula (stage 11) caps injected as above with primerson morphogenesis using an animal cap assay. Animalamplifying the mesodermal marker Xbra (Figure 4B). Ascaps were dissected from injected embryos and cul-
tured in the presence or absence of activin. Under expected, HAV\ control caps showed little Xbra expres-

Developmental C ell694
Figure 4. XtSproutys Inhibit Morphogenesis and F G F-Induced C a2+ and PK Cδ Signaling
(A) Animal caps cultured with and without activin until mid-neurula stages. C aps injected with HAV\ control extended when treated withactivin, but Xtspry2- or Xtsprd1-injected caps did not.(B) RT-P C R results from stage 11 animal caps injected as above. Activin treatment increased Xbra expression in HAVØ controls. Xtspry2increased Xbra expression in untreated caps, but had no effect after activin treatment. Xtsprd1 inhibited Xbra expression with activin.(C) Oocyte assay of C a2+ signaling. Release of 45C a2+ from oocytes coinjected with constitutively active F G FR (CIXR) and the indicated RNAswas counted after 10 min. Addition of CIXR increased 45C a2+ release 5-fold (HAV\). C oexpression of dominant-negative CIXR (CIXD), Xtspry1,Xtspry2, or X. laevis Sprouty2 (Xspry2) effectively blocked this increase. Xtsprd1 had significantly less effect, and Xtsprd2 and Xtsef had noeffect. Error bars represent standard deviation.(D) C onfocal images of PK Cδ-G F P expressed in animal caps and cultured for 10 min in the presence of either 1 M PMA, 100 ng/ml F G F2, orcoexpressed XtSpry2 or XtSprd1 as indicated. PK Cδ was activated by both PMA and F G F2 and localized to the membrane. XtSpry2 inhibitedF G F-induced localization of PK Cδ but XtSprd1 did not.
sion unless treated with activin to induce mesoderm. in the absence of activin. In comparison, injection ofXtsprd1 inhibited the induction of Xbra after activinXtspry2 had no effect on Xbra induction with activin,
and surprisingly caused an increase in Xbra expression treatment. Therefore, morphogenetic extension was

F G F Signal Interpretation by Sprys/Sprds695
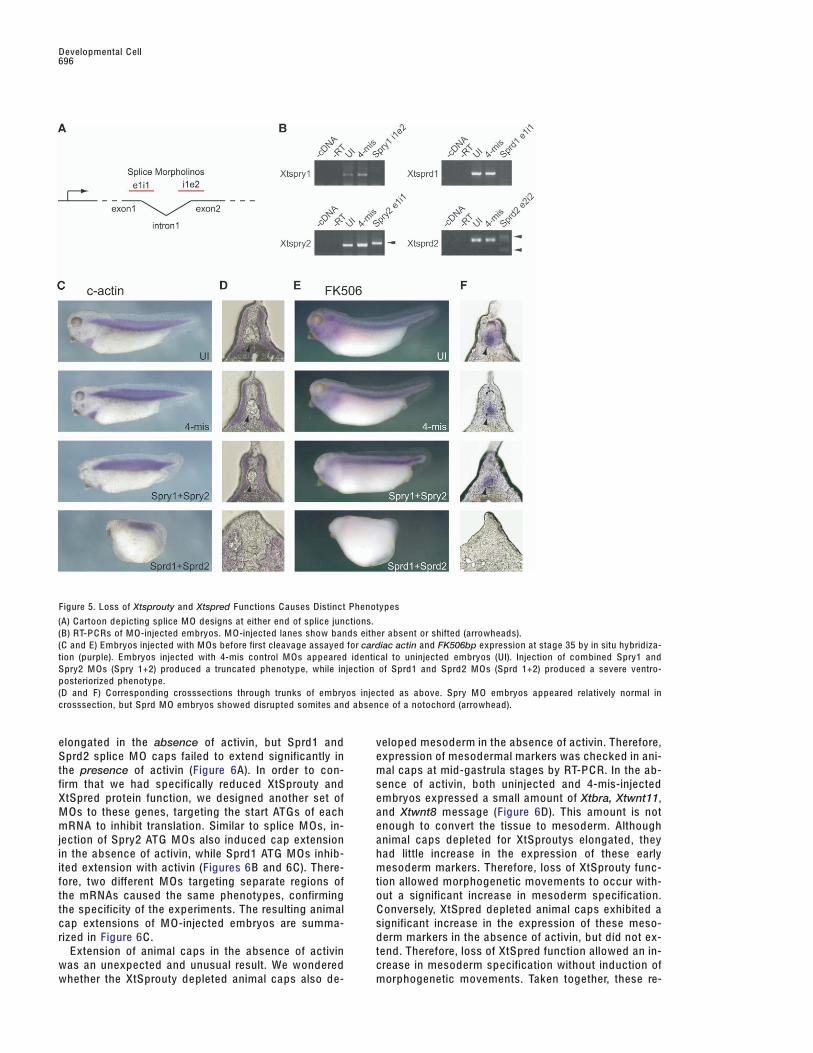
blocked in Xtspry2- injected caps even though they still Loss of XtSprouty and XtSpred Functions CausesDistinct Phenotypescontained mesoderm, while the Xtsprd1-injected caps
did not extend, but also lacked mesoderm. Loss-of-function experiments were conducted for Xt-Sprouty and XtSpred proteins using antisense morpho-lino oligonucleotides (M Os). We have previously usedXtSproutys Are More Effective Inhibitors of Ca2+this technique to effectively abrogate RNA translationSignaling than XtSpredsand/or splicing by injecting M Os directly into early em-The MAPK cascade is only one of several signalingbryos (Kenwrick et al., 2004). M O sequences were de-pathways that can be induced following F G FR activa-signed to specifically target splice junctions for eachtion. Another prominent downstream pathway actsXtsprouty and Xtspred pre-mRNA. A cartoon depictingthrough phospholipase Cγ (PLCγ). PLCγ hydrolyzesthe design and targeting of splice morpholinos isphosphatidylinositol into diacylglycerol (DA G) and ino-shown in Figure 5A. Each M O sequence was desig-sitol trisphosphate (IP3), which activates the IP3 recep-nated by its target location (e.g., e1i1 for the exon1-tor in the endoplasmic reticulum, causing an efflux ofintron1 boundary, i1e2 for the intron1-exon2 boundary,C a2+ into the cytoplasm (Schlessinger, 2000). Localizedetc.). We confirmed the effectiveness of the morpho-C a2+ signaling has been linked to convergent extensionlinos by assaying for splicing interference by perform-movements during Xenopus gastrulation (Wallingford eting RT-P C R using primers across each splice junction.al., 2001). We have previously reported that X. laevisAffected transcripts either amplified at different sizesSprouty2 inhibits F G F-induced C a2+ mobilization in anor failed to amplify (Figure 5B).efflux assay using Xenopus oocytes (Nutt et al., 2001).
X. tropicalis embryos were used for these experi-Using the same assay we compared the effect of thements as their diploid genome allows for much simplerXtSprouty and XtSpred proteins on F G FR-induced C a2+
loss-of-function interpretation than the allotetraploid X.release. Both XtSpry1 and XtSpry2 were as effective aslaevis. In each experiment, M Os were injected into zy-X. laevis Spry2 in blocking C a2+ release (Figure 4C). Ingotes before first cleavage. As a control, a M O contain-contrast, XtSprd1 only partially inhibited C a2+ releaseing four mismatches to the Xtspry2 sequence (4-mis)and XtSprd2 was ineffective. A negative control waswas used. Injection of M Os targeted to the Xtsproutyprovided by XtSef, an inhibitor of F G FR signaling thatand Xtspred genes developed markedly different phe-targets the Ras/MAPK pathway and bears no homologynotypes (Figures 5C and 5E). In these experiments,to the Sprouty family (Furthauer et al., 2002). These re-combinations of splice M Os were used in order to re-sults provide additional evidence that XtSprouty andduce the effects of functional redundancy betweenXtSpred proteins target different signaling pathwaysfamily members. A combination of Xtspry1 and Xtspry2downstream of F G FR activation.M Os (Spry1 i1e2 and Spry2 e1i1) caused anterior-pos-terior truncations. These results were similar to theFGF-Induced PKC! Localization Is Inhibitedgain-of-function effect (Figure 3A), which might be ex-by XtSproutys but Not by XtSpredspected if coordination of morphogenesis was dis-Another important secondary messenger produced byrupted. In contrast, M Os targeting the Xtspred genesPLCγ in addition to IP3 is DA G. The most prominent(using a combination of Sprd1 e1i1 and Sprd2 e2i2intracellular targets of DA G belong to the protein kinaseM Os) produced a more severe phenotype, with em-C (PK C) family. Kinoshita et al. (2003) recently demon-bryos appearing ventro-posteriorized (Figures 5C andstrated that upon activation, PKCδ, a member of the5E). This effect is reminiscent of that reported for em-novel (nPK C) subfamily, translocates to the cell mem-bryos with overactive F G FR signaling (Isaacs et al.,brane where it is an essential component of the nonca-1994). Xtsprd M O-injected embryos had diffuse andnonical Wnt, or planar cell polarity, signaling pathway.weak cardiac actin staining and had either absent orInhibition of PKCδ disrupts convergent extension move-undifferentiated notochords (Figures 5C and 5D). Simi-ments during gastrulation, but does not block the expres-larly, Xtsprd M O embryos showed no staining for thesion of mesodermal markers (Kinoshita et al., 2003).notochord marker FK506 binding protein (Spokony andGiven the similarities of these results to our observa-Saint-Jeannet, 2000) (Figures 5E and 5F). In contrast,tions with XtSproutys, and that nPK C proteins relyXtspry M O-injected embryos stained normally forsolely on DA G for activation (Gschwendt, 1999), wecardiac actin expression and the notochord was clearlytested whether F G FR signaling can activate PK Cδ.visible (Figures 5C–5F).Membrane localization of G F P-tagged PK Cδ was ob-
served in animal caps using confocal microscopy (Fig-ure 4D). PK Cδ-G F P signal was diffuse in untreated XtSprouty Depletion Causes Animal Caps
to Undergo Morphogenetic Extension, whilecaps, but addition of phorbol 12-myristate 13-acetate(PMA), a DA G analog, caused its translocation to cell XtSpred Depletion Inhibits Extension
Animal caps isolated from MO-injected embryos were in-membranes. Interestingly, addition of FG F2 also causedPK Cδ-G F P to localize to the membrane. This localiza- cubated with and without activin (10 ng/ml) and assayed
for extension. Both uninjected and 4-mis-injected capstion was inhibited if XtSpry2 was coexpressed withPK Cδ-G F P, but not when coexpressed with XtSprd1 remained as rounded balls when left untreated but elon-
gated in the presence of activin (Figure 6A). However,(Figure 4D). These results provide additional evidencethat XtSproutys target a distinct PLCγ-mediated signal- animal caps isolated from Xtsprouty and Xtspred de-
pleted embryos had strikingly different outcomes. C apsing pathway compared with XtSpreds downstream ofthe F G FR. injected with combined Spry1 and Spry2 splice M Os
Developmental C ell696
Figure 5. Loss of Xtsprouty and Xtspred Functions C auses Distinct Phenotypes
(A) C artoon depicting splice M O designs at either end of splice junctions.(B) RT-P C Rs of M O-injected embryos. M O-injected lanes show bands either absent or shifted (arrowheads).(C and E) Embryos injected with M Os before first cleavage assayed for cardiac actin and FK506bp expression at stage 35 by in situ hybridiza-tion (purple). Embryos injected with 4-mis control M Os appeared identical to uninjected embryos (UI). Injection of combined Spry1 andSpry2 M Os (Spry 1+2) produced a truncated phenotype, while injection of Sprd1 and Sprd2 M Os (Sprd 1+2) produced a severe ventro-posteriorized phenotype.(D and F) C orresponding crosssections through trunks of embryos injected as above. Spry M O embryos appeared relatively normal incrosssection, but Sprd M O embryos showed disrupted somites and absence of a notochord (arrowhead).
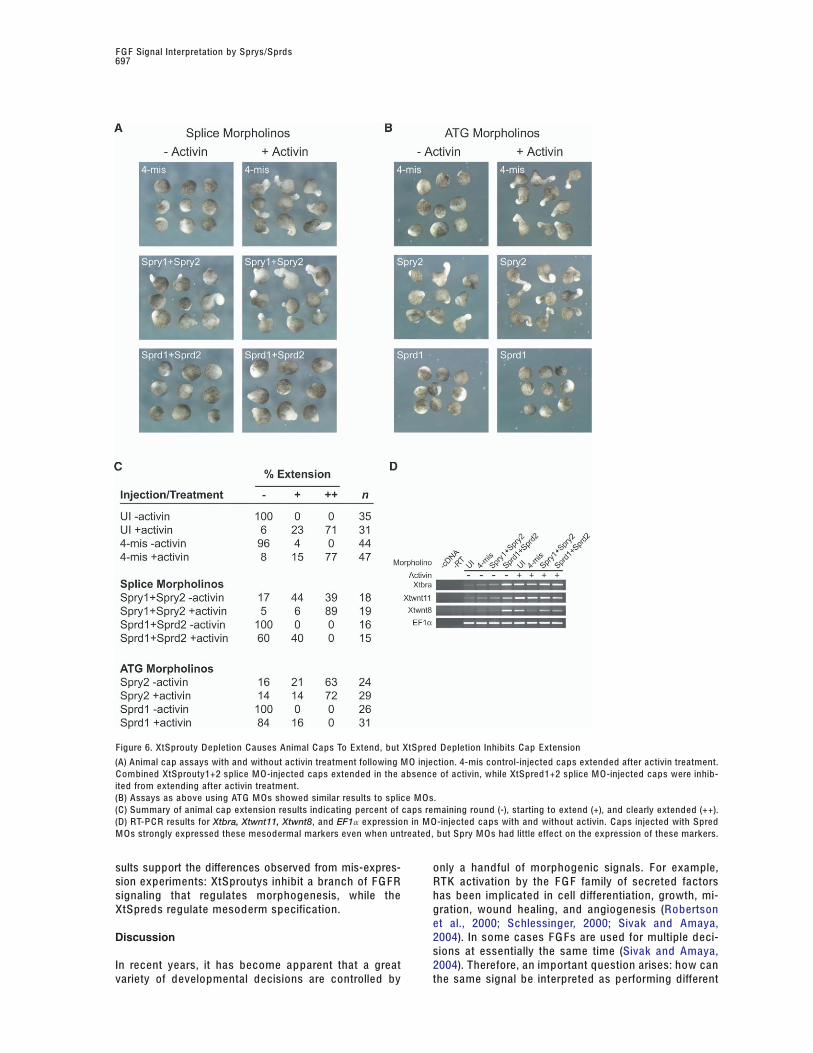
elongated in the absence of activin, but Sprd1 and veloped mesoderm in the absence of activin. Therefore,expression of mesodermal markers was checked in ani-Sprd2 splice M O caps failed to extend significantly in
the presence of activin (Figure 6A). In order to con- mal caps at mid-gastrula stages by RT-P C R. In the ab-sence of activin, both uninjected and 4-mis-injectedfirm that we had specifically reduced XtSprouty and
XtSpred protein function, we designed another set of embryos expressed a small amount of Xtbra, Xtwnt11,and Xtwnt8 message (Figure 6D). This amount is notM Os to these genes, targeting the start ATGs of each
mRNA to inhibit translation. Similar to splice M Os, in- enough to convert the tissue to mesoderm. Althoughanimal caps depleted for XtSproutys elongated, theyjection of Spry2 ATG M Os also induced cap extension
in the absence of activin, while Sprd1 ATG M Os inhib- had little increase in the expression of these earlymesoderm markers. Therefore, loss of XtSprouty func-ited extension with activin (Figures 6B and 6C). There-
fore, two different M Os targeting separate regions of tion allowed morphogenetic movements to occur with-out a significant increase in mesoderm specification.the mRNAs caused the same phenotypes, confirming
the specificity of the experiments. The resulting animal C onversely, XtSpred depleted animal caps exhibited asignificant increase in the expression of these meso-cap extensions of M O-injected embryos are summa-
rized in Figure 6C . derm markers in the absence of activin, but did not ex-tend. Therefore, loss of XtSpred function allowed an in-Extension of animal caps in the absence of activin
was an unexpected and unusual result. We wondered crease in mesoderm specification without induction ofmorphogenetic movements. Taken together, these re-whether the XtSprouty depleted animal caps also de-
F G F Signal Interpretation by Sprys/Sprds697
Figure 6. XtSprouty Depletion C auses Animal C aps To Extend, but XtSpred Depletion Inhibits C ap Extension
(A) Animal cap assays with and without activin treatment following M O injection. 4-mis control-injected caps extended after activin treatment.C ombined XtSprouty1+2 splice M O-injected caps extended in the absence of activin, while XtSpred1+2 splice M O-injected caps were inhib-ited from extending after activin treatment.(B) Assays as above using ATG M Os showed similar results to splice M Os.(C) Summary of animal cap extension results indicating percent of caps remaining round (-), starting to extend (+), and clearly extended (++).(D) RT-P C R results for Xtbra, Xtwnt11, Xtwnt8, and EF1a expression in M O-injected caps with and without activin. C aps injected with SpredM Os strongly expressed these mesodermal markers even when untreated, but Spry M Os had little effect on the expression of these markers.
sults support the differences observed from mis-expres- only a handful of morphogenic signals. For example,RTK activation by the F G F family of secreted factorssion experiments: XtSproutys inhibit a branch of F G FR
signaling that regulates morphogenesis, while the has been implicated in cell differentiation, growth, mi-gration, wound healing, and angiogenesis (RobertsonXtSpreds regulate mesoderm specification.et al., 2000; Schlessinger, 2000; Sivak and Amaya,2004). In some cases F G Fs are used for multiple deci-Discussionsions at essentially the same time (Sivak and Amaya,2004). Therefore, an important question arises: how canIn recent years, it has become apparent that a great
variety of developmental decisions are controlled by the same signal be interpreted as performing different

Developmental C ell698
Figure 7. Model of Sprouty and Spred Regu-lated Switching of F G FR Signal Pathways
(A) Xtsproutys are expressed first, resulting ininhibition of the PLCγ/morphogenesis pathwaydownstream of the F GFR while the MAPK/mesoderm specification pathway proceeds.(B) As mesoderm specification finishes,Xtspred expression increases, resulting in in-hibition of the mesoderm specification path-way. At this point expression of Xtsproutysdiminishes, allowing the morphogenesispathway to proceed.
functions, often within the same cells? The related major outcomes of PLCγ activation. Importantly, PK Cδhas recently been demonstrated to be an essentialSprouty and Spred families are recently discovered pro-component of the wnt/planar cell polarity pathway in-teins that function to directly regulate RTK signalingvolved in regulating morphogenesis (Kinoshita et al.,downstream of receptor activation (Christofori, 2003;2003). C a2+ signaling has also been implicated in con-Guy et al., 2003). Here we have described divergenttrol of convergent extension movements without af-roles for Sproutys and Spreds in regulation of F G FRfecting cell fate (Wallingford et al., 2001). Therefore,signaling during vertebrate gastrulation: XtSprouty pro-these results provide evidence of a mechanism directlyteins inhibit morphogenesis, PK Cδ activation, and C a2+
linking F G FR signaling to cell migration/cell polaritysignaling, and XtSpreds inhibit mesoderm specificationpathways.and MAPK activation. In this manner XtSproutys and
In contrast, XtSpreds inhibited mesoderm differentia-XtSpreds can switch between activation of two F G FRtion through strong inhibition of MAPK activation andsignal pathways to coordinate distinct developmentalhad little effect on C a2+ signaling or PK Cδ activation.events.Embryos injected with Xtspred1 and Xtspred2 RNAsdid not develop mesoderm and displayed an openFunctional Divergence of Xtsproutyblastopore phenotype reminiscent of the effect ofand Xtspred ProteinsdnF G FR (Amaya et al., 1991). Likewise, XtSpred pro-Gain- and loss-of-function experiments revealed markedteins blocked MAPK activity and extension in animaldifferences between XtSprouty and XtSpred functionscaps. These results are consistent with studies show-in vivo. Xtsprouty misexpression resulted in shorteneding that mammalian Spreds inhibit Raf activation up-embryos and inhibited morphogenic movements but leftstream of MAPK (Kato et al., 2003; Wakioka et al.,mesoderm formation intact. FG FR-dependent MAPK ac-2001). Interestingly, the timing of expression of thetivation was only weakly affected, but PKCδ activationXtspred genes correlates with the loss of MAPK activityand Ca2+ signaling were strongly inhibited. Notably, Xbra in embryos, suggesting that the decrease in MAPK ac-
was induced in untreated animal caps overexpressing tivity is due to the expression of the Xtspred genes.Xtspry2, suggesting that in this context XtSprouty pro- Depletion of XtSpreds resulted in ventro-posteriorizedteins can actually enhance the mesoderm specification embryos that lacked a notochord, similar to the effectpathway. Conversely, XtSprouty depletion caused un- of constitutive F G FR signaling (Isaacs et al., 1994).treated animal caps to undergo morphogenetic exten- Loss of XtSpreds in animal caps inhibited elongationssions without an accompanying increase in mesoderm. with activin treatment. However, increased mesodermThis striking result is similar to the one observed when was detected in untreated Xtspred-depleted caps, indi-an activated version of protein-tyrosine phosphatase cating excessive mesoderm formation consistent withSHP-2 is missexpressed in animal caps (O’Reilly et al., XtSpreds acting as MAPK inhibitors.2000). Interestingly, this same activated version of SHP-2 Taken together, these results describe a molecularhas recently been shown to dephosphorylate and inac- branching of F G FR signaling during mesoderm forma-tivate Sprouty (Hanafusa et al., 2004). Therefore, it may tion and morphogenesis: a Sprouty-sensitive pathwaybe possible that untreated animal caps expressing acti- affecting PK Cδ activation and C a2+ signaling and medi-vated SHP-2 elongate because Sprouty is inactivated, ating morphogenesis, and a Spred-sensitive pathwaya question that is now being investigated. affecting MAPK activity and mesoderm specification.
In other model systems Sproutys have also consis- With the identification of these two regulatory pathwaystently been associated with morphogenesis and cell it will now be possible to dissect the various down-migration, processes not normally associated with stream elements involved in each branch. One impor-MAPK signaling (Lim et al., 2000; Yigzaw et al., 2001, tant aspect to address will be the functional roles con-2003). In particular, depletion of Sproutys has been re- tributed by the various XtSprouty and XtSpred proteinported to cause excessive morphogenesis in mice and domains in order to identify the elements regulatingflies (Hacohen et al., 1998; Tefft et al., 1999). The animal their specificity. As there is considerable divergencecap extensions we observed after XtSprouty depletion between Drosophila and vertebrate Sprouty sequences,is reminiscent of these other cases. We have shown Sprouty and Spred functions may have become spe-here that XtSproutys are efficient inhibitors of F G FR- cialized to accomplish specific signaling tasks during
vertebrate evolution.dependent C a2+ signaling and PK Cδ activation, two

F G F Signal Interpretation by Sprys/Sprds699
TG as112n24), Xtspred1 (Xtsprd1; TNeu018m17), and Xtspred2A Switching Mechanism for FGFR(Xtsprd2; TG as019j20). Full-length clones were purified from corre-Signal Interpretationsponding EST libraries (Gilchrist et al., 2004). In some cases extra-F G FR-dependent activation of the Ras/MAPK pathwayneous sequence was deleted to remove unwanted regulatory ele-
is necessary for specification of mesoderm in vivo (Si- ments. The Xtspry1 clone TG as112n24 was missing 86 bases ofvak and Amaya, 2004). However, we have demon- coding sequence at the 5# end, which was replaced by searching
the JGI X. tropicalis G enomic Database to identify the correspond-strated a role for F G FR signaling in the morphogeneticing sequence. G enB ank accession numbers are as follows:cell movements that follow as well. Several otherXtspry1, AY714335; Xtspry2, AY714336; Xtsprd1, AY714338;studies have also demonstrated a role of F G FR signal-Xtsprd2, AY714337). In situ hybridizations were performed accord-ing in morphogenesis during gastrulation (Ciruna anding to established protocols using antisense DIG-labeled probes.
Rossant, 2001; Nutt et al., 2001; Yang et al., 2002). No- Dominant-negative F G F receptor (XF D), nonfunctional F G F re-tably, we have shown here that although F G FR function ceptor (HAV\), constitutively active F G FR1 (p CIXR), and its corre-
sponding dominant-negative (p CIXD) have been previously de-is required, MAPK is not active during the majority ofscribed (Amaya et al., 1991; Nutt et al., 2001). Full-length X. laevismorphogenesis in the early embryo. Therefore a devel-PK Cδ1 was obtained from the I.M.A.G.E. C onsortium (Invitrogen/opmental switch to a different F G FR signaling pathwayResgen, 4969220) and subcloned into the StuI and XhoI sites of themust be made to mediate this function.p C SG F P3-TAA vector to generate an N-terminal PK Cδ-G F P fusion.
All four Xtsproutys and Xtspreds analyzed had largelyoverlapping expression patterns. However, we found RNA Synthesis and Embryo Microinjectionsvariations over time, which has important implications C apped mRNA was synthesized using the mMessage Machine kit
according to the manufacturer’s directions (Ambion). Embryos werefor the regulation of F G FR signaling. Expression ofcultured and injected with RNAs according to established protocol.Xtspry1 and 2 are relatively stronger during early gas-Developmental staging was assessed as described (Nieuwkooptrulation, peaking by stage 10.5, after which expressionand F aber, 1967). G enerally, 100 pg of nuclear localized β-galacto-levels are reduced. In contrast, Xtsprd1 and 2 are ex- sidase (pnucβ-gal) or G F P RNA were coinjected as a lineage tracer.
pressed at lower levels during early gastrulation but For animal cap experiments, embryos were injected at animal polesrise after stage 12, when morphogenesis predominates. and cap tissue was excised at stage 8. C aps were then incubated
in either 0.4× MMR (for extension experiments) or 1× MMR (forWhen these temporal differences are compared withMAPK assay). Human recombinant activin A (R&D Systems) wasthe signaling pathways targeted by each gene family, aadded at 10 ng/ml, and F G F2 (a gift of Dr. Harry Isaacs) was addedmodel of F G FR signal switching can be proposed (Fig-at 100 ng/ml.ure 7).
During the early stages of gastrulation, F G FR signal- FGFR and MAPK Inhibitor Treatmentsing is necessary to maintain specification of mesoder- Embryos were cultured to stage 8 or 12.5 and injected into themal tissues. At this time, Xtsproutys are strongly ex- blastocoel with 10 nl of a 2 mM DMSO stock of either SU5402
(C albiochem), an F G FR1 kinase activity inhibitor, or U0126 (C albio-pressed and inhibit the PLCγ pathway regulatingchem), a MEK1 inhibitor, and cultured in 0.1× MMR with 20 !M ofmorphogenetic movements. C onsequently, the pre-the corresponding inhibitor. Embryos treated at stage 8 were fixeddominant signal transduced downstream of the F G FRat stages 11.5 and 29–30 for in situ hybridization. Embryos treatedis the ras/MAPK pathway, leading to transcription of at stage 12.5 were fixed at stage 31 and photographed.
mesodermal genes such as Xbra. As gastrulation pro-ceeds, Xtsprouty transcription decreases and Xtspred Cellular Localization Assaytranscription increases. As a result, the ras/MAPK path- 100 pg PK Cδ-G F P mRNA was coinjected into X. laevis animal poles
with Xtspry2 or Xtsprd1. Animal caps were excised at stage 9 in 1×way is inhibited and the predominant pathway ismodified Danilchik’s medium (D FA) to prevent healing (Sater et al.,switched toward regulation of morphogenesis. A key1994) and were cultured for 10 min in 1× D FA with or without 100advantage of this mechanism is that F G F signal inter-ng/ml F G F2 or 1 !M PMA (Sigma). C aps were lightly fixed in formal-
pretation occurs relatively quickly at the cell membrane dehyde for 20 min and G F P localization was determined by confo-to allow for plastic and responsive changes to events cal microscopy.during a rapid early developmental timescale. However,this model may be complicated by positive and/or Morpholino Design and Microinjections
M Os were purchased from G ene Tools LLC . RNA splice junctionsnegative feedback due to the fact that Sproutys, andwere identified by alignment of cDNA sequences with matchinglikely Spreds, are themselves downstream targets ofgenomic sequences. X. tropicalis embryos were injected with 10FG FR signaling (Minowada et al., 1999; Nutt et al., 2001).ng total M Os before first cleavage, and were generally coinjected
In conclusion, we have uncovered consistent func- with a fluorescent control M O (Li std; G ene Tools). M O sequencestional differences between XtSprouty and XtSpred are listed as follows. ATG M Os: 4-mis Spry2, 5#-G C CTTTTAGproteins downstream of the F G FR that are used to co- TA C T C T C GTGT C CTT C -3# (mismatch bases are bold); Spry2 ATG,
5#-G C C ATTTTGTA C T C T C GT C T C C AT C -3#; Sprd1 ATG, 5#- C C TGTTordinate F GF signal interpretation during vertebrate mor-C G C C G C T C ATTGT C C C C TT-3#; Sprd2 ATG, 5#-T C C T C G C T C ATTphogenesis. It will be interesting to discover if similarTTGT C C C TG C T C A-3#; Splice M Os (intron sequences are lowermechanisms are involved in regulating other examplescase): Spry1 i1e2, 5#- C AT C TG AAAA C ctgccgatcaaaac-3#; Spry2
of RTK activity. e1i1, 5#- gtacttac C GTG A C C T C C T C G C C C C -3#; Sprd1 e1i1, 5#-ggtggcactcacT C TG G C T C C TGTT-3#; Sprd2 e2i2, 5#-ttcctta C C A GTTTGT C ATT C A GT C -3#.Experimental Procedures
Isolation of Xtsprouty and Xtspred Genes RNA Isolation and RT-PCRTotal RNA was isolated from embryos and animal caps using theand Construct Design
X. tropicalis Sprouty-related homologs were identified through TRIzol reagent according to manufacturer’s instructions (Invitro-gen). cDNA was synthesized using AMV reverse transcriptasesearches of the Wellcome Trust Full Length Database against hu-
man and X. laevis EST sequences. Four genes were identified: (Roche), and P C R reactions were performed using Taq polymerase(Roche) according to established protocol. Primer sequences areXtsprouty1 (Xtspry1; clone TNeu048o16), Xtsprouty2 (Xtspry2;

Developmental C ell700
written 5# to 3# as follows. Xtspry1 forward, C G C A GTT C C G AT C G Sef is a feedback-induced antagonist of Ras/MAPK-mediated F G Fsignalling. Nat. C ell Biol. 4, 170–174.G ATTTG C; reverse, C A C TATTTGTG C TA C C A G AA C; Xtspry2 f, C AT
G C G AATT C ATG G A G A C G A G A GTA C; r, GTGTG G C GTA GT C TGT C G Gilchrist, M.J., Zorn, A.M., Voigt, J., Smith, J.C ., Papalopulu, N.,TG G; Xtsprd1 f, C G C A C TT C C C ATATAA C C T C; r, C C TGTG GT C C A and Amaya, E. (2004). Defining a large set of full-length clones fromT C C T C A G AA G; Xtsprd2 f, G A C G GT C C T C TT C G ATG C TG C; r, C C A a Xenopus tropicalis EST project. Dev. Biol. 271, 498–516.A G C T C TG C C T C ATTATG G; Xbra f, A G A C AT C TTG G ATG A G G G; r, Gschwendt, M. (1999). Protein kinase C delta. Eur. J. Biochem. 259,G AA G G GTA C TG A C TTG A G. Xtwnt11 f, TA C T C AT C TTGTG C TG C T C 555–564.C A G G; r, A C AA G C A C G A G C AATG GTATG G; Xtwnt8 f, C TG AA G AT C
Gupta, R.W., and Mayer, B.J. (1998). Dominant-negative mutants ofAA G C A C G A C C A; r, C A G C T C C TT C TTT C C C A C TG. C ontrol primersthe SH2/SH3 adapters Nck and Grb2 inhibit MAP kinase activationfor E F1α (Hemmati-Brivanlou and Melton, 1994) and ornithine de-and mesoderm-specific gene induction by e F G F in Xenopus. Onco-carboxylase (O D C) (Heasman et al., 2000) have been previously de-gene 17, 2155–2165.scribed. Real-time RT-P C R analysis was performed using a Light-Guy, G.R., Wong, E.S., Yusoff, P., Chandramouli, S., Lo, T.L., Lim,Cycler SystemTM (Roche).J., and Fong, C .W. (2003). Sprouty: how does the branch managerwork? J. C ell Sci. 116, 3061–3068.45Ca2+ Efflux AssayHacohen, N., Kramer, S., Sutherland, D., Hiromi, Y., and Krasnow,Oocytes were isolated, injected, and cultured for 48 hr in modifiedM.A. (1998). sprouty encodes a novel antagonist of F G F signalingB arth’s saline (M BSH) at 16°C and then assayed for 45C a2+ effluxthat patterns apical branching of the Drosophila airways. C ell 92,as described (Musci et al., 1990). 10 ng of each RNA was injected253–263.along with 1 ng of p CIXR, a constitutively active F G F receptor (Nutt
et al., 2001). Hanafusa, H., Torii, S., Yasunaga, T., and Nishida, E. (2002).Sprouty1 and Sprouty2 provide a control mechanism for the Ras/MAPK signalling pathway. Nat. C ell Biol. 4, 850–858.Immunoblotting
For assays of MAPK activity, 10 embryos or animal caps were ho- Hanafusa, H., Torii, S., Yasunaga, T., Matsumoto, K., and Nishida,mogenized in 1× RIPA lysis buffer with the addition of C omplete E. (2004). Shp2, an SH2-containing protein-tyrosine phosphatase,protease inhibitor cocktail (Roche) and 50 mM sodium fluoride and positively regulates receptor tyrosine kinase signaling by dephos-10 mM sodium orthovanadate to inhibit phosphatase activity. In phorylating and inactivating the inhibitor Sprouty. J. Biol. Chem.whole embryos, freon was used to extract yolk proteins according 279, 22992–22995.to established protocol. Western blots were probed with 1:10000 Harmer, N.J., Sivak, J.M., Amaya, E., and Blundell, T. (2005). 1.65 Åmouse anti-dpMAPK (clone MAPK-YT, Sigma) or 1:2000 mouse Crystal structure of the X. tropicalis Spred1 Enabled/Vasodilator-anti-panMAPK (Clone 16, Transduction Laboratories) according to stimulated phosphoprotein homology-1 domain. F E BS Lett. 579,established protocols. 1161–1166.
Heasman, J., Kofron, M., and Wylie, C . (2000). B eta-catenin signal-ing activity dissected in the early Xenopus embryo: a novel anti-
Acknowledgments sense approach. Dev. Biol. 222, 124–134.
Hemmati-Brivanlou, A., and Melton, D.A. (1994). Inhibition of activinThe authors would like to acknowledge the advice and suggestionsreceptor signaling promotes neuralization in Xenopus. C ell 77,of Andrew Chalmers and Nancy Papalopulu. Also, thanks to273–281.Michael Gilchrist for constructing and maintaining the X. tropicalisIsaacs, H.V., Pownall, M.E., and Slack, J.M. (1994). e F G F regulatesEST database, and thanks to Jun-An Chen for providing the X.Xbra expression during Xenopus gastrulation. EM B O J. 13, 4469–tropicalis F K506 clone. This work was funded by a Wellcome Trust4481.Prize Studentship (L.F.P.) and a Wellcome Trust Senior Research
F ellowship (E.A.). Kato, R., Nonami, A., Taketomi, T., Wakioka, T., Kuroiwa, A., Mat-suda, Y., and Yoshimura, A. (2003). Molecular cloning of mammalianSpred-3 which suppresses tyrosine kinase-mediated Erk activa-
Received: O ctober 22, 2004 tion. Biochem. Biophys. Res. C ommun. 302, 767–772.Revised: January 5, 2005
Kenwrick, S., Amaya, E., and Papalopulu, N. (2004). Pilot morpho-Accepted: F ebruary 28, 2005lino screen in Xenopus tropicalis identifies a novel gene involved inPublished: April 3, 2005head development. Dev. Dyn. 229, 289–299.
Kinoshita, N., Iioka, H., Miyakoshi, A., and Ueno, N. (2003). PK CReferencesdelta is essential for Dishevelled function in a noncanonical Wntpathway that regulates Xenopus convergent extension move-Amaya, E., Musci, T.J., and Kirschner, M.W. (1991). Expression of aments. G enes Dev. 17, 1663–1676.dominant negative mutant of the F G F receptor disrupts mesodermKroll, K.L., and Amaya, E. (1996). Transgenic Xenopus embryosformation in Xenopus embryos. C ell 66, 257–270.from sperm nuclear transplantations reveal F G F signaling require-
C asci, T., Vinos, J., and Freeman, M. (1999). Sprouty, an intracellularments during gastrulation. Development 122, 3173–3183.
inhibitor of Ras signaling. C ell 96, 655–665.Lim, J., Wong, E.S., Ong, S.H., Yusoff, P., Low, B.C ., and Guy, G.R.
Christen, B., and Slack, J.M. (1997). F G F-8 is associated with ante- (2000). Sprouty proteins are targeted to membrane ruffles uponroposterior patterning and limb regeneration in Xenopus. Dev. Biol. growth factor receptor tyrosine kinase activation. Identification of192, 455–466. a novel translocation domain. J. Biol. Chem. 275, 32837–32845.Christofori, G. (2003). Split personalities: the agonistic antagonist MacNicol, A.M., Muslin, A.J., and Williams, L.T. (1993). Raf-1 kinaseSprouty. Nat. C ell Biol. 5, 377–379. is essential for early Xenopus development and mediates the in-Ciruna, B., and Rossant, J. (2001). F G F signaling regulates meso- duction of mesoderm by F G F. C ell 73, 571–583.derm cell fate specification and morphogenetic movement at the Minowada, G., Jarvis, L.A., Chi, C .L., Neubuser, A., Sun, X., Haco-primitive streak. Dev. C ell 1, 37–49. hen, N., Krasnow, M.A., and Martin, G.R. (1999). Vertebrate SproutyCiruna, B.G., Schwartz, L., Harpal, K., Yamaguchi, T.P., and Ros- genes are induced by F G F signaling and can cause chondrodys-sant, J. (1997). Chimeric analysis of fibroblast growth factor recep- plasia when overexpressed. Development 126, 4465–4475.tor-1 (Fgfr1) function: a role for F G FR1 in morphogenetic movement Musci, T.J., Amaya, E., and Kirschner, M.W. (1990). Regulation ofthrough the primitive streak. Development 124, 2829–2841. the fibroblast growth factor receptor in early Xenopus embryos.C ornell, R.A., and Kimelman, D. (1994). Activin-mediated meso- Proc. Natl. Acad. Sci. USA 87, 8365–8369.derm induction requires F G F. Development 120, 453–462. Nieuwkoop, P.D., and F aber, J. (1967). Normal Table of Xenopus
Laevis (Amsterdam: North-Holland).Furthauer, M., Lin, W., Ang, S.L., Thisse, B., and Thisse, C . (2002).

F G F Signal Interpretation by Sprys/Sprds701
Nutt, S.L., Dingwell, K.S., Holt, C .E., and Amaya, E. (2001). XenopusSprouty2 inhibits F G F-mediated gastrulation movements but doesnot affect mesoderm induction and patterning. G enes Dev. 15,1152–1166.
O’Reilly, A.M., Pluskey, S., Shoelson, S.E., and Neel, B.G. (2000).Activated mutants of SHP-2 preferentially induce elongation ofXenopus animal caps. Mol. C ell. Biol. 20, 299–311.
Robertson, S.C ., Tynan, J., and Donoghue, D.J. (2000). RTK muta-tions and human syndromes: when good receptors turn bad.Trends G enet. 16, 368.
Sasaki, A., Taketomi, T., Wakioka, T., Kato, R., and Yoshimura, A.(2001). Identification of a dominant negative mutant of Sprouty thatpotentiates fibroblast growth factor- but not epidermal growthfactor-induced ERK activation. J. Biol. Chem. 276, 36804–36808.
Sater, A.K., Alderton, J.M., and Steinhardt, R.A. (1994). An increasein intracellular pH during neural induction in Xenopus. Development120, 433–442.
Schlessinger, J. (2000). C ell signaling by receptor tyrosine kinases.C ell 103, 211–225.
Sivak, J.M., and Amaya, E. (2004). F G F signaling during gastrula-tion. In G astrulation, C . Stern, ed. (Woodbury, NY: C old Spring Har-bor Laboratory Press), pp. 463–473.
Song, J., and Slack, J.M. (1996). XF G F-9: a new fibroblast growthfactor from Xenopus embryos. Dev. Dyn. 206, 427–436.
Spokony, R., and Saint-Jeannet, J.P. (2000). Xenopus F K 506-bind-ing protein, a novel immunophilin expressed during early develop-ment. Mech. Dev. 94, 205–208.
Tang, T.L., Freeman, R.M., Jr., O’Reilly, A.M., Neel, B.G., and Sokol,S.Y. (1995). The SH2-containing protein-tyrosine phosphatase SH-PTP2 is required upstream of MAP kinase for early Xenopus devel-opment. C ell 80, 473–483.
Tefft, J.D., Lee, M., Smith, S., Leinwand, M., Zhao, J., Bringas, P.,Jr., Crowe, D.L., and Warburton, D. (1999). C onserved function ofmSpry-2, a murine homolog of Drosophila sprouty, which nega-tively modulates respiratory organogenesis. Curr. Biol. 9, 219–222.
Wakioka, T., Sasaki, A., Kato, R., Shouda, T., Matsumoto, A., Miyo-shi, K., Tsuneoka, M., Komiya, S., B aron, R., and Yoshimura, A.(2001). Spred is a Sprouty-related suppressor of Ras signalling. Na-ture 412, 647–651.
Wallingford, J.B., Ewald, A.J., Harland, R.M., and Fraser, S.E.(2001). C alcium signaling during convergent extension in Xenopus.Curr. Biol. 11, 652–661.
Whitman, M., and Melton, D.A. (1992). Involvement of p21ras inXenopus mesoderm induction. Nature 357, 252–254.
Wong, E.S., Fong, C .W., Lim, J., Yusoff, P., Low, B.C ., Langdon,W.Y., and Guy, G.R. (2002). Sprouty2 attenuates epidermal growthfactor receptor ubiquitylation and endocytosis, and consequentlyenhances Ras/ERK signalling. EM B O J. 21, 4796–4808.
Yang, X., Dormann, D., Munsterberg, A.E., and Weijer, C .J. (2002).C ell movement patterns during gastrulation in the chick are con-trolled by positive and negative chemotaxis mediated by F G F4 andF G F8. Dev. C ell 3, 425–437.
Yigzaw, Y., C artin, L., Pierre, S., Scholich, K., and Patel, T.B. (2001).The C terminus of sprouty is important for modulation of cellularmigration and proliferation. J. Biol. Chem. 276, 22742–22747.
Yigzaw, Y., Poppleton, H.M., Sreejayan, N., Hassid, A., and Patel,T.B. (2003). Protein-tyrosine phosphatase-1B (PTP1B) mediates theanti-migratory actions of Sprouty. J. Biol. Chem. 278, 284–288.
Yusoff, P., Lao, D.H., Ong, S.H., Wong, E.S., Lim, J., Lo, T.L., Leong,H.F., Fong, C .W., and Guy, G.R. (2002). Sprouty2 inhibits the Ras/MAP kinase pathway by inhibiting the activation of Raf. J. Biol.Chem. 277, 3195–3201.





























