Embed Size (px)
Citation preview

Vol. 166, No. 2, 1990
January 30, 1990
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Pages 873-878
DETERMINATION OF FOUR APPARENT EERCURY INTERACTION SITES
IN PHOTOSYSTEM II BY USING A NEW MODIFICATION OF THE
STERN-VOLMER ANALYSIS
Guy SAMSON, Jean-Claude MORISSETTE and Radovan POPOVIC*
Departement de Chimie, Universite du Quebec h Montreal Case postale 8888, succursale A Montrbal Qc, Canada H3C 3P8
Received December 8, 1989
We used the Stern-Volmer method to analyze the mercury fluorescence quenching effect in the green alga Dunaliella tertiolecta. To this end, we introduced a new modification of the Stern-Volmer equation on the basis of the Lineweaver-Burk analysis used to characterize allosteric enzyme activity. This modification was useful to determine the Stern-Volmer constant, the parameter indicating the fraction of PSI1 fluorescence susceptible to the mercury quenching effect (Fs), and to estimate the apparent number of mercury binding sites (Napp = 3.72) on PSI1 which af feet the variable fluorescence. This value of Napp indicates the possibility of four mercury binding sites in the PSI1 complex. We suggested that this may be related to the mercury inhibition of the oxygen-evolving complex containing four Hn active sites. Q 1990 Academic Press, Inc.
Significant mercury inhibitory effects on photosystem II (PSI11 in algal
cells and thylakoid membranes were previously reported. The inhibition of PSI1
activity is consequent to a mercury effect on the PSI1 oxidizing side at the
water-splitting system (l-4). Recently, we indicated that the mercury
quenching effect on variable fluorescence is related to the inactivation of
some PSI1 reaction centers due to the loss of the functional interaction
between the water-splitting system and the PSI1 reaction centers (4). Further
study of mercury fluorescence quenching requires new parameters which are
necessary to describe the mechanisms of the PSI1 inhibition by mercury.
Earlier, the Stern-Volmer method was shown to be suitable to provide useful
parameters for the understanding of chlorophyll fluorescence quenching
mechanisms in chloroplasts (5,6). In this report, we used the Stern-Volmer
method to analyze the mercury fluorescence quenching effect in the green alga
*To whom correspondence should be addressed.
ABBREVIATIONS: Chl: chlorophyll: DCMU: 3-(3,4-Dichlorophenyl)-1, 1-dimethyl urea: PO: non-photochemical fluorescence; Fmax: maximum yield of fluorescence in the presence of DCMU; Fvmax = Fmax-Fo; PSII: Photosystem II.
0006-291X/90 $1.50
873 Copyright 0 1990 by Academic Press, Inc.
All rights of reproduction in any form reserved.
Vol. 166, No. 2, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Dunaliella tertiolecta. In order to further characterize the mercury in-
hibitory effect on PSI1 photochemistry, we introduced a new modification of
the Stern-Volmer equation.
EATERIALS AND EETHODS
The alga p. tertiolecta Butcher was cultured and harvested as previously described (4). Total chlorophyll concentration was estimated in 80% acetone as by Strain & d (7). Cells were dark-adapted for 30 min and were then incubated for 15, 30, 45, 60 and 75 min in the dark with various concentra- tions of mercuric chloride. Algal cells were treated with 20 pE DCEU for 60 set prior to fluorescence measurements. Haximum yield of variable fluores- cence (Fvmax = Fmax - Fo) (with algal containing 7pg Chl tot/ml) was measured by the method described (8,9). Fmax represents the maximum yield of fluores- cence in the presence of DCEU, and Fo is the constant fluorescence level not related to PSI1 photochemistry.
RESULTS AND DISCUSSION
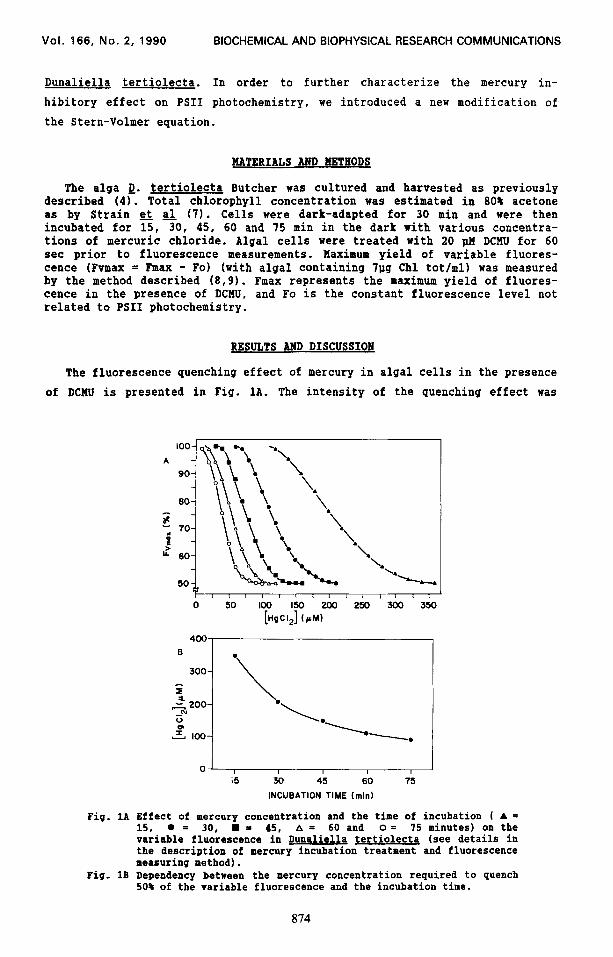
The fluorescence quenching effect of mercury in algal cells in the presence
of DCMU is presented in Fig. 1A. The intensity of the quenching effect was
r 2
3
”
9
200 1
100
00 I5 30 45 60 75
INCUBATION TIME (min)
Fig. 1A Effect of mercury concentration and the time of incubation ( A = 15. l = 30, w = 45, A = 60 and o = 75 minutes) on the variable fluorescence in Dunaliella tertiolecta (see details in the description of mercury incubation treatment and fluorescence measuring method).
Pig. 1B Dependency between the mercury concentration required to quench 50% of the variable fluorescence and the incubation time.
874

Vol. 166, No. 2, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
dependent on the incubation time and on the mercury concentration. Lower
mercury concentrations at a longer time of incubation were sufficient to
induce an equivalent quenching effect seen for higher mercury concentrations
at a shorter time of incubation. The shape of the curves for the different
incubation times was similar, indicating that the incubation time did not
influence the pattern of mercury interaction with PSII. Regardless of the
incubation time and mercury concentration, 50% of the initial variable
fluorescence can not be quenched by mercury. Since this fluorescence quenching
was attributed to a mercury inhibitory effect at the water-splitting system
(2,3), it indicates the presence of an unsusceptibility of some PSI1 to the
mercury inhibitory effect. The PSI1 fluorescence which was not susceptible to
the mercury effect was also reported recently (4). We noticed that the mercury
concentration required to quench 50% of the variable fluorescence decreases as
the incubation time increases (Fig. 1B). However, more than 75 min of
incubation does not change the concentration of mercury required to quench 50%
of the variable fluorescence, indicating a steady-state quenching effect. The
exponential relationship observed between the mercury concentration effect and
the incubation time is similar to the kinetic of ion diffusion into the algal
cell (10). Therefore, it seems that the intracellular concentration of AgClz
(responsible for fluorescence quenching) reaches an equilibrium shortly after
75 min of incubation.
To analyze the mercury quenching effect on variable fluorescence suscep-
tible and non-susceptible to mercury, the modified Stern-Volmer equation was
used :
10 = 1 + 1 - AI FsKsvQ Fs (1)
where 10 is the maximum variable fluorescence yield (Fvmax) in the absence of
the quencher (mercury), AI represents the difference of Fvmax in the absence
and in the presence of mercury, Ksv is the Stern-Volmer quenching constant,
[Q] is the quencher effect and Fs is the fluorescence fraction susceptible to
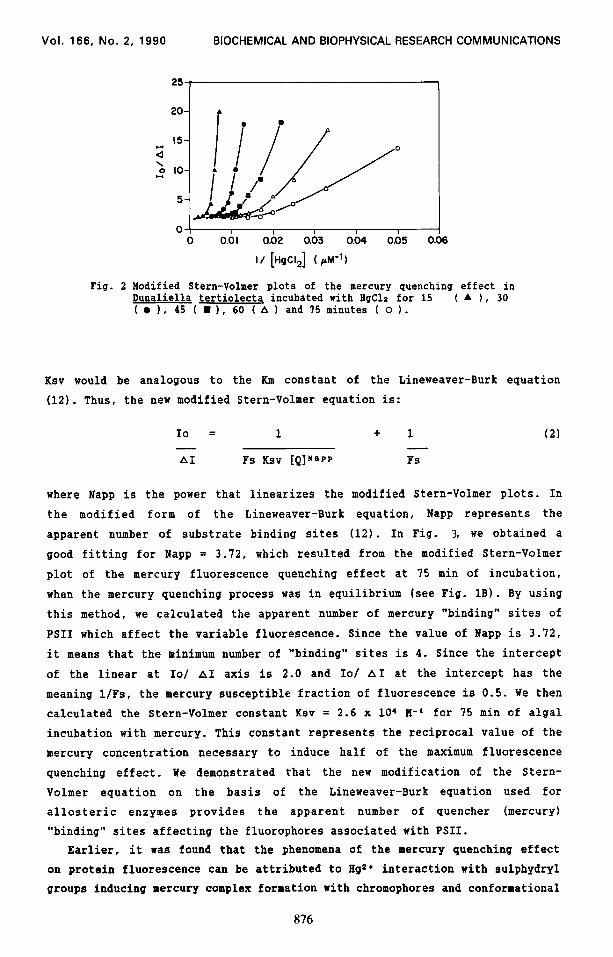
the quencher effect (5,6,11). All plots of IO/ AI versus l/[HgCla] are curved
and tend to an intercept value of 2.0 at the IO/ AI-axis (Fig. 2). Thus, the
mercury fluorescence quenching effect does not adhere to the linear relation-
ship defined by the modified Stern-Volmer equation (eq. 1). However, we
observed that the modified Stern-Volmer plots of the mercury quenching data
are analogous to the Lineweaver-Burk plot derived from the substrate activity
curve for an allosteric enzyme (12). Therefore, by the analogy of curve
behaviour and related equations, we replaced the substrate concentration with
the quencher concentration and the enzyme reaction velocity with the fluores-
cence quenching intensity. In this case, the Stern-Volmer quenching constant
875
Vol. 166, No. 2, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
25
0 I I I I I 0 0.01 0.02 0.03 0.04 0.05 0
I/ [H&Id ( /A-‘)
6
Fig. 2 Modified Stern-Volmer plots of the mercury quenching effect in Dunaliella tertiolecta incubated with HgClz for 15 ( A ), 30 ( l 1, 45 ( n ), 60 ( A 1 and 75 minutes ( o 1.
Ksv would be analogous to the Km constant of the Lineweaver-Burk equation
(12). Thus, the new modified Stern-Volmer equation is:
10 = 1 + 1 (2) - - AI Fs Ksv iQINsPP Fs
where Napp is the power that linearizes the modified Stern-Volmer plots. In
the modified form of the Lineweaver-Burk equation, Napp represents the
apparent number of substrate binding sites (12). In Fig. 3, we obtained a
good fitting for Napp = 3.72, which resulted from the modified Stern-Volmer
plot of the mercury fluorescence quenching effect at 75 min of incubation,
when the mercury quenching process was in equilibrium (see Fig. 18). By using
this method, we calculated the apparent number of mercury “binding” sites of
PSI1 which affect the variable fluorescence. Since the value of Napp is 3.72,
it means that the minimum number of “binding” sites is 4. Since the intercept
of the linear at IO/ AI axis is 2.0 and IO/ AI at the intercept has the
meaning l/Fs, the mercury susceptible fraction of fluorescence is 0.5. We then
calculated the Stern-Volmer constant Ksv = 2.6 x 104 K-I-’ for 75 min of algal
incubation with mercury. This constant represents the reciprocal value of the
mercury concentration necessary to induce half of the maximum fluorescence
quenching effect. We demonstrated that the new modification of the Stern-
Volmer equation on the basis of the Lineweaver-Burk equation used for
allosteric enzymes provides the apparent number of quencher (mercury)
“binding” sites affecting the fluorophores associated with PSII.
Earlier, it was found that the phenomena of the mercury quenching effect
on protein fluorescence can be attributed to Hg 2+ interaction with sulphydryl
groups inducing mercury complex formation with chromophores and conformational

Vol. 166, No. 2, 1990 BlOCHEMlCAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
0 5 IO
’ ’ [ H9C$1 3.72 -6
x IO
5
Fig. 3 Linearized form of the modified Stern-Volmer plot of the mercury quenching effect in Dunaliella tertiolecta incubated for 75 minutes with BgC12.
change of protein (13,14). Also, the copper quenching effect of fluorescence
in chloroplasts was shown as a consequence of copper binding to a protein
component close to the water-splitting system (15.16). Taking into account
these specificities of metal-protein interaction in view of our results, we
assume that the mercury quenching effect results from mercury binding with
protein associated with the water-splitting system. Our results support the
idea of using Lineweaver-Burk analysis to estimate the apparent number of
quencher binding sites. The apparent number of four mercury binding sites
suggests that this may be related to water-splitting proteins associated with
four manganese atoms (17,18). Further experiments will be required to define
the nature of mercury binding process with PSI1 complex.
ACKNOWLEDGIENTS: This work was supported by the Natural Sciences and Engineering Research Council of Canada (NSERC) through grants no. G-1929, G- 1930 and A-3047, and by technical assistance of the Centre de Recherche en Photobiophysique at the U.Q.T.R.
REFERENCES
1. Kimimura, N., and Katoh, S. (1972) Biochim. Biophys. Acta 283, 279-292.
2. Honeycutt, R.C.. and Krogmann, D.W. (1972) Plant Physiol. 49, 376-380.
3. de Filippis, L-P., Hampp, R., and Ziegler, H. (1981) Arch. Hicrobiol. 128. 407-411.
4. Samson, G., and Popovic, R. (in press) J. Photochem. Photobiol.
5. Karuktis, K-K., Boegeman, S.C., Fruetel, J-A., Gruber, S.M., and Terris, M.H. (1987) Biochim. Biophys. Acta 891, 256-264.
6. Karuktis, K-K-, Gruber, S-M., Fruetel, J.A., and Boegeman, S.C. (1988) Biochim. Biophys. Acta 932, 84-90.
877

Vol. 166, No. 2, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
1.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
Strain, H-H., Cope, B.T., and Svec, W-A. (1972) in Methods in Enzymology 23, 452-416.
Morissette, J.C., Eeunier, P-C., and Popovic, R. (1988) Rev. Sci. Instrum. 59, 934-936.
Horissette, J-C., and Popovic, R. (1987) Biochem. Biophys. Res. Comm. 149, 385-390.
Walker, B.A., and Pitman, M.G. (1976) in: Encvclooedia of Plant Physio- logy (LUttge, U. and Pitman, M.G., eds) vol. ZA, pp. 93-126, Springer- Verlag, Berlin.
Lakowicz, J-R. (1984) Principles of Fluorescence Spectroscopy, Plenum Press, New York.
Segel, I.H. (1975) Enzyme Kinetics, John Wiley & Sons, New York.
Chen, R.F. (1971) Arch. Biochim. Biophys. 142, 552-564.
Chen, R.F. (1976) in: Biochemical Fluorescence: Concepts, (Chen, R.F. and Edelhoch, Ii., eds), vol. 2, pp. 573-606, Marcel Dekker Inc., New York.
Vierke, G., and Struckmeier, P. (1977) 2. Naturforsh. 32c, 605-610.
Vierke, G. and Struckmeier, P. (1978) 2. Naturforsh. 33c, 266-270.
Govindjee, Kambara, T., and Coleman, W. (1986) Photochem. Photobiol. 42, 187-210.
Enami, I., Miyaoka, T., Mochizuki, Y., Shen, J-R., Satoh, K., and Katoh, S. (1989) Biochim. Biophys. Acta 973, 5-40.
878





























