Embed Size (px)
Citation preview

J. Plant Res. 112: 237-244, 1999 Journal of Plant Research 0 by The Botanical Society of Japan 1999
JPR Symposium
Determination of the Role of Cold Acclimation-Induced Diverse Changes in Plant Cells from the Viewpoint of Avoidance of Freezing Injury
Seizo Fujikawa’*, Yutaka Jitsuyama’ and Katsushi Kuroda3
7 Environmental Cryobiology Group, Institute of Low Temperature Science, Hokkaido University, Sapporo, 060-0819 Japan
3 Department of Forest Science, Faculty of Agriculture, Hokkaido University, Sapporo, 060-8589 Japan Department of Agrobiology and Bioresorces, Faculty of Agriculture, Hokkaido University, Sapporo, 060-8589 Japan
Cold acclimation is a complex adaptive mechanism by which plants survive freezing in winter. During cold accli- mation, diverse intracellular and extracellular changes occur. Although most of these changes are related to the acquirement of freezing tolerance, the exact role of these changes in the attainment of freezing tolerance is still unclear. In this review, we suggest the possible role of some of these cold acclimation-induced changes in relation with increased freezing tolerance from the viewpoint of inhibition of freezing injury produced by close approach of membranes.
Key words: Cell wall - Cold acclimation - Extracellular freezing - Freeze fracture replica electron microscopy - Plasma membrane
In the natural environment, water in the apoplastic spaces of plant tissues freezes when the temperatures drops to and below around -2 C. However, intracellular water does not freeze at this temperature because plasma membranes act as a barrier against ice penetration and because the cyto- plasm does not contain active ice-nucleating substances, at least at this high subzero temperature. Consequently, intracellular water is temporarily supercooled. Due to the difference in vapor pressures between supercooled water and ice, intracellular water is dehydrated toward extracellular ice to make an equlibrium between intracellular water and extracellular ice. The response of cells to further reduction in temperature varies according to the rate of temperature reduction. When the rate of temperature reduction is too rapid for an equilibrium to be established by dehydration, intracellular water is excessibly supercooled and finally results in freezing within cells. This intracellular freezing is lethal to all biological materials. On the other hand, when the rate of temperature reduction is sufficiently slow to allow for an equilibrium to be established by dehydration, cellular water is gradually dehydrated to avoid excessive supercool- ing, causing shrinkage and deformation of cells. In this type
* Corresponding author.
of extracellular freezing, cells are subjected to a higher degree of dehydration with reduction of freezing tempera- tures, and the cells that tolerate dehydration can survive under freezing. Because the rate of temperature reduction in the natural environment is sufficiently slow to allow for an equilibrium to be established, freezing of almost all plant tissue cells in a natural environment results in extracellular freezing.
Mechanism of Freezing Injury by Extracellular Freezing
Hypotheses on the mechanisms of freezing injury As the cause of freezing injury of plant tissue cells by
extracellular freezing, many hypotheses have been proposed, such as freezing-induced dehydration, and the increased concentration of the cell sap due to freezing dehydration (see, Levitt 1980). Furthermore, mechanical stresses by growth of extracellular ice may be the cause of freezing injury. Although all these hypotheses stress the important role of the plasma membranes as a primary site of injury, the direct evidence of injury is unclear.
Ultrastructural changes in plasma membranes due to freezing and their relation to the Occurrence of freezing injury
Studies using a freeze-fracture replica electron micro- scopy have shown the occurrence of distinct ultrastructural changes in the plasma membranes of plant tissues when the plant tissues are subjected down to lethal freezing tempera- tures. Freezing of plant tissues, including protoplasts iso- lated from rye leaves (Gordon-Kamm and Steponkus 1984, Fujikawa and Steponkus 1990), tissue cells of rye leaves (Webb and Steponkus 1993), protoplasts isolated from spring oat (Webb et a/. 1994), cortical parenchyma cells of mulberry (Fujikawa 1994), protoplasts isolated from Arabidopsis thaliana leaves (Uemura et a/. 1995), and shoot apex cells of asparagus (Jitsuyama et a/. 1995), as well as tertiary hyphae of edible mushrooms (Fujikawa and Miura 1986, Fujikawa 1987a, 1987b, 1990,1991) and vegetative cells of fresh water algae, Tribonema bombycinum (Nagao et a/.), produced plasma membrane ultrastructural changes showing either
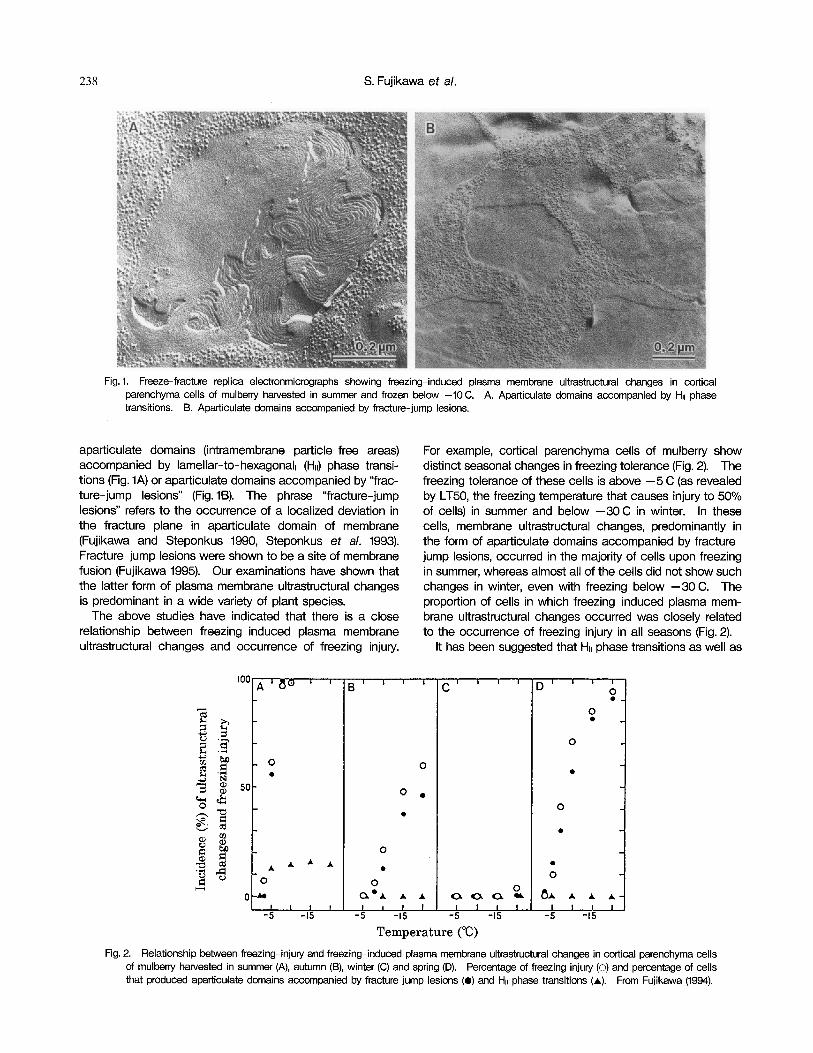
238 S. Fujikawa et a/.
Fig. 1. Freeze-fracture replica electronmicrographs showing freezing-induced plasma membrane ultrastructural changes in cortical parenchyma cells of mulberry harvested in summer and frozen below -10 C. A. Aparticulate domains accompanied by HII phase transitions. B. Aparticulate domains accompanied by fracture-jump lesions.
aparticulate domains (intramembrane particle-free areas) accompanied by lamellar-to- hexagonal11 (HII) phase transi- tions (Fig. IA) or aparticulate domains accompanied by “frac- ture-jump lesions” (Fig. 16). The phrase “fracture-jump lesions” refers to the occurrence of a localized deviation in the fracture plane in aparticulate domain of membrane (Fujikawa and Steponkus 1990, Steponkus et a/. 1993). Fracture-jump lesions were shown to be a site of membrane fusion (Fujikawa 1995). Our examinations have shown that the latter form of plasma membrane ultrastructural changes is predominant in a wide variety of plant species.
The above studies have indicated that there is a close relationship between freezing-induced plasma membrane ultrastructural changes and occurrence of freezing injury.
For example, cortical parenchyma cells of mulberry show distinct seasonal changes in freezing tolerance (Fig. 2). The freezing tolerance of these cells is above -5 C (as revealed by LT50, the freezing temperature that causes injury to 50% of cells) in summer and below -3OC in winter. In these cells, membrane ultrastructural changes, predominantly in the form of aparticulate domains accompanied by fracture- jump lesions, occurred in the majority of cells upon freezing in summer, whereas almost all of the cells did not show such changes in winter, even with freezing below -3OC. The proportion of cells in which freezing-induced plasma mem- brane ultrastructural changes occurred was closely related to the occurrence of freezing injury in all seasons (Fig. 2).
It has been suggested that HII phase transitions as well as
Temperature (“C) Fig. 2. Relationship between freezing-injury and freezing-induced plasma membrane ultrastructural changes in cortical parenchyma cells
of mulberry harvested in summer (A), autumn (B), winter (C) and spring (D). Percentage of freezing injury (0) and percentage of cells that produced aparticulate domains accompanied by fracture jump lesions (0) and HII phase transitions (A). From Fujikawa (1994).

Role of Cold Acclimation to Avoid Freezing Injury 239
membrane fusions, as revealed by the formation of fracture- jump lesions, may result in a loss of the semi-permeable property of the plasma membrane and thus lead to injury (Steponkus et a/. 1993). Our freeze-fracture studies showed that frozen cells in which these plasma membrane ultras- tructural changes occurred were severely damaged by producing numerous vesiculations of plasma membranes upon thawing (data not shown).
The cause of freezing -induced ultrastructural changes Plasma membrane ultrastructural changes due to freezing,
which are closely related to the occurrence of injury, was previously reported to be simply a result of freezing-induced dehydration (Gordon-Kamm and Steponkus 1984). In terti- ary hyphae of the mushroom, however, we verified that these freezing-induced ultrastructural changes occurred only in areas where the two membranes had closely approached each other (Fujikawa and Miura 1996, Fujikawa 1987a, 1987b, 1990,1991,1994). In areas where membranes were not closely approached, low temperature stress, osmotic stress, and a combination of both did not produce plasma mem- brane ultrastructural changes. Thus, freezing-induced plasma membrane ultrastructural changes that are closely related to the occurrence of freezing injury are now thought to be a result of interbilayer effects, which are produced by the close approach of membranes (Fujikawa and Miura 1986, Steponkus and Lynch 1989, Fujikawa and Steponkus 1990).
The molecular mechanism of membrane ultrastructural changes by the freezing-induced close approach of mem- branes was proposed by Steponkus et a/. (1993) based upon theoretical studies by Siege1 (1987). It has been suggested that when lipid bilayers are brought into close approach they may form either HII phase or inter lamellar attachment (ILA) and fusion from a common structural intermediate, referred to as an inverted micellar intermediate (IMI). Whether the IMls are converted to the HII phase or ILA and fusion is thought to be dependent on the intrinsic curvature of the constituent monolayers of lipids and the temperatures at which such conversion occurs. The monolayers with a high intrinsic curvature form the HII phase, whereas those with a low intrinsic curvature form ILA and fusion.
The stresses causing dose approach of membranes Steponkus and his-colleagues (Steponkus et a/. 1993) and
Bryant and Wolfe (1989,1992) believe that the close approach of membranes is due to cellular shrinkage caused by freezing-induced dehydration. In the case of protoplasts isolated from rye leaves (Gordon-Kamm and Steponkus 1984) and those from A. thaliana leaves (Uemura et a/. 1995), freezing-induced and osmotic manipulation-induced de- hydration produced a similar degree of plasma membrane ultrastructural change as well as a similar degree of freezing injury.
However, the above situation is apparently different in many plant tissues or plant cultured cells with cell walls. A comparison of the incidences of plasma membrane ultras- tructural changes due to freezing and osmotic manipulation at the corresponding osmolality showed that freezing caused
a significantly higher incidence of occurrence of plasma membrane ultrastructural changes as well as a higher inci- dence of occurrence of injury than did osmotic manipulation in tertiary hyphae of mushroom (Fujikawa and Miura 1986). In cortical parenchyma cells of mulberry harvested in autumn, plasma membrane ultrastructural changes occurred in almost 100% of cells due to freezing to -10 C but in only about 20% of cells due to osmotic manipulation with 5.4 osm sorbitol solution (corresponding to osmolality with -10 C freezing).
The higher incidence of occurrence of plasma membrane ultrastructural changes caused by freezing than that caused by osmotic manipulation is apparently related to the degree of cellular deformation. Freezing produced a distinct defor- mation of cells in addition to shrinkage (Fig. 3A), whereas osmotic manipulation produced generally an almost symmet- rical shrinkage of cells (Fig. 3B). Due to the distinct defor- mation and shrinkage of cells, freezing caused a close approach not only between plasma membranes and intracel- lular organelle membranes but also even between plasma membranes in the same cells. On the other hand, osmotic manipulation caused only a close approach between plasma membranes and intracellular organelle membranes. This suggests that freezing causes a higher incidence of close approach of membranes and consequently a higher inci- dence of freezing injury due to deformation of cells, at least in tertiary hyphae of mushrooms and in cortical parenchyma cells of mulberry.
The deformation of cells due to freezing is apparantly produced by the deformation of cell walls due to mechanical stress from the growth of large extracellular ice crystals (Fig. 3B). Thus, the property of cell walls may be closely related to the occurrence of freezing injury. On the one hand, the significant connection of the cell walls has been indicated to increase freezing injury in previous studies that showed a lesser degree of freezing injury in protoplasts than in tissue cells in cultured potato cells (Tao eta/. 1983), in cultured rice cells of low cryoprotectable cell lines (Watanabe et a/. 1995), and in Jerusalem artichoke tubers (Murai and Yoshida 1998). On the other hand, the significant connection of cell walls has been indicated to reduce freezing injury in previous studies that showed a higher degree of freezing injury in protoplasts than in tissue cells in cultured cells of Pyrus communis, Distichlis spicata and Spartina pectinata (Bartoro et a/. 1987). All these results (Tao et a/. 1983, Bartoro et a/. 1987, Watanabe et a/. 1995, Murai and Yoshida 1998) as well as our results about tertiary hyphae of mushrooms and cortical parenchyma cells of mulberry, together with the results in protoplasts showing no difference in the freezing tolerance between freezing and osmotic manipulation (Gor- don-Kamm and Steponkus 1984, Uemura et a/. 1995), indi- cate that the property of cell walls is significantly related to the freezing tolerance of plant cells.

240 S. Fujikawa et al.
Fig. 3. Freeze-fracture replica electronmicrographs showing different responces of tertiary hyphae of mushroom by freezing to -1OC (A) and by osmotic manipulation with 5.4 osm sorbitol solutions (B).
The Role of Cold Acclimation-induced Cellular Changes in Relation to the Inhibition of
Freezing Injury
Considering that freezing injury of plant cells is produced by the close approach of membranes, any changes to inhibit or reduce such close approach of membranes during freez- ing should improve freezing tolerance. Diverse structural and compositional changes occur in cells in cold acclimation (Levitt 1980, Sakai and Larcher 1987). Changes in the lipid composition in the plasma membranes as well as accumula- tion of soluble carbohydrates as a result of cold acclimation have been found to be the most closely related to the acquirement of freezing tolerance in many plant cells. These two changes may at least partially improve freezing tolerance by inhibiting or reducing the close approach of membranes.
Effects of lipid composition changes in the plasma mem- brane
The relationship between changes in membrane lipid composition and increased freezing tolerance as a result of cold acclimation has long been recognized in many plant cells (Yoshida 1984). Steponkus and his colleagues (Webb and Steponkus 1993, Webb et al. 1994, Uemura et al. 1995) using protoplasts isolated from rye and A. thaliana have proposed the hypothesis that increased freezing tolerance due to a change in membrane lipid composition is related to the occurrence of the close approach of membranes.
They hypothesized that the close approach of membranes is influenced by the hydration characteristics of the mem- brane lipids, which influence the spacial separation between bilayers at a given freezing temperature. An increase in bilayer hydration, which results in a greater spacial separa- tion between bilayers at a given freezing temperature, can be affected by either an increase in a highly hydrated lipid class or a decrease in the proportion of poorly hydrated classes, or
by a combination of both. In the materials they used, cold acclimation resulted in an increased level of phosphatidyl- choline (hydrated lipid) and a decreased level of cerebroside and free sterol (poorly hydrated lipid), suggesting the reduc- tion of close approach at a given freezing temperature as a result of cold acclimation. They also hypothesized that these compositional changes of lipids may affect the shift of membrane ultrastructural changes from K phase transitions to ILA and membrane fusion by close approach.
Our freeze fracture replica examinations using plasma membrane vesicles isolated from cortical parenchyma cells of mulberry harvested in winter, however, showed that these isolated plasma membrane vesicles produced close approach of membranes by freezing to -20 C (Fig. 4B), while the same freezing temperature did not produce close approach of membranes in the tissue cells (Fig. 4A). Thus, we suggest that the close approach of membranes is affected not only by compositional changes of membrane lipids but also by other factors produced by cold acclimation.
Effects of the accumulation of soluble carbohydrates Numerous studies have demonstrated that the content of
soluble carbohydrates within cells may increase several-fold in association with an increase in the freezing tolerance as a result of cold acclimation of plants (Siminovitch eta/. 1953, Parker 1962, Sakai and Yoshida 1968, Yelenosky and Guy 1977). We examined the role of accumulation of sugars in increased freezing tolerance by using a model system in which soluble sugars were artificially incorporated inside the cells. By using this system, the effect of intracellular sugars may be judged by eliminating or minimizing the other effects produced by cold acclimation.
We incubated asparagus shoot apices with a medium containing 0.5 M of glucose for 2days at 22C. By this incubation, the intracellular content of total sugars increased from 7g/mg FW before treatment to 40g/mg FW after treatment, with an accompanying increase in freezing toler-
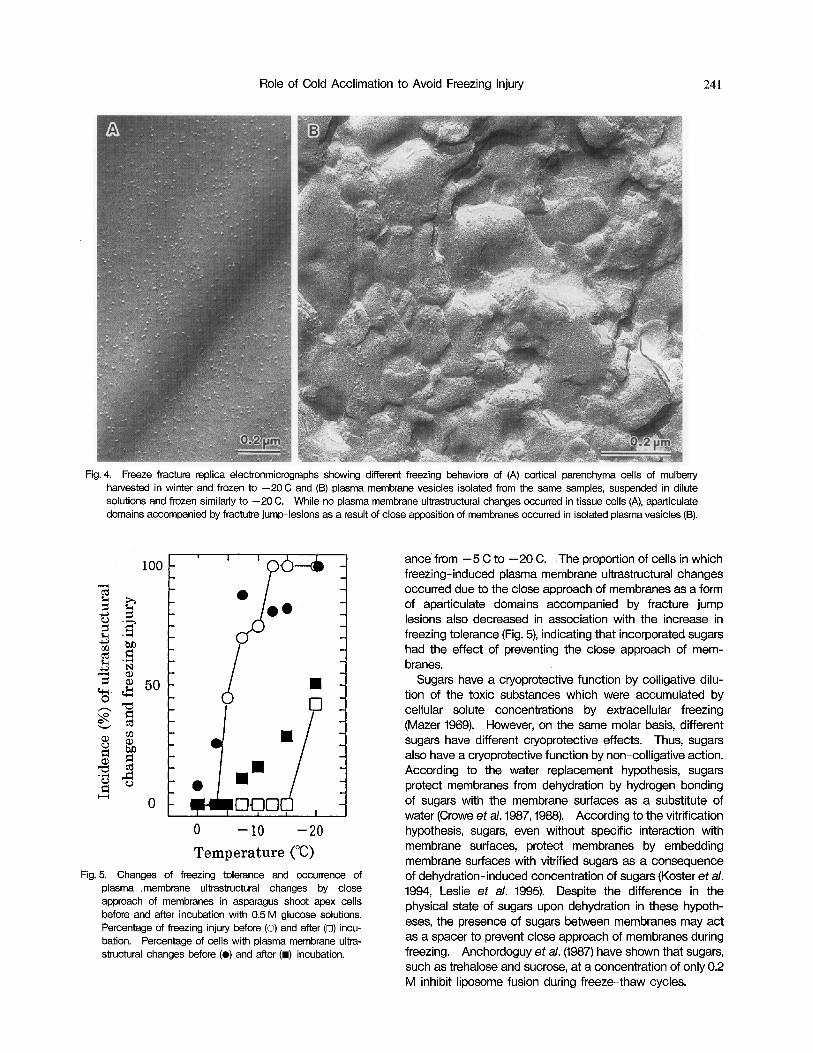
Role of Cold Acclimation to Avoid Freezing Injury 24 1
Fig. 4. Freeze-fracture replica electronmicrographs showing different freezing behaviors of (A) cortical parenchyma cells of mulbeny harvested in winter and frozen to -20 C and (B) plasma membrane vesicles isolated from the same samples, suspended in dilute solutions and frozen similarly to -20 C. While no plasma membrane ultrastructural changes occurred in tissue cells (A), aparticulate domains accompanied by fractutre jump-lesions as a result of close apposition of membranes occurred in isolated plasma vesicles (6).
I
100 ‘064 -I I- I
0 -10 -20 Temperature (“C)
Fig. 5. Changes of freezing tolerance and occurrence of plasma membrane ultrastructural changes by close approach of membranes in asparagus shoot apex cells before and after incubation with 0.5 M glucose solutions. Percentage of freezing injury before (0) and after (0) incu- bation. Percentage of cells with plasma membrane ultra- structural changes before (.) and after (.) incubation.
ance from -5 C to -20 C. The proportion of cells in which freezing-induced plasma membrane ultrastructural changes occurred due to the close approach of membranes as a form of aparticulate domains accompanied by fracture jump lesions also decreased in association with the increase in freezing tolerance (Fig. 5), indicating that incorporated sugars had the effect of preventing the close approach of mem- branes.
Sugars have a cryoprotective function by colligative dilu- tion of the toxic substances which were accumulated by cellular solute concentrations by extracellular freezing (Mazer 1969). However, on the same molar basis, different sugars have different cryoprotective effects. Thus, sugars also have a cryoprotective function by non-colligative action. According to the water replacement hypothesis, sugars protect membranes from dehydration by hydrogen bonding of sugars with the membrane surfaces as a substitute of water (Crowe et a/. 1987,1988). According to the vitrification hypothesis, sugars, even without specific interaction with membrane surfaces, protect membranes by embedding membrane surfaces with vitrified sugars as a consequence of dehydration-induced concentration of sugars (Koster et a/. 1994, Leslie et a/. 1995). Despite the difference in the physical state of sugars upon dehydration in these hypoth- eses, the presence of sugars between membranes may act as a spacer to prevent close approach of membranes during freezing. Anchordoguy et a/. (1987) have shown that sugars, such as trehalose and sucrose, at a concentration of only 0.2 M inhibit liposome fusion during freeze-thaw cycles.

242 S. Fujikawa et al.
The Role of Cold Acclimation-Induced Changes in the Cell Wall Property and Changes in the
Composition of Apoplastic Solutions in Relation to the Inhibition of Freezing Injury
Cold acclimation produces changes not only in cytoplas- mic and plasma membrane structures, as described above, but also in the property of cell walls and even in the composition of apoplastic solutions, that is, accumulation of anti-freezing proteins (AFPs). These changes in cell wall and apoplastic solutions are considered to be an additional significant phenomenon that prevents or reduces the freez- ing-induced close approach of membranes by reducing the deformation of the cells.
Effect of cell wall propem changes Cold acclimation resulted in increased rigidity of plant cell
walls (Bartolo et a/. 1987). Several biochemical changes in cell walls, including phenolic crosslinking between cell wall polymers (Fry 1983), deposition of lipids on the wall (Hunter et a/. 1981) and deposition of extensin on the wall (Stafstrom and Staehelin 1986, Kozbial et al. 1998), as a result of cold acclimation may produce cell wall rigidity. Moreover, thick- ening of cell walls as a result of cold acclimation has been found in rye leaves (Hunter et al. 1981). Cell wall rigidity has been shown to affect certain freezing characteristics of plant tissues, such as freezing-induced cell dehydration and the temperature of ice nucleation (Rajashekar and Lafta 1996). The changes of cell wall property by cold acclimation are evident in xylem ray parenchyma cells of woody plants (Fujikawa et al. 1994,1996,1999, Kuroda et a/. 1997,1999). Increased cell wall rigidity may prevent cell deformation during extracellular freezing, while fragile cell walls may facilitate cell deformation which is one of the main causes of the close approach of membranes.
The fact that cold-acclimation induces distinct changes in the cell wall property may indicate that not only cellular shrinkage due to dehydration but also cell deformation due to growth of extracellular ice crystals play a significant role in the occurrence of freezing injury by close approach of membranes. It is suggested, on the other word, that because the occurrence of freezing injury is related to cell wall deformation, cold acclimation produces rigidity of the cell walls in order to reduce the deformation.
Effect of the accumulation of AFP in sap Recently, it has been shown that accumulation of AFP or
the antifreeze activity in apoplastic sap is widespread in the plant kingdom, including vascular and nonvascular plants, as a result of cold acclimation (Kurkela and Franck 1990, Griffith et al. 1992, Urrutia et al. 1992, Duman and Olsen 1993, Hoshino et al. 1999). The plant AFPs control and reduce their growth and recrystallization of extracellular ice, rather than producing large thermal hysteresis (Griffith and Anti- kainen 1996). Marentes et al. (1993) showed that if the sap including AFPs is extracted from the cold-acclimated rye leaves before freezing, the leaves exhibit increased injury as compared to that of leaves with sap. Although the exact
role of AFPs in reducing freezing injury is unknown, we suggest that the accumulation of AFP may inhibit the growth of large extracellular ice crystals, thereby reducing cell deformation. Together with the increased rigidity of cell walls as a result of cold acclimation, accumulation of AFPs in plant sap may play a significant role in preventing cell deformation and consequent inhibition or reduction of close approach of membranes. The accumulation of AFPs as a result of cold acclimation may also implicate possible partici- pation of cell deformation to the occurrence of close approach of membranes.
Conclusion
The present article showed that diverse intracellular and extracellular changes as a result of cold acclimation may concurrently play a role in preventing freezing injury in the form of close approach of membranes. Before or without the occurrence of freezing injury in this form, however, frozen cells are also subjected to dehydration and consequent exposure to solutions with concentrated toxic substances, which may produce denaturation of cell components such as proteins. Cold acclimation, thus, also functions to protect cell components from such freezing stresses. Endo-geneous accumulation of soluble sugars, prolines, betaines and hydrophilic-boiling stable polypeptides, which accumulate in many cells as a result of cold acclimation, may play a role in protecting cell components from such dehydration-related stresses. Although soluble sugars have been shown to have the additional function of protecting cells from the close approach of membranes, further study is needed to determine whether prolines, betaines and hydro- philic-boiling stable polypeptides have a similar dual role.
This work was supported in part by a Grant-in-Aid for Science Research from the Ministry of Education, Science, Sports and Culture of Japan to S.F. and by Cooperative Recearch Funds from the Institute of Low Temperature Science, Hokkaido University. K.K. is a post-doctoral research fellow supported by the Japan Society for the Promotion of Science.
References
Anchordoguy, T.J., Rudolph, A.S., Carpenter, J.F. and Crowe, J.H. 1987. Modes of interaction of cryoprotectants with membrane phospholipids during freezing. Cryobiology 24: 324-331.
Bartolo, M.E., Wallner, S.J. and Ketchum, R.E. 1987. Com- parison of freezing tolerance in cultured plant cells and their respective protoplasts. Cryobiology. 24: 53-57.
Bryant, G. and Wolfe, J. 1989. Can hydration forces induce lateral phase separations in lamellar phases ? Eur. Biophys. J. 16: 369-374.
Bryant, G. and Wolfe, J. 1992. Interfacial forces in cryobiology and anhydrobiology. Cryo-Letters 13: 23- 36.
Crowe, J.H., Crowe, L.M., Carpenter, J.F. and Wistrom, C.A.

Role of Cold Acclimation to Avoid Freezing Injury 243
1987. Stabilization of dry phospholipid bilayers and proteins by sugars. Biochem. J. 242: 1-10,
Crowe, J.H., Crowe, LM., Carpehter, J.F., Rudolph, A.S., Wistrom, C.A., Spargo, B.J. and Anchordoguy, T.J. 1988. Interaction of sugars with membranes. Bio- chim. Biophys. Acta. 947: 367-384.
Dumann, G.J. and Olsen, T.M. 1993. Thermal hysteresis protein activity in bacteria, fungy, and phylogenetically diverse plants. Cryobiology. 30: 322-328.
Fry, S.C. 1983. Feruloylated pectins from the primary cell wall. Their structure and possible functions. Planta
Fujikawa, S. 1987a. lntramembrane particle aggregation caused by membrane-to-membrane direct contact by freezing. J. Electron Microsc. 36: 224-227.
Fujikawa, S. 1987b. Mechanical force by growth of extracellular ice crystals is widespread cause for slow freezing injury in tertiary hyphae of mushrooms. Cryo- Letters 8: 156-161.
Fujikawa, S. 1990. Cryo-scanning electron microscope and freeze-replica study on the occurrence of slow freezing injury. J. Electron Microsc. 39: 80-85.
Fujikawa, S. 1991. Lamellar to hexagonal II phase transi- tions of mushroom hyphae caused by mechanical stress resulting from the formation of extracellular ice crystals. Cryobiology 28 191-202.
Fujikawa, S. 1994. Seasonal ultrastructural alterations in the plasma membrane produced by slow freezing in cortical tissues of mulberry (Morus bombycis Koidz. cv. Goroji). Trees 8: 288-296.
Fujikawa, S. 1995. A freeze-fracture study designed to clarify the mechanisms of freezing injury due to the freezing-induced close apposition of membranes in cortical parenchyma cells of mulberry. Cryobiology 32: 444454.
Fujikawa, S. and Miura, K. 1986. Plasma membrane ultras- tructural changes caused by mechanical stress in the formation of extracellular ice as a primary cause of slow freezing injury in fruit-bodies of Basidiomycetes (Lyo- phylum ulmarium (Fr.) Kuhner). Cryobiology 23: 371- 382.
Fujikawa, S. and Steponkus, P.L. 1990. Freeze-induced alterations in the ultrastructure of the plasma membrane of rye protoplasts isolated from cold-acclimated leaves. Cryobiology 27: 665-666.
Fujikawa, S., Kuroda, K. and Fukazawa, K. 1994. Ultras- tructural study of deep supercooling of xylem ray paren- chyma cells from Styrax obassia. Micron 25: 241-252.
Fujikawa, S., Kuroda, K. and Ohtani, J. 1996. Seasonal changes in the low-temperature behaviour of xylem ray parenchyma cells in red osier dogwood (Cornus sericea L.) with respect to extracellular freezing and supercoo- ling. Micron 27: 181-191.
Fujikawa, S., Kuroda, K., Jitsuyama, Y., Sano, Y. and Ohtani, J. 1999. Freezing behavior of xylem ray parenchyma cells in softwood species with differences in the organi- zation of cell walls. Protoplasma 206: 31-40.
Gordon-Kamm, W.J. and Steponkus, P.L. 1984. Lamellar- to-hexagonal II phase transitions in the plasma mem- brane of isolated protoplasts after freeze-induced de- hydration. Proc. Natl. Acad. Sci. USA. 81: 6373-6377.
157: 111-123.
Griffith, M., Ala, P., Yang, D.S.C., Hon, W.C. andMoffatt, B.A. 1992. Antifreeze protein produced endogenously in winter rye leaves. Plant Physiol. 100: 593-596.
Griffith, M. and Antikainen, M. 1996. Extracellular ice for- mation in freezing-tolerant plants. Adv. Low-Temp. Biol. 3: 107-139.
Hoshino, T., Odaira, M., Yoshida, M. and Tsuda, S. 1999. Physiological and biochemical significance of anti- freeze substances in plarlts. J. Plant Res. 112: 255- 261.
Hunter, N.P.A., Palta, J.P., Li, P.H. and Carter, J.V. 1981. Anatomical changes in leaves of puma rye in responce to growth at cold-hardening temperatures. Bot. Gaz.
Jitsuyama, Y., Suzuki, T., Harada, T. and Fujikawa, S. 1995. Effects of pretreatments on cryopreservation in aspara- gus shoot apices. Cryobiol. Cryotech. 41: 50-57 (in Japanese).
Koster, K., Webb, M.S., Bryant, G. and Lynch, D.V. 1994. Interactions between soluble sugars and POPC (I-pal- mitoyl-2-oleoylphosphatidylcholine) during dehydration. Vitrification of sugars alters the phase behavior of the phospholipid. Biochim. Biophys. Acta. 1193: 143150.
Kozbial, P.Z., Jerzrnanowski, A., Shirsat, A.H. and Kacper- ska, A. 1998. Transient freezing regulates expression of extensin-type genes in winter oilseed rape. Physiol. Plant. 103: 264-270.
Kurkera, S. and Franck, M. 1990. Cloning and characteri- zation of a cold- and ABA-inducible Arabidopsis gene. Plant Mol. Biol. 15: 137-144.
Kuroda, K., Ohtani, J. and Fujikawa, S. 1997. Supercooling of xylem ray parenchyma cells in tropical and subtropi- cal hardwood species. Trees 12: 97-106.
Kuroda, K., Ohtani, J., Kubota, M. and Fujikawa, S. 1999. Seasonal changes in the freezing behavior of xylem ray parenchyma cells in four boreal hardwood species. Cryobiology 38: 81-88.
Leslie, S.B., Israeli, E., Lighthart, B., Crowe, J.H. and Crowe, L.M. 1995. Trehalose and sucrose protect both mem- branes and proteins in intact bacteria during drying. Appl. Environ. Microbiol. 61: 3592-3597.
Levitt, J. 1980. Responses of Plants to Environmental Stresses. Vol. I. Chilling, Freezing, and High Tempera- ture Stresses. Academic Press, New York.
Marentes, E., Griffith, M., Mlynarz, A. and Brush, R.A. 1993. Proteins accumulate in the apoplast of winter rye leaves during cold acclimation. Physiol. Plant. 87: 499-507.
Mazer, P. 1969. Freezing injury in plants. Annual Rev. Plant Physiol. 2 0 419-448.
Murai, M. and Yoshida, S. 1998. Evidence for the cell wall involvement in temporal changes in freezing tolerance of Jerusalem artichoku (Helianthus tuberosus L.) tubers during cold acclimation. Plant Cell Physiol. 39: 97- 105.
Nagao, M., Arakawa, K., Takezawa, D., Yoshida, S. and Fujikawa, S. 1999. Akinete formation in Tribonema bombycinum Derbes et Solier (Xanthophyceae) in rela- tion to freezing tolerance. J. Plant Res. 112: in press.
Parker, J. 1962. Relationships among cold hardiness, water-soluble protein, anthocyanins, and free sugars in Hedera helix L. Plant Physiol. 37: 809-813.
142: 55-62.

244 S. Fujikawa et a/.
Rajashekar, C.B. and Lafta, A. 1996. Cell-wall changes and cell tension in response to cold acclimation and exogenous abscisic acid in leaves and cell cultures. Plant. Physiol. 111: 605-612.
Sakai, A. and Yoshida, S. 1968. The role of sugar and related compounds in variations of freezing resistance. Cryobiology 5: 160-174.
Sakai, A. and Larcher, W. 1987. Frost Survival of Plants. Responses and Adaptation to Freezing Stress. Sprin- ger-Verlag, Berlin.
Siegel, D.P. 1987. Membrane-membrane interactions via intermediates in lamellar-to- hexagonal phase transi- tions. In A.E. Sowers, ed., Cell Function, Plenum Press, New York, pp. 81-208.
Siminovitch, D., Wilson, C.M. and Briggs, D.R. 1953. Studies on the chemistry of the living bark of the black locust in relation to its frost hardiness. V. Seasonal transformations and variations in the carbohydrates. Starch-sucrose interconversions. Plant Physiol. 28: 383-399.
Stafstrom, J.P. and Staehelin, L.A. 1986. The role of carbo- hydrate in maintaining extensin in an extended confor- mation. Plant Physiol. 81: 242-246.
Steponkus, P.L. and Lynch, D.V. 1989. Freeze/thaw-in- duced destabilization of the plasma membrane and the effects of cold acclimation. J. Bioenerg. Biomembr. 21:
Steponkus, P.L., Uemura, M. and Webb, M.S. 1993. A contrast of the cryostability of the plasma membrane of winter rye and spring oat. Two species that widely differ in their freezing tolerance and plasma membrane
21 -41.
lipid composition. Adv. Low Temp. Biol. 2: 211-312. Tao, D., Li, P.H. and Carter, J.V. 1983. Roll of cell wall in
freezing tolerance of cultured potato cells and their protoplasts. Physiol. Plant. 58: 527-532.
Uemura, M., Joseph, R.A. and Steponkus, P.L. 1995. Cold acclimation of Arabidopsis thaliana. Effect on plasma membrane lipid composition and freeze-induced lesions. Plant Physiol. 109: 15-30.
Urrutia, M.A., Duman, J.G. and Knight, C.A. 1992. Plant thermal hysteresis proteins. Biochim. Biophys. Acta.
Watanabe, K., Kawai, F. and Kanamori, M. 1995. Factors affecting cryoprotectability of cultured rice (Oryza sativa L.) cells. Cell wall and cell-aggregate size. Cryo- Letters 16: 147-156.
Webb, M.S. and Steponkus, P.L. 1993. Freeze-induced membrane ultrastructural alterations in rye (Secale cere- ale) leaves. Plant Physiol. 101: 955-963.
Webb, M.S., Uemura, M.and Steponkus, P.L. 1994. A com- parison of freezing injury in oat and rye. Two cereals at the extremes of freezing tolerance. Plant Physiol.
Yelenosky, G. and Guy, C.L. 1977. Carbohydrate accumu- lation in leaves and stems of Valencia orange at pro- gressively colder temperatures. Bot. Gaz. 138: 13-17.
Yoshida, S. 1984. Chemical and biophysical changes in the plasma membrane during cold acclimation of mul- berry bark cells (Morus bombycis Koidz. cv Goroji). Plant Physiol. 76: 257-265.
1121: 199-206.
104: 467-478.
(Received January 4, 1999; accepted April 7, 1999)





























