Embed Size (px)
Citation preview

Ana Clara Bragada Sequeira
2014
DEPARTAMENTO DE CIÊNCIAS DA VIDAFACuldAde de CiênCiAS e TeCnologiA
univerSidAde de CoimBrA
Treadmill exercise exerts an antidepressant-like effect to mice submitted to a neurotoxic
methamphetamine dose
Dissertação apresentada à Universidade de Coimbra para cumprimento dos requisitos necessários à obtenção do grau de Mestre em Bioquímica, realizada sob a orientação científica do Professor Doutor Frederico Pereira (Universidade de Coimbra) e do Professor Doutor Rui de Carvalho (Universidade de Coimbra)

This work was developed in the following institution:
Pharmacology and Experimental Therapeutics, Institute for Biomedical Imaging and Life
Sciences, Faculty of Medicine, University of Coimbra, Coimbra


Agradecimentos
É com grande prazer que aqui expresso, a mais profunda gratidão a todos
aqueles que tornaram este trabalho possível.
Esta tese não teria sido realizada sem a ajuda, apoio e paciência do meu
principal orientador Prof. Dr. Frederico Pereira. Quero agradecer-lhe por
me ter recebido no seu laboratório, pela liberdade que me foi dada para a
execução de todos os experimentos, pelos seus esforços para me motivar
nesta pesquisa, por abrir a minha mente, pelo seu entusiasmo e constante
bom humor e, acima de tudo, pela amizade.
Os bons conselhos, apoio e amizade do meu segundo orientador, Prof.
Dr. Rui de Carvalho, têm sido de grande valor tanto a nível académico
como a nível pessoal, pelo qual sou extremamente grata.
Quero agradecer ao Prof. Doutor Carlos Fontes Ribeiro por me ter
acolhido no Laboratório de Farmacologia e Terapêutica Experimental da
Faculdade de Medicina da Universidade de Coimbra.
Também quero agradecer a todos os companheiros e colegas de
laboratório, por todos os bons momentos e companheirismo. Um
agradecimento especial à Cristina Lemos, Sofia Viana, Doutora Catarina
Gomes e também ao Filipe Matheus e Prof. Dr. Rui Prediger
(Universidade Federal de Santa Catarina-USFC-Florianópolis, Santa
Catarina, Brasil) por todo o apoio, dicas e paciência demonstrada em
todos os ensinamentos, que foram fundamentais para realizar todo o meu
trabalho. Para a minha colega Inês Pita, por toda a ajuda, apoio e
momentos de alegria.
Agradeço às minhas amigas e colegas - Raquel Fonseca e Jani Almeida -
por todos os momentos passados na vossa companhia. Obrigado por
tudo, obrigado pela vossa amizade.
Obrigada a todos os meus amigos, que me têm incentivado, que me
apoiaram nos momentos difíceis, e que foram brilhantes e
compreensivos quando eu precisei que eles o fossem.
Por último, mas não menos importante, gostaria de expressar a minha
mais sentida gratidão à minha família. Nada disto teria sido possível sem
o amor e a paciência dos meus pais e irmã, que têm sido uma fonte de
amor constante, preocupação, apoio e força em todos estes anos.
Obrigada por terem fé em mim e por me terem incentivando em cada
decisão que tomei na minha vida, sempre acreditando nas minhas
escolhas que me levaram até onde estou agora.
Aos meus avós, e ao resto da minha família, pelo amor, amizade, alegria
e todos os momentos experienciados que fizeram tudo valer a pena.
Finalmente, quero agradecer meu namorado Ricardo, pelo seu amor,
amizade, apoio e toda a felicidade que me proporcionou ao longo destes

anos. Obrigada por acreditares em mim, mesmo quando eu duvidei
de mim mesma. Adoro-te!
A todos aqueles que tornaram isto possível!

i
Index
Abbreviations iii
List of Figures v
List of Tables vii
Resumo viii
Abstract x
CHAPTER 1 1
INTRODUCTION AND AIMS 1
1. Drug Addiction 2
1.1 Amphetamine-type stimulants 3
1.2 Epidemiology of abuse amphetamine-type stimulants 4
1.3 Methamphetamine and Serotonin 7
1.3.1. Methamphetamine: pharmacokinetics and pharmacodynamics 7
1.3.2. Serotonin Biosynthesis 9
1.3.3. 5-HT Receptors 14
1.3.4. Methamphetamine action in serotonergic systems 15
2. Depression 18
2.1. Depression: Focusing on 5-HT 19
2.2. Depressive behaviour during withdrawal from METH 20
3. Physical Exercise 22
3.1. Physical Exercise: a non-pharmacological approach in depressive disorders 22
3.2. Physical Exercise: managing METH addiction and neurotoxicity 23
4. Aims of this thesis 25
CHAPTER 2 26
MATERIAL AND METHODS 26
1. Animals 27
2. Drugs and Chemicals 27
3. Experimental Design 28
3.1. Treadmill adaptation 28
3.2. Methamphetamine administration 29
3.3. Exercise protocol 31
4. Behavioural Tests 32
4.1. Open-field test 33
4.2. Splash test 34

ii
4.3. Forced-swim test 35
5. Frontal Cortex and Hippocampus Isolation 36
6. Determination of 5-HT by HPLC 36
7. Statistical analysis 38
CHAPTER 3 39
RESULTS 39
1. Effects of methamphetamine and/or chronic physical exercise on body weight 40
2. Effect of treadmill exercise on general exploratory locomotion in METH-exposed
mice 41
2.1. Open-field test 41
3. Effect of treadmill exercise on depressive-like behavior in METH-exposed mice43
3.1. Splash test 43
3.2. Forced-swim test 44
4. Effect of treadmill exercise on 5-HT total content in frontal cortex and
hippocampi from METH-exposed mice 45
CHAPTER 4 46
Discussion 47
CHAPTER 5 51
Conclusion 52
CHAPTER 6 53
References 54

iii
Abbreviations
5-HIAA – Hydroxyindoleacetic acid
5-HT – 5-hydroxytryptamine; serotonin
5-HTP – 5-hyfroxytryptophan
AADC – L-amino acid decarboxylase
AADC – L-aromatic amino acid decarboxylase
ACTH – Adrenocorticotropic hormone
ADHD – Attention deficit hyperactivity disorder
AMPH – Amphetamine
ATS – Amphetamine-type stimulants
BBB – Blood brain barrier
BDNF – Brain-derived neurotrophic factor
BH4 – Tetrahydrobiopterin
CNS – Central nervous system
Cont – Control
CREB – cAMP response element-binding protein
CRF – Corticotropin-releasing factor
DA – Dopamine
DAT – Dopamine transporter
DOPAC – 3,4 – dihydroxyphenylacetic acid
ENS – Enteric nervous system
FST – Forced-swim test
GDNF – Glial cell–derived neurotrophic factor
HPA – Hypothalamic-pituitary-adrenal
HPLC – High pressure liquid chromatography
HVA – Homovanillic acid
IBS – Irritable bowel syndrome
L-TRP- L-tryptophan
MAO – Monoamine oxidase
MAO-A – Monoamine oxidase A
MAOIs – Monoamine oxidase inhibitors
MDD – Major depressive disorder
MDD – Major depressive disorder

iv
MDMA – 3-4-methylenedioxymethamphetamine
METH – Methamphetamine
NA – Norepinephrine
Nac – Nucleus accumbens
OF – Open-field test
PFC – Prefrontal cortex
PNS – Peripheral nervous system
SAL – Saline
SED – Sedentary
SERT – Serotonin transporter
SNS – Sympathetic nervous system
SSRIs – Serotonin-specific reuptake inhibitor
TCA – Tricyclic antidepressants
TH – Tyrosine hydroxylase
TPH – Tryptophan hydroxylase
TPH2 – Tryptophan hydroxylase 2
UNODC – United Nations Office on Drugs and Crime
VMAT2 – Vesicular monoamine transporter 2
VTA – Ventral tegmental area
WHO – World Health Organization

v
List of Figures
Figure 1 – Neurocircuitry schematic illustrating the combination of neuroadaptations in
the brain circuitry for the three stages of the addiction cycle that promote drug-seeking
behavior in the addicted state. 2
Figure 2 – Trends in the prevalence of different drugs, 2009-2011. 6
Figure 3 – Use of ATS in 2011 (or latest year available). 6
Figure 4 – Chemical structure of methamphetamine (1), as well as the closely related
psychostimulant d-amphetamine (2). 8
Figure 5 – Synthesis and metabolism of 5-HT. 11
Figure 6 – 5-HT pathways. 13
Figure 7 – The impact of METH on a serotonergic synapse. 16
Figure 8 – Example of a C57BL/6 mice. 27
Figure 9 – Mice adaptation to treadmill protocol. 29
Figure 10 – Intraperitoneal administration of a single dose of METH (30 mg/kg). 30
Figure 11 – Physical exercise protocol for the respective exercise groups, SAL/EX and
METH/EX. 31
Figure 12 – Training of the mice belonging to exercise group on the treadmill, separarated
by acrylic divisions, providing four individual lanes to run. 32
Figure 13 – Mice in open-field test apparatus. 33
Figure 14 – Splash test: A - mice in the cage 2 is grooming his dorsal coat in his home
cage; B - schematic illustration of dorsal mice grooming. 34
Figure 15 – Forced-swim test. Mice placed into an open cylindrical container. 36
Figure 16 – HPLC system used to quantify 5-HT in this study. 37
Figure 17 – Chromatogram illustrating a 5-HT peak relative to a frontal cortex sample
from a METH/EX mice group. 37
Figure 18 – A: Evolution of the mice body-weight over 61 days (total duration of
experimental protocol). B: Mice body-weight at 3 different time-points: day 1, begining of
training; day 14: METH administration; day 61: last day of physical exercise program.
40
Figure 19 – Effect of treadmill exercise on the locomotor activity and exploratory
behavior (open field test) in mice injected with a single dose of METH (30 mg/kg, i.p.).
41

vi
Figure 20 – Effect of treadmill exercise on grooming behavior (splash test) in mice
injected with a single dose of METH (30 mg/kg, i.p.). 43
Figure 21 – Effect of treadmill exercise on the depair-like behavior (forced swimming
test) in mice injected with a single dose of METH (30 mg/kg, i.p.). 44
Figure 22 – Effect of treadmill exercise on hippocampal (A,B) and frontal cortical (C)
serotonin levels (5-HT measured from mice at 3 (A) and 49 days (B, C) post-METH
injection). 45

vii
List of Tables
Tabel 1 – Drug Consumption in 2010, (UNODC). 5
Tabel 2 – Experimental groups and number of used mice (n). 28

viii
Resumo
Demonstrámos recentemente, que uma dose única e elevada de
metanfetamina (METH) causou um fenótipo depressivo de longa duração, aferido pelo
aumento do tempo de imobilidade no teste de tail suspension. Tem sido demonstrado que o
exercício físico alivia os sintomas depressivos.
Propomo-nos confirmar e caracterizar mais detalhadamente o fenótipo depressivo
causado por uma única injecção neurotóxica de METH e investigar a correlação entre a
homeostase de 5-HT no córtex frontal e hipocampo e o efeito antidepressivo de exercício
físico.
Murganhos adultos C57BL/6 foram submetidos a um programa de exercício em
treadmill (cinco dias por semana durante sete semanas) a partir de 24 h após uma única
dose elevada de METH (30 mg/kg, i.p.), tendo sido organizados da seguinte forma
(n=8/grupo): salino/sedentário; METH/sedentário; salino/exercício e METH/exercício. Em
seguida, avaliou-se a exploração/actividade locomotora, anedonia e desespero
(comportamentos do tipo depressivo) numa fase tardia da privação de METH (49 dias)
através dos testes open-field, splash e forced-swim, respectivamente. Os teores totais de 5-
HT no cortex-frontal e hipocampo (49 dias pós-METH) foram avaliados por cromatografia
líquida de fase reversa e de alta eficiência (HPLC). Avaliámos o impacto inicial (3 dias
pós-METH) de METH nos níveis totais de 5-HT do hipocampo noutro grupo de
murganhos (16).
É importante registar que a METH diminuiu o tempo de grooming dos murganhos
sedentários, no teste splash. Isto demonstra que esta dose de METH induz um
comportamento do tipo anedónico. Por outro lado, confirmou-se ainda que a METH induz
um comportamento do tipo depressivo de longa duração. Além disso, o exercício em
treadmill reverteu o tempo de grooming no teste splash, 7 semanas após a injecção de
METH. Este efeito antidepressivo foi ainda aferido por uma diminuição do tempo de
imobilidade e um aumento do tempo de natação no teste forced-swim exibido pelos
murganhos, control ou submetidos à METH. Adicionalmente, o exercício aumentou a
actividade locomotora, o comportamento exploratório e a razão distância central - distância
total nos murganhos control ou submetidos à METH no teste open-field. Isto é consistente
com o facto de o exercício físico induzir um estado de bem-estar emocional,
independentemente do tratamento farmacológico. Por outro lado, a METH não alterou
significativamente os parâmetros motores de murganhos sedentários. Isto é relevante, visto

ix
que sugere que a actividade locomotora não contribuiu para o comportamento do tipo
anedónico observado.
A análise neuroquímica mostrou que a METH provocou uma deplecção robusta de
5-HT no hipocampo nos primeiros 3 dias após a injecção de METH, que foi recuperada 49
dias após o tratamento. Além disso, o exercício físico não teve um impacto significativo
nos níves hipocampais de 5-HT. Adicionalmente, nem o exercício físico nem a METH
alteraram significativamente os teores toatis de 5-HT.
É importante destacar, que demostrámos que um programa de sete semanas de
exercício em treadmill promoveu um efeito antidepressivo em murganhos intoxicados com
METH. No entanto, este benefício do exercício não está associado a altenações na
homeostase serotonérgica fronto-cortical/hipocampal.
Palavras-chave: metanfetamina, neurotoxicidade, depressão, serotonina, exercício
físico, córtex frontal, hipocampo, murganho.

x
Abstract
Recently we demonstrated that a single high neurotoxic dose of methamphetamine
(METH) caused a long-lasting depressive phenotype as gauged by increased immobility
time in the tail suspension test. It has been shown that physical exercise alleviates
depressive symptoms.
This study aims to further characterize the long-term depressive phenotype caused
by a single neurotoxic METH-injection and to probe for a correlation between frontal-
cortical and hippocampal 5-HT homeostasis and the antidepressant effect of physical
exercise.
Adult C57BL/6 mice were submitted to a treadmill exercise program (five days a
week for seven weeks) starting 24 h post-single high dose of METH (30 mg/kg, i.p.) and
were organized as follows (n=8/group): saline/sedentary; METH/sedentary; saline/exercise
and METH/exercise. Then, we assessed the exploration/locomotor activity, anhedonia and
despair (depressive-like behaviours) at a late stage of METH withdrawal (49 days) by
open-field, splash and forced-swim tests, respectively. Frontal cortical and hippocampal
(49 days post-METH) levels of 5-HT were evaluated by reversed-phase high performance
liquid chromatography (HPLC). Another set of mice (16) was used to evaluate early
impact (3 days post-METH) of METH on 5-HT hippocampal levels.
Remarkably METH decreased the time that sedentary mice spent grooming. This
shows that this METH dose induces anhedonia-like behavior. This further confirms that
METH induces a long-lasting depressive-like behaviour. Remakably treadmill exercise
reversed the time mice spent grooming in the splash test 7 weeks post-injection. This
antidepressant-like effect was further gauged by a decreased immobility time and increased
swimming time in the forced swimming test exhibited by both saline- and METH-
exercised animals. Additionally, exercise increased locomotor activity, exploratory
behavior and the central distance – total distance ratio in both saline and METH treated
mice in the open field. This is consistent with physical exercise inducing a high-mood
state, irrespectively of drug treatment. On the other hand, METH did not alter significantly
the motor parameters from sedentary mice. This is relevant since locomotor activity did not
contribute to the anhedonic-like behavior seen herein. The neurochemical analysis showed
that METH triggered a robust 5-HT hippocampal depletion as early as 3 days post-METH,
which recovered 49 days post-tretament. Additionally, physical exercise failed to have any

xi
significant impact on hippocampal 5-HT levels. Moreover, neither METH nor physical
exercise altered frontal cortical 5-HT levels.
Importantly, we present new evidence that a 7 weeks treadmill program provided
an antidepressant effect to METH-intoxicated mice. This exercise benefit is not associated
with changes in the frontal cortical/hippocampal serotonergic homeostasis.
Keywords: methamphetamine, neurotoxicity, depression, serotonin, physical exercise,
frontal cortex, hippocampus, mice.

CHAPTER 1
INTRODUCTION AND AIMS

CHAPTER 1 – Introduction and Aims
2
1. Drug Addiction
Drug addiction is a major health and social issue due to high rates of morbidity and
mortality that are accompanied by bouts of violence and legal problems, thus imposing
burdening individual and societal costs (Al-Haggar, 2014). This medical and societal
problem is considered to be a chronic, relapsing brain disorder. Particulalrly, brain
addiction is envisaged as a malfunction of the brain’s hedonic tone regulation and the
motivational system (Müller & Homberg, 2014). This psychiatric disorder is characterized
by compulsive drug seeking, by their continued use despite serious negative
socioeconomic and health consequences and marked withdrawal symptoms (Cami & Farré,
2003; National Institute on Drug Abuse (NIDA, 2012); Koob & Volkow, 2010 and
McAvoy, 2009).
Collapsing the cycles of impulsivity and compulsivity yields a composite addiction
cycle composed of three stages-binge/intoxication, withdrawal/negative affect,
preoccupation/anticipation-in which impulsivity often dominates at the early stages and
impulsivity combined with compulsivity dominates at the later stages (Koob & Volkow,
2010). As an individual moves from impulsivity to compulsivity, a shift occurs from
positive reinforcement driving the motivated behavior to negative reinforcement and
automaticity driving the motivated behavior. These three stages are conceptualized as
interacting with each other, becoming more intense, and ultimately leading to the
pathological state known as addiction (Figure 1).

CHAPTER 1 – Introduction and Aims
3
Figure 1 – Neurocircuitry schematic illustrating the combination of neuroadaptations in the brain circuitry
for the three stages of the addiction cycle that promote drug-seeking behavior in the addicted state. Note the
activation of the ventral striatum/dorsal striatum/extended amygdala driven by cues through the hippocampus
and basolateral amygdala and stress through the insula. The frontal cortex system is compromised, producing
deficits in executive function and contributing to the incentive salience of drugs compared to natural
reinforcers. Dopamine systems are compromised, and brain stress systems such as CRF are activated to reset
further the salience of drugs and drug-related stimuli in the context of an aversive dysphoric state (taken from
Koob & Volkow, 2010).
The binge–intoxication stage of the addiction cycle dependes upon the reinforcing
effects of drugs that may engage reward neurotransmitters (including dopamine and opioid
peptides) and associative mechanisms in the nucleus accumbens shell and core and then
engage stimulus–response habits that depend on the dorsal striatum (Koob & Volkow
2010).
The preoccupation–anticipation (craving) stage involves key glutamatergic
projections to the extended amygdala and nucleus accumbens from the prefrontal cortex
(for drug-induced reinstatement of drug seeking) and from the basolateral amygdala (for
cue-induced reinstatement of drug seeking) (Kalivas et al., 2003). Compulsive drug-
seeking behaviour is thought to engage ventral striatal–ventral pallidal–thalamic–cortical
loops that could subsequently engage dorsal striatal–dorsal pallidal–thalamic–cortical
loops (Vanderschuren et al., 2005), both of which are exaggerated by concomitant
decreased activity in reward circuits (Koob et al., 2005, 2008).
The neural substrates and neuropharmacological mechanisms for the negative
motivational effects of the withdrawal–negative affect stage of the addiction cycle may
involve not only disruption of the neural systems implicated in the positive reinforcing
effects of drugs, but also recruitment of brain stress systems (Koob et al., 2008).
Importantly, a new functional model also suggests that specific adaptations in the 5-
HT system render the nervous system susceptible to the transition to compulsive drug use
behaviours (Müller & Homberg, 2014).
1.1 Amphetamine-type stimulants
Amphetamine (1-methyl-2-phenethylamine) (AMPH), which is the first member of
a group of compounds that have similar structures and biological properties and are
collectively called “amphetamine-type stimulants” (ATS), was first synthesized by a
Romanian chemist named Lazar Edeleanu at the University of Berlin in 1887. However, it

CHAPTER 1 – Introduction and Aims
4
was not used clinically until Gordon A. Alles re-synthesized the drug in the 1920s for use
in medical settings as a bronchodilator to treat asthma and colds (Benzedrine©) (Menhard,
2006; Berman et al., 2008; Cunha-Oliveira et al., 2013).
The most popular ATS include methamphetamine - synthesized six years later - 3-
4-methylenedioxymethamphetamine (MDMA) and methylphenidate (Ritalin©, Concerta©,
Methylin©), patented in 1914 and in 1954, respectively (McDowel & Kleber, 1994;
Morton & Stockon, 2000, McCornack & Buckley, 2006). While methamphetamine is more
potent than the parent compound, amphetamine has moderate hallucinogenics properties
(Cunha-Oliveira et al, 2013).
ATS are indirect sympathomimetic and act in the nervous sytem by blocking
transporter-mediated reuptake of biogenic amines dopamine (DA), serotonin (5-HT) and
noradrenaline (NA) and by triggering an aberrant release of these amines from the
presynaptic terminal into the synaptic cleft through the reversal of the amine transporters
(Sulzer et al., 2005). Therefore, ATS produce a series of effects in mind and body
including increasing energy, heart rate, respiration, and mental alertness, elevating mood
and self-confidence, decreasing fatigue and sleep, producing euphoria and appetite
suppression (Cunha-Oliveira et al., 2013; Berman et al., 2008).
While amphetamine, METH and methylphenidate have long been used for treating
various disorders such as attention deficit hyperactivity disorder (ADHD), narcolepsy, and
obesity, they also may cause addition, profound effects on mental function and behavior,
and can produce neurodegeneration (Sulzer et al., 2005; National Institute on Drug Abuse
(NIDA; 2012); Koob & Volkow, 2010 and McAvoy, 2009).
1.2 Epidemiology of abuse amphetamine-type stimulants
According to the World Drug Report 2012, which was issued by UNODC,
amphetamines (amphetamine and methamphetamine) are the prescription drugs most
commonly abused by adolescents and young adults, and illicit amphetamines abuse ranks
second place in young adults, with an estimated prevalence of 0.3-1.2% in 2010 (between
14.3 million and 52.5 million consumers) (Table 1; Berman et al., 2008).

CHAPTER 1 – Introduction and Aims
5
Table 1 – Drug Consumption in 2010, (UNODC). Information taken from World Drug Report (2012).
In 2011, between 3.6 and 6.9 per cent of the adult population were estimated to
have used an illicit substance in the preceding year. The prevalence of illicit drug use and
the numbers of drug users with drug use disorders or dependence have remained stable
(World Drug Report, 2013). While the prevalence of use of cocaine, amphetamines and
“ecstasy”- group substances appears to have followed a declining trend between 2009 and
2011, the prevalence of cannabis, opioids, and opiates use has risen up (Figure 2).
Nevertheless, since 2008 there has been an overall 18 per cent increase in the estimated
total number of people who had used an illicit substance in the preceding year, which to
some extent reflects both an increase in the global population and a slight increase in the
prevalence of illicit drug use.

CHAPTER 1 – Introduction and Aims
6
Figure 2 – Trends in the prevalence of different drugs, 2009-2011. Information taken from World Drug
Report (2013).
According to World Drug Report (2013), Oceania, North America and Central
America represent the regions with the highest prevalence of abuse of amphetamines.
Moreover Southeast and Central Asia have been witnessing a growth in its abuse in recent
years (Figure 3).
Figure 3 – Use of ATS in 2011 (or latest year available). Information taken from World Drug Report (2013).

CHAPTER 1 – Introduction and Aims
7
According to the National Survey on the Use of Psychoactive Substances in the
General Population, Portugal 2012, amphetamines use prevalence increased 0.4% from 2001
to 2007 and youngest population is increasing its use in Portugal. The Algarve region,
followed by Lisbon and Tagus Valley, has the highest rates of amphetamines use in both
consumption throughout life and in continuation rates (Amaral & Guimarães 2012).
Relatively to METH, nearly 25 million people worldwide are estimated to have used
methamphetamine (Buxton & Dove, 2008; McAvoy, 2009), a total that exceeds the number
of people who abuse heroin and cocaine and makes METH the second most widely abused
drug after cannabis. This is consistent with METH having become very popular due to its
inexpensive production, low cost of acquisition, and durability in terms of effects (Krasnova
& Cadet, 2009; Koob & Volkow, 2010), becoming a global epidemic (Barr et al., 2006).
1.3 Methamphetamine and Serotonin
1.3.1. Methamphetamine: pharmacokinetics and pharmacodynamics
Methamphetamine was introduced in the 1930s as a bronchodilator for the
treatment of nasal and bronchial congestion associated with colds (Meng et al., 1999;
Guerreiro & Carmo et al., 2011), and was originally synthesized from ephedrine by the
Japanese pharmacologist Nagayoshy Nagai in 1893, but it was not widely used until World
War II when Japan, Germany, and the United States gave it to military personnel to combat
fatigue and increase endurance and performance (Freese et al., 2002; Meredith et al.,
2005). METH can also be synthesized through pseudoephedrine reduction or a
condensation of phenylacetone and methylamine (Cho, 1990, 2001). The produced METH
is a lipid-soluble pure base form, which is volatile and evaporates if left exposed to room
air. The producer, therefore, tipically uses hydrochloride to convert it to
methamphetamine-HCL powder, which is water-soluble. This METH salt is marketed on
the streets as “speed”, “crank”, “go”, “crystal” or methamphetamine (Derlet, 1990;
McAvoy, 2009).
The additional methyl group found in METH confers higher lipid solubility to this
molecule, comparatively to amphetamine, which results in greater blood brain barrier
(BBB) permeability (Aron & Paulus, 2007), enhanced stability against enzymatic
degradation by monoamine oxidase (MAO), and hence longer duration of action (Barr et
al., 2006) (Figure 4).

CHAPTER 1 – Introduction and Aims
8
Figure 4 – Chemical structure of methamphetamine (METH) (1), as well as the closely related
psychostimulant d-amphetamine (AMPH) (2). Adapted from Barr et al., 2006.
The clinical toxicology of drugs of abuse depends on the administration pathway,
which affects its bioavailability (affecting the onset and extent of the psychotropic effects),
the biodistribution (and therefore the exposure of target organs), and biotransformation or
metabolism, which occurs mainly in the liver (affecting the nature and concentration of
toxic compounds in the organism). The intensity and onset of a drug’s effects are
determined by the rapidity of its delivery to the central nervous system (CNS). Drug users
learn to optimize the delivery of the drug to the brain and to maximize the bioavailability
of the drug by adapting the methods and routes of administration. (Cunha-Oliveira et al.,
2013).
In addition to the traditional routes of administration, such as oral ingestion,
intravenous injection, and nasal snorting, METH can be smoked and also dissolved
sublingually, or solubilized and consumed in a beverage (Meng et al., 1999). When
administered intravenously or smoked, small doses of METH have prominent central
stimulant effects, acting almost immediately and causing intense pleasure (rush or flash)
that last only a few minutes (Schepers, 2003; Kish, 2008). Thus, these two pathways
consumption allow greater drug concentration at the sites of action at the level of the CNS
and, therefore, provide greater potential for addiction and increase the risk of overdose
(McAvoy, 2009). When consumed intranasally or orally, METH effect is neither so
immediate nor so intense when compared to intravenous and inhalation routes, because of
lower bioavailability of the former routes. After oral ingestion, METH is rapidly absorbed,
due to being highly lipid soluble, with peak plasma levels occurring within 2.6–3.6 h

CHAPTER 1 – Introduction and Aims
9
(Cunha-Oliveira et al., 2013). Overall, effects of METH typically persist for 4-8h, but the
residual effects can last up to 12h (McAvoy, 2009). A typical daily dose of oral METH for
the treatment of ADHD in children is 20-25 mg (Kish, 2008). However, the required dose
to produce a euphoric effect, typical of this drug, is 40 to 60 mg/day. Common abused
doses of METH are 100-1000 mg daily, and up to 5000 mg in chronic binge use (McAvoy,
2009).
The desired acute effects include well-being, increased alertness, increased activity,
excitement, and decreased appetite and anxiety. METH intoxication initially produces
excessive stimulation of the sympathetic nervous system, resulting in marked tachycardia,
hypertension, pupillary dilation, diaphoresis, tachypnea, peripheral hyperthermia and
hyperpyrexia (Meredith et al., 2005; Homer et al., 2008). These effects can last for several
hours because the elimination half-line of METH ranges from 10 to 12h (Schepers et al.,
2003).
High doses may result in restlessness and agitation, and excessive doses may
produce stereotypic behaviors (repetitive and automatic acts). Medical problems associated
with excessive dosages include cerebral hemorrhage, stroke, seizure, hyperthermia,
arrhythmias, coma, and death (NIDA, 1998). Chronic METH addicts often present altered
brain structure and function underlying deficits in attention, working memory and
decision-making and altered behavioral and cognitive functions (Krasnova & Cadet, 2009).
1.3.2. Serotonin Biosynthesis
Serotonin (5-hydroxytryptamine, 5-HT) is one of the ubiquitous molecules acting
as messengers, well known as a neurotransmitter and neuromodulator (Berumen et al.,
2011). 5-HT is phylogentically ancient and evolved prior to the apperanece of neurons
(Muller & Jacobs, 2010). This biogenic amine was first isolated from mammalian
organism in 1946 and then from the brain 7 years later (Hoyer et al., 2001; Hannon &
Hoyer, 2008) and was named by Rapport et al. (1948).
The molecular machinery governing serotonin biosynthesis across serotonergic
neurons is well characterized and also well conserved among vertebrates and invertebrates
(Curran & Chalasani, 2012).
Figure 5 depicts synthesis and metabolism of 5-HT. This is a monoamine having an
indoleamine group (Nishida, 2007) and being synthesized from L-tryptophan. This
precursor is converted into L-5-hydroxytryptophan (L-5-HTP) by an enzymatic reaction

CHAPTER 1 – Introduction and Aims
10
catalyzed by tryptophan hydroxylase-2 (TPH2). Then, the enzyme aromatic L-amino acid
decarboxylase (AADC) converts L-5-HTP to 5-HT. Subsequently, vesicle monoamine
transporters (type 2) (VMAT2) are responsible for packaging serotonin into vesicles.
Evidence suggests that stored 5-HT is released in an even sprinkler-type fashion termed
volume transmission, and functional concentrations of this neurotransmitter are maintained
several microns from release sites (Doubert & Condron, 2010). Extracellular 5-HT binds to
serotonin receptors on pre-and post-synaptic sites.
The signal is terminated when unbound serotonin is taken back into the presynaptic
cells by reuptake transporters (SERT), thereby limiting 5-HT extracellular concentration.
Finally, the enzymatic degradation of brain 5-HT is mainly mediated by monoamine
oxidase MAO-A and, in the absence of this enzyme, by its cognate isoenzyme MAO-B.
(Bortolato er al., 2008). In addition, since MAO-B is the only isoenzyme expressed in
platelets (Bond & Cundall, 1977), it may play an important function in the regulation of
plasma 5-HT levels. This metabolic step origins the non-active aldehyde derivative 5-
hydroxyindoleacetic acid (5-HIAA) (Chase & Koelle, 2007; Curran & Chalasani, 2012).
Several genes encoding the synthetic and recycling enzymes for the synthesis of co-factor
tetrahydrobiopterin (BH4) for TPH2 in the synthesis of 5-THP are also represented in
figure 5.

CHAPTER 1 – Introduction and Aims
11
Figure 5 – Synthesis and metabolism of serotonin. 5-HT neurons coexpress genes directing 5-HT synthesis
(Tph2, Aadc), reuptake (Sert), vesicular transport (Vmat2), autoreceptor signaling (Htr1a, Htr1b) and
metabolism (Maoa, Maob). Tetrahydrobiopterin (BH4), an essential cofactor for tryptophan hydroxylase 2
(Tph2) in the synthesis of 5-hydroxytryptophan (5-HTP), is synthesized (red pathway) de novo from
guanosine triphosphate (GTP). It is also recycled through a regeneration pathway (orange). Genes Gch1,
Gfrp, Ptps, Qdpr, Pcbd and Spr are directly involved in these BH4 red and orange pathways. Aldehyde
dehydrogenase (Aldh) converts 5-hydroxyindolealdehyde (5-HIAL) into 5-hydroxyindoleacetic acid (5-
HIAA). Synaptic 5-HT modulates 5-HT neuron firing through somatodendritic Htr1a autoreceptors (5-
HT1a), 5-HT release from the presynaptic terminal through Htr1b autoreceptors (5-HT1b) and stimulates
neurotransmission through postsynaptic 5-HT receptors (5-HT1–7). Aadc, aromatic l-amino acid
decarboxylase; Sert, serotonin transporter; Vmat2, vesicular monoamine transporter 2; Maoa, monoamine
oxidase; Gfrp, GTP cyclohydrolase I feedback regulator; Gch1, GTP cyclohydrolase 1; Ptps, 6-pyruvoyl-
tetrahydropterin synthase; Spr, sepiapterin reductase; Pcbd, pterin-4-alpha-carbinolamine dehydratase; Qdpr,
quinoid dihydropteridine reductase. BH4 synthetic intermediates: H2NTP, 7,8-dihydroneopterin
triphosphate; PTP, 6-pyruvoyl-5,6,7,8-tetrahydropterin; BH4αC, tetrahydrobiopterin-4α-carbinolamine;
qBH2, quinoid dihydrobiopterin (Taken from Deneris & Wyler, 2012).
Serotonin content in the central nervous system constitutes only 1–2% of the whole
pool of this monoamine in the organism and it does not cross blood-brain barrier (Filip et
al., 2005). Serotonergic CNS neurons are found primarily in nine clusters located mostly in
the raphe nuclei of the midbrainm pons, and medulla (Figure 6a). The more rostral raphe
nuclei contain the principal dorsal raphe groups (B6 and B7; depicted in yellow) and the
median raphe groups (B5 and B8; depicted in green) that project to overlapping brain areas

CHAPTER 1 – Introduction and Aims
12
structures, including amygdala, thalamus, hypothalamus, hippocampus, and frontal cortex.
The more caudal nuclei (B1–B3) in the medulla project axons to the spinal cord and the
periphery. On the other hand, in the periphery, a partially separate serotonergic enteric
neural system (ENS) exists within the gut and interacts with other CNS and peripheral
serotonergic mechanisms (PNS), that includes lung, heart, blood-vessel and pancreatic
tissue, as well as platelets. Notably, these tissues normally take up and store serotonin in
vesicles, and can release it in response to local stimuli (Figure 6b).
5-HT also play a role in the neuroendocrine serotonergic system (NSS) that
includes the hypothalamo–pituitary–adrenocortical (HPA) system. There is an interaction
between 5-HT and hormones such as prolactin, ACTH, corticosterone and oxytocin within
this NSS system (Murphy & Lesch, 2008).

CHAPTER 1 – Introduction and Aims
13
Figure 6 – 5-HT pathways. a – Central serotonergic system: CNS serotonin neuron cell-body groups in the
nine raphe nuclei, B1–B9. The more caudal nuclei (B1–B3) in the medulla project axons to the spinal cord
and the periphery, whereas the more rostral raphe nuclei contain the principal dorsal raphe groups (B6 and
B7; depicted in yellow) and the median raphe groups (B5 and B8; depicted in green), which project to
different but overlapping brain areas. b – Peripheral serotonergic system: Serotonin also functions in the
enteric nervous system (ENS), the hypothalamo–pituitary–adrenocortical (HPA) system, the
adrenomedullary neuroendocrine serotonin system (NSS) and the peripheral serotonin system (PSS), which
includes the lungs, the heart, the blood vessels, the pancreas and platelets. DRN, dorsal raphe nucleus; MFB,
medial frontal bundle; MRN, median raphe nucleus. (Taken from Murphy & Lesch, 2008).

CHAPTER 1 – Introduction and Aims
14
Therefore, central and peripheral 5-HT system fulfill a significant role in the
regulation of many vital functions of the organism including sleep, circadian rhythm,
emotional, feeding, cognitive and reproductive behaviors, thermoregulation, nociceptive
transmission, motor, endocrine, cardiovascular and respiratory functions, and intestinal
peristalsis. Moreover, serotonin plays a major role in the etiology of the related
pathological states, including depression, anxiety, schizophrenia, obsessive –compulsive
and panic disorders, autism, in addition to migraine, hypertension, pulmonary
hypertension, eating disorders, vomiting and, irritable bowel syndrome (IBS) (Filip et al.,
2005).
1.3.3. 5-HT Receptors
Based on structural (amino acid sequence), biochemical (postreceptor mechanisms
of signal transduction) and pharmacological differences, 5-HT receptors were classified
into 7 classes and 16 different subtypes. A majority of these receptors belong to the
metabotropic receptor family, except for 5-HT3 (5-HT3A, 5-HT3B and 5-HT3C) receptors,
which are included in the ionotropic receptor family. 5-HT1 (5-HT1A, 5-HT1B, 5-HT1D, 5-
HT1E and 5-HT1F) receptors inhibit adenylate cyclase; 5-HT2 (5-HT2A, 5-HT2B and 5-HT2C)
receptors stimulate phospholipase C; 5-HT4, 5-HT6 and 5-HT7 receptors stimulate
adenylate cyclase, while the mechanism of signal transduction via 5-HT5 (5-HT5A and 5-
HT5B) has not been yet satisfactorily defined (Filip et al., 2005; Curran & Chalasani, 2012;
Rojas et al., 2014).
Preclinical studies have shown an increase of 5-HT1A receptor-mediated
hippocampal transmissions after long-term treatment with selective serotonin reuptake
inhibitors (SSRIs) and other antidepressant drug classes, suggesting that this receptor is
involved in mood disorders. However, most selective 5-HT1A agonists developed so far
have failed to demonstrate clinical effectiveness (Celada et al., 2004). On the other hand,
de Angelis (2002) reviewed data presenting evidence that there is an association between
the 5-HT2A receptor gene and psychiatric disorders including schizophrenia, tardive
dyskinesia, major depression, suicidality, anorexia nervosa and obsessive-compulsive
disorder. Furthermore, chronic administration of 5-HT2A antagonists results in a
paradoxical downregulation of 5-HT2A receptors. This was suggested to be of benefit in the
treatment of depression (Glennon & Dukat, 2002). Also 5-HT2A receptors mediate the
primary effects of hallucinogenic drugs (Vollenweider et al., 1998; Gonzalez-Maeso et al.,

CHAPTER 1 – Introduction and Aims
15
2007). Interestingly, hallucinations and cognitive impairment are the typical clinical
symptoms of METH psychosis.
1.3.4. Methamphetamine action in serotonergic systems
Serotonin has been shown to be of essential importance for the hedonic tone
(O’Leary & Cryan, 2010), motivational (Lee & Clifton, 2010) and reinforcement processes
(McBride, 2010), and for learning and memory (Cassel, 2010). Accordingly, it is not
surprising that drugs of abuse, which induce profound changes in extra-cellular 5-HT
activity and 5-HT receptor function, change the behaviour-organizing circuitry of the brain
directly by modulating the 5-HT system, or, indirectly, by 5-HT effects on other
transmitter systems (Adell et al., 2010). Chronic drug exposure alters 5-HT tissue levels,
basal extracellular activity, and responsiveness of the 5-HT system to acute drug
administration. These effects may serve as neurochemical mechanisms for the acute
behavioural and subjective effects. They may also contribute to the transition to addiction.
Neurochemical studies showed that almost all psychoactive drugs with abuse
potential acutely increase 5-HT activity throughout the brain (Muller & Homberg, 2014).
In particular, it has been reported that METH acutely increases extracellular 5-HT levels in
the nucleus accumbens (Nac), dorsal striatum, ventral hippocampus and prefrontal cortex
(PFC) (Kuczenski et al., 1995; Segal et al., 1997; Rocher & Gardier, 2001; Ago et al.,
2006; Ikeda et al., 2011 and Yubero-Lahoz et al., 2012) from rodents. Studies in rodents
further showed that the rapid increase in the concentration of 5-HT METH-induced is
mediated by different mechanisms: in addition to stimulating the release of 5-HT, it
inhibits its reuptake (Brodkin et al., 1993) (Figure 7).
Neurotoxic METH doses following an acute aberrant 5-HT release also cause long-
term deleterious effects to serotonergic neuronal systems (Krasnova & Cadet, 2009).

CHAPTER 1 – Introduction and Aims
16
Figure 7 – The impact of METH on a serotonergic synapse (Adapted from Torres et al., 2003).
In fact, experimental neurotoxic rodent models including single-day multiple-dose
and sub-chronic METH administration have consistently shown that dysfunctional
serotonin nerve terminals are in place (Rusyniak, 2011; Marshall et al., 2007; Belcher et
al., 2008 Ladenheim et al., 2000; Fumagalli et al., 1998). These serotonergic changes
include a persistent fall in the levels of 5-HT as well as 5-HT transporter (SERT) within
several brain regions including the striatum, hippocampus, medial prefrontal and
somatosensory cortices, hypothalamus and amygdala (Kish et al., 2009; Sekine et al, 2006;
Gluck et al., 2001; Friedman et al., 1998; Baldwin et al., 1993; Ohmori et al., 1993; Bakhit
et al., 1981). Furthermore a significant decrease in SERT binding in the anterior cingulate,
nucleus accumbens, amygdala, hippocampus, somatosensory cortex, hypothalamus,
thalamus and septum was also reported (Armstrong & Noguchi, 2004; Guilarte et al.,
2003).
Finally, these neurotoxic METH dosing regimens also disturb TPH activity in the
cortex, hippocampus and nucleus accumbens from rodents (Bakhit et al., 1981; Hotchkiss
& Gibb, 1980; Morgan & Gibb, 1980). These experimental models are consistent with
terminal serotonergic toxicity measured in human addicts (see section Depressive

CHAPTER 1 – Introduction and Aims
17
behaviour during withdrawal from METH). Moreover, Chiu et al. (2014) demonstrated that
a single-day 'binge' METH dosing regimen caused an up-regulation of 5-HT2A receptor in
the medial frontal cortex, a brain region involved in perception, cognition and mood. This
is suggestive that this dosing regimen, while producing detrimental effects to serotonergic
nerve terminals, also changes 5-HT receptor density.
It has also been shown that METH, can also cause other neurodegenerative changes
that include dopaminergic terminal toxicity, loss of gray matter accompanied by
hypertrophy of the white matter, gliosis (astrocytic and microglia proliferation and
reactivity, in different brain areas), cortical and hippocampal BBB changes and oxidative
stress (Krasnova & Cadet, 2010; Pereira et al., 2004, 2006, 2012; Bowyer & Ali, 2006;
Bowyer et al., 2008; Martins et al. 2011, 2013; Silverstein et al., 2011; Ramirez et al.,
2009; Park et al., 2012 and Moszczynska & Yamamoto, 2011).

CHAPTER 1 – Introduction and Aims
18
2. Depression
Depressive disorders including major depressive disorder (MDD) rank within the
most prevalent forms of psychiatric disorders and are among the leading causes of
disability (Andrews et al., 2000). Today, depression is estimated to affect 350 million
people (WHO, 2012; Wen et al., 2014) and has a 12-month prevalence of 6.6% and a
lifetime prevalence of 16.2%, is twice as common in women as in men, and causes
considerable impairment (Kupfer et al., 2012; Belmaker & Agam, 2008). The World
Health Organization (WHO) estimates that depression will be the second most prevalent
cause of illness-induced disability by the year 2020 (Murray & Lopez, 1997).
Depression presents with depressed mood, loss of interest or pleasure (anhedonia),
irritability, decreased energy, feelings of guilt or low self-worth, disturbed sleep or
appetite, and poor concentration (Warner-Schmidt & Duman, 2006). Moreover, depression
often comes with symptoms of anxiety. These problems can become chronic or recurrent
and lead to substantial impairments in an individual’s ability to take care of his or her
everyday responsibilities. MDD is the leading cause of suicide worldwide. Almost 1
million lives are lost yearly due to suicide, which translates to 3000 suicide deaths every
day. For every person who completes a suicide, 20 or more may attempt to end his or her
life (WHO, 2012; Wen et al., 2014). Most likely, depression is caused by a combination of
genetic, biological, environmental, and psychological factors. Thus, vulnerable and/or
exposed individuals to certain endogenous and exogenous stressors, may develop,
cognitive, behavioral and somatic emotional dysfunction that translates into a depressive
syndrome (Tsuang et al. 2004).
Although the identification of specific neural substrates for depressive disorders is
still under investigation, hippocampus has received a great deal of attention (Warner-
Schmidt & Duman, 2006). Particularly, preclinical and clinical studies demonstrate that
reductions of the total volume of neurons and neuronal atrophy or loss occur in the adult
hippocampus in a context of stress and depression. Hippocampal circuitry functions
include the control of learning and memory and regulation of the hypothalamic-pituitary-
adrenal (HPA) axis, which are altered in depression. In addition, the hippocampus interacts
with the amygdala and frontal cortex, regions that are more directly involved in emotion
cand cognition and thereby contribute to major symptoms of depression (Duman &
Monteggia, 2006).

CHAPTER 1 – Introduction and Aims
19
2.1. Depression: Focusing on 5-HT
The 5-HT hypothesis of depression stretch back over 40 years to the discovery that
the first generation of antidepressant drugs blocked 5-HT reuptake or metabolism as part of
their pharmacological effect (Sharp & Cowen, 2002). Although the nature of the 5-HT
defect remains elusive, the best evidence that 5-HT contributes to the pathophysiology of
depression comes from studies of tryptophan depletion, which show that lowering brain 5-
HT levels can induce acute symptomatic relapse in recovered depressed patients (Cowen,
2008; Ruhé et al., 2007; Smith et al., 1997).
This link between 5-HT and depression provided a rich source of discoveries
including the development of newest and popular selective 5-HT reuptake (SSRIs), which
are currently the first choice antidepressant drug treatment. SSRIs block 5-HT reuptake,
leading to an increase in extracellular 5-HT levels (Blier, 2001; Quesseveur et al., 2013).
This monomine increase activates feedback mechanisms mediated by 5-HT1A (cell body)
and 5-HT1B (terminal) autoreceptors, which, respectively, reduce the firing in dorsal raphe
5-HT neurons and decrease the amount of 5-HT released per action potential, resulting in
attenuated 5-HT neurotransmission in regions such as frontal cortex and hippocampus.
Long-term treatment desensitizes the inhibitory 5-HT1 autoreceptors, and 5-HT
neurotransmission is enhanced. The time course of these events is similar to the delay of
clinical action.
Overall SSRIs seem to allow what might be an important convergence of
neurobiological and psychological explanations of the role of 5-HT in antidepressant
action and depression (Sharp & Cowen, 2002). The oldest class of antidepressant
medications is monoamine oxidase inhibitors (MAOIs). Tricyclics are other types of older
antidepressants. Tricyclics antidepressants (TCA) inhibit non-selectively 5-HT and
noradrenaline uptake (Katzung et al., 2001). Interestingly, previously referred hippocampal
alterations can be reversed by chronic antidepressant treatment (Lee & Kim, 2010).
However the nature of post-synaptic signaling and the contribution of glia to the
antidepressant pharmacological effects are largely unknown. Both TCAs and MAOIs are
potentially lethal in overdose, require titration to achieve a therapeutic dose, have serious
drug interactions and have many troublesome adverse effects. Consequently, these drugs
are now reserved for patients who do not respond to other agents.
Moreover, due to its long therapeutic time delay, low rates of remission, and
exposure to relevant side effects, including gastrointestinal, hepatic and sexual, the

CHAPTER 1 – Introduction and Aims
20
research for more effective agents in the treatment of these disorders has been greatly
stimulated (Manji et al., 2008; Berton & Nestler, 2006). For example, physical exercise,
being able to relieve symptoms of depression (APA, DSM-V-TR, 2013), seems to be a
non-pharmacological alternative strategy in the treatment of depression.
Finally, although not addressed in our work, other hypothesis pointing towards a
role for other neurotransmitters including NA, inflammation, neuroplasticty and epigenetic
regulation have also been implied in depressive disorders (Han & Yun, 2014; Massart et
al., 2012; Krishnan & Nestler, 2008).
2.2. Depressive behaviour during withdrawal from METH
Mood disorders, including depression and bipolar disorders, are the most common
psychiatric comorbidities among patients with substance use disorders (Quello et al., 2005;
NIH, 2011). For example, diminished interest or pleasure in rewarding stimuli or
anhedonia is one of the core symptoms of both depression and psychostimulant
withdrawal. Importantly, METH withdrawal and major depression share many behavioral
commonalities in humans (Cryan et al., 2003). In detail, repeated METH use produces a
withdrawal syndrome marked more by psychiatric complaints than by physical
manifestations. These withdrawal syndrome typically manifests as anxiety, depression with
severe dysphoria, irritability and melancholia, social isolation, fatigue with hypersomnia,
psychomotor dysfunction, mood disturbances, impaired social functioning, intense craving
for the drug and even paranoia or aggression, as well as attention deficits and memory in
making decisions (London et al., 2004; Meredith et al., 2005; Darke et al., 2008; Homer et
al., 2008, Sutcliffe et al., 2008; Krasnova & Cadet, 2009; Daniulaityte et al., 2010;
Guerreiro & Carmo et al., 2011).
Although the severity of the abstinence syndrome appears to be related to the
frequency of use, it tends to resolve spontaneously, while the depressive symptoms persist
for months or years after discontinuation of drug use (Barr et al., 2002; Cami & Farre,
2003). Moreover, the prevalence of depression among METH users is higher than the
general population, with the majority of METH users reporting a lifetime history of
depression and over a third reporting a lifetime diagnosis of depression. However, the
temporal relationship between depression and METH use is unclear. In fact, it is unknown
whether experiencing depressive symptoms promotes METH use, whether depression

CHAPTER 1 – Introduction and Aims
21
results from or is enhanced by METH use, or whether it is bidirectional (Sutcliffe et al.,
2009).
Addressing this issue is clinically relevant as it may inform about treatment options
for both METH dependence and mental health (Pecke et al., 2005). Therefore, the
examination of the behavioral effects of METH withdrawal in rodents may provide
insights into the neurobiological mechanisms underlying both disorders. For example,
Cryan et al. (2003) showed that drug-withdrawn rodents displayed increased immobility in
forced swim and tail suspension tasks (24 hours after deprivation of METH; 5 mg/kg/day
for 7 days), suggesting that there is a depressive-like behavior. Recently, Jang et al. (2013)
found that chronic METH self-administration produced a depressive-like withdrawal state
in rats during early withdrawal.
We also recently provided novel data that a single high neurotoxic dose of METH
(30 mg/kg i.p.) evoked depressive-like behavior as gauged by increased immobility in the
tail-suspension task at 3 days and 7 weeks post-METH treatment (Silva et al., 2014). This
negative affect-like behavior was underlined by prolonged frontostriatal dopaminergic
deficits including DA and TH. Importantly we also demonstrated that this METH regimen
evoked 5-HT depletion in frontal cortex at 3 and 49 days post-METH injection. However
we failed to probe METH impact in hippocampus. This is suggestive that this 5-HT
homeostasis disruption might also contribute to the depressive-like behavior.
Interestingly, abstinent METH abusers showed reduced brain 5-HT transporter
density and cerebral metabolic abnormalities associated with depressive symptoms
(London et al., 2004; Sekine et al., 2006; Panenka et al., 2012). It is worthwhile stress that
psychostimulant withdrawal seems to provide the basis for the development of an animal
model of depressive symptoms, such as despair, anhedonia, lethargy and anxiety (Paulson
et al., 1991; Cryan et al., 2003), thus allowing the screening of new pharmacological or
non-pharmacological approaches (Barr et al., 2002). This is particularly relevant since
there is no reliable treatment that can reverse the effects of psychostimulant withdrawal
rapidly and completely at present.

CHAPTER 1 – Introduction and Aims
22
3. Physical Exercise
3.1. Physical Exercise: a non-pharmacological approach in
depressive disorders
Data from epidemiological studies suggest a strong association between physical
inactivity and high levels of depressive symptoms (Farmer et al., 1988; Camacho et al.,
1991; Conn et al., 2010). In fact, low levels of physical activity lead to an increase in
symptoms of depression in young (Motl et al., 2004; Strong et al., 2005; Neissaar et al.,
2011) and older adults (Blumenthal et al., 1999; Lampinen et al., 2000; Brosse et al., 2002;
Conn et al., 2010).
On the other hand, human studies suggest that exercise may be useful in the
prevention and treatment of psychiatric disorders such as depression (Conn et al., 2010)
and anxiety disorders (Dunn et al., 2010), Furthermore, Lawlor & Hopker (2001)
reviewed data suggesting that exercise has antidepressant effects that are of the same
magnitude as cognitive therapy in the management of mild-to-moderate mental health
diseases. It also was demonstrated that the antidepressant effects of exercise were
prolonged beyond the period of treatment with benefits until 6 (Babyak et al., 2000) and 21
months (Singh et al., 2001) after stopping the exercise. Trivedi et al. (2006) showed that
exercise reduced depressive symptoms or reduced side effects when combined with a
pharmacological approach. In addition, Frazer et al., 2005 indicate chronic exercise as one
of the best non pharmacological treatments for depression, besides the fact that it is low
cost, have positive effects on cardiovascular disease risk, and be a potential prevention
agent for future depressive episodes. Others further proposed that exercise was as effective
as antidepressant medications (Blumenthal et al., 1999; Strawbridge et al., 2002).
However, habitual physical activity has not been shown to prevent the onset of depression
(Paluska & Schwenk, 2000). One possible neurobiological mechanism underlying the
positive effects of exercise is the increased synthesis of 5-HT (Matta et al., 2013). This is
the studied hypothesis in the present work.
Despite the relationship between the efficacy of physical exercise in reducing the
symptoms of depression already has been widely documented, there is still a lack of data
on the type of physical exercise that induces better antidepressant response (Mead et al.,
2008). On one hand, aerobic exercise is associated with higher benefits (Ernst et al., 2006).
For example, a program of 12 weeks of aerobic exercise functioned as an effective

CHAPTER 1 – Introduction and Aims
23
treatment for depression of mild to moderate severity (Dunn et al., 2002, 2005).
Furthermore, increased aerobic exercise or strength training has been shown to
significantly reduce depressive symptoms (Paluska & Schwenk, 2000; Craft & Perna,
2004). On the other hand, Greenwood et al. (2013) were the first to suggest that individuals
who have made forced physical exercise can still benefit from its protective effects against
psychiatric disorders related to stress.
Although effective pharmacological interventions are available, depression remains
inadequately treated. Thus, physical exercise seems to be a non-pharmacological tool in
the treatment of psychiatric diseases such as depression and, therefore, in the promotion of
a better mental health.
A cautionary note should be added. In fact, it was suggested that excessive physical
activity may lead to overtraining and generate psychological symptoms that mimic
depression (Paluska & Schwenk, 2000).
3.2. Physical Exercise: managing METH addiction and
neurotoxicity
To date, there are no medications approved for the treatment of METH dependence
(Mooney et al., 2014 and Karila et al., 2010). Behavioral approaches, such as cognitive
behavioral therapy and contingency management, have proven modestly effective and
remain the standard treatment. However, there are substantial proportions of individuals
dropping out early in the treatment (Rawson et al., 2004; Peirce et al., 2006; Roll et al.,
2006; Rawson et al., 2006).
Notably, there is an increasing amount of literature suggesting that physical
exercise is as a potential treatment for METH-dependent individuals (Mooney et al., 2014).
For example, it was demonstrated that physical exercise improved cognitive deficits found
in chronic METH users (Simon et al., 2000). Recently, a study in humans reported for the
first time that 8-weeks of endurance and resistance exercise training triggered positive
adaptations on individuals recovering from METH-dependence, such as substantial
improvements in aerobic exercise performance, muscle strength and endurance, and body
composition with exercise training. These changes are consistent with those seen in the
general population, suggesting that structured and supervised exercise training should be
considered as part of the overall treatment plan in recovering METH patients (Dolezal et
al., 2013). Another study examined whether a combination of aerobic exercise and

CHAPTER 1 – Introduction and Aims
24
resistance training could facilitate relapse prevention and maintenance of abstinence from
METH after discharge from residential treatment (Mooney et al., 2014). These authors
suggested that exercise may provide a reinforcing behavior that offers an alternative to
drug use as a means of enhancing positive mood states.
Furthermore, exercise may have a salutary effect on reducing cardiovascular risk
factors, such as hypertension, that are associated with METH use (Mooney et al., 2009;
Turnipseed et al., 2003). Exercise also improves sleep (Youngstedt et al., 2005) and
performance on cognitive tasks, which may be impaired in METH users and in long-term
METH users in early phases of abstinence (Mooney et al., 2014).
On the other hand, O’Dell et al. (2012) demonstrated that running wheel exercise
ameliorates methamphetamine-induced damage to forebrain DA and 5-HT terminals in
rats. Moreover, it was shown that exercise protected against cerebrovascular toxicity of
METH mice (Toborek et al., 2009). Remakably, we have recently shown that treadmill
exercise reversed the long-lasting behavioural phenotype triggered by single-METH
neurotoxic dose as demonstrated by decreased immobility time in the tail suspension test
when compared to sedentary METH-intoxicated mice (Pereira et al., 2014, unpublished
data).

CHAPTER 1 – Introduction and Aims
25
4. Aims of this thesis
This study aims to further characterize the long-lasting depressive phenotype
caused by a single neurotoxic METH-injection and to probe for a correlation between
frontal-cortical and hippocampal 5-HT homeostasis and e the antidepressant effect of
physical exercise.

CHAPTER 2
MATERIAL AND METHODS

CHAPTER 2 – Material and Methods
27
1. Animals
In this investigation we used 44 C57BL/6 male mice (10 weeks old, 21-26 g,
Charles River Laboratories, Barcelona, Spain) (Figure 8), divided into 11 cages (4/cage)
and maintained in the animal house of the Faculty of Medicine, University of Coimbra
(FMUC) under controlled environmental conditions (temperature, 22 ± 1 ° C, humidity 50
± 10 %, light cycle, 12/12 hours), with food and water provided ad libitum. Their weight
was monitored weekly to assess the evolution of the different experimental groups.
All experimental procedures followed the rules imposed by the Standards of
Animal protection techniques used for experimental and other scientific purposes
(Ordinance No 129/92 of 6 July), as well as the standards of the European Convention on
Animal Welfare (Ordinance No 1005/92) and in accordance with the guidelines of the
European Community (2010/63/EU). All efforts to minimize animal suffering and to use
the smallest possible number of animals were made.
Figure 8 – Example of a C57BL/6 mouse. Taken from http://jaxmice.jax.org/strain/002020.html.
2. Drugs and Chemicals
We were issued permission to import methamphetamine.HCL (METH) for Sigma-
Aldrich (St. Louis, MO, USA9 by INFARMED, Portugal (National Authority of
Medicines and Healths Products). Standards for serotonin (5-HT) were purchased from
Sigma-Aldrich. The other used chemicals (ultrapure and pro analysis quality) were
purchased from Sigma-Aldrich and Merck AG (Darmstadt, Germany).

CHAPTER 2 – Material and Methods
28
3. Experimental Design
32 animals were randomly divided into 4 groups, with 8 animals per group: (1)
sedentary saline group (Cont/SED), (2) exercise saline group (Cont/EX), (3) METH
sedentary group (METH/SED) and (4) exercise METH group (METH/EX) (table 2). The
animals were previously identified with ink stripes on the tail and housed in their
respective cages, having been subjected to an adaptation period of 2 weeks in controlled
vivarium conditions (mentioned above). The weight of the mice was recorded weekly,
using an analytical balance (Kern CB 6 K1, Germany). However, the weighing was done
daily on subsequent administration of the neurotoxin week.
Table 2 – Experimental groups and number of mice used (n).
Mice (n=32)
Exercise (n=16) Sedentary (n=16)
METH/EX
(n=8)
Cont/EX
(n=8)
METH/SED
(n=8)
Cont/SED
(n=8)
3.1. Treadmill adaptation
Herein, two treadmills were used [the LE8700 model, serial number 2187/07 and
the LE8706 model, serial number 8589/04, both with 50 W, 110/120 V and 50/60 Hz;
Panlab, SL, Barcelona, Spain,] for the implementation of aerobic exercise. Separating each
lane with transparent acrylic, thus diminishing the width of each corridor and resized these
models, which are suitable for mice. Therefore, it was possible to exercise 4 mice per
treadmill at the same time (Figure 12).
Mice submitted to exercise (SAL/EX and METH/EX groups) were putin a daily
workout plan aiming to foster treadmill adaptation for 2 weeks (5 days/week). The
adaptation protocol consisted in four phases (Figure 9), with a progressive increase in the
total time (20 min at the beginning to 40 min on the last week) and intensity (daily
maximum speed of 20 to 30 cm/s) of each training session until achieving the values used
in the exercise protocol.
The treadmill was turned on to a minimum speed of 5cm/seg only after and the
animals were placed in the lanes. Thereafter, the speed was gradually increased (about
1cm/seg) until the desired speed, thus comprising the initial stage of the session - heating

CHAPTER 2 – Material and Methods
29
phase. After the time and intensity required for the training session were applied, the speed
periodically decreased at the same rate until the treadmill is turned off -– cooling phase.
Both phases are relevant because they aim to prevent injury.
Figure 9 – Mice adaptation protocol to treadmill. Tr, training and R, rest.
3.2. Methamphetamine administration
Subsequently to the adaptation protocol to the treadmill and prior to the beggining
of the exercise protocol, all mice were weighed again to calculate METH dose for each
mouse.
Control groups (SAL/SED and SAL/EX groups) were administered a saline
solution (NaCl 0.9%, 250μl.), while METH/SED and METH/EX groups were injected
with 30 mg/kg methamphetamine.HCL (METH; 3mg/ml). Animals were dosed with a
single intraperitoneal injection (i.p.) (Figure 10). We have recently demonstrated that this
dose induces astrogliosis, as well as dopaminergic and serotonergic terminal toxicity to
striatal-cortical regions in mice as soon as 3 days post-METH (Pereira et al. 2012; Silva et
al. 2014). However, we failed to characterize serotonergic homeostasis in hippocampus
from these neurotoxic models at this time-point. Therefore we used another set of 12 mice
to probe METH-impact on hippocampal total 5-HT levels at 3 days post-injection. Half of
the animals were injected with saline.

CHAPTER 2 – Material and Methods
30
Figure 10 – Intraperitoneal administration of a single dose of METH (30 mg/kg).
After injection, the animals were carefully observed, at 0.5, 1 and 3 time-points.
Animals receiving this neurotoxin were extremely agitated, which is consistent with what
is reported in the literature (Meredith et al., 2005).
Two of the mice belonging to the METH/EX group were found dead the following
day. This may be due to hyperthermia caused by METH.
Infarmed Portugal (National Authority of Medicines and Health Products IP) issued
necessary authorizations for FMUC to acquire METH from Life Science and High
Technology Company Sigma-Aldrich (St. Louis, MO, USA).

CHAPTER 2 – Material and Methods
31
3.3. Exercise protocol
Figure 11 – Physical exercise protocol for the respective exercise groups, SAL/EX and METH/EX. Ex,
exercise; R, rest and S, sacrifice.
Twenty-four hours follwoing METH/NaCl administration, the exercise groups
(SAL/EX and METH/EX) underwent a protocol of daily running (Figure 11), 5 days per
week for 7 weeks (35 days of exercise) according to the following planning: 1) 5 min
heating at 20 cm/s; 2) 30 min of running at 30 cm/s; 3) 5 min cooling at 20 cm/s. Exercise
was always performed on the same morning period. The inclination of the treadmill was
always null (0%). Mice were prompted to run by gentle manual strokes instead of using
electric shocks, thus minimizing stress imposed to animals, which is considered to be a
possible factor stress that is not usually associated with exercise.
Protocol:
- Ex: Velocity – 20cm/s – 5 min; 30cm/s – 30 min; 20cm/s – 5 min
- R: Rest
- BT: Behavioural test
- BT/S: Behavioural test/Sacrifice

CHAPTER 2 – Material and Methods
32
Figure 12 – Mice training on a treadmill separated by acrylic divisions, providing four individual
lanes.
The animals in the sedentary groups ran only once a week for 10 minutes at
minimum speed of 5 cm/s to experience the same stressful conditions felt by the animals of
the exercise groups. Woods et al. (2003) proposed that the engine noise of the treadmill,
vibration, treadmill texture, water and food deprivation and frequent handling of animals
are some of the stressful factors.
4. Behavioural Tests
Behavioural tests, Open-field (OF), Splash and Forced-swim test (FST) were
performed, 48 and days after administration of METH (30 mg/kg, i.p.) or saline to assess
mice emotional states. The first test was performed in the first day, whereas the Splash and
the FST were conducted on the second day.
Tests were all performed between 9 and 17h, in a sound-attenuated room lit with
low intensity Light (12 lx). Mice were transferred to this room 1h before the start of the
testing to get used to the environment. The behaviour was monitored by a video camera
positioned above the equipment and the images were subsequently analyzed with the
system of video monitoring ANY Maze (Stoelting Co., Wood Dale, IL, USA) by an
experienced experimenter who was unaware of the experimental group being tested.

CHAPTER 2 – Material and Methods
33
4.1. Open-field test
The open-field (OF) test procedure consists of subjecting an animal to an unknown
environment from which escape is prevented by surrounding walls (Walsh and Cummins,
1976). We applied herein a 5-min test length to assess the effect of treadmill exercise on
novel environment exploration and general locomotor activity in METH-treated METH
mice (Bayley & Crawley, 2009; Machado et al., 2012.
Mice were individually placed in the center of a wooden box (40×40×50 cm3), and
the following behavioral items were recorded: horizontal locomotion (total distance
travelled), central distance, central-total distance ratio, number of rearing (sometimes
termed vertical activity) and average speed. Tipically, mice prefer spontaneously the
periphery of the apparatus to activity in the central parts of the open field (Figure 13).
Indeed, rodents walk close to the walls, a behavior called thigmotaxis. Therefore, increase
of time spent in the central part, as well as of the ratio central/total locomotion are
indications of anxiolytic-like behavior (Prut & Belzung, 2003).
The apparatus was cleaned with a solution of ethanol 10% between tests in order to
remove animal odors or clues.
Figure 13 – Open Field test: mice in open-field test apparatus.

CHAPTER 2 – Material and Methods
34
4.2. Splash test
This pharmacological test was adapted from Yalcin et al. (2005) and was used
herein to evaluate the impact of treadmill exercise on grooming behaviour in mice injected
with METH as an index of self-care and motivational behaviour phenotype of the
experimental groups. For this purpose, 10% sucrose solution was squirted on the dorsal
coat of mice in their home cage. Following this viscous solution dirtying the mice fur,
animals initiate grooming behaviour. The latency for the first grooming and the total
grooming time were recorded in an acrylic chamber (40 x 40x 40 cm) during 5 minutes
(Figure 14).
Grooming bouts were recorded including nose/face grooming (strokes along the
snout), head washing (semicircular movements over the top of the head and behind the
ears) and body grooming (body fur licking) (Kaluelf & Tuohimaa, 2004). Anhedonic
symptoms were characterized by increased latency (midle time between spray and
initiation of grooming) and decreased time spent grooming (d’Audiffret et al., 2010).
Figure 14 – Splash test: A - mice in the cage 2 is grooming his dorsal coat in his home cage; B - schematic
illustration of dorsal mice grooming. Adapted from http://mybrainnotes.com/bdd-trichotillomania-skin-
picking.html.
B
A
1 2

CHAPTER 2 – Material and Methods
35
4.3. Forced-swim test
The Forced-swim test (FST) has been used as a model predictive of antidepressant
effect (Cryan et al., 2002). The test procedure was carried out according Porsolt et al.
(1977) with some modifications. Mice were individually forced to swim in an open
cylindrical container (21 cm height × 12 cm internal diameter) containing fresh water till a
height of 15 cm at 23±1 °C; the total duration of immobility was recorded during a 5
minute period (Figure 15). The water was changed and the cylinder rinsed with clean water
after each mice. A trained observer that was blind to the treatment measured the
immobility time.
During the 5 min swimming test session, the following behavioral responses were
recorded by a trained observer: the immobility time (i.e. the time spent floating in the
water without struggling, making only those movements necessary to keep its head above
water), swimming (time spent actively swimming around in circles), and climbing
behavior, which is defined as upward directed movements of the forepaw along the
cylinder walls. This test involved the scoring of active (swimming and climbing) and
passive (immobility) behavior when mice were forced to swim in a cylinder from which
there is no escape. A decrease in the duration of immobility time and an increase in
swimming time is indicative of an antidepressant-like effect (Porsolt et al., 1977), while
time of climbing was used as a predictor of altered motor activity scored directly in the
forced swimming test (Vieira et al., 2008). Typically, after the initial 2-3 minutes of
vigorous activity the animals showed a period of immobility by floating with minimum
movements. An animal is considered to be immobile whenever it remained floating
passively in the water in a slightly hunched but up right position, its nose above the water
surface (Khulbe et al., 2013).

CHAPTER 2 – Material and Methods
36
Figure 15 – Forced-swim test. Mice placed into an open cylindrical container.
5. Frontal Cortex and Hippocampus Isolation
Following behavioural tests, animals were sacrificed by cervical dislocation and
decapitated - that is 48 h after the end of the protocol of physical exercise (on the second
day of rest) or 49 days after administration of METH/saline.
The brains were rapidly removed, and hippocampus and the frontal cortex were
dissected on icebased on the coordinates for the mouse brain described by Paxinos &
Franklin (2004). The biological samples were immediately frozen in dry ice and stored at -
80 ° C until 5-HT analysis by high pressure liquid chromatography with coulometric
electrochemical detection (HPLC).
6. Determination of 5-HT by HPLC
A reversed-phase high-performance liquid chromatography (HPLC) method with
coulometric electrochemical detection was applied to quantify 5-HT brain levels (Figure
17) (Pereira et al., 2011). The used equipment is illustrated in figure 16 and included a
Gilson pump (model 307), an autosampler Gilson (model 234; 50mL loop), a detector
Gilson (coulometric) and v5.11 software.

CHAPTER 2 – Material and Methods
37
Figure 16 – HPLC system used to quantify 5-HT in this study. A - Gilson pump, B - autosampler Gilson, C -
detector Gilson and D - v5.11 software. (Picture taken at Laboratory of Pharmacology and Experimental
Therapeutics, Faculty of Medicine, University of Coimbra).
Figure 17 – Chromatogram illustrating a 5-HT peak relative to a frontal cortex sample from a METH/EX
mice group. The obtained 5-HT retention time was between 8 - 9 min.
B D
A
C
5-HT

CHAPTER 2 – Material and Methods
38
perchloric acid 0.2 M. The homogenates were centrifuged (13,000 rpm for 7 min, 4 ° C)
and filtered with 0.22 mM Nylon microfilter (Spin-X ® Centrifuge Tube Filter, Costar at
10,000 rpm for 4 minutes, 4° C). The pellets were resuspended in NaOH 1 M and used for
quantification of the protein by the bicinchoninic acid method (BCA) and reading by
ELISA. The filtered supernatants and the pellets were stored at -80 ºC.
Serotonin was separated on a reversed-phase, Grace Platinum EPS C18 column ®
(4.6 x 150 mm, particle size: 5 mM) with a mobile, degassed and filtered phase consisting
of monopotassium phosphate (25 mM), 1-heptanesulphonic acid (0.4 mM), EDTA (50
µM), 3% acetonitrile (pH 2.5). The flow rate was 1 mL/minute. Samples were quantified
by an analytical cell (model 5011, ESA Analytical, Dorton Aylesbury, Buckinghamshire,
UK) attached to a Coulochem-II electrochemical detector (ESA, Analytical). The
analytical cell was set at E1=+250 mV (sensitivity - 200 nA).
The concentration of 5-HT in each sample was calculated taking as reference the
standard curves of 5-HT. The results were expressed in ng/mg protein.
7. Statistical analysis
The results are expressed as mean ± standard error (SEM). Body weight data were
analysed by one-way ANOVA followed by Newman–Keuls multiple comparison.
Behavioural and neurochemical data were analysed by two-way ANOVA (drug x
exercise) followed by Bonferroni multiple comparison. Significant differences were
defined at P<0.05. The statistical analyses were performed using GraphPad Prism 6.0
software.

CHAPTER 3
RESULTS

CHAPTER 3 – Results
40
1. Effects of methamphetamine and/or chronic physical exercise
on body weight
The mice body weight significantly increased in all groups (P<0.05) over the total
duration of the experimental protocol (61 days, Figure 18 A,B). Interestingly, mice body
weight from each group is not significantly different from each other at the last time
evaluated (P> 0.05) (Figure 18 B).
Figure 18 – A: Evolution of the mice body-weight over 61 days (total duration of experimental protocol). B:
Mice body-weight at 3 different time-points: day 1: beginning of training; day 14: METH administration; day
61: last day of physical exercise program. The results are expressed as mean ± S.E.M. of 6-8 animals per
group. Control sedentary mice body-weight at the 61 days is statistically different from the other time points
($P<0.05). #P<0.05. Statistical analysis was performed using one-way ANOVA followed by Newman–Keuls
multiple comparison.
A
B

CHAPTER 3 – Results
41
2. Effect of treadmill exercise on general exploratory
locomotion in METH-exposed mice
2.1. Open-field test
Mice activity was quantified in the open-field over 5 minutes to assess novel
environment exploration 49 days post-METH injection. METH-sedentary mice showed
normal total distance (Figure 19 A) and central distance travelled (Figure 19 B), normal
central distance–total distance ratio (Figure 19 C), number or rearings (Figure 19 D) and
average speed (Figure 19 E). This is suggestive that the general exploratory behavior of
these animals is not statistically different from that showed by the respective controls
(SAL/SED) (P>0.05). Exercise increased all these measured parameters in both METH and
Cont (P<0.05). This is highly suggestive that physical activity enhanced both locomotor
activity and exploratory behavior.
0
10
20
30
sedentary
Cont METH
exercise
Cont METH
# #
dis
tan
ce (
m)
0
2
4
6
8
sedentary
Cont METH
exercise
Cont METH
##
cen
tral
dis
tan
ce (
m)
0
5
10
15
20
25
sedentary
Cont METH
exercise
Cont METH
##
nu
mb
er
of
reari
ng
s
0
2
4
6
8
10
sedentary
Cont METH
exercise
Cont METH
# #
av
era
ge
sp
ee
d (
cm
/s)
0
10
20
30
sedentary
Cont METH
exercise
Cont METH
# #
dis
tan
ce (
m)
0
2
4
6
8
sedentary
Cont METH
exercise
Cont METH
##
cen
tral
dis
tan
ce (
m)
0
5
10
15
20
25
sedentary
Cont METH
exercise
Cont METH
##
nu
mb
er
of
reari
ng
s
0
2
4
6
8
10
sedentary
Cont METH
exercise
Cont METH
# #
av
era
ge
sp
ee
d (
cm
/s)
A B
0
10
20
30
sedentary
Cont METH
exercise
Cont METH
# #
dis
tan
ce (
m)
0
2
4
6
8
sedentary
Cont METH
exercise
Cont METH
##
cen
tral
dis
tan
ce (
m)
0
5
10
15
20
25
sedentary
Cont METH
exercise
Cont METH
##
nu
mb
er
of
reari
ng
s
0
2
4
6
8
10
sedentary
Cont METH
exercise
Cont METH
# #
av
era
ge
sp
ee
d (
cm
/s)
C D
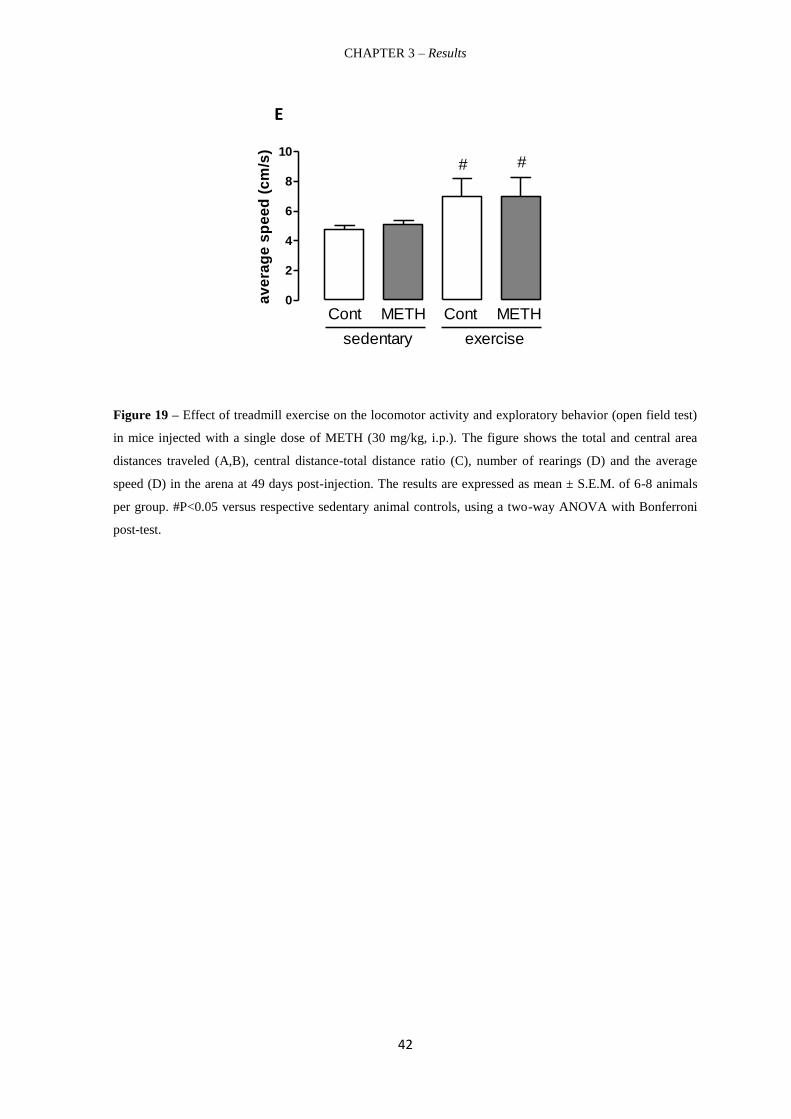
CHAPTER 3 – Results
42
Figure 19 – Effect of treadmill exercise on the locomotor activity and exploratory behavior (open field test)
in mice injected with a single dose of METH (30 mg/kg, i.p.). The figure shows the total and central area
distances traveled (A,B), central distance-total distance ratio (C), number of rearings (D) and the average
speed (D) in the arena at 49 days post-injection. The results are expressed as mean ± S.E.M. of 6-8 animals
per group. #P<0.05 versus respective sedentary animal controls, using a two-way ANOVA with Bonferroni
post-test.
0
10
20
30
sedentary
Cont METH
exercise
Cont METH
# #d
ista
nce (
m)
0
2
4
6
8
sedentary
Cont METH
exercise
Cont METH
##
cen
tral
dis
tan
ce (
m)
0
5
10
15
20
25
sedentary
Cont METH
exercise
Cont METH
##
nu
mb
er
of
reari
ng
s
0
2
4
6
8
10
sedentary
Cont METH
exercise
Cont METH
# #
av
era
ge
sp
ee
d (
cm
/s)
E
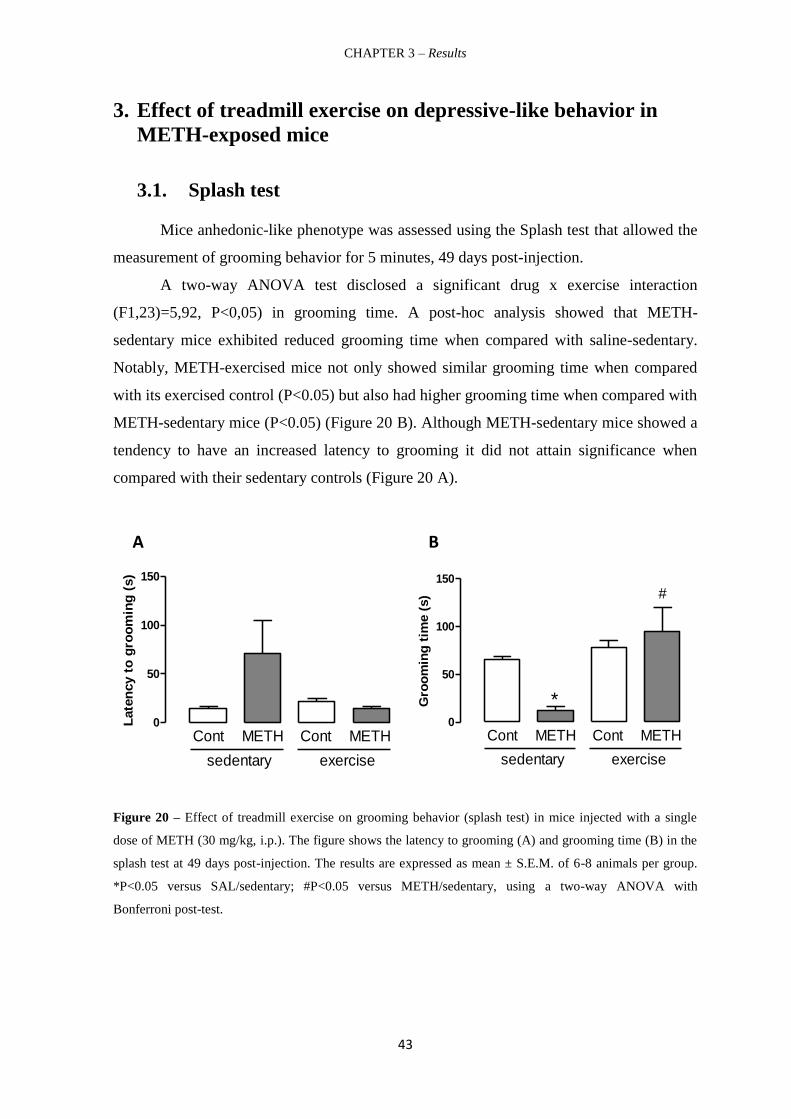
CHAPTER 3 – Results
43
3. Effect of treadmill exercise on depressive-like behavior in
METH-exposed mice
3.1. Splash test
Mice anhedonic-like phenotype was assessed using the Splash test that allowed the
measurement of grooming behavior for 5 minutes, 49 days post-injection.
A two-way ANOVA test disclosed a significant drug x exercise interaction
(F1,23)=5,92, P<0,05) in grooming time. A post-hoc analysis showed that METH-
sedentary mice exhibited reduced grooming time when compared with saline-sedentary.
Notably, METH-exercised mice not only showed similar grooming time when compared
with its exercised control (P<0.05) but also had higher grooming time when compared with
METH-sedentary mice (P<0.05) (Figure 20 B). Although METH-sedentary mice showed a
tendency to have an increased latency to grooming it did not attain significance when
compared with their sedentary controls (Figure 20 A).
Figure 20 – Effect of treadmill exercise on grooming behavior (splash test) in mice injected with a single
dose of METH (30 mg/kg, i.p.). The figure shows the latency to grooming (A) and grooming time (B) in the
splash test at 49 days post-injection. The results are expressed as mean ± S.E.M. of 6-8 animals per group.
*P<0.05 versus SAL/sedentary; #P<0.05 versus METH/sedentary, using a two-way ANOVA with
Bonferroni post-test.
0
50
100
150
sedentary
Cont METH
exercise
Cont METH
Late
ncy t
o g
roo
min
g (
s)
0
50
100
150
sedentary
Cont METH
exercise
Cont METH
#
*Gro
om
ing
tim
e (
s)
0
50
100
150
sedentary
Cont METH
exercise
Cont METH
Late
ncy t
o g
roo
min
g (
s)
0
50
100
150
sedentary
Cont METH
exercise
Cont METH
#
*Gro
om
ing
tim
e (
s)
A B

CHAPTER 3 – Results
44
3.2. Forced-swim test
Mice despair-like behaviour was probed using forced swim-test for 5 minutes, 49
days post-METH injection.
METH-sedentary mice did not show a despair-like behavior as gauged by
immobility, swimming and climbing times not being statistically different from those
shown by their respective controls (SAL/SED) (P>0.05; Figure 21 A, B, C). Notably,
treadmill exercise decreased the immobility time (Figure 21 A) and increased the
swimming time (Figure 21 B) in both SAL and METH groups when compared to their
sedentary controls (P<0.05; Figure 22 A, B, C). Nonetheless exercise failed to change the
climbing time (C) relatively to sedentary groups (P>0.05; Figure 21 C).
Figure 21 – Effect of treadmill exercise on the depair-like behavior (forced swimming test) in mice injected
with a single dose of METH (30 mg/kg, i.p.). The figure shows the immobility (A), swimming (B) and
climbing (C) times in the forced swimming test at 49 days post-injection. The results are expressed as mean ±
S.E.M. of 6-8 animals per group. #P<0.05 versus respective sedentary control animals using a two-way
ANOVA followed by with Bonferroni post-test.
0
50
100
150
200
sedentary
Cont METH
exercise
Cont METH
#
#
Imm
obilit
y t
ime (
s)
0
50
100
150
200
sedentary
Cont METH
exercise
Cont METH
# #
Sw
imm
ing
tim
e (
s)
0
50
100
150
sedentary
Cont METH
exercise
Cont METH
Clim
bin
g t
ime (
s)
0
50
100
150
200
sedentary
Cont METH
exercise
Cont METH
#
#
Imm
obilit
y t
ime (
s)
0
50
100
150
200
sedentary
Cont METH
exercise
Cont METH
# #
Sw
imm
ing
tim
e (
s)
0
50
100
150
sedentary
Cont METH
exercise
Cont METH
Clim
bin
g t
ime (
s)
A B
0
50
100
150
200
sedentary
Cont METH
exercise
Cont METH
#
#
Imm
obilit
y t
ime (
s)
0
50
100
150
200
sedentary
Cont METH
exercise
Cont METH
# #
Sw
imm
ing
tim
e (
s)
0
50
100
150
sedentary
Cont METH
exercise
Cont METH
Clim
bin
g t
ime (
s)
C

CHAPTER 3 – Results
45
4. Effect of treadmill exercise on 5-HT total content in frontal
cortex and hippocampi from METH-exposed mice
Serotonin is a neurochemical fingerprint from depressive-like behavior and was
measured herein by HPLC. Hippocampal 5-HT levels were quantified 3 and 49 days post-
METH injection. Frontal cortical 5-HT levels were only determined at the final end-point.
METH imposed a hippocampal 5-HT depletion as seen in Figure 22 A. However, neither
drug nor exercise had a significant effect on cortical and hippocampal 5-HT total content
(P>0.05; Figure 22 B,C).
Figure 22 – Effect of treadmill exercise on hippocampal (A,B) and frontal cortical (C) serotoninlevels (5-HT
measured from mice at 3 (A) and 49 days (B, C) post-METH injection). The results are expressed as mean ±
S.E.M. of 6-8 animals per group. *P<0,05 versus saline-treated animals using unpaired t-test.
A: 3 days post - METH injection
B: 49 days post - METH injection C: 49 days post - METH injection

CHAPTER 4
DISCUSSION

CHAPTER 4 - Discussion
47
Discussion
Although clinical studies have shown the relevance of the symptoms of depression
in consumers of METH on abstinence (Glasner-Edwards et al. 2009 and Volkow et al.,
2001; Rusyniak, 2011), this psychiaytric abnormality in METH abuse context has been
scarcely studied. In fact, the study of the long-term mood behavioural and underlying
neurochemical consequences of METH consumption is still poorly characterized.
Therefore herein we offer a comprehensive characterization of the long-term emotional
state of mice following a single-neurotoxic dose of METH using Splash, forced swim test
(FST) and open field (OF). We want ultimately to robustly confirm that METH induces
depressive-like behaviour. We further aimed to establish a relationship between 5-HT
homeostasis in frontal-cortex and hippocampus and the putative antidepressant effect
induced by physical exercise.
The single high METH dose regimen that we use in the present study recapitulates
the detrimental effects found in METH users including corticalstriatal dopaminergic and
astroglial dysfunction (Cappon et al., 2000; Imam & Ali, 2001; Xu et al., 2005; Zhu et al.,
2005; Krasnova & Cadet, 2009; O’Callaghan et al., 2008; Kitamura et al., 2010; Sailasuta
et al., 2010; Pereira et al., 2012; Silva et al., 2014). In addition, this model reduces the
inherent complexity present in repeated dosage regimens and reduces the occurrence of
seizures and high mortality seen with multiple doses regimen (Davidson et al., 2001;
Miller & O’Callaghan, 2003). In fact, in the present study, only two mice died and none
showed convulsions or weight reductions. It is noteworthy to stress that animals from the
exercise group started to put on weight following 14 days of adaptation to the treadmill.
This might suggest that the animals had to cope with a stressful situation that they
overcome as shown by weight evolution and behavioral phenotype (see above).
The splash test is a direct measure of self-care and motivational behavior and has
been reported to probe for mood disorders in rodent models. We observed for the first
time, that the METH regimen induced a decreased grooming behaviour in the METH-
sedentary mice, when compared to the respective control, which is indicative of a long-
term anhedonic-like behaviour that mimics apathy observed in clinical depression
(Willner, 2005). Furthermore, METH-sedentary mice showed a tendency to had increased
latency to grooming, further indicating loss of self-care and motivational behavior. This is
consistent with the fact that some chronic METH users exhibit an anhedonic and

CHAPTER 4 - Discussion
48
depressive behavior in a persistent manner after a few years follwoing the end of
consumption (Rawson et al. 2002). Physical exercise, by itself, didn’t alter the grooming
time of the mice, but recovered grooming-time in METH-intoxicated mice. This result
suggests that the practice of physical exercise is a non-pharmacologic strategy with anti-
anhedonic potential (Willner et al., 2005). We further analysed negative mice behaviour
using FST. This test was designed by Porsolt, as a primary screening test for
antidepressants with a strong predictive validity (Porsolt et al., 1977). Porsolt et al. (1977)
further proposed that the immobility time observed in this test reflected a state of lowered
mood or hopelessness in animals (Cryan et al., 2002).
FST showed that METH-sedentary mice exhibited normal immobility, swimming
and climbing times when compared with their sedentary controls. This is apparently
contractidory with METH-intoxicated mice having increased immobility time in the TST
at the same time-point (Silva et al., 2014). However, it was demonstrated that TST is more
sensitive than FST in illustrating neurochemical abnormalities (St´eru et al., 1985;
Castagn´e, et al., 2010; Chatterjee et al., 2011). On the other hand this might also further
suggest that TST and FST might reflect different pathophysiological mechanisms
underlying immobility as proposed by Chatterjee et al. (2011).
Finally, Castagn´e, et al. (2010) described numerous advantages of using TST over
FST such as no induction of hypothermia and the animals once taken out from the
experimental set up resume instantly normal spontaneous activity. More importantly,
treadmill exercise decreased immobility time and increased swimming time, irrespectively
of drug treatment, when compared to their sedentary controls. These behavioural
observations clearly demonstrate that physical exercise is endowed with
antidespair/antidepressant-like properties. This further corroborates our TST data showing
that treadmill exercise decreased the immobility time spent by METH-intoxicated mice 49
days post-METH (Pereira et al., 2014, unpublished). These antidepressant-like properties
are independent of locomotor behaviour activity since treadmill exercise had no effect on
climbing time from both METH- and Cont- trated mice. We also evaluated novel
environment exploration of mice 49 days post-METH injection using the OF test during 5
minutes. We demonstrated that METH regimen did not significantly alter the general
physical motor abilities and level of interest in the novelty of the environment as gauged
by similar total distance travelled, average speed and number of rearings compared to
saline-treated animals, in the sedentary group. Interestingly, Timár et al. (2003) reported a
decrease of spontaneous locomotor activity in rats 3 days after METH treatment (10

CHAPTER 4 - Discussion
49
mg/kg, 4 subcutaneous injections every 2 hours). However, this effect was transient since
1, 2 and 4 weeks after METH administration, animals revealed a normal locomotor
behavior. This further suggests that the anhedonic-like behavior showed by METH-
intoxicated mice is independent of locomotor activity. However, chronic exercise increased
all the measured OF parameters in both METH and Cont, suggesting that treadmill
enhanced both locomotor activity and exploratory behavior. These results could be related
with the high-mood state triggered by physical exercise as shown by both Splash and FST,
irrespectively of drug treatment. Consistently, these mice exhibited an anxiolytic-like
behaviour as gauged by increased center distance–total distance ratio compared to
sedentary mice.
Articulating the three behavioral tests used herein, one can propose that treadmill
exercise offers an antidepressant-like effect to mice submitted to a neurotoxic METH dose.
This pre-clinical approach gives further credence for the reported benefitial effects of
physical exercise to METH-abstinent individuals (Mooney et al., 2014).
We hypothesized that this beneficial impact of treadmill exercise is dependent upon
its modulatory impact on 5-HT homeostasis in both hippocampus and frontal cortex from
METH-intoxicated mice. Although METH imposed a robust hippocampal 5-HT depletion
(circa 45%) at 3 days post-injection, this neurochemical detrimental effect was reversed 49
days post-injection. On the other hand, treadmill exercise failed to significantly change 5-
HT levels in both saline and METH groups. Importantly, this means that physical exercise
had no effect on 5-HT hippocampal terminal recovery in METH-intoxicated mice.
Additionally, frontal cortical 5-HT levels from METH sedentary-intoxicated mice were not
statistically different from their control saline mice. On the other and, Silva et al. (2014)
showed that, METH imposed a 5-HT depletion of circa 25% in frontal cortex. Although
apparently contradictory, these results stress that the 5-HT depletion on frontal cortex was
not robust enough to be reproducible. One can also add that this METH regimen is more
detrimental to dopaminergic systems compared to serotonergic systems (Pereira et al.,
2012; Silva et al., 2014). Consistently with hippocampal 5-HT data, treadmill exercise
failed to significantly change 5-HT levels in frontal cortex from both saline and METH
groups. Overall, one can discard the hypothesis that treadmill exercise exerted an
antidepressant-like effect through modulating 5-HT synthesis in both of hippocampal and
frontal-cortex. However, one cannot disregard the hypothesis that METH-intoxicated mice
had changes in 5-HT extracellular basal levels and signaling. This warrants further
scrutiny.

CHAPTER 4 - Discussion
50
Disruption in the neurotrophic factors, which play a major role role in
neuroplasticity, could be also held responsible for the depressive-like behavior METH-
induced (Matta et al., 2013). Interestingly, it was also postulated that a reduction of
neurotrophic factors in limbic structures, including the hippocampus and prefrontal cortex
is directly involved in the pathophysiology of depressive disorders (Duman & Monteggia,
2006). Therefore, antidepressants might mediate their therapeutic benefit, in part, by
increasing levels of these factors in the hippocampus.
Importantly, it was shown that serum brain-derived neurotrophic factor (BDNF)
levels were significantly downregulated in METH abusers during the first 3 weeks of
withdrawal (Chen et al., 2012). Another study found that chronic METH abusers who had
been abstinent for at least 1 month had significantly higher plasma BDNF levels compared
to healthy control subjects (Kim et al., 2005).
Taken together, these studies strongly suggest that BDNF, was shown to be
decreased first and then increased after METH withdrawal in humans. These findings are
compatible with findings from animal studies. For example, neurotoxic-METH treatment
in rat increased BDNF in multiple brain regions from rat including frontal cortex and
hippocampal CA1 (Braun et al., 2011). Therefore probing the role of BDNF in the
beneficial treadmill exercise in METH-intoxicated mice is warranted.
On the other hand, there is also substantial evidence showing that regular physical
exercise is associated with a low reactivity of the sympathetic nervous system (SNS) and
the HPA axis, that plays a critical role in developing responses to psychological and
physical stressors (Stranahan et al., 2008 and Rimmele et al., 2009). Specifically,
physically active subjects showed lower cortisol increase (Rimmele et al., 2007), to
psychological laboratory stressors in comparison with less active controls. Thus, exercise-
induced changes in HPA axis might modulate negative affects in humans. Importantly,
adolescent METH use is associated with greater psychiatric symptoms and enhanced
cortisol secretion following a social stressor, particularly in younger female METH users
(King et al. 2010). Therefore the relevance of the HPA axis to the antidepressive-like effect
of treadmill exercise also warrants further analysis. Additionally other brain regions
including amygdala should be scrutinized.

CHAPTER 5
CONCLUSION

CHAPTER 5 - Conclusion
52
Conclusion
This study is relevant because it further demonstrated that a single-high METH
dose imposes a long-lasting depressive-like symptoms such as anhedonia. Furthermore, we
unequivocally show that physical exercise is a non-pharmacologic strategy that offers
antidepressive-like effects to METH-intoxicated mice. We also demonstrated that this
treadmill effect was independent of 5-HT homeostasis in both frontal-cortex and
hippocampus. Therefore, further studies are warranted to understand the neurochemical
basis for this positive behavioral effect.

CHAPTER 6
REFERENCES

CHAPTER 6 - References
54
References
Aguiar Jr, A. S., Castro, A. A., Moreira, E. L., Glaser, V., Santos, A. R., Tasca, C. I.,
Latini, A., & Prediger, R. D. (2011). Short bouts of mild-intensity physical exercise
improve spatial learning and memory in aging rats: Involvement of hippocampal
plasticity via AKT, CREB and BDNF signaling. Mechanisms of ageing and
development, 132(11), 560-567.
Al-Haggar, M. (2014). SNPs as Co-morbid Factors for Drug Abuse and Ischemic Heart
Disease. Gene Technology, 3(107), 2.
Allen, S. J., Watson, J. J., Shoemark, D. K., Barua, N. U., & Patel, N. K. (2013). GDNF,
NGF and BDNF as therapeutic options for neurodegeneration. Pharmacology &
therapeutics, 138(2), 155-175.
Amaral, A. S & Guimarães, M. I (2012). Manifestações orais do uso de metanfetaminas.
Rev Port de Estomatol, Med Dent Maxilofac, 53 (3):175-180..
Andrews G, Sanderson K, Corry J, Lapsley HM (2000): Using epidemiological data to
model efficiency in reducing the burden of depression*. J Ment Health Policy Econ
3:175-186.
Angelucci, F., Gruber, S. H., El Khoury, A., Tonali, P. A., & Mathé, A. A. (2007). Chronic
amphetamine treatment reduces NGF and BDNF in the rat brain. European
Neuropsychopharmacology, 17(12), 756-762.
Angelucci, F., Ricci, V., Spalletta, G., Caltagirone, C., Mathé, A. A., & Bria, P. (2009).
Effects of psychostimulants on neurotrophins: implications for psychostimulant-
induced neurotoxicity. International review of neurobiology, 88, 1-24.
Aron, J. L., & Paulus, M. P. (2007). Location, location: using functional magnetic
resonance imaging to pinpoint brain differences relevant to stimulant
use. Addiction, 102(s1), 33-43.
Audrain-McGovern, J., Rodriguez, D., & Moss, H. B. (2003). Smoking progression and
physical activity. Cancer Epidemiology Biomarkers & Prevention, 12(11), 1121-
1129.
Autry, A. E., & Monteggia, L. M. (2012). Brain-derived neurotrophic factor and
neuropsychiatric disorders. Pharmacological reviews, 64(2), 238-258.
Babyak, M., Blumenthal, J. A., Herman, S., Khatri, P., Doraiswamy, M., Moore, K., ... &
Krishnan, K. R. (2000). Exercise treatment for major depression: maintenance of
therapeutic benefit at 10 months. Psychosomatic medicine, 62(5), 633-638.

CHAPTER 6 - References
55
Barden, N. (2004). Implication of the hypothalamic–pituitary–adrenal axis in the
physiopathology of depression. Journal of Psychiatry and Neuroscience, 29(3),
185.
Barr, A. M., Markou, A., & Phillips, A. G. (2002). A ‘crash’course on psychostimulant
withdrawal as a model of depression. Trends in pharmacological sciences, 23(10),
475-482.
Barr, A. M., Panenka, W. J., MacEwan, G. W., Thornton, A. E., Lang, D. J., Honer, W. G.,
& Lecomte, T. (2006). The need for speed: an update on methamphetamine
addiction. Journal of Psychiatry and Neuroscience, 31(5), 301.
Bartha C, Parker C, Thomson C, Kitchen K. (1999) Depressive Illness: A Guide for People
with Depression and Their Families. 1999. Centre for addiction and mental health,
2185:3-6.
Berman, S., O'Neill, J., Fears, S., Bartzokis, G., & London, E. D. (2008). Abuse of
amphetamines and structural abnormalities in the brain. Annals of the New York
Academy of Sciences, 1141(1), 195-220.
Berton, O., & Nestler, E. J. (2006). New approaches to antidepressant drug discovery:
beyond monoamines. Nature Reviews Neuroscience, 7(2), 137-151.
Bhagwagar, Z., Whale, R., & COWEN, P. J. (2002). State and trait abnormalities in
serotonin function in major depression. The British Journal of Psychiatry, 180(1),
24-28.
Blair, S. N., Jacobs Jr, D. R., & Powell, K. E. (1985). Relationships between exercise or
physical activity and other health behaviors. Public health reports,100(2), 172.
Blier, P. (2000). Pharmacology of rapid-onset antidepressant treatment strategies. The
Journal of clinical psychiatry, 62, 12-17.
Blumenthal, J. A., Babyak, M. A., Moore, K. A., Craighead, W. E., Herman, S., Khatri, P.,
... & Krishnan, K. R. (1999). Effects of exercise training on older patients with
major depression. Archives of internal medicine, 159(19), 2349-2356.
Bobinski, F., Martins, D. F., Bratti, T., Mazzardo-Martins, L., Winkelmann-Duarte, E. C.,
Guglielmo, L. G. A., & Santos, A. R. S. (2011). Neuroprotective and
neuroregenerative effects of low-intensity aerobic exercise on sciatic nerve crush
injury in mice. Neuroscience, 194, 337-348.
Boutelle, K. N., Murray, D. M., Jeffery, R. W., Hennrikus, D. J., & Lando, H. A. (2000).
Associations between exercise and health behaviors in a community sample of
working adults. Preventive Medicine, 30(3), 217-224.

CHAPTER 6 - References
56
Bowyer, J. F., & Ali, S. (2006). High doses of methamphetamine that cause disruption of
the blood-brain barrier in limbic regions produce extensive neuronal degeneration
in mouse hippocampus. Synapse, 60(7), 521-532.
Bowyer, J. F., Robinson, B., Ali, S., & Schmued, L. C. (2008). Neurotoxic-.related
changes in tyrosine hydroxylase, microglia, myelin, and the blood-brain barrier in
the caudate-putamen from acute methamphetamine exposure. Synapse, 62(3), 193-
204.
Bowyer, J. F., Thomas, M., Schmued, L. C., & Ali, S. F. (2008). Brain Region–Specific
Neurodegenerative Profiles Showing the Relative Importance of Amphetamine
Dose, Hyperthermia, Seizures, and the Blood–Brain Barrier. Annals of the New
York Academy of Sciences, 1139(1), 127-139.
Boyle, R. G., O'Connor, P., Pronk, N., & Tan, A. (2000). Health behaviors of smokers, ex-
smokers, and never smokers in an HMO. Preventive medicine,31(2), 177-182.
Braun, Amanda A., et al. "Neurotoxic (+)-methamphetamine treatment in rats increases
brain-derived neurotrophic factor and tropomyosin receptor kinase B expression in
multiple brain regions." Neuroscience 184 (2011): 164-171.
Bremner, J. D., Narayan, M., Anderson, E. R., Staib, L. H., Miller, H. L., & Charney, D. S.
(2000). Hippocampal volume reduction in major depression. American Journal of
Psychiatry, 157(1), 115-118.
Brière, F. N., Fallu, J. S., Janosz, M., & Pagani, L. S. (2012). Prospective associations
between meth/amphetamine (speed) and MDMA (ecstasy) use and depressive
symptoms in secondary school students. Journal of epidemiology and community
health, 66(11), 990-994.
Brodkin, J., Malyala, A., & Frank Nash, J. (1993). Effect of acute monoamine depletion on
3, 4-methylenedioxymethamphetamine-induced neurotoxicity. Pharmacology
Biochemistry and Behavior, 45(3), 647-653.
Brosse, A. L., Sheets, E. S., Lett, H. S., & Blumenthal, J. A. (2002). Exercise and the
treatment of clinical depression in adults. Sports medicine, 32(12), 741-760.
Brunoni, A. R., Lopes, M., & Fregni, F. (2008). A systematic review and meta-analysis of
clinical studies on major depression and BDNF levels: implications for the role of
neuroplasticity in depression. The International Journal of
Neuropsychopharmacology, 11(08), 1169-1180.
Buchert, R., Obrocki, J., Thomasius, R., Väterlein, O., Petersen, K., Jenicke, L., ... &
Clausen, M. (2001). Long-term effects of ‘ecstasy’abuse on the human brain
studied by FDG PET. Nuclear medicine communications, 22(8), 889-897.
Calfas, K. J., & Taylor, W. C. (1994). Effects of physical activity on psychological
variables in adolescents. Pediatric exercise science, 6, 406-406.

CHAPTER 6 - References
57
Camacho, T. C., Roberts, R. E., Lazarus, N. B., Kaplan, G. A., & Cohen, R. D. (1991).
Physical activity and depression: evidence from the Alameda County
Study. American journal of epidemiology, 134(2), 220-231.
Cami, J., & Farré, M. (2003). Drug addiction. New England Journal of Medicine, 349(10),
975-986.
Cappon, G. D., Pu, C., & Vorhees, C. V. (2000). Time-course of methamphetamine-
induced neurotoxicity in rat caudate-putamen after single-dose treatment. Brain
research, 863(1), 106-111.
Carboni, L. (2013). Peripheral Biomarkers in Animal Models of Major Depressive
Disorder. Disease markers, 35(1), 33-41.
Carnicella, S., & Ron, D. (2009). GDNF-a potential target to treat addiction.
Pharmacology & therapeutics, 122(1), 9-18.
Cass, W. A., Peters, L. E., Harned, M. E., & Seroogy, K. B. (2006). Protection by GDNF
and Other Trophic Factors Against the Dopamine‐Depleting Effects of Neurotoxic
Doses of Methamphetamine. Annals of the New York Academy of
Sciences, 1074(1), 272-281.
Chang, L., Alicata, D., Ernst, T., & Volkow, N. (2007). Structural and metabolic brain
changes in the striatum associated with methamphetamine abuse.
Addiction, 102(s1), 16-32.
Chase, D. L., & Koelle, M. R. (2007). Biogenic amine neurotransmitters in C. elegans.
Chatterjee, M., Jaiswal, M., & Palit, G. (2012). Comparative evaluation of forced swim test
and tail suspension test as models of negative symptom of schizophrenia in
rodents. ISRN psychiatry, 2012.
Chen, P. H., Huang, M. C., Lai, Y. C., Chen, P. Y., & Liu, H. C. (2012). Serum
brain‐derived neurotrophic factor levels were reduced during methamphetamine
early withdrawal. Addiction biology.
Chiu, H. Y., Chan, M. H., Lee, M. Y., Chen, S. T., Zhan, Z. Y., & Chen, H. H. (2014).
Long-lasting alterations in 5-HT2A receptor after a binge regimen of
methamphetamine in mice. The international journal of
neuropsychopharmacology/official scientific journal of the Collegium
Internationale Neuropsychopharmacologicum (CINP), 1-12.
Cho A. et al. (2001) Patterns of Methamphetamine Abuse and Their Consequences,
Journal of Addictive Diseases, 21:1, 21-34.
Cho, A. K. (1990). Ice: a new dosage form of an old drug. Science, 249(4969), 631-634.

CHAPTER 6 - References
58
Conn, V. S. (2010). Depressive symptom outcomes of physical activity interventions:
meta-analysis findings. Annals of behavioral Medicine, 39(2), 128-138.
Cotman, C. W., & Berchtold, N. C. (2002). Exercise: a behavioral intervention to enhance
brain health and plasticity. Trends in neurosciences, 25(6), 295-301.
Cowen, P. J. (2008). Serotonin and depression: pathophysiological mechanism or
marketing myth?. Trends in pharmacological sciences, 29(9), 433-436.
Craft, L. L., & Landers, D. M. (1998). The effect of exercise on clinical depression and
depression resulting from mental illness: A meta-analysis. Journal of Sport and
Exercise Psychology, 20, 339-357.
Craft, L. L., & Perna, F. M. (2004). The benefits of exercise for the clinically
depressed. Primary care companion to the Journal of clinical psychiatry, 6(3), 104.
Crews, D. J., & Landers, D. M. (1987). A meta-analytic review of aerobic fitness and
reactivity to psychosocial stressors. Medicine & Science in Sports & Exercise.
Cryan, J. F., Markou, A., & Lucki, I. (2002). Assessing antidepressant activity in rodents:
recent developments and future needs. Trends in pharmacological sciences, 23(5),
238-245.
Cunha-Oliveira, T., Rego, A. C., Carvalho, F., & Oliveira, C. R. (2013). Chapter 17 –
Medical Toxicology of Drugs of Abuse. Principles of Addiction: Comprehensive
Addictive Behaviors and Disorders. Vol 1, 159-175.
Curran, K. P., & Chalasani, S. H. (2012). Serotonin circuits and anxiety: what can
invertebrates teach us? Invertebrate Neuroscience, 12(2), 81-92.
d'Audiffret, A. C., Frisbee, S. J., Stapleton, P. A., Goodwill, A. G., Isingrini, E., & Frisbee,
J. C. (2010). Depressive behavior and vascular dysfunction: a link between clinical
depression and vascular disease?. Journal of Applied Physiology, 108(5), 1041-
1051.
Daniulaityte, R., Falck, R., Wang, J., Carlson, R. G., Leukefeld, C. G., & Booth, B. M.
(2010). Predictors of depressive symptomatology among rural stimulant
users. Journal of psychoactive drugs, 42(4), 435-445.
Darke, S., Darke, S., Kaye, S., Darke, S., Kaye, S., McKetin, R., ... & Duflou, J. (2008).
Major physical and psychological harms of methamphetamine use. Drug and
alcohol review, 27(3), 253-262.
Daubert, E. A., & Condron, B. G. (2010). Serotonin: a regulator of neuronal morphology
and circuitry. Trends in neurosciences, 33(9), 424-434.
Dawn D Han., et al. Comparison of the monoamine transportes from human and mousse in
their sensitivities to psychostimulant drugs. BMC Pharmacology (2006) 6:6.

CHAPTER 6 - References
59
de Angelis, L. (2002). 5-HT2A antagonists in psychiatric disorders. Current opinion in
investigational drugs (London, England: 2000), 3(1), 106-112.
De Kloet, E. R., Joëls, M., & Holsboer, F. (2005). Stress and the brain: from adaptation to
disease. Nature Reviews Neuroscience, 6(6), 463-475.
Delgado, P. L., Price, L. H., Miller, H. L., Salomon, R. M., Licinio, J., Krystal, J. H., ... &
Charney, D. S. (1991). Rapid serotonin depletion as a provocative challenge test for
patients with major depression: relevance to antidepressant action and the
neurobiology of depression. Psychopharmacology Bulletin.
Derlet, R. W., & Heischober, B. (1990). Methamphetamine. Stimulant of the 1990s?
Western Journal of Medicine, 153(6), 625.
Deslandes, A., Moraes, H., Ferreira, C., Veiga, H., Silveira, H., Mouta, R., ... & Laks, J.
(2009). Exercise and mental health: many reasons to move.
Neuropsychobiology, 59(4), 191-198.
Dluzen, D. E. (2004). The effect of gender and the neurotrophin, BDNF, upon
methamphetamine-induced neurotoxicity of the nigrostriatal dopaminergic system
in mice. Neuroscience letters, 359(3), 135-138.
Dluzen, D. E., Anderson, L. I., McDermott, J. L., Kucera, J., & Walro, J. M. (2002).
Striatal dopamine output is compromised within+/− BDNF mice.Synapse, 43(2),
112-117.
Dolezal, B. A., Chudzynski, J., Storer, T. W., Abrazado, M., Penate, J., Mooney, L., ... &
Cooper, C. B. (2013). Eight weeks of exercise training improves fitness measures in
methamphetamine-dependent individuals in residential treatment. Journal of
addiction medicine, 7(2), 122-128.
Duman, R. S., & Monteggia, L. M. (2006). A neurotrophic model for stress-related mood
disorders. Biological psychiatry, 59(12), 1116-1127.
Dunn, A. L. (2010). Review: exercise programes reduce anxiety symptoms in sedentary
patients with chronic illnesses. Evid Based Ment Health. 13, 95.
Dunn, A. L., Trivedi, M. H., Kampert, J. B., Clark, C. G., & Chambliss, H. O. (2002). The
DOSE study: a clinical trial to examine efficacy and dose response of exercise as
treatment for depression. Controlled clinical trials, 23(5), 584-603.
Dunn, A. L., Trivedi, M. H., Kampert, J. B., Clark, C. G., & Chambliss, H. O. (2005).
Exercise treatment for depression: efficacy and dose response. American journal of
preventive medicine, 28(1), 1-8.
Erickson, K. I., Voss, M. W., Prakash, R. S., Basak, C., Szabo, A., Chaddock, L., ... &
Kramer, A. F. (2011). Exercise training increases size of hippocampus and

CHAPTER 6 - References
60
improves memory. Proceedings of the National Academy of Sciences, 108(7),
3017-3022.
Ernst, C., Olson, A. K., Pinel, J. P., Lam, R. W., & Christie, B. R. (2006). Antidepressant
effects of exercise: evidence for an adult-neurogenesis hypothesis? Journal of
psychiatry & neuroscience, 31(2), 84.
Escobedo, L. G., Marcus, S. E., Holtzman, D., & Giovino, G. A. (1993). Sports
participation, age at smoking initiation, and the risk of smoking among US high
school students. JAMA: the journal of the American Medical Association, 269(11),
1391-1395.
Etnier, J. L., Nowell, P. M., Landers, D. M., & Sibley, B. A. (2006). A meta-regression to
examine the relationship between aerobic fitness and cognitive performance. Brain
research reviews, 52(1), 119-130.
Farmer, M. E., Locke, B. Z., Moscicki, E. K., Dannerberg, A. L., Larson, D. B., & Radloff,
L. S. (1988). Physical activitu and depressive symptoms: the Nhanes I
Epidemiologic Follow-up Study. Amercian Journal of Epidemiology, 128(6), 1340-
1351.
Filip, M., Frankowska, M., Zaniewska, M., Golda, A., & Przegaliñski, E. (2005). The
serotonergic system and its role in cocaine addiction. Pharmacol Rep, 57(6), 685-
700.
Fontes-Ribeiro, C. A., Marques, E., Pereira, F. C., Silva, A. P., & Macedo, T. A. (2011).
May exercise prevent addiction? Current neuropharmacology, 9(1), 45.
Frazer, C. J., Christensen, H., & Griffiths, K. M. (2005). Effectiveness of treatments for
depression in older people. Med J Aust, 182(12), 627-32.
Freese T.E et al. (2002) The effects and consequences of selected club drugs. Journal of
substance, 23:151-156.
Fukumura, M., Cappon, G. D., Pu, C., Broening, H. W., & Vorhees, C. V. (1998). A single
dose model of methamphetamine-induced neurotoxicity in rats: effects on
neostriatal monoamines and glial fibrillary acidic protein. Brain research, 806(1),
1-7.
Fumagalli, F., Gainetdinov, R. R., Valenzano, K. J., & Caron, M. G. (1998). Role of
dopamine transporter in methamphetamine-induced neurotoxicity: evidence from
mice lacking the transporter. The Journal of neuroscience,18(13), 4861-4869.
Gardner, E. L. (2011). Addiction and brain reward and antireward pathways.
Ghitza, U. E., Zhai, H., Wu, P., Airavaara, M., Shaham, Y., & Lu, L. (2010). Role of
BDNF and GDNF in drug reward and relapse: a review. Neuroscience &
Biobehavioral Reviews, 35(2), 157-171.

CHAPTER 6 - References
61
Glasner-Edwards, S., Marinelli-Casey, P., Hillhouse, M., Ang, A., Mooney, L. J., &
Rawson, R. (2009). Depression among methamphetamine users: association with
outcomes from the Methamphetamine Treatment Project at 3-year follow-up. The
Journal of nervous and mental disease, 197(4), 225.
Glasner-Edwards, S., Mooney, L. J., Marinelli-Casey, P., Hillhouse, M., Ang, A., &
Rawson, R. (2010). Anxiety Disorders among Methamphetamine Dependent
Adults: Association with Post--Treatment Functioning. The American Journal on
Addictions, 19(5), 385-390.
Glasner-Edwards, Suzettee, Mooney, L. J., Marinelli-Casey, Patricia, Hillhouse, M., Ang,
A., & Rawson, R. A. (2010). Psychopathology in methamphetamine-edependent
adults 3 years after treatment. Drug and alcohol review, 29(1), 12-20.
Gluck, M. R., Moy, L. Y., Jayatilleke, E., Hogan, K. A., Manzino, L., & Sonsalla, P. K.
(2001). Parallel increases in lipid and protein oxidative markers in several mouse
brain regions after methamphetamine treatment. Journal of neurochemistry, 79(1),
152-160.
Goldwater, D. S., Pavlides, C., Hunter, R. G., Bloss, E. B., Hof, P. R., McEwen, B. S., &
Morrison, J. H. (2009). Structural and functional alterations to rat medial prefrontal
cortex following chronic restraint stress and recovery. Neuroscience, 164(2), 798-
808.
Gomes da Silva, S., Unsain, N., Mascó, D. H., Toscano‐Silva, M., de Amorim, H. A., Silva
Araujo, B. H., ... & Arida, R. M. (2012). Early exercise promotes positive
hippocampal plasticity and improves spatial memory in the adult life of
rats. Hippocampus, 22(2), 347-358.
Gonul, A. S., Akdeniz, F., Taneli, F., Donat, O., Eker, Ç., & Vahip, S. (2005). Effect of
treatment on serum brain-derived neurotrophic factor levels in depressed
patients. European archives of psychiatry and clinical neuroscience,255(6), 381-
386.
Gonzalez, R., et al. (2004) Neurocognitive performance of methamphetamine users
discordant for history of marijuana exposure. Drug Alcohol Depend. 76, 181-190.
Grace, C. E., Schaefer, T. L., Herring, N. R., Graham, D. L., Skelton, M. R., Gudelsky, G.
A., ... & Vorhees, C. V. (2010). Effect of a neurotoxic dose regimen of (+)-
methamphetamine on behavior, plasma corticosterone, and brain monoamines in
adult C57BL/6 mice. Neurotoxicology and teratology, 32(3), 346-355.
Greenwood, B. N., Spence, K. G., Crevling, D. M., Clark, P. J., Craig, W. C., & Fleshner,
M. (2013). Exercise-induced stress resistance is independent of exercise
controllability and the medial prefrontal cortex. European Journal of Neuroscience.

CHAPTER 6 - References
62
Griebel, G. (1995). 5-Hydroxytryptamine-interacting drugs in animal models of anxiety
disorders: more than 30 years of research. Pharmacology & therapeutics, 65(3),
319-395.
Guerreiro, D. F., Carmo, A. L., da Silva, J. A., Navarro, R., & Góis, C. (2011). Club drugs:
Um Novo Perfil de Abuso de Substâncias em Adolescentes e Jovens Adultos. Acta
Médica Portuguesa, 24(5), 739-56.
Hannon, J., & Hoyer, D. (2008). Molecular biology of 5-HT receptors. Behavioural brain
research, 195(1), 198-213.
Hartig, P. R. (1989). Molecular biology of 5-HT receptors. Trends in pharmacological
sciences, 10(2), 64-69.
Haughey, H. M., Fleckenstein, A. E., Metzger, R. R., & Hanson, G. R. (2000). The effects
of methamphetamine on serotonin transporter activity. Journal of
neurochemistry, 75(4), 1608-1617.
Hillman, C. H., Erickson, K. I., & Kramer, A. F. (2008). Be smart, exercise your heart:
exercise effects on brain and cognition. Nature Reviews Neuroscience,9(1), 58-65.
Hindmarch, I. (2002). Beyond the monoamine hypothesis: mechanisms, molecules and
methods. European Psychiatry, 17, 294-299.
Hirata, H., & Cadet, J. L. (1997). p53-knockout mice are protected against the long-term
effects of methamphetamine on dopaminergic terminals and cell bodies. Journal of
neurochemistry, 69(2), 780-790.
Hirata, H., Ladenheim, B., Rothman, R. B., Epstein, C., & Cadet, J. L. (1995).
Methamphetamine-induced serotonin neurotoxicity is mediated by superoxide
radicals. Brain research, 677(2), 345-347.
Hirschfeld, R. (2000). History and evolution of the monoamine hypothesis of
depression. Journal of Clinical Psychiatry.
Hodges, K., Alto, N. M., Ramaswamy, K., Dudeja, P. K., & Hecht, G. (2008). The
enteropathogenic Escherichia coli effector protein EspF decreases sodium hydrogen
exchanger 3 activity. Cellular microbiology, 10(8), 1735-1745.
Homer, B. D., Solomon, T. M., Moeller, R. W., Mascia, A., DeRaleau, L., & Halkitis, P.
N. (2008). Methamphetamine abuse and impairment of social functioning: a review
of the underlying neurophysiological causes and behavioral implications.
Psychological bulletin, 134(2), 301.
Hopkins, M. E., Nitecki, R., & Bucci, D. J. (2011). Physical exercise during adolescence
versus adulthood: differential effects on object recognition memory and brain-
derived neurotrophic factor levels. Neuroscience, 194, 84-94.

CHAPTER 6 - References
63
Hoyer, D., Hannon, J. P., & Martin, G. R. (2002). Molecular, pharmacological and
functional diversity of 5-HT receptors. Pharmacology Biochemistry and
Behavior, 71(4), 533-554.
HU, G., Pekkarinen, H., HÄNNINEN, O., YU, Z., Guo, Z., & Tian, H. (2002).
Commuting, leisure-time physical activity, and cardiovascular risk factors in
China. Medicine and science in sports and exercise, 34(2), 234-238.
Huang, E. J., & Reichardt, L. F. (2001). Neurotrophins: roles in neuronal development and
function. Annual review of neuroscience, 24, 677.
Hyman, S. E., & Nestler, E. J. (1993). The molecular foundations of psychiatry. American
Psychiatric Pub.
Imam, S. Z., & Ali, S. F. (2001). Aging increases the susceptiblity to methamphetamine-
induced dopaminergic neurotoxicity in rats: correlation with peroxynitrite
production and hyperthermia. Journal of neurochemistry, 78(5), 952-959.
Jackson, E. M., & Dishman, R. K. (2006). Cardiorespiratory fitness and laboratory stress:
A meta-regression analysis. Psychophysiology, 43(1), 57-72.
Jang, C. G., Whitfield, T., Schulteis, G., Koob, G. F., & Wee, S. (2013). A dysphoric-like
state during early withdrawal from extended access to methamphetamine self-
administration in rats. Psychopharmacology, 225(3), 753-763.
Kalivas, P. W., & McFarland, K. (2003). Brain circuitry and the reinstatement of cocaine-
seeking behavior. Psychopharmacology, 168(1-2), 44-56.
Kalueff, A. V., & Tuohimaa, P. (2004). Experimental modeling of anxiety and
depression. Acta neurobiologiae experimentalis, 64(4), 439-448.
Karege, F., Perret, G., Bondolfi, G., Schwald, M., Bertschy, G., & Aubry, J. M. (2002).
Decreased serum brain-derived neurotrophic factor levels in major depressed
patients. Psychiatry research, 109(2), 143-148.
Karila, L., Weinstein, A., Aubin, H. J., Benyamina, A., Reynaud, M., & Batki, S. L.
(2010). Pharmacological approaches to methamphetamine dependence: a focused
review. British journal of clinical pharmacology, 69(6), 578-592.
Katzung, B. G. (Ed.). (2001). Basic & clinical pharmacology (Vol. 8). Toronto:: Lange
Medical Books/McGraw-Hill, 12th
edition - antidepressant agents.
Khulbe, A., Pandey, S., & Sah, S. P. (2013). Antidepressant-like action of the
hydromethanolic flower extract of Tagetes erecta L. in mice and its possible
mechanism of action. Indian journal of pharmacology, 45(4), 386.

CHAPTER 6 - References
64
Kim, D. J., Roh, S., Kim, Y., Yoon, S. J., Lee, H. K., Han, C. S., & Kim, Y. K. (2005).
High concentrations of plasma brain-derived neurotrophic factor in
methamphetamine users. Neuroscience letters, 388(2), 112-115.
Kim, S. J., Lyoo, I. K., Hwang, J., Chung, A., Hoon Sung, Y., Kim, J., ... & Renshaw, P. F.
(2006). Prefrontal grey-matter changes in short-term and long-term abstinent
methamphetamine abusers. The International Journal of
Neuropsychopharmacology, 9(02), 221-228.
Kish, S. J. (2008). Pharmacologic mechanisms of crystal meth. Canadian Medical
Association Journal, 178(13), 1679-1682.
Kish, S. J., Furukawa, Y., Ang, L., Vorce, S. P., & Kalasinsky, K. S. (2000). Striatal
serotonin is depleted in brain of a human MDMA (Ecstasy) user. Neurology, 55(2),
294-296.
Kitamura, O., Tokunaga, I., Gotohda, T., & Kubo, S. I. (2007). Immunohistochemical
investigation of dopaminergic terminal markers and caspase-3 activation in the
striatum of human methamphetamine users. International journal of legal
medicine, 121(3), 163-168.
Kitaoka, R., Fujikawa, T., Miyaki, T., Matsumura, S., Fushiki, T., & Inoue, K. (2010).
Increased noradrenergic activity in the ventromedial hypothalamus during treadmill
running in rats. Journal of nutritional science and vitaminology, 56(3), 185-190.
Kokoshka, J. M., Metzger, R. R., Wilkins, D. G., Gibb, J. W., Hanson, G. R., &
Fleckenstein, A. E. (1998). Methamphetamine treatment rapidly inhibits serotonin,
but not glutamate, transporters in rat brain. Brain research, 799(1), 78-83.
Koob G., Volkow N., 2010. Neurocircuitry of addiction. 35: 217-38.
Koob, G. F., & Le Moal, M. (2005). Plasticity of reward neurocircuitry and the'dark side'of
drug addiction. Nature neuroscience, 8(11), 1442-1444.
Koob, G. F., & Le Moal, M. (2008). Addiction and the brain antireward system. Annu.
Rev. Psychol., 59, 29-53.
Kousik, S. M., Graves, S. M., Napier, T. C., Zhao, C., & Carvey, P. M. (2011).
Methamphetamine-induced vascular changes lead to striatal hypoxia and dopamine
reduction. Neuroreport, 22(17), 923.
Krasnova I. N. and Cadet J. L. (2009) Methamphetamine toxicity and messengers of death.
Brain research reviews, 60: 379-407.
Krishnan, V., & Nestler, E. J. (2008). The molecular neurobiology of
depression. Nature, 455(7215), 894-902.

CHAPTER 6 - References
65
Kujala, U. M., Kaprio, J., & Rose, R. J. (2007). Physical activity in adolescence and
smoking in young adulthood: a prospective twin cohort study. Addiction, 102(7),
1151-1157.
Lampinen, P., Heikkinen, R. L., & Ruoppila, I. (2000). Changes in intensity of physical
exercise as predictors of depressive symptoms among older adults: an eight-year
follow-up. Preventive medicine, 30(5), 371-380.
LaVoie, M. J., Card, J. P., & Hastings, T. G. (2004). Microglial activation precedes
dopamine terminal pathology in methamphetamine-induced neurotoxicity.
Experimental neurology, 187(1), 47-57.
Lawlor, D. A., & Hopker, S. W. (2001). The effectiveness of exercise as an intervention in
the management of depression: systematic review and meta-regression analysis of
randomised controlled trials. BMJ: British Medical Journal, 322(7289), 763.
Lee, B. H., & Kim, Y. K. (2010). The roles of BDNF in the pathophysiology of major
depression and in antidepressant treatment. Psychiatry investigation,7(4), 231-
Lee, R., Kermani, P., Teng, K. K., & Hempstead, B. L. (2001). Regulation of cell survival
by secreted proneurotrophins. Science, 294(5548), 1945-1948.
Lessmann, V., Gottmann, K., & Malcangio, M. (2003). Neurotrophin secretion: current
facts and future prospects. Progress in neurobiology, 69(5), 341-374.
Lewin, G. R., & Barde, Y. A. (1996). Physiology of the neurotrophins. Annual review of
neuroscience, 19(1), 289-317.
Logan, B. K. (2002). Methamphetamine-effects on human performance and behavior.
Forensic Science Review, 14(1), 133-151.
London, E. D., Berman, S. M., Voytek, B., Simon, S. L., Mandelkern, M. A., Monterosso,
J., ... & Ling, W. (2005). Cerebral metabolic dysfunction and impaired vigilance in
recently abstinent methamphetamine abusers. Biological psychiatry, 58(10), 770-
778.
Low, D. A., Vichayanrat, E., Iodice, V., & Mathias, C. J. (2014). Exercise hemodynamics
in Parkinson's disease and autonomic dysfunction. Parkinsonism & related
disorders.
Machado, D. G. (2012). Investigação do potencial antidepressivo de Rosmarinus
officinalis.
Mandyam, C. D., Wee, S., Crawford, E. F., Eisch, A. J., Richardson, H. N., & Koob, G. F.
(2008). Varied access to intravenous methamphetamine self-administration
differentially alters adult hippocampal neurogenesis. Biological psychiatry, 64(11),
958-965.

CHAPTER 6 - References
66
Mandyam, C. D., Wee, S., Crawford, E. F., Eisch, A. J., Richardson, H. N., & Koob, G. F.
(2008). Varied access to intravenous methamphetamine self-administration
differentially alters adult hippocampal neurogenesis. Biological psychiatry, 64(11),
958-965.
Marcus M., Yasamay M. T., Ommeren M., Chisholm D., Saxena S. (2011). Depression: A
global public health concern. WHO, 6-8.
Marcus, B. H., Albrecht, A. E., Niaura, R. S., Taylor, E. R., Simkin, L. R., Feder, S. I., ...
& Thompson, P. D. (1995). Exercise enhances the maintenance of smoking
cessation in women. Addictive Behaviors, 20(1), 87-92.
Martins, T., Baptista, S., Gonçalves, J., Leal, E., Milhazes, N., Borges, F., ... & Silva, A. P.
(2011). Methamphetamine transiently increases the blood–brain barrier
permeability in the hippocampus: Role of tight junction proteins and matrix
metalloproteinase-9. Brain research, 1411, 28-40.
Martins, T., Burgoyne, T., Kenny, B. A., Hudson, N., Futter, C. E., Ambrósio, A. F., ... &
Turowski, P. (2013). Methamphetamine-induced nitric oxide promotes vesicular
transport in blood–brain barrier endothelial cells. Neuropharmacology, 65, 74-82.
Mather, A. S., Rodriguez, C., Guthrie, M. F., McHARG, A. M., Reid, I. C., &
McMURDO, M. E. (2002). Effects of exercise on depressive symptoms in older
adults with poorly responsive depressive disorder Randomised controlled trial. The
British Journal of Psychiatry, 180(5), 411-415.
Mathew, S. J., H. K. Manji, et al. (2008). Novel drugs and therapeutic targets for severe
mood disorders. Neuropsychopharmacology, 33(9), 2080-92.
Matta Mello Portugal, E., Cevada, T., Sobral Monteiro-Junior, R., Teixeira Guimarães, T.,
da Cruz Rubini, E., Lattari, E., ... & Camaz Deslandes, A. (2013). Neuroscience of
Exercise: From Neurobiology Mechanisms to Mental
Health. Neuropsychobiology, 68(1), 1-14.
McAvoy, B. R. (2009). Methamphetamine–what primary care practitioners need to know.
Journal of Primary Health Care, 1(3), 170-176.
McCann, U. D., Szabo, Z., Scheffel, U., Dannals, R. F., & Ricaurte, G. A. (1998). Positron
emission tomographic evidence of toxic effect of MDMA (“Ecstasy”) on brain
serotonin neurons in human beings. The Lancet,352(9138), 1433-1437.
McCann, U. D., Wong, D. F., Yokoi, F., Villemagne, V., Dannals, R. F., & Ricaurte, G. A.
(1998). Reduced striatal dopamine transporter density in abstinent
methamphetamine and methcathinone users: evidence from positron emission
tomography studies with [11C] WIN-35,428. The Journal of neuroscience, 18(20),
8417-8422.

CHAPTER 6 - References
67
Mcfadden, L. M., Hanson, G. R., & Fleckenstein, A. E. (2013). The effects of
methamphetamine self-administration on cortical monoaminergic deficits induced
by subsequent high-dose methamphetamine administrations. Synapse, 67(12), 875-
881.
Mead, G. E., Morley, W., Campbell, P., Greig, C. A., McMurdo, M., & Lawlor, D. A.
(2009). Exercise for depression. Cochrane Database Syst Rev, 3.
Meng, Y., Dukat, M., Troy Bridgen, D., Martin, B. R., & Lichtman, A. H. (1999).
Pharmacological effects of methamphetamine and other stimulants via inhalation
exposure. Drug and Alcohol Dependence, 53(2), 111-120.
Meredith C. W. et al. (2005) Implications of chronic methamphetamine use: a literature
review. Harv Rev Psychiatry, 13:141-154.
Mooney, L. J., Cooper, C., London, E. D., Chudzynski, J., Dolezal, B., Dickerson, D., ... &
Rawson, R. A. (2014). Exercise for methamphetamine dependence: Rationale,
design, and methodology. Contemporary clinical trials, 37(1), 139-147.
Mooney, L. J., Glasner-Edwards, S., Marinelli-Casey, P., Hillhouse, M., Ang, A., Hunter,
J., ... & Rawson, R. (2009). Health conditions in methamphetamine-dependent
adults 3 years after treatment. Journal of addiction medicine, 3(3), 155-163.
Moszczynska, A., & Yamamoto, B. K. (2011). Methamphetamine oxidatively damages
parkin and decreases the activity of 26S proteasome in vivo. Journal of
neurochemistry, 116(6), 1005-1017.
Motl, R. W., Birnbaum, A. S., Kubik, M. Y., & Dishman, R. K. (2004). Naturally
occurring changes in physical activity are inversely related to depressive symptoms
during early adolescence. Psychosomatic Medicine, 66(3), 336-342.
Mowla, S. J., Farhadi, H. F., Pareek, S., Atwal, J. K., Morris, S. J., Seidah, N. G., &
Murphy, R. A. (2001). Biosynthesis and post-translational processing of the
precursor to brain-derived neurotrophic factor. Journal of Biological Chemistry,
276(16), 12660-12666.
Muller, C. P., & Jacobs, B. (Eds.). (2009). Handbook of behavioral neurobiology of
serotonin (Vol. 21). Academic Press.
Murnane, K. S., Perrine, S. A., Finton, B. J., Galloway, M. P., Howell, L. L., &
Fantegrossi, W. E. (2012). Effects of exposure to amphetamine derivatives on
passive avoidance performance and the central levels of monoamines and their
metabolites in mice: Correlations between behavior and neurochemistry.
Psychopharmacology, 220(3), 495-508.
National Institute on Drug Abuse webpages: Commonly Abused
Drugs[http://www.nida.nih.gov/DrugPages/DrugsofAbuse.html]

CHAPTER 6 - References
68
National Institutes of Health (NIH) (2011). Depression. NIH Publication No. 11-3561
Revised.
Neissaar, I., & Raudsepp, L. (2011). Changes in physical activity, self-efficacy and
depressive symptoms in adolescent girls. Pediatric exercise science, 23(3).
Nishida S., 2007. Mecanismos de comunicação entre os neurónios e dos neurónios com os
órgãos efetuadores. Departamento de Fisiologia, IB Unesp-Botucetu. 37-53.
Northrop, N. A., & Yamamoto, B. K. (2012). Persistent neuroinflammatory effects of
serial exposure to stress and methamphetamine on the blood-brain barrier. Journal
of Neuroimmune Pharmacology, 7(4), 951-968.
O'Callaghan, J. P., Sriram, K., & Miller, D. B. (2008). Defining
“neuroinflammation”. Annals of the New York Academy of Sciences, 1139(1), 318-
330.
O'dell, S. J., Galvez, B. A., Ball, A. J., & Marshall, J. F. (2012). Running wheel exercise
ameliorates methamphetamine-induced damage to dopamine and serotonin
terminals. Synapse, 66(1), 71-80.
O'hearn, E., Battaglia, G., De Souza, E. B., Kuhar, M. J., & Molliver, M. E. (1988).
Methylenedioxyamphetamine (MDA) and methylenedioxymethamphetamine
(MDMA) cause selective ablation of serotonergic axon terminals in forebrain:
immunocytochemical evidence for neurotoxicity. The Journal of
neuroscience, 8(8), 2788-2803.
Obrocki, J., Schmoldt, A., Buchert, R., Andresen, B., Petersen, K., & Thomasius, R.
(2002). Specific neurotoxicity of chronic use of ecstasy. Toxicology letters, 127(1),
285-297.
Owens, M. J. (2004). Selectivity of antidepressants: from the monoamine hypothesis of
depression to the SSRI revolution and beyond. Journal of Clinical Psychiatry.
Paluska, S. A., & Schwenk, T. L. (2000). Physical activity and mental health. Sports
medicine, 29(3), 167-180.
Park, M., Hennig, B., & Toborek, M. (2012). Methamphetamine alters occludin expression
via NADPH oxidase-induced oxidative insult and intact caveolae. Journal of
cellular and molecular medicine, 16(2), 362-375.
Paulson, P. E., Camp, D. M., & Robinson, T. E. (1991). Time course of transient
behavioral depression and persistent behavioral sensitization in relation to regional
brain monoamine concentrations during amphetamine withdrawal in
rats. Psychopharmacology, 103(4), 480-492.
Paxinos, G. (2004). The mouse brain in stereotaxic coordinates. Gulf Professional
Publishing.

CHAPTER 6 - References
69
Peirce, J. M., Petry, N. M., Stitzer, M. L., Blaine, J., Kellogg, S., Satterfield, F., ... & Li, R.
(2006). Effects of lower-cost incentives on stimulant abstinence in methadone
maintenance treatment: a National Drug Abuse Treatment Clinical Trials Network
study. Archives of General Psychiatry, 63(2), 201.
Pereira, F. C., Cunha-Oliveira, T., Viana, S. D., Travassos, A. S., Nunes, S., Silva, C., ... &
Ribeiro, C. A. F. (2012). Disruption of striatal glutamatergic/GABAergic
homeostasis following acute methamphetamine in mice. Neurotoxicology and
teratology, 34(5), 522-529.
Pereira, F. C., Lourenço, E. S., Borges, F., Morgadinho, T., Ribeiro, C. F., Macedo, T. R.,
& Ali, S. F. (2006). Single or multiple injections of methamphetamine increased
dopamine turnover but did not decrease tyrosine hydroxylase levels or cleave
caspase-3 in caudate-putamen. Synapse, 60(3), 185-193.
Pereira, F. C., Macedo, T. R., Imam, S. Z., Ribeiro, C. F., & Ali, S. F. (2004). Lack of
hydroxyl radical generation upon central administration of methamphetamine in rat
caudate nucleus: a microdialysis study. Neurotoxicity research, 6(2), 149-152.
Peretti-Watel, P., Guagliardo, V., Verger, P., Pruvost, J., Mignon, P., & Obadia, Y. (2003).
Sporting activity and drug use: alcohol, cigarette and cannabis use among elite
student athletes. Addiction, 98(9), 1249-1256.
Petit-Demouliere, B., Chenu, F., & Bourin, M. (2005). Forced swimming test in mice: a
review of antidepressant activity. Psychopharmacology, 177(3), 245-255.
Picavet, H. S., Wendel-Vos, G. C., Vreeken, H. L., Schuit, A. J., & Verschuren, W. M.
(2011). How stable are physical activity habits among adults? The Doetinchem
Cohort Study. Medicine and science in sports and exercise, 43(1), 74-79.
Ploughman, M. (2008). Exercise is brain food: the effects of physical activity on cognitive
function. Developmental neurorehabilitation, 11(3), 236-240.
Porsolt, R. D., Le Pichon, M., & Jalfre, M. (1977). Depression: a new animal model
sensitive to antidepressant treatments. Nature, 266(5604), 730-732.
Prut, L., & Belzung, C. (2003). The open field as a paradigm to measure the effects of
drugs on anxiety-like behaviors: a review. European journal of
pharmacology, 463(1), 3-33.
Ramirez, S. H., Potula, R., Fan, S., Eidem, T., Papugani, A., Reichenbach, N., ... &
Persidsky, Y. (2009). Methamphetamine disrupts blood–brain barrier function by
induction of oxidative stress in brain endothelial cells. Journal of Cerebral Blood
Flow & Metabolism, 29(12), 1933-1945.
Rawson, R. A., Huber, A., Brethen, P., Obert, J., Gulati, V., Shoptaw, S., & Ling, W.
(2001). Status of methamphetamine users 2–5 years after outpatient
treatment. Journal of Addictive Diseases, 21(1), 107-119.

CHAPTER 6 - References
70
Rawson, R. A., Marinelli-Casey, P., Anglin, M. D., Dickow, A., Frazier, Y., Gallagher, C.,
... & Zweben, J. (2004). A multi-site comparison of psychosocial approaches for the
treatment of methamphetamine dependence. Addiction, 99(6), 708-717.
Rawson, R. A., McCann, M. J., Flammino, F., Shoptaw, S., Miotto, K., Reiber, C., & Ling,
W. (2006). A comparison of contingency management and cognitive-behavioral
approaches for stimulant-dependent individuals. Addiction,101(2), 267-274.
Reback, C. J., Yang, M. X., Rotheram-Fuller, M. E., & Shoptaw, S. (2005). Sustained
reductions in drug use and depression symptoms from treatment for drug abuse in
methamphetamine-dependent gay and bisexual men. Journal of Urban
Health, 82(1), i100-i108.
Reneman, L., Booij, J., de Bruin, K., Reitsma, J. B., de Wolff, F. A., Gunning, W. B., ... &
van den Brink, W. (2001). Effects of dose, sex, and long-term abstention from use
on toxic effects of MDMA (ecstasy) on brain serotonin neurons. The
lancet, 358(9296), 1864-1869.
Rimmele, U., Seiler, R., Marti, B., Wirtz, P. H., Ehlert, U., & Heinrichs, M. (2009). The
level of physical activity affects adrenal and cardiovascular reactivity to
psychosocial stress. Psychoneuroendocrinology, 34(2), 190-198.
Rimmele, U., Zellweger, B. C., Marti, B., Seiler, R., Mohiyeddini, C., Ehlert, U., et al.
(2007) Trained men show lower cortisol, heart rate and psychological responses to
psychosocial stress compared with untrained men. Psychoneuroendocrinology. 32,
627-635.
Roll, J., Petry, N., Stitzer, M., Brecht, M., Peirce, J., McCann, M., ... & Kellogg, S. (2006).
Contingency management for the treatment of methamphetamine use
disorders. American Journal of Psychiatry, 163(11), 1993-1999.
Russo-Neustadt, A., Ha, T., Ramirez, R., & Kesslak, J. P. (2001). Physical activity-
antidepressant treatment combination: Impact on brain-derived neurotrophic factor
and behavior in an animal model. Behavioural brain research.
Rusyniak, D. E. (2011). Neurologic manifestations of chronic methamphetamine
abuse. Neurologic clinics, 29(3), 641.
Sailasuta, N., Abulseoud, O., Harris, K. C., & Ross, B. D. (2010). Glial dysfunction in
abstinent methamphetamine abusers. Journal of Cerebral Blood Flow &
Metabolism, 30(5), 950-960.
Sari, Y. (2004). Serotonin 1B receptors: from protein to physiological function and
behavior. Neuroscience & Biobehavioral Reviews, 28(6), 565-582.
Sasco, A. ., Laforest, L., Benha m-Luzon, V., Poncet, M., & Little, R. E. (2002). Smoking
and its correlates among preadolescent children in France. Preventive
medicine, 34(2), 226-234.

CHAPTER 6 - References
71
Schepers, R. J., Oyler, J. M., Joseph, R. E., Cone, E. J., Moolchan, E. T., & Huestis, M. A.
(2003). Methamphetamine and amphetamine pharmacokinetics in oral fluid and
plasma after controlled oral methamphetamine administration to human volunteers.
Clinical chemistry, 49(1), 121-132.
Schumann, A., Hapke, U., RUMPF, H. J., Meyer, C., & John, U. (2001). The association
between degree of nicotine dependence and other health behaviours Findings from
a German general population study. The European Journal of Public Health, 11(4),
450-452.
Sekine, Y., Ouchi, Y., Takei, N., Yoshikawa, E., Nakamura, K., Futatsubashi, M., ... &
Mori, N. (2006). Brain serotonin transporter density and aggression in abstinent
methamphetamine abusers. Archives of General Psychiatry, 63(1), 90-00.
Sharp, T., & Cowen, P. J. (2011). 5-HT and depression: is the glass half-full? Current
opinion in pharmacology, 11(1), 45-51.
Shimizu, E., Hashimoto, K., Okamura, N., Koike, K., Komatsu, N., Kumakiri, C., ... & Iyo,
M. (2003). Alterations of serum levels of brain-derived neurotrophic factor (BDNF)
in depressed patients with or without antidepressants. Biological psychiatry, 54(1),
70-75.
Silva, C. D., Neves, A. F., Dias, A. I., Freitas, H. J., Mendes, S. M., Pita, I., ... & Pereira, F.
C. (2013). A single neurotoxic dose of methamphetamine induces a long-lasting
depressive-like behaviour in mice. Neurotoxicity research, 1-10.
Silverstein, P. S., Shah, A., Gupte, R., Liu, X., Piepho, R. W., Kumar, S., & Kumar, A.
(2011). Methamphetamine toxicity and its implications during HIV-1
infection. Journal of neurovirology, 17(5), 401-415.
Simon, S. L., Domier, C., Carnell, J., Brethen, P., Rawson, R., & Ling, W. (2000).
Cognitive impairment in individuals currently using methamphetamine.The
American Journal on Addictions, 9(3), 222-231.
Singh, N. A., Clements, K. M., & Singh, M. A. F. (2001). The Efficacy of Exercise as a
Long-term Antidepressant in Elderly Subjects A Randomized, Controlled Trial.The
Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 56(8),
M497-M504.
Sjösten, N., & Kivelä, S. L. (2006). The effects of physical exercise on depressive
symptoms among the aged: a systematic review. International journal of geriatric
psychiatry, 21(5), 410-418.
Smith, A. D., & Zigmond, M. J. (2003). Can the brain be protected through exercise?
Lessons from an animal model of parkinsonism☆. Experimental Neurology, 184(1),
31-39.

CHAPTER 6 - References
72
Sobel, B. P. (2001). Bingo vs. physical intervention in stimulating short-term cognition in
Alzheimer's disease patients. American journal of Alzheimer's disease and other
dementias, 16(2), 115-120.
Spalding, T. W., Lyon, L. A., Steel, D. H., & Hatfield, B. D. (2004). Aerobic exercise
training and cardiovascular reactivity to psychological stress in sedentary young
normotensive men and women. Psychophysiology, 41(4), 552-562.
Stahl, S. M. (1998). Mechanism of action of serotonin selective reuptake inhibitors:
serotonin receptors and pathways mediate therapeutic effects and side
effects. Journal of affective disorders, 51(3), 215-235.
Stathopoulou, G., Powers, M. B., Berry, A. C., Smits, J. A., & Otto, M. W. (2006).
Exercise interventions for mental health: a quantitative and qualitative
review. Clinical Psychology: Science and Practice, 13(2), 179-193.
Stranahan, A. M., Lee, K., & Mattson, M. P. (2008). Central mechanisms of HPA axis
regulation by voluntary exercise. Neuromolecular medicine, 10(2), 118-127.
Strawbridge, W. J., Deleger, S., Roberts, R. E., & Kaplan, G. A. (2002). Physical activity
reduces the risk of subsequent depression for older adults. American Journal of
Epidemiology, 156(4), 328-334.
Ströhle, A. (2009). Physical activity, exercise, depression and anxiety disorders. Journal of
neural transmission, 116(6), 777-784.
Strong, W. B., Malina, R. M., Blimkie, C. J., Daniels, S. R., Dishman, R. K., Gutin, B., ...
& Trudeau, F. (2005). Evidence based physical activity for school-age youth. The
Journal of pediatrics, 146(6), 732-737.
Sulzer, D., Sonders, M. S., Poulsen, N. W., & Galli, A. (2005). Mechanisms of
neurotransmitter release by amphetamines: a review. Progress in neurobiology,
75(6), 406-433.
Sutcliffe, C. G., German, D., Sirirojn, B., Latkin, C., Aramrattana, A., Sherman, S. G., &
Celentano, D. D. (2009). Patterns of methamphetamine use and symptoms of
depression among young adults in northern Thailand. Drug and alcohol
dependence, 101(3), 146-151.
Takemura, Y., Sakurai, Y., Inaba, Y., & Kugai, N. (2000). A cross-sectional study on the
relationship between leisure or recreational physical activity and coronary risk
factors. Tohoku Journal of Experimental Medicine, 192(3), 227-237.
Taylor, A. H., Ussher, M. H., & Faulkner, G. (2007). The acute effects of exercise on
cigarette cravings, withdrawal symptoms, affect and smoking behaviour: a
systematic review. Addiction, 102(4), 534-543.

CHAPTER 6 - References
73
Thomas, D. M., Dowgiert, J., Geddes, T. J., Francescutti-Verbeem, D., Liu, X., & Kuhn,
D. M. (2004b). Microglial activation is a pharmacologically specific marker for the
neurotoxic amphetamines. Neuroscience letters, 367(3), 349-354.
Thomas, D. M., Walker, P. D., Benjamins, J. A., Geddes, T. J., & Kuhn, D. M. (2004a).
Methamphetamine neurotoxicity in dopamine nerve endings of the striatum is
associated with microglial activation. Journal of Pharmacology and Experimental
Therapeutics, 311(1), 1-7.
Thompson, P. M., Hayashi, K. M., Simon, S. L., Geaga, J. A., Hong, M. S., Sui, Y., ... &
London, E. D. (2004). Structural abnormalities in the brains of human subjects who
use methamphetamine. The Journal of neuroscience, 24(26), 6028-6036.
Timar, J., Gyarmati, S., Szabo, A., & Fürst, S. (2003). Behavioural changes in rats treated
with a neurotoxic dose regimen of dextrorotatory amphetamine
derivatives. Behavioural pharmacology, 14(3), 199-206.
Toborek, M., Seelbach, M. J., Choi, Y. J., Andras, I. E., Chen, L., Hennig, B., & Esser, K.
A. (2009). Voluntary exercise protects against methamphetamine-induced oxidative
stress in brain microvasculature and disruption of the blood-brain barrier. The
FASEB Journal.
Torres, G. E., Gainetdinov, R. R., & Caron, M. G. (2003). Plasma membrane monoamine
transporters: structure, regulation and function. Nature Reviews Neuroscience, 4(1),
13-25.
Trivedi, M. H., Greer, T. L., Grannemann, B. D., Church, T. S., Galper, D. I., Sunderajan,
P., ... & Carmody, T. J. (2006). TREAD: TReatment with Exercise Augmentation
for Depression: study rationale and design. Clinical Trials, 3(3), 291-305.
Tsuang, M. T., Bar, J. L., Stone, W. S., & Faraone, S. V. (2004). Gene-environment
interactions in mental disorders. World Psychiatry, 3(2), 73.
Turnipseed, S. D., Richards, J. R., Kirk, J. D., Diercks, D. B., & Amsterdam, E. A. (2003).
Frequency of acute coronary syndrome in patients presenting to the emergency
department with chest pain after methamphetamine use. The Journal of emergency
medicine, 24(4), 369-373.
UNODC, World Drug Report. (2012). United Nations publication, Sales No. E.12.XI.1.
UNODC, World Drug Report. (2013). United Nations publication, Sales No. E.13.XI.6.
van der Staay, F. J., Arndt, S. S., & Nordquist, R. E. (2009). Evaluation of animal models
of neurobehavioral disorders. Behavioral and Brain Functions,5(1), 11.
van Praag, H. (2008). Neurogenesis and exercise: past and future directions.
Neuromolecular medicine, 10(2), 128-140.

CHAPTER 6 - References
74
Van Praag, H. M. (2001). Past expectations, present disappointments, future hopes or
psychopathology as the rate-limiting step of progress in
psychopharmacology. Human Psychopharmacology: Clinical and Experimental,
16(1), 3-7.
Van Rensburg, K. J., Taylor, A., & Hodgson, T. (2009). The effects of acute exercise on
attentional bias towards smoking-related stimuli during temporary abstinence from
smoking. Addiction, 104(11), 1910-1917.
Volkow, N. D., Chang, L., Wang, G. J., Fowler, J. S., Leonido-Yee, M., Franceschi, D., ...
& Miller, E. N. (2001). Association of dopamine transporter reduction with
psychomotor impairment in methamphetamine abusers. American Journal of
Psychiatry, 158(3), 377-382.
Walsh, R. N., & Cummins, R. A. (1976). The open-field test: A critical review.
Psychological bulletin, 83(3), 482.
Warburton, D. E., Nicol, C. W., & Bredin, S. S. (2006). Health benefits of physical
activity: the evidence. Canadian medical association journal, 174(6), 801-809.
Willner, P. (2005). Chronic mild stress (CMS) revisited: consistency and behavioural-
neurobiological concordance in the effects of CMS. Neuropsychobiology, 52(2),
90-110.
Wilson, J. M., Kalasinsky, K. S., Levey, A. I., Bergeron, C., Reiber, G., Anthony, R. M., ...
& Kish, S. J. (1996). Striatal dopamine nerve terminal markers in human, chronic
methamphetamine users. Nature medicine, 2(6), 699-703.
Woods, J. A., Ceddia, M. A., Zack, M. D., Lowder, T. W., & Lu, Q. (2003). Exercise
training increases the näive to memory T cell ratio in old mice. Brain, behavior,
and immunity, 17(5), 384-392.
Xu, W., Zhu, J. P., & Angulo, J. A. (2005). Induction of striatal pre-and postsynaptic
damage by methamphetamine requires the dopamine receptors. Synapse, 58(2),
110-121.
Yalcin, I., Aksu, F., & Belzung, C. (2005). Effects of desipramine and tramadol in a
chronic mild stress model in mice are altered by yohimbine but not by
pindolol. European journal of pharmacology, 514(2), 165-174.
Youngstedt, S. D. (2005). Effects of exercise on sleep. Clinics in sports medicine, 24(2),
355-365.
Zhu, J. P., Xu, W., & Angulo, J. A. (2005). Disparity in the temporal appearance of
methamphetamine-induced apoptosis and depletion of dopamine terminal markers
in the striatum of mice. Brain research, 1049(2), 171-181.

CHAPTER 6 - References
75
Zweben, J. E., Cohen, J. B., Christian, D., Galloway, G. P., Salinardi, M., Parent, D., &
Iguchi, M. (2004). Psychiatric symptoms in methamphetamine users. The American
Journal on Addictions, 13(2), 181-190.