Embed Size (px)
Citation preview

1022-7954/04/4012- © 2004
MAIK “Nauka
/Interperiodica”1371
Russian Journal of Genetics, Vol. 40, No. 12, 2004, pp. 1371–1377. Translated from Genetika, Vol. 40, No. 12, 2004, pp. 1654–1661.Original Russian Text Copyright © 2004 by Galaev, Babayants, Sivolap.
INTRODUCTION
During the past decades, the limited capacity ofgenetic pool of industrial wheat cultivars to providegenes for resistance to biotic and abiotic stress factorsmotivated the search for methods of enhancing thewheat genome at the expense of genetic material of itswild relatives. Species closely related to cultured wheatare a source of agronomically valuable genes thatwhich can be used in wheat breeding. Many commer-cial characters, including resistance to fungal diseases,were transferred to bread wheat from various
Aegilops
species via distant hybridization [1–4].Rybalka undertook experiments on introgression of
Aegilops cylindrica
genome (CCDD; 2
n
= 28) elementsinto
Triticum aestivum
(AABBDD; 2
n
= 42). Theresulting material was used to select wheat forms(BC
1
F
9
) manifesting complex resistance to a number ofwheat fungal diseases [5–7].
It is known that the C genome of
Ae. cylindrica
, sim-ilar to the C genome of
Ae. caudata
[8], is capable ofpartially inhibiting the wheat diploidization system(
Ph
genes). Under partial suppression of
Ph
genes, thechromosomes of
Ae. cylindrica
D and C genomes canbe integrated into homeologous associations, and char-acteristically, chromosomes of the
Ae. cylindrica
Dgenome are capable of pairing with chromosomes of Aand B wheat genome, while chromosomes of
Ae. cylin-drica
C genome pair with chromosomes of A wheatgenome [9].
Avsenin
et al.
[9] believe that genes for diseaseresistance transmitted from
Ae. cylindrica
[5–7] to
wheat are located not only in D, but also in theA genome, because it is the C genome that ensuresresistance to diseases in
Aegilops
species [10].For effective employment of forms carrying frag-
ments of the
Ae. cylindrica
genome related to a gene (orgenes) of resistance to fungal diseases [5–7]), it is ofinterest to conduct a comparative analysis of recipientand donor genomes and to establish the variation typeof introgressive wheat forms resulting from distanthybridization.
To find a solution to these tasks, we used two sys-tems of PCR analysis, ISSR and SSR–PCR. The lattersystem operates with the help of microsatellite markersthat display various advantages: the monolocus type,multiallelism, and codominance [11, 12]. In ISSR–PCR analysis, the primers used are tandem 2–4 nucle-otide repeats complementary to short microsatellitesequences with an additional selective nucleotide at the5' or 3' ends [13], which possess such properties as mul-tilocus character, biallelism, and dominance [14, 15].The aim of this study was to conduct a comparativeanalysis of introgressive and parental forms and iden-tify donor genome sites in wheat that received genes ofresistance to fungal diseases from
Ae. cylindrica.
MATERIALS AND METHODS
We examined DNA of 30 individual plants BC
1
F
9
ofintrogressive lines 5/55-91 and 5/20-91 (15 plants perline) and DNA of parental forms: 20 individual plantsof cultivar Odesskaya semidwarf, 24
Ae. cylindrica
Detection of the Introgression of Genome Elements of the
Aegilops cylindrica
Host into the
Triticum aestivum
L. Genome by ISSR and SSR Analysis
A. V. Galaev
1
, L. T. Babayants
2
, and Yu. M. Sivolap
1
1
South Plant Biotechnology Center, Academy of Agrarian Sciences of Ukraine, Odessa, 65036 Ukraine; e-mail: [email protected]
2
Institute of Breeding and Genetics, Odessa, 65036 Ukraine
Received July 19, 2003
Abstract
—To reveal sites of the donor genome in wheat crossed with
Aegilops cylindrica
, which acquired con-ferred resistance to fungal diseases, a comparative analysis of introgressive and parental forms was conducted.Two systems of PCR analysis, ISSR and SSR–PCR, were employed. Upon use of 7 ISSR primers in genotypesof 30 individual plants BC
1
F
9
belonging to lines 5/55-91 and 5/20-91, 19 ISSR loci were revealed and assignedto introgressive fragments of
Aegilops cylindrica
genome in
Triticum aestivum.
The 40 pairs of SSR primersallowed the detection of seven introgressive alleles; three of these alleles were located on common wheat chro-mosomes in the B genome, while four alleles, in the D genome. Based on data of microsatellite analysis, it wasassumed that the telomeric region of the long arm of common wheat chromosome 6A also changed. ISSR andSSR methods were shown to be effective for detecting variability caused by introgression of foreign geneticmaterial into the genome of common wheat.
PLANTGENETICS

1372
RUSSIAN JOURNAL OF GENETICS
Vol. 40
No. 12
2004
GALAEV
et al
.
lines (DNA mixtures of 5 individual plants per mixturewere applied in ISSR analysis), and 20 individual
Ae.cylindrica
plants (in SSR analysis).
DNA was isolated from etiolated seedlings with aSTAV buffer [16]. A Thercyc thermocycler (DNA-Technology, Russia) was used for amplification.
Amplification was conducted with 7 ISSR primers((AG)
9
C, (AGC)
6
G, (ACC)
6
G, (GAG)
6
C, (CTC)
6
A,(GA)
9
C, and (CT)
9
G) along with 40 pairs of primers formicrosatellite loci (Table 1). In agreement with the dataobtained by Roder
et al.
[17], of the 40 microsatelliteloci studied, 11 were located on chromosomes of thecommon wheat A genome; 10 loci, on chromosomes ofthe B genome; 18 loci were located in the D genome;the location of one locus is unknown. SSR analysis wasconducted using microsatellite loci residing in theD genome as well as in A and B wheat genomes, sinceit is known that translocations of the
Ae. cylindrica
genome into the A or B genome of
T. aestivum
mayoccur [8, 9]. The composition of the reaction mixtureand amplification conditions for conducting ISSR–PCR and SSR–PCR are described in [18] and [11],respectively.
ISSR–PCR amplification products in 1
×
TBEbuffer were separated by electrophoresis in a 2% agar-ose gel 10 cm in length or in a 7% nondenaturating
0.75-mm-thick polyacrylamide gel 17.5
×
22 in size at500 V for 1.0–1.5 h.
SSR–PCR fragments in 1
×
TBE buffer were frac-tionated in a 2% agarose gel 10 cm in length or in a 10%nondenaturating 0.75-mm-thick polyacrylamide gel17.5
×
22 in size at 500 V for 2 to 2.5 h.Immediately prior to electrophoretic separation of
amplification products, agarose gels were stained withethidium bromide, and after electrophoresis, gels werephotographed using 127-TASMA-2-81-B film. DNAfragments in polyacrylamide gels were stained with sil-ver according to a Silver sequence TM DNA Sequenc-ing System Technical Manual (Promega). The size ofthe DNA amplification fragments (in bp) was deter-mined using the Image Master 1D program. Calibrationof molecular mass was made using standards pGEMand
λ
/
Pst
1.DNA fragments typical for the donor were assigned to
introgressive loci (ISSR–PCR) and alleles (SSR–PCR).
RESULTS AND DISCUSSION
ISSR and SSR Analyses of Polymorphismin Parental Forms
ISSR analysis of 20 individual plants of cultivarOdesskaya semidwarf conducted with seven primers
Table 1.
Primers to wheat microsatellite loci and their location
Primer Locus Chromosomelocalization Primer Locus Chromosome
localization
WMS 3 Xgwm 3 3DL WMS 325 Xgwm 325 6DS
WMS 5 Xgwm 5 3AL WMS 341 Xgwm 341 3DL
WMS 13 Xgwm 13 Unknown WMS 357 Xgwm 357 1AL
WMS 18 Xgwm 18 1BS WMS 383 Xgwm 383 3DL
WMS 46 Xgwm 46 7BS WMS 389 Xgwm 389 3BS
WMS 52 Xgwm 52 3DL WMS 408 Xgwm 408 5BL
WMS 95 Xgwm 95 2AS WMS 427 Xgwm 427 6AL
WMS 155 Xgwm 155 3AL WMS 437 Xgwm 437 7DL
WMS 160 Xgwm 160 4AL WMS 456 Xgwm 456 3DL
WMS 161 Xgwm 161 3DS WMS 458 Xgwm 458 1DL
WMS 165 Xgwm 165 4AS, 4BL, 4DL WMS 493 Xgwm 493 3BS
WMS 169 Xgwm 169 6AL WMS 513 Xgwm 513 4BL
WMS 186 Xgwm 186 5AL WMS 533 Xgwm 533 3BS
WMS 190 Xgwm 190 5DS WMS 577 Xgwm 577 7BL
WMS 192 Xgwm 192 5DS WMS 619 Xgwm 619 2BL
WMS 194 Xgwm 194 4DL WMS 631 Xgwm 631 7AS
WMS 212 Xgwm 212 5DL WMS 645 Xgwm 645 3DL
WMS 261 Xgwm 261 2DS WMS 664 Xgwm 664 3DL
WMS 304 Xgwm 304 5AS WMS 680 Xgwm 680 6BS
WMS 314 Xgwm 314 3DL WMS 695 Xgwm 695 4A*
* The arm of chromosome in which microsatellite locus was located is unknown.
RUSSIAN JOURNAL OF GENETICS
Vol. 40
No. 12
2004
DETECTION OF THE INTROGRESSION OF GENOME ELEMENTS 1373
revealed 131 amplified fragments, 49 of which werepolymorphic. The mean intracultivar heterogeneityparameter was 37.4% (Table 2). Primer (AG)
9
C thatdetected the greatest number of polymorphic ISSR lociseems to be most informative for the characterization ofpolymorphism.
In molecular genetic study of the heterogeneity in24
Ae. cylindrica
lines from the local population, 122ISSR loci, 54 of which were polymorphic, weredetected through amplification. The mean intraspeciesheterogeneity parameter of 24
Ae. cylindrica
lines was43.2% with the use of 7 ISSR primers, which is by5.8% greater than in cultivar Odesskaya semidwarf(Table 2). The greatest number of polymorphic ISSRloci in 24
Ae. cylindrica
lines was detected when usingprimers (AG)
9
C and (CTC)
6
A.
SSR analysis allowed the detection of 91 alleles, 49of which were found in
Ae. cylindrica
and 56, in culti-var Odesskaya semidwarf. For 18 microsatellite mark-ers, 22 monomorphic alleles were detected: 8 of thesealleles were localized on chromosomes of commonwheat in the A genome, 5 alleles, in the B genome, 8, inthe D genome, and the location of the monomorphicallele at locus Xgwm 13 is unknown. Specific primersfor the
Ae. cylindrica
genome were 26 alleles, whereasfor the genome of
T. aestivum
cultivar Odesskaya semi-dwarf, 34 alleles were specific.
Individual
Ae. cylindrica
plants in the examinedsample lacked the following loci: those located on chro-mosomes of common wheat in the A genome (Xgwm 5,Xgwm 427 (in 10 of 20 plants), Xgwm 631), in the Bgenome (408, Xgwm 513, Xgwm 680), and in Dgenome (Xgwm 52).
The genetic identity index I of analyzed
Ae. cylin-drica
forms and cultivar Odesskaya semidwarf deter-mined according to Nei and Lee [19] with respect to
allelic variants of 40 microsatellite loci was 43.81%.Chebotar and Sivolap [20] showed that I obtained forcultivar Odesskaya semidwarf and
Ae. cylindrica
inRAPD analysis constitutes 55.56%, which is by11.75% higher than the value obtained on the basis ofmicrosatellite analysis. A lower genetic identity esti-mate obtained in SSR analysis for
Ae. cylindrica
andcultivar Odesskaya semidwarf is connected with highrecombination frequency of reiterated genome regionsidentified using microsatellite markers, whereas RAPDis a method for the identification of mostly uniquesequences. Sivolap
et al.
[11] calculated the averagevalue of discrimination ability (
PIC
) measuring discrim-ination vigor of the locus with respect to the number andrelative frequency of identified alleles. According to thedata in [11], it was 0.59 for SSR markers among 28 com-mon wheat cultivars, which is higher than the value cal-culated in RAPD analysis (
PIC
= 0.32).
ISSR and SSR Analyses of Introgression of Aegilops Genome Elements into Common Wheat Genome
With the use of seven ISSR primers, the lociencountered both in
Ae. cylindrica
and in plants of cul-tivar Odesskaya semidwarf were revealed to preciselyassign the identified ISSR loci of introgressive lines5/55-91 and 5/20-91 to common wheat or to
Ae. cylin-drica.
In genotypes of individual plants belonging toline 5/55-91, 137 ISSR loci were found (Table 2), six ofwhich were attributed to introgressive loci, i.e., theybelong to
Ae. cylindrica
(Table 3). In individual plantsof line 5/20-91, 142 loci were found with seven ISSRprimers, and 17 of them were attributed to introgressiveloci. In total, 19 ISSR loci typical for
Ae. cylindrica
were detected in genotypes of 30 BC
1
F
9
plants belong-ing to two introgressive lines. Primer (AGC)
6
G (Table 3)seems to be most informative in terms of detecting
Table 2.
ISSR analysis of molecular–genetic polymorphism in analyzed plants
ISSR primer
Cultivar, line, form
Odesskaya semidwarf
Aegilops cylindrica
5/55-91 5/20-91
number of ISSR loci
detected polymor-phic detected polymor-
phic detected polymor-phic detected polymor-
phic
(AG)
9
C 28 11 29 13 28 11 28 7
(AGC)
6
G 20 6 23 11 24 18 28 16
(ACC)
6
G 14 7 12 5 16 8 15 5
(GAG)
6
C 11 2 16 4 11 3 11 2
(CTC)
6
A 23 9 23 13 21 6 26 14
(GA)
9
C 17 5 11 6 21 5 20 6
(CT)
9
G) 18 9 11 2 16 5 14 7
Total 131 49 122 54 137 56 142 57
Mean polymorphism, % 37.4 43.2 40.9 40.1
1374
RUSSIAN JOURNAL OF GENETICS
Vol. 40
No. 12
2004
GALAEV
et al
.
introgression, since due to the use of this primer, eightintrogressive ISSR loci were detected in plants of line5/20-91. In contrast, (AG)
9
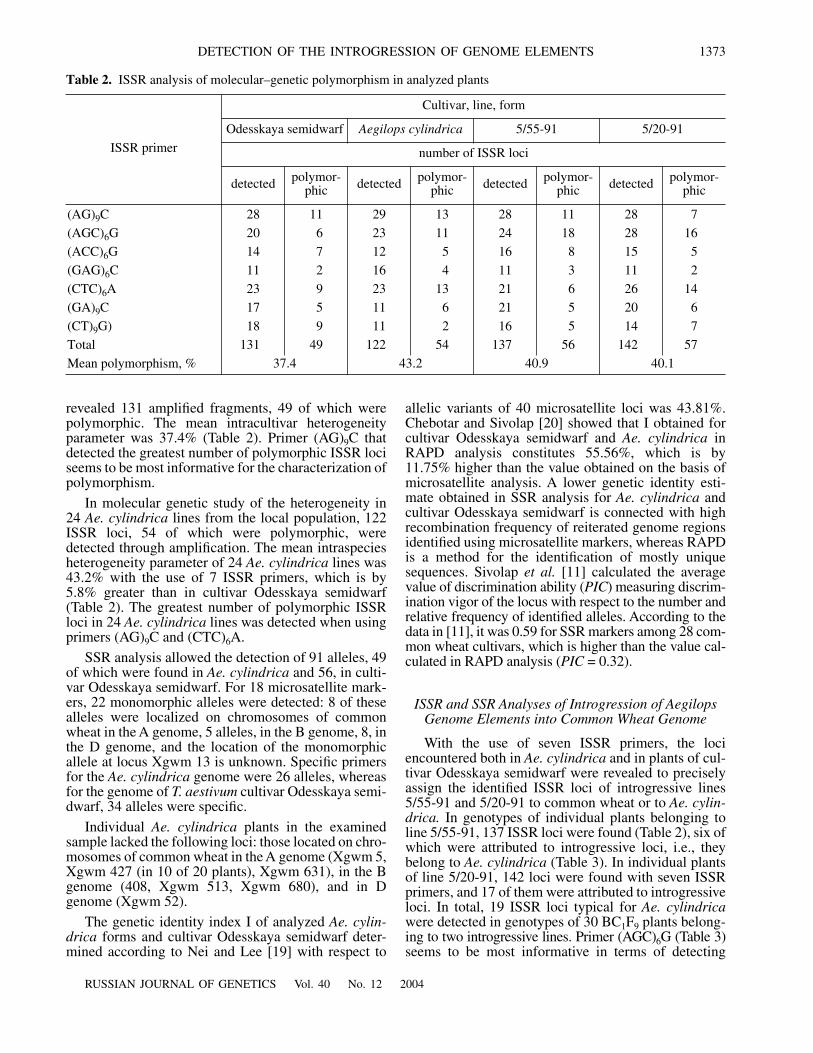
C is apparently the leastinformative primer, since no introgressive clone wasdetected in lines 5/55-91 and 5/20-91. All plants of line5/55-91 contained introgressive loci, having 3545- and823-bp MM (Fig. 1), which were detected with primer(ACC)
6
G. With primers (AGC)
6
G, (ACC)
6
G, and(GA)
9
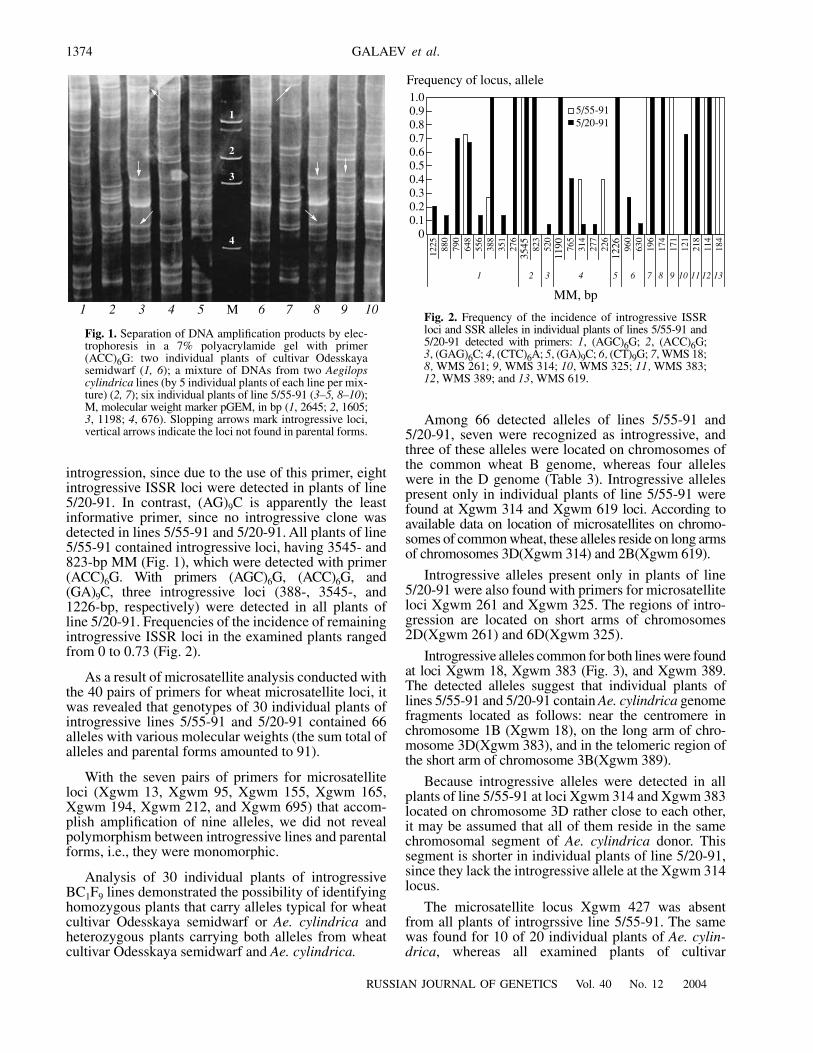
C, three introgressive loci (388-, 3545-, and1226-bp, respectively) were detected in all plants ofline 5/20-91. Frequencies of the incidence of remainingintrogressive ISSR loci in the examined plants rangedfrom 0 to 0.73 (Fig. 2).
As a result of microsatellite analysis conducted withthe 40 pairs of primers for wheat microsatellite loci, itwas revealed that genotypes of 30 individual plants ofintrogressive lines 5/55-91 and 5/20-91 contained 66alleles with various molecular weights (the sum total ofalleles and parental forms amounted to 91).
With the seven pairs of primers for microsatelliteloci (Xgwm 13, Xgwm 95, Xgwm 155, Xgwm 165,Xgwm 194, Xgwm 212, and Xgwm 695) that accom-plish amplification of nine alleles, we did not revealpolymorphism between introgressive lines and parentalforms, i.e., they were monomorphic.
Analysis of 30 individual plants of introgressiveBC
1
F
9 lines demonstrated the possibility of identifyinghomozygous plants that carry alleles typical for wheatcultivar Odesskaya semidwarf or Ae. cylindrica andheterozygous plants carrying both alleles from wheatcultivar Odesskaya semidwarf and Ae. cylindrica.
Among 66 detected alleles of lines 5/55-91 and5/20-91, seven were recognized as introgressive, andthree of these alleles were located on chromosomes ofthe common wheat B genome, whereas four alleleswere in the D genome (Table 3). Introgressive allelespresent only in individual plants of line 5/55-91 werefound at Xgwm 314 and Xgwm 619 loci. According toavailable data on location of microsatellites on chromo-somes of common wheat, these alleles reside on long armsof chromosomes 3D(Xgwm 314) and 2B(Xgwm 619).
Introgressive alleles present only in plants of line5/20-91 were also found with primers for microsatelliteloci Xgwm 261 and Xgwm 325. The regions of intro-gression are located on short arms of chromosomes2D(Xgwm 261) and 6D(Xgwm 325).
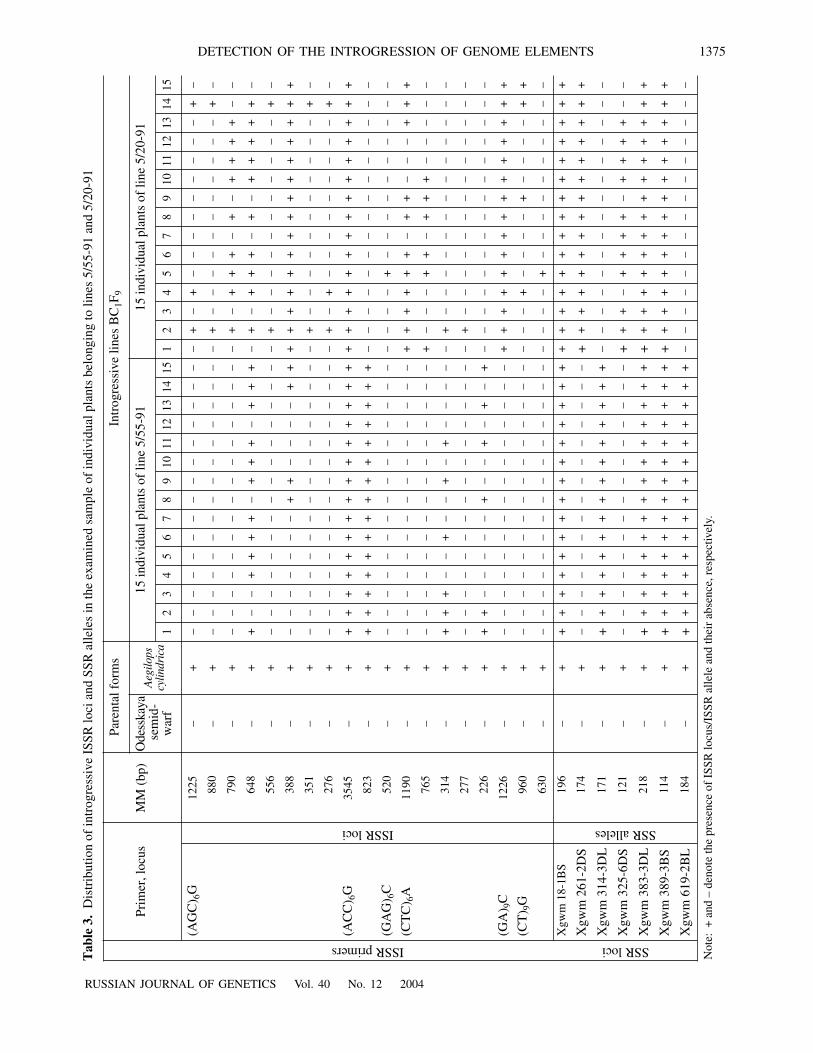
Introgressive alleles common for both lines were foundat loci Xgwm 18, Xgwm 383 (Fig. 3), and Xgwm 389.The detected alleles suggest that individual plants oflines 5/55-91 and 5/20-91 contain Ae. cylindrica genomefragments located as follows: near the centromere inchromosome 1B (Xgwm 18), on the long arm of chro-mosome 3D(Xgwm 383), and in the telomeric region ofthe short arm of chromosome 3B(Xgwm 389).
Because introgressive alleles were detected in allplants of line 5/55-91 at loci Xgwm 314 and Xgwm 383located on chromosome 3D rather close to each other,it may be assumed that all of them reside in the samechromosomal segment of Ae. cylindrica donor. Thissegment is shorter in individual plants of line 5/20-91,since they lack the introgressive allele at the Xgwm 314locus.
The microsatellite locus Xgwm 427 was absentfrom all plants of introgrssive line 5/55-91. The samewas found for 10 of 20 individual plants of Ae. cylin-drica, whereas all examined plants of cultivar
1 2 3 4 5 6 7 8 9 10M
1
2
3
4
Fig. 1. Separation of DNA amplification products by elec-trophoresis in a 7% polyacrylamide gel with primer(ACC)6G: two individual plants of cultivar Odesskayasemidwarf (1, 6); a mixture of DNAs from two Aegilopscylindrica lines (by 5 individual plants of each line per mix-ture) (2, 7); six individual plants of line 5/55-91 (3–5, 8–10);M, molecular weight marker pGEM, in bp (1, 2645; 2, 1605;3, 1198; 4, 676). Slopping arrows mark introgressive loci,vertical arrows indicate the loci not found in parental forms.
1225 88
079
064
855
638
835
127
635
45 823
520
1190 76
531
427
722
612
26 960
630
196
174
171
121
218
114
184
1.00.90.80.70.60.50.40.30.20.1
0
1 2 3 4 5 6 7 8 9 10 11 12 13
MM, bp
Frequency of locus, allele
5/55-915/20-91
Fig. 2. Frequency of the incidence of introgressive ISSRloci and SSR alleles in individual plants of lines 5/55-91 and5/20-91 detected with primers: 1, (AGC)6G; 2, (ACC)6G;3, (GAG)6C; 4, (CTC)6A; 5, (GA)9C; 6, (CT)9G; 7, WMS 18;8, WMS 261; 9, WMS 314; 10, WMS 325; 11, WMS 383;12, WMS 389; and 13, WMS 619.
RUSSIAN JOURNAL OF GENETICS Vol. 40 No. 12 2004
DETECTION OF THE INTROGRESSION OF GENOME ELEMENTS 1375
Tab
le 3
. D
istr
ibut
ion
of in
trog
ress
ive
ISSR
loci
and
SSR
alle
les
in th
e ex
amin
ed s
ampl
e of
indi
vidu
al p
lant
s be
long
ing
to li
nes
5/55
-91
and
5/20
-91
Pr
imer
, loc
usM
M (
bp)
Pare
ntal
for
ms
Intr
ogre
ssiv
e lin
es B
C1F
9
Ode
sska
ya s
emid
-w
arf
Aeg
ilops
cylin
dric
a
15 in
divi
dual
pla
nts
of li
ne 5
/55-
9115
indi
vidu
al p
lant
s of
line
5/2
0-91
12
34
56
78
910
1112
1314
151
23
45
67
89
1011
1213
1415
ISSR primers
(AG
C) 6
G
ISSR loci
1225
–+
––
––
––
––
––
––
––
––
+–
+–
––
––
––
––
+–
880
–+
––
––
––
––
––
––
––
––
+–
––
––
––
––
––
+–
790
–+
––
––
––
––
––
––
––
––
+–
++
+–
+–
++
++
––
648
–+
+–
–+
++
+–
++
+–
++
+–
+–
++
+–
+–
++
++
+–
556
–+
––
––
––
––
––
––
––
––
+–
––
––
––
––
––
+–
388
–+
––
––
––
–+
+–
––
–+
++
++
++
++
++
++
++
++
351
–+
––
––
––
––
––
––
––
––
+–
––
––
––
––
––
+–
276
–+
––
––
––
––
––
––
––
––
+–
+–
––
––
––
––
+–
(AC
C) 6
G35
45–
++
++
++
++
++
++
++
++
++
++
++
++
++
++
++
+
823
–+
++
++
++
++
++
++
++
+–
––
––
––
––
––
––
––
(GA
G) 6
C52
0–
+–
––
––
––
––
––
––
––
––
––
+–
––
––
––
––
–
(CT
C) 6
A11
90–
+–
––
––
––
––
––
––
––
++
++
++
–+
+–
––
++
+
765
–+
––
––
––
––
––
––
––
–+
––
–+
+–
++
+–
––
––
314
–+
++
+–
–+
––
+–
+–
––
––
+–
––
––
––
––
––
––
277
–+
––
––
––
––
––
––
––
––
+–
––
––
––
––
––
––
226
–+
++
––
––
–+
––
+–
+–
+–
––
––
––
––
––
––
––
(GA
) 9C
1226
–+
––
––
––
––
––
––
––
–+
++
++
++
++
++
++
++
(CT
) 9G
960
–+
––
––
––
––
––
––
––
––
––
+–
––
–+
––
––
++
630
–+
––
––
––
––
––
––
––
––
––
–+
––
––
––
––
––
SSR loci
Xgw
m 1
8-1B
S
SSR alleles
196
–+
++
++
++
++
++
++
++
++
++
++
++
++
++
++
++
Xgw
m 2
61-2
DS
174
–+
––
––
––
––
––
––
––
–+
++
++
++
++
++
++
++
Xgw
m 3
14-3
DL
171
–+
++
++
++
++
++
++
++
+–
––
––
––
––
––
––
––
Xgw
m 3
25-6
DS
121
–+
––
––
––
––
––
––
––
–+
++
–+
++
+–
++
++
––
Xgw
m 3
83-3
DL
218
–+
++
++
++
++
++
++
++
++
++
++
++
++
++
++
++
Xgw
m 3
89-3
BS
114
–+
++
++
++
++
++
++
++
++
++
++
++
++
++
++
++
Xgw
m 6
19-2
BL
184
–+
++
++
++
++
++
++
++
+–
––
––
––
––
––
––
––
Not
e:+
and
– d
enot
e th
e pr
esen
ce o
f IS
SR lo
cus/
ISSR
alle
le a
nd th
eir
abse
nce,
res
pect
ivel
y.

1376
RUSSIAN JOURNAL OF GENETICS Vol. 40 No. 12 2004
GALAEV et al.
Odesskaya semidwarf and line 5/20-91 contained amicrosatellite allele having 204-bp MM at this locus.This fact may be explained by the introgression of anAe. cylindrica genome region lacking locus Xgwm 427into wheat chromosome 6A. The translocation site islocated in the telomeric region of the long arm of chro-mosome 6A.
Analyzing individual plants of lines 5/55-91 and5/20-91 by ISSR and SSR methods, we detected ISSRloci and SSR alleles, which were not found in the exam-ined sample of parental forms. Thus, with ISSR primers(AG)9C, (AGC)6G, (ACC)6G (Fig. 1), and (GA)9C,7 new loci were detected. Nine alleles, which are non-typical for parental forms, were found at nine microsat-ellite loci (Xgwm 3, Xgwm 160, Xgwm 169, Xgwm192, Xgwm 325, Xgwm 408, Xgwm 437, Xgwm 577,and Xgwm 619). The seven detected ISSR loci and nineSSR alleles are probably connected with the reorgani-zation of parental genomes in the genome of hybridplants occurring under the influence of distant hybrid-ization. Resistance of lines 5/55-91 and 5/20-91 to fun-gal diseases may have occurred not only from intro-gression of the Ae. cylindrica genome site carrying newresistance genes but also due to chromosomal rear-rangements caused by distant hybridization. Therefore,these loci and alleles must be considered in the searchfor resistance markers.
A 191-bp allele of Xgwm 192 locus detected inplants belonging to line 5/20-91, which is absent in theexamined sample of parental forms, was shown byRoder and coworkers [17] to be present in commonwheat variety Chinese Spring. Meanwhile, a 171-bpintrogressive allele (Xgwm 314) detected by us wasfound in synthetic amphihexaploid wheat (obtainedafter crossing T. tauschii (DD) with T. durum (AABB)).According to Chebotar’ and Sivolap [21], commonwheat cultivars Odom and Mironovskaya 808 contain a174-bp allele (Xgwm 261) detected by us in all plantsof line 5/20-91. This suggests that the above allele is notan introgressive element of the Ae. cylindrica genome,because common wheat cultivars contain this allele.
The examined introgressive lines 5/55-91 and 5/20-91have a high level of complex resistance to a number of
cereal diseases transferred from Ae. cylindrica to thesusceptible cultivar Odesskaya semidwarf [7, 8] as pos-sible sources of resistance gene (genes) marker. Thismotivated a great deal of interest in studying the intro-gressive ISSR loci and SSR alleles that are character-ized by the highest frequency of the incidence in theexamined sample of individual plants belonging to bothlines. According to the constructed histogram (Fig. 2),the following introgressive loci and alleles had thehighest frequency of the incidence in plants: ISSR lociwith MM of 648 bp (primer (AGC)6G) and 3545 bp(primer (ACC)6G), and SSR alleles of 196 bp (Xgwm18), 218 bp (Xgwm 383), and 114 bp (Xgwm 389).
Thus, the results of ISSR and SSR analyses demon-strated the following changes in the genome of distanthybrids belonging to lines 5/55-91 and 5/20-91: (1) lossof amplification fragments typical for one of the parents(cultivar Odesskaya semidwarf and Ae. cylindrica);(2) the appearance in the amplification spectrum of newfragments nontypical for both parents; (3) a completechange in the spectrum of fragment distribution (thepresence in the amplification spectrum of only a newfragment in the absence of fragments typical for par-ents).
The changes revealed in A and B genomes of indi-vidual plants belonging to lines 5/55-91 and 5/20-91,which were connected with introgression of Ae. cylin-drica genome elements allow us to draw the conclusionabout homeologous conjugation with participation ofthe D genome and, possibly, C genome of Ae. cylin-drica in the examined hybrids.
ACKNOWLEDGMENTS
The authors are grateful to A.I. Rybalka for supply-ing experimental material and to S.V. Chebotar for pro-viding SSR primers.
REFERENCES1. Innes, R.L. and Kerber, E.R., Resistance to Wheat Leaf
Rust and Stem Rust in Triticum tauschii and Inheritancein Hexaploid Wheat of Resistance Transferred fromTriticum tauschii, Genome, 1994, vol. 37, pp. 813–822.
2. Bonhomme, A., Gale, M.D., Koebner, R.M.D., et al.,RFLP Analysis of an Aegilops ventricosa ChromosomeThat Carries a Gene Conferring Resistance to Leaf Rust(Puccinia recondita) When Transferred to HexaploidWheat, Theor. Appl. Genet., 1995, vol. 90, pp. 1042–1048.
3. Harjit-Singh, Tsujimoto, H., Sakhuja, P.K., et al., Trans-fer of Resistance to Wheat Pathogens from Aegilops tri-uncialis into Bread Wheat, Wheat Inf. Serv., 2000,no. 91, pp. 5–20.
4. Aghaee-Sarbarzeh, M., Dhaliwal, H.S., Chhuneja, P.,and Harjit-Singh, Suppression of Resistance Genes fromDistantly Related Species in Triticum durum–AegilopsAmphiploids, Wheat Inf. Serv., 2001, no. 92, pp. 12–16.
5. Babayants, L.T., Ribalka, O.Z., and Aksel’rud, D.V.,A New Source of Wheat Resistance to Major Diseases, in
1 2 3 4 5 6 7 8 9 10 11 12M
Fig. 3. Separation of DNA amplification products by elec-trophoresis in agarose gel with primers for microsatellitelocus Xgwm 383: cultivar Odesskaya semidwarf (1–3); Ae.cylindrica (4–6); line 5/55-91 (7–9); line 5/20-91; (10–12);M, molecular weight marker λ/PstI. Slopping arrows markintrogressive loci.

RUSSIAN JOURNAL OF GENETICS Vol. 40 No. 12 2004
DETECTION OF THE INTROGRESSION OF GENOME ELEMENTS 1377
Realizatsiya potentsiinikh mozhlivostei sortiv i gibridivSelektsiino-genetichnogo institutu v umovakh Ukraïni(Realization of the Potential of Cultivars and Hybrids ofthe Institute of Breeding and Genetics under Conditionsof Ukraine), Odessa, 1996, pp. 111–116.
6. Babayants, L.T., Dubinina, L.A., and Yushchenko, G.M.,Identification of the Resistance Genes That ConferResistance to Tilletia caries (DC) Tull. and Are Nonal-lelic to Known Ones in Wheat Lines by InterspecificHybridization Triticum aestivum × Aegilops cylindrica,Tsitol. Genet., 2000, vol. 34, no. 4, pp. 32–40.
7. Babayants, L.T., Vasil’ev, A.A., and Novitskaya, N.A.,The Genetic Basis of Resistance to Puccinia reconditaRob. ex Desm. f. sp. tritici in Interspecific WheatHybrids, Tsitol. Genet., 1998, vol. 32, no. 6, pp. 20–26.
8. Sechnyak, A.L. and Simonenko, V.K., Cytogenetics ofHybrids of Durum Wheat Triticum durum Desf. andAegilops caudate L., Tsitol. Genet., 1999, vol. 33, no. 4,pp. 49–55.
9. Avsenin, V.I., Motsnyi, I.I., Rybalka, A.I., and Fait, V.I.,Hybrids of Aegilops cylindrica Host. With Triticumdurum Desf. and T. aestivum L., Tsitol. Genet., 2003, no. 1,pp. 11–17.
10. Bochev, B., Kunovski, Zh., and Ganeva, G., The GenusAegilops L. as a Source of Fungal Diseases in WheatBreeding, Tr. Prikl. Bot., Genet. Selekts. VNII Raste-nievod., 1982, vol. 73, no. 3, pp. 111–120.
11. Sivolap, Yu.M., Chebotar’, S.V., Topchieva, E.A., et al.,RAPD and SSRP Analyses of Molecular-Genetic Poly-morphism in Triticum aestivum L. Cultivars, Genetika(Moscow), 1999, vol. 35, no. 12, pp. 1665–1673.
12. Plachke, J., Ganal, M.W., and Roder, M.S., Detection ofGenetic Diversity in Closely Related Bread Wheat UsingMicrosatellite Markers, Theor. Appl. Genet., 1995, vol. 91,pp. 1001–1007.
13. Zietkiewicz, E., Rafalski, A., and Labuda, D., GenomeFingerprinting by Simple Sequence Repeat (SSR)-Anchored Polymerase Chain Reaction Amplification,Genomics, 1994, vol. 20, pp. 176–183.
14. Tsumura, Y., Ohba, K., and Strauss, S.H., Diversity andInheritance of Inter-Simple Sequence Repeat Polymor-phisms in Douglas Fir (Pseudotsuga menziesii) and Sugi(Cryptomeria japonica), Theor. Appl. Genet., 1996,vol. 92, pp. 40–45.
15. Wu, K.-S., Jones, R., Danneberger, L., and Scolnik, P.A.,Detection of Microsatellite Polymorphisms withoutCloning, Nucleic Acids Res., 1994, vol. 22, pp. 3257–3258.
16. Sivolap, Yu.M., Ispol’zovanie PTsR-analiza v genetiko-selektsionnykh issledovaniyakh (nauchno-metodicheskoerukovodstvo) (PCR-Based Analyses in Studies on Geneticsand Breeding (Methodical Recommendations)), Kiev:Agrarna Nauka, 1998.
17. Roder, M.S., Korzun, V., Plachke, J., et al., A Microsat-ellite Map of Wheat, Genetics, 1998, vol. 149, pp. 2007–2023.
18. Kuts, O.O., Chebotar’, S.V., Sivolap, Yu.M., andTots’kii, V.M., Molecular Genetic Polymorphism ofTriticum aestivum L. as Revealed by Inter-SSR PCR,Visnik Odessk. D. Univ., 2000, vol. 5, no. 1, pp. 97–102.
19. Nei, M. and Li, W.-H., Mathematical Model for StudyingGenetics Variation in Terms of Restriction Endonucleases,Proc. Natl. Acad. Sci. USA, 1979, vol. 76, pp. 5269–5273.
20. Chebotar’, S.V., Molecular Genetic Analysis of Intro-gression of Aegilops Genome Elements into the Com-mon Wheat Genome, Cand. Sci. (Biol.) Dissertation,Odessa, 1995.
21. Chebotar’, S.V. and Sivolap, Yu.M., Differentiation,Identification, and Database Construction of UkrainianT. aestivum L. Cultivars on the Basis of STMS Analysis,Tsitol. Genet., 2001, vol. 35, no. 6, pp. 18–27.





























