Embed Size (px)
Citation preview

Pest Management Science Pest Manag Sci 61:917–921 (2005)DOI: 10.1002/ps.1097
Degradation of simazine by microorganismsisolated from soils of Spanish olive fields†
Raquel Santiago-Mora,1∗ Fabrice Martin-Laurent,2 Rafael de Prado 3
and Antonio R Franco1
1Dpto de Bioquımica y Biologıa Molecular, Universidad de Cordoba, Campus de Rabanales, 14071 Cordoba, Spain2Microbiologie et Geochimie des Sols, UMR 1229 INRA/Universite de Bourgogne INRA/CMSE, Dijon, France3Dpto de Quımica Agrıcola y Edafologıa, Universidad de Cordoba, Campus de Rabanales, 14071 Cordoba, Spain
Abstract: The capability of the microbial flora isolated from an olive field soil from Andalusia to mineralizesimazine has been analyzed. From this soil, a group of bacteria capable of degrading 60 mg simazine litre−1
in less than a week has been isolated. These microorganisms showed a low capacity for degrading thisherbicide to carbon dioxide. When total DNA was isolated from this group of bacteria, we were ableto detect by PCR the presence of only the atzC and the trzN genes. Some components of this bacterialpopulation have been identified by sequencing of specific fragments from bacterial 16S rDNA, includingVariovorax sp, Pseudoxanthomonas mexicana Thierry et al, Acidovorax sp and Methylopila capsulataDoronina et al. These data suggest that this consortium of bacteria performs an incomplete degradationof the simazine 2005 Society of Chemical Industry
Keywords: simazine; microbial degradation; soil
1 INTRODUCTIONSimazine (6-chloro-N2,N4-diethyl-1,3,5-triazine-2,4-diamine) is an s-triazine herbicide used for controllingbroad-leaved weeds in various crops. This herbicidehas a moderate persistence in the soil, and both itswidespread use and its mobility have led to tracesof simazine in rivers,1 groundwaters2 and soils.3 Theeffects of the simazine on living organisms have beenstudied, although it remains a matter of debate whetherthe accumulation of simazine or its derivatives is anecological risk.4–6
For many decades, simazine was extensively used onthe olive fields in Andalusia (south Spain). However,in January 2003, an European Union directive forbadethe application of this herbicide in this crop.7,8
Simazine can be degraded in the soil with a half-lifeencompassing a few days or several months, dependingon the soil characteristics and the number of previousapplications of the herbicide.9,10 Degradation ofsimazine relies on both biotic and abiotic processes,11
but microorganisms are mainly responsible for thecomplete or partial degradation of the herbicide.12
Gram-positive and -negative bacteria,13 together withsome fungi that are able to use s-triazine herbicidesas a carbon and/or a nitrogen source for growthare involved in this degradation. Some isolates of
these microorganisms can assimilate simazine bythemselves, while others need to be components ofmicro-organism consortia to degrade it.14 The best-known degradation pathway of an s-triazine herbicideis the degradation of atrazine by Pseudomonas ADP.15
In this mineralization process the atrazine is degradedby six enzymes encoded by the plasmidic atz familyof genes. Some variations of this pathway have beendescribed. In Nocardioides sp a new s-triazine hydrolaseenzyme, C190, which is encoded by the trzN genehas been discovered,16 and in Rhodococcus strainss-triazines are degraded by another set of enzymesencoded by the trz gene family.17
This work was conducted to show and analyze thecapacity of soil microflora from an olive soil fromAndalusia (Spain) to mineralize simazine. Severalmicroorganisms which can degrade simazine wereidentified. The degradation rate, the genes involvedand the possible degradation pathway were analyzed.
2 MATERIALS AND METHODS2.1 Soil samplesThe soil was collected from the surface (0–15 cm) ofa plot in the olive fields called Hornerico, in the areaof Baena, Cordoba (Andalusia, south Spain). This
∗ Correspondence to: Raquel Santiago-Mora, Dpto de Bioquımica y Biologıa Molecular, Universidad de Cordoba, Campus de Rabanales,14071 Cordoba, SpainE-mail: [email protected]†Based on a paper presented at the 4th International Symposium on Environmental Aspects of Pesticide Microbiology: Pesticides, MicrobialFunctions and Biodiversity in Soil, held at Thessaloniki, Greece on 4–9 September 2004Contract/grant sponsor: Junta de Andalucıa (Spain); contract/grant number: CAO01-027(Received 26 November 2004; revised version received 31 March 2005; accepted 26 May 2005)Published online 8 July 2005
2005 Society of Chemical Industry. Pest Manag Sci 1526–498X/2005/$30.00 917
R Santiago-Mora et al
soil had been repeatedly treated with simazine formany years. Soil samples were partially dried, sievedto 2 mm, and stored at 4 ◦C until its use.
2.2 Soil mineralization assaysThe capability of this soil to biodegrade simazinewas assessed in triplicate by a mineralization assay.Samples (10 g) of soil received 1 ml of an aqueoussolution containing [ring-U-14C]-simazine (specificactivity ≥185 GBq mol−1, purity >95%; Sigma) andunlabelled simazine. At the beginning of this assay,each sample received 814 Bq and the equivalentamount of unlabelled simazine, up to 1.5 mg kg−1
soil. The same amount of unlabelled simazine wasadded every 28 days throughout the entire assay. Thevessel with the soil was placed in a 1-litre hermeticjar together with a vial containing aqueous sodiumhydroxide (0.2 M; 5 ml) that trapped any [14C]-carbondioxide resulting from the mineralization of simazine.The jars were incubated at 20 ◦C in the dark. Thealkaline solution was replaced periodically, and theevolved [14C]-carbon dioxide was measured in aliquid scintillation counter (Packard) using 5 ml ofAC-SII scintillation fluid (Amersham). The data wereprocessed and statistically analyzed using the programSigmaPlot version 8.0.
2.3 Isolation of simazine-degradingmicroorganismsA simazine enrichment procedure was used to isolatethe microorganisms responsible for degrading theherbicide. Soil (10 g) was added to 90 ml of minimumsalt medium18 containing 60 mg simazine litre−1.The mixture was incubated at 25 ◦C with maximumshaking, to prevent the soil sedimentation, and inthe dark. New cultures were made by dilutionof the original medium into fresh medium every15 days, until a final dilution of 10−5 relative tothe first medium was reached. Aliquots from the soilsolutions (10−3 to 10−5) were plated on minimum saltmedium supplemented with traces of vitamins (HCl-thiamine and biotin) and agar 16 g, sodium citrate1 g, cycloheximide 0.1 g and simazine (that formed achalky suspension) 2 g per litre of medium. Coloniesthat grew on plates were inoculated in minimum saltliquid medium with 60 mg simazine litre−1. The samemedium without inoculation was used as a control.The incubations conditions were 25 ◦C, 120 rpm anddark. Simazine degradation was assessed by periodicanalysis of medium by HPLC. The chromatographymethod used was that described previously.19 Thismethod allows the detection and quantification ofseveral s-triazine herbicides. In this case, the run timewas changed from 32 to 10 min.
2.4 PCR amplification of genes involved insimazine degradationBacterial DNA was isolated by breaking the bacteriacells and digesting for 2 h with 1mg litre−1 of proteinaseK in Tris-HCl (0.5 M; pH 8) at 55 ◦C. The enzyme was
then inactivated by heating the mixture at 80 ◦C for5 min. After centrifugation at 13 000 rpm for 10 min,the cellular debris was discarded and the supernatantswith the DNA were kept at −20 ◦C until use.
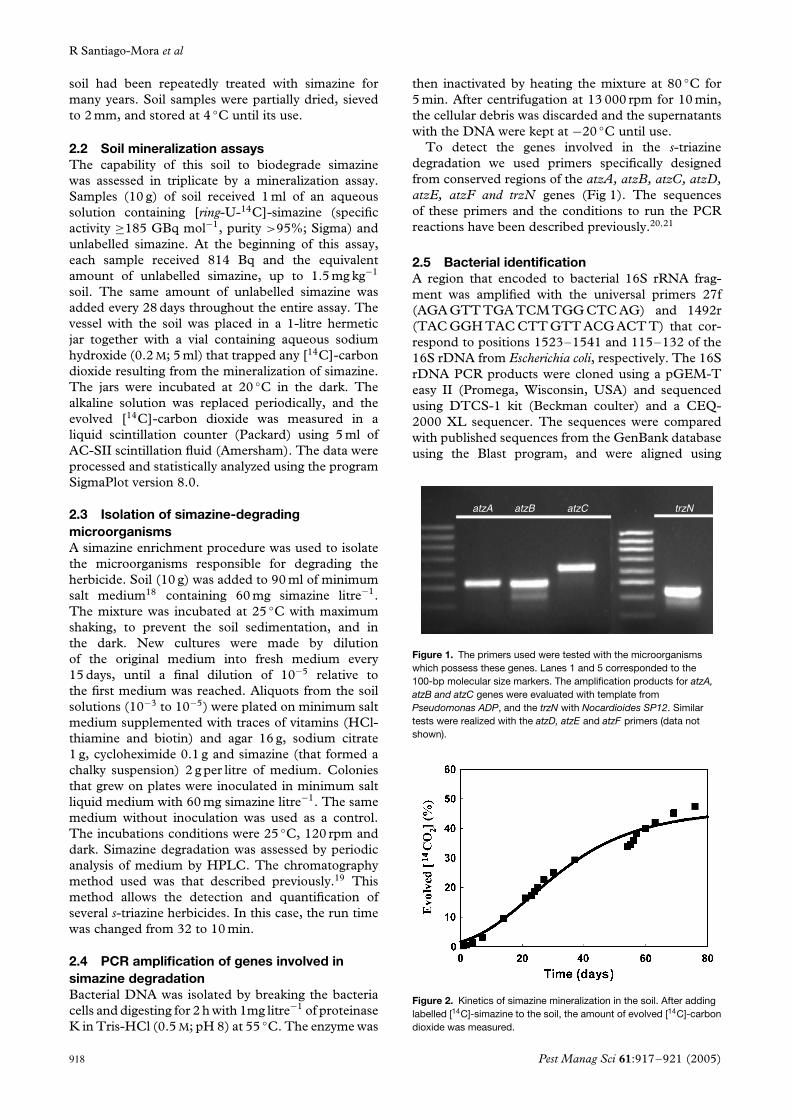
To detect the genes involved in the s-triazinedegradation we used primers specifically designedfrom conserved regions of the atzA, atzB, atzC, atzD,atzE, atzF and trzN genes (Fig 1). The sequencesof these primers and the conditions to run the PCRreactions have been described previously.20,21
2.5 Bacterial identificationA region that encoded to bacterial 16S rRNA frag-ment was amplified with the universal primers 27f(AGA GTT TGA TCM TGG CTC AG) and 1492r(TAC GGH TAC CTT GTT ACG ACT T) that cor-respond to positions 1523–1541 and 115–132 of the16S rDNA from Escherichia coli, respectively. The 16SrDNA PCR products were cloned using a pGEM-Teasy II (Promega, Wisconsin, USA) and sequencedusing DTCS-1 kit (Beckman coulter) and a CEQ-2000 XL sequencer. The sequences were comparedwith published sequences from the GenBank databaseusing the Blast program, and were aligned using
atzA atzB atzC trzN
Figure 1. The primers used were tested with the microorganismswhich possess these genes. Lanes 1 and 5 corresponded to the100-bp molecular size markers. The amplification products for atzA,atzB and atzC genes were evaluated with template fromPseudomonas ADP, and the trzN with Nocardioides SP12. Similartests were realized with the atzD, atzE and atzF primers (data notshown).
Figure 2. Kinetics of simazine mineralization in the soil. After addinglabelled [14C]-simazine to the soil, the amount of evolved [14C]-carbondioxide was measured.
918 Pest Manag Sci 61:917–921 (2005)
Degradation of simazine by microorganisms from olive field soils
the ClustawX (v1.81) program. The phylogeneticanalyses were conducted with TreeView (v1.6.6). Theaccession numbers of the sequences are AY692 427,AY692 428, AY692 429 and AY692 430
3 RESULTS AND DISCUSSION3.1 Simazine soil mineralizationApproximately 40% of the simazine added to thesoil was completely degraded to carbon dioxide after80 days incubation (Fig 2). The data clearly fittedto a Gompertz sigmoid curve (R2 = 0.98).22 Themineralization constant in this soil was estimatedas 18.8 day−1. These results are in accordance witha mineralization kinetic typical of soils adaptedto the herbicide, and they suggest the existenceof endogenous microflora with the capability ofdegrading simazine.
3.2 Isolation of a simazine-degradingconsortiumAfter the enrichment procedure, different bacteriawere isolated and tested for their capability of
Figure 3. Degradation of simazine by the bacterial consortium inliquid medium. The simazine remaining in the medium was measured(°) in the negative control medium without bacteria or (ž) inoculatedwith the consortium of bacteria.
Figure 4. Kinetics of mineralization of simazine by the bacterialconsortium. These experiments were done in liquid medium, and theevolved [14C]-carbon dioxide was measured.
degrading simazine. Among these bacteria we selecteda bacterial consortium that could degrade simazine ata rate substantially faster than the rest of the isolatedbacteria (data not shown). Moreover, this populationgrew with simazine as a sole nitrogen and carbonsource. After 6 days of incubation, no simazine wasdetected by HPLC in the liquid medium (Fig 3).However, the mineralization assays showed that they
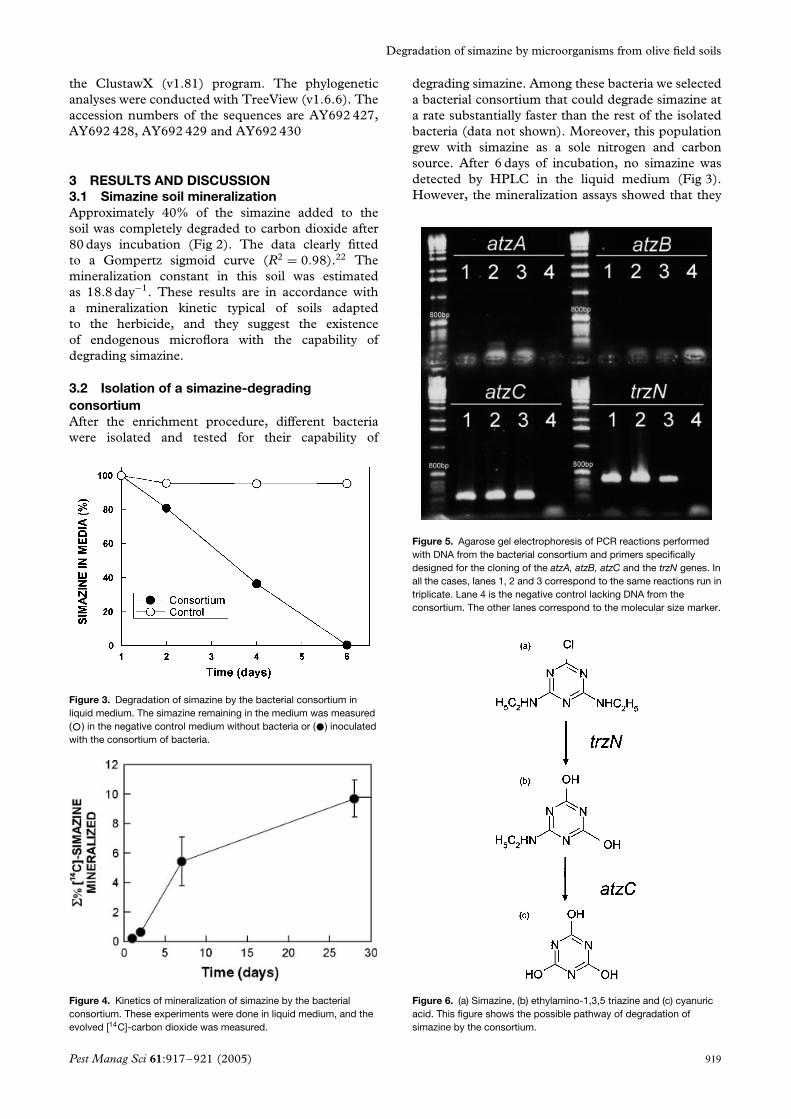
Figure 5. Agarose gel electrophoresis of PCR reactions performedwith DNA from the bacterial consortium and primers specificallydesigned for the cloning of the atzA, atzB, atzC and the trzN genes. Inall the cases, lanes 1, 2 and 3 correspond to the same reactions run intriplicate. Lane 4 is the negative control lacking DNA from theconsortium. The other lanes correspond to the molecular size marker.
Figure 6. (a) Simazine, (b) ethylamino-1,3,5 triazine and (c) cyanuricacid. This figure shows the possible pathway of degradation ofsimazine by the consortium.
Pest Manag Sci 61:917–921 (2005) 919

R Santiago-Mora et al
Figure 7. Phylogenetic analysis of 16S rDNA sequences obtained using template DNA from bacteria isolated from the consortium (underlinednames). The sequences of the bacteria composing the consortium were compared with the 16 rDNA sequences from Pseudomonas sp ADP(AF326383); Agrobacterium sp (AY196 141), Rhizobium sp (AF514801), Rhodococcus sp (AY336 559), Nocardioides sp C190 (AF253510) andNocardioides sp SP12 (AF537327).
had a low capacity to break down the simazine ringsince, after 28 days of incubation, only around 10% ofthe labelled simazine evolved to [14C]-carbon dioxide(Fig 4). This suggests that this bacterial consortiumprobably uses the ethylamino side-chain of simazinefor growth.
3.3 PCR amplification of genes involvedin simazine degradationPCR amplified bands with template DNA from thebacterial consortium were only obtained when primerstargeting for the atzC and tzrN genes were used. Noamplification products were obtained with atzA, atzB,atzD atzE and atzF primers (Fig 5). To date, thetrzN gene has been detected in Nocardioides (strainsC190 and SP12).16,21 None of these Nocardioidesstrains can completely degrade the s-triazine ringto carbon dioxide, and have a different set ofcomplementary degrading genes. Indeed, Nocardioidessp C190 lacks the atzA, atzB or atzC genes, whereasNocardioides SP12 contains the atzB, atzC and trzNgenes. The enzyme encoded by the trzN gene hasboth clorohydrolase and desaminase activities.23 Theother gene detected in the consortium, atzC, encodesan enzyme with isopropylammelide isopropylaminehydrolase activity that can hydrolyze the N-ethylaminogroup of simazine.24
Both the activities of the trzN product and thehydrolase activity of the enzyme encoded by atzCsuggest that the consortium could first degradesimazine by the action of TRZN enzyme to renderethylamino-1,3,5 triazine, which could then betransformed to cyanuric acid by the ATZC enzyme.(Fig 6).
3.4 Identification of the bacterial consortiumThe 16S rDNA PCR products were cloned andsequenced, four different sequences of 809, 1487,1464 and 1445 bp were obtained. These sequenceswere compared with those available in the Genbankdatabase, using the Blast program. The sequences of
809, 1464 and 1487 bp were more than 98% identicalto those of Variovorax sp, Pseudoxanthomonas mexicanaThierry et al and Acidovorax sp 16S rDNA sequences,respectively. The 1445-bp sequence was 95% identicalto that of the Methylopila capsulata Doronina et al16s rDNA sequence.
These sequences were aligned and phylogeneticallyanalyzed with 16S rDNA sequences from others-triazine-degrading bacteria (Fig 7). The speciesfrom the bacterial consortium fit into three differentbranches in the phylogenetical tree. Acidovorax sp andVariovorax sp aligned closely and P mexicana is closeto the Pseudomonas ADP sequence.
ACKNOWLEDGEMENTSThis project is supported by the grant CAO01-027of Junta de Andalucıa (Spain). DNA sequences wererealized using the facilities offered by the Service ofSequencing and Genotyping (SSG).
REFERENCES1 Lopez-Flores R, Quintana XD, Salvado V, Hidalgo M, Sala L
and Moreno-Amich R, Comparison of nutrient and contam-inant fluxes in two areas with different hydrological regimes(Emporda Wetlands, NE Spain). Water Res 37:3034–3046(2003).
2 Vandecasteele K, Gaus I, Debreuck W and Walraevens K, Iden-tification and quantification of 77 pesticides in groundwaterusing solid phase coupled to liquid-liquid microextrac-tion and reversed-phase liquid chromatography. Anal Chem72:3093–3101 (2000).
3 Cogger CG, Bristow PR, Stark JD, Getzin LW and Mont-gomery M, Transport and persistence of pesticides in alluvialsoils: I. Simazine. J Environ Qual 27:543–550 (1998).
4 Kligerman AD, Doerr CL, Tennant AH and Zucker RM,Cytogenetic studies of three triazine herbicides. I. In vitrostudies. Mutat Res 465:53–59 (2000).
5 Tennant AH, Peng B and Kligerman AD, Genotoxicity studiesof three triazine herbicides: in vivo studies using the alkalinesingle cell gel (SCG) assay. Mutat Res 493:1–10 (2001).
6 Gojmerac T, Uremovic M, Uremovic Z, Curic S andBilandzic N, Reproductive disturbance caused by anS-triazine herbicide in pigs. Acta Vet Hung 47:129–135(1999).
920 Pest Manag Sci 61:917–921 (2005)

Degradation of simazine by microorganisms from olive field soils
7 Council directive 91/414/EEC, concerning the registration andselling of Plant Protection Products. http://europa.eu.int/eur-lex/
8 Commission regulation (EC) No 451/2000. http://europa.eu.int/eur-lex/
9 Kristensen GB, Johannesen H and Aamand J, Mineralizationof aged atrazine and mecoprop in soil and aquifer chalk.Chemosphere 45:927–934 (2001).
10 Abdelhafid SHEB, How increasing availability of carbon andnitrogen affect atrazine behaviour in soils. Biol Fertil Soils30:333–340 (2000).
11 Evgenidou E and Fytianos K, Photodegradation of triazineherbicides in aqueous solutions and natural waters. J AgricFood Chem 50:6423–6427 (2002).
12 Strandberg MT and Scott-Fordsmand JJ, Field effects ofsimazine at lower trophic levels—a review. Sci Total Environ296:117–137 (2002).
13 Rousseaux S, Hartmann A and Soulas G, Isolation and charac-terisation of new Gram-negative and Gram-positive atrazinedegrading bacteria from different French soils. FEMS Micro-biol Ecol 36:211–222 (2001).
14 Mandelbaum RT, Wackett LP and Allan DL, Mineralizationof the s-triazine ring of atrazine by stable bacterialmixed cultures. Appl Environ Microbiol 59:1695–1701(1993).
15 Martinez B, Tomkins J, Wackett LP, Wing R and Sad-owsky MJ, Complete nucleotide sequence and organizationof the atrazine catabolic plasmid pADP-1 from Pseudomonassp strain ADP. J Bacteriol 183:5684–5697 (2001).
16 Topp E, Mulbry WM, Zhu H, Nour SM and Cuppels D,Characterization of S-triazine herbicide metabolism by aNocardioides sp isolated from agricultural soils. Appl EnvironMicrobiol 66:3134–3141 (2000).
17 Behki R, Topp E, Dick W and Germon P, Metabolism ofthe herbicide atrazine by Rhodococcus strains. Appl EnvironMicrobiol 59:1955–1959 (1993).
18 Topp E, Zhu H, Nour SM, Houot S, Lewis M and Cuppels D,Characterization of an atrazine-degrading Pseudaminobactersp isolated from Canadian and French agricultural soils. ApplEnviron Microbiol 66:2773–2782 (2000).
19 Martinez MJ, Dıaz MA and Pena FdPR, Extraccion y detr-minacion de triazinas por cromatografıa lıquida de altaresolucion (HPLC), in Uso de herbicidas en la agriculturadel siglo XXI, ed by de Prado R and Jorrın VJ. Servicio depublicaciones de la Universidad de Cordoba, pp 665–663(2001).
20 Martin-Laurent F, Piutti S, Hallet S, Wagschal I, Philippot L,Catroux G and Soulas G, Monitoring of atrazine treatmenton soil bacterial, fungal and atrazine-degrading communitiesby quantitative competitive PCR. Pest Manag Sci 59:259–268(2003).
21 Piutti S, Semon E, Landry D, Hartmann A, Dousset S, Licht-fouse E, Topp E, Soulas G and Martin-Laurent F, Isolationand characterisation of Nocardioides sp SP12, an atrazine-degrading bacterial strain possessing the gene trzN from bulk-and maize rhizosphere soil. FEMS Microbiol Lett 221:111–117(2003).
22 Rousseaux S, Hartmann A, Lagacherie B, Piutti S, Andreux Fand Soulas G, Inoculation of an atrazine-degrading strain,Chelatobacter heintzii Cit1, in four different soils: effectsof different inoculum densities. Chemosphere 51:569–576(2003).
23 Mulbry WW, Zhu H, Nour SM and Topp E, The triazinehydrolase gene trzN from Nocardioides sp strain C190:cloning and construction of gene-specific primers. FEMSMicrobiol Lett 206:75–79 (2002).
24 Sadowsky MJ, Tong Z, de SM and Wackett LP, AtzC is anew member of the amidohydrolase protein superfamily andis homologous to other atrazine-metabolizing enzymes. JBacteriol 180:152–158 (1998).
Pest Manag Sci 61:917–921 (2005) 921





























