Embed Size (px)
Citation preview

Biochimica et Biophysica Acta 1777 (2008) 1370–1377
Contents lists available at ScienceDirect
Biochimica et Biophysica Acta
j ourna l homepage: www.e lsev ie r.com/ locate /bbab io
Defective assembly of a hybrid vacuolar H+-ATPase containing the mousetestis-specific E1 isoform and yeast subunits
Kazuhiro Hayashi a,b, Ge-Hong Sun-Wada d, Yoh Wada c, Mayumi Nakanishi-Matsui a,b, Masamitsu Futai a,b,⁎a Department of Biochemistry, Faculty of Pharmaceutical Sciences, Iwate Medical University, Iwate 028-3694, Japanb Futai Special Laboratory, Microbial Chemistry Research Center, Shinagawa, Tokyo 141-0021, Japanc Division of Biological Sciences, ISIR, Osaka University, Ibaraki, Osaka 567-0047, Japand Faculty of Pharmaceutical Sciences, Doshisha Women's College, Kyotanabe, Kyoto 610-0395, Japan
Abbreviations: V-ATPase, vacuolar-type proton pumlineethanesulfonic acid; E1, testis-specific V-ATPase subV-ATPase subunit E isoform⁎ Corresponding author. Department of Biochemist
Sciences, Iwate Medical University, 2-1-1 Nishitokuta,028-3694, Japan. Tel.: +81 19 651 5111; fax: +81 19 698
E-mail address: [email protected] (M. Futai).
0005-2728/$ – see front matter © 2008 Elsevier B.V. Adoi:10.1016/j.bbabio.2008.06.011
a b s t r a c t
a r t i c l e i n f oArticle history:
Mammalian vacuolar-type p Received 21 May 2008Received in revised form 26 June 2008Accepted 30 June 2008Available online 9 July 2008Keywords:V-ATPaseE1 isoformTestisSubunit assembly
roton pumping ATPases (V-ATPases) are diverse multi-subunit proton pumps.They are formed from membrane Vo and catalytic V1 sectors, whose subunits have cell-specific or ubiquitousisoforms. Biochemical study of a unique V-ATPase is difficult because ones with different isoforms are presentin the same cell. However, the properties of mouse isoforms can be studied using hybrid V-ATPases formedfrom the isoforms and other yeast subunits. As shown previously, mouse subunit E isoform E1 (testis-specific)or E2 (ubiquitous) can form active V-ATPases with other subunits of yeast, but E1/yeast hybrid V-ATPase isdefective in proton transport at 37 °C (Sun-Wada, G.-H., Imai-Senga, Y., Yamamoto, A., Murata, Y., Hirata, T.,Wada, Y., and Futai, M., 2002, J. Biol. Chem. 277, 18098–18105). In this study, we have analyzed the propertiesof E1/yeast hybrid V-ATPase to understand the role of the E subunit. The proton transport by the defectivehybrid ATPase was reversibly recovered when incubation temperature of vacuoles or cells was shifted to30 °C. Corresponding to the reversible defect of the hybrid V-ATPase, the Vo subunit a epitope was exposed tothe corresponding antibody at 37 °C, but became inaccessible at 30 °C. However, the V1 sector was stillassociated with Vo at 37 °C, as shown immunochemically. The control yeast V-ATPase was active at 37 °C, andits epitope was not accessible to the antibody. Glucose depletion, known to dissociate V1 from Vo in yeast, hadonly a slight effect on the hybrid at acidic pH. The domain between Lys26 and Val83 of E1, which containseight residues not conserved between E1 and E2, was responsible for the unique properties of the hybrid.These results suggest that subunit E, especially its amino-terminal domain, plays a pertinent role in theassembly of V-ATPase subunits in vacuolar membranes.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Vacuolar-type ATPase (V-ATPase) is similar to F-ATPase (ATPsynthase) in complex subunit organization, and the coupling betweenATPase catalysis and proton pumping through the rotation of subunits(for reviews, see Refs. [1–5]). V-ATPase, an essential mammalianmembrane enzyme [6,7], pumps protons into the lumens of diverseorganelles such as vacuoles, lysosomes, endosomes, and secretorygranules [1–5]. The lumenal acidic pH or electrochemical protongradient thus formed is required for many biological processesincluding endocytosis, exocytosis, fertilization, bone resorption, etc.
V-ATPase has thirteen different subunits (A, B, C, D, E, F, G, H, a, c,c', c'', and d), A-H and a-d for the V1 and Vo sectors, respectively, with
ping ATPase; MES, 4-morpho-unit E isoform; E2, ubiquitous
ry, Faculty of PharmaceuticalYahaba-cho, Shiwa-gun, Iwate1843.
ll rights reserved.
defined stoichiometries [1–5]. As expected from the diverse inside-acidic compartments in mammals, isoforms are found in unique cellsor organelles. The V-ATPases with subunit a isoform a1, a2, or a3 arefound ubiquitously in different endomembrane organelles, whereasthose with a4 are specific to the plasma membranes of kidneyintercalated cells [8–11]. Similarly, two and three isoforms have beenidentified for subunits E and C or G, respectively [12–15]. The mouseE1 and E2 isoforms encoded by the Atp6e1 and Atp6e2 genes,respectively, exhibit more than 70% amino acid identity to eachother [12]. E1 is specifically expressed in germ cells of testis, whereasE2 is found ubiquitously, i. e., in all tissues examined. They reside inthe stalk domain connecting V1 and Vo [1–5,7], and function as a rotoror stator during the subunit rotation [16]. It is of interest to determinethe roles of isoforms such as E1 in V-ATPase. However, the biochemicalproperties of a specific V-ATPase are difficult to establish because thesame cell contains ones with different isoforms.
The hybrid V-ATPases formed from a mammalian isoform andother yeast subunits will be useful to study the properties of theisoform. Mouse cDNA encoding the E, C, and G subunits, respectively,can complement Saccharomyces cerevisiae mutants lacking the

Fig. 1. Acidic vacuolar compartments of Δvma4 cells expressing the mouse E1 isoform or yeast Vma4p. Log-phase yeast Δvma4 cells carrying cDNA for mouse E1 [Δvma4 (E1)cells] or the yeast gene for Vma4p [Δvma4 (VMA4) cells] were incubated for 15 min at 30 °C or 37 °C in YPED medium buffered with Na2HPO4 (pH 7.5). The Δvma4 (E1) cellsincubated at 37 °C were also transferred to 30 °C medium for 15 min (37°→30°). Cells were stained with quinacrine, and fluorescent images (right panel) are shown togetherwith Nomarski ones (left panel).
1371K. Hayashi et al. / Biochimica et Biophysica Acta 1777 (2008) 1370–1377
corresponding genes [12–15], indicating that each mouse subunitcould form an active hybrid V-ATPase with other yeast subunits. Thus,a null mutant (Δvma4) harboring cDNA for the subunit E1 gene couldgrow at 30 °C. However, the same cells could not grow at 37 °C,although those with E2 cDNA could. The altered properties of these
Fig. 2. Formation of an electrochemical proton gradient in vacuolar vesicles expressing the m(E1) cells or Δvma4 (VMA4) cells were incubated at 30 °C or 37 °C for 5, 10, or 15 min in pH(37 °C→30 °C). The formation of an electrochemical proton gradient was assayed at theaddition of ATP (open arrowhead) or NH4Cl (closed arrowhead).
cells suggest the defective energy coupling between ATP hydrolysisand proton transport of E1/yeast hybrid V-ATPase [12]. Further studiesof the hybrid V-ATPase may shed light on the role of the E subunit.
In this study, we observed that the mouse E1/yeast hybridV-ATPase showed a reversible change in energy coupling depending
ouse E1/yeast hybrid V-ATPase. Vacuolar vesicles (10 μg protein) derived from Δvma47.5 buffer; or incubated at 37 °C for 15 min followed by a further 10 or 15 min at 30 °Cindicated temperatures. The incubation temperatures are indicated together with the

1372 K. Hayashi et al. / Biochimica et Biophysica Acta 1777 (2008) 1370–1377
on the temperature shift, and exhibited altered assembly properties.Its subunit a epitope was exposed to the anti-subunit a antibody at37 °C, although the V1 sector was not released from Vo. Upon shiftingof the medium to a lower temperature or to an acidic pH, the epitopebecame inaccessible to the antibody, due to the in vivo reassembly of afunctional V-ATPase. The dissociation of V1 from yeast V-ATPase,shown in glucose-depleted cells [17], was not observed with thehybrid V-ATPase at acidic pH. The altered assembly properties of thehybrid suggest that subunit E plays a pH-dependent role pertinent tothe assembly of the V1 and Vo sectors.
2. Experimental procedures
2.1. Construction of recombinant plasmids, cell growth, and staining
pKT10-N-myc-VMA4 and pKT10-N-myc-E1 carrying the entire openreading frames for Vma4p (yeast E subunit) and mouse E1, respectively,were constructed [12]. The DNA fragments of VMA4, Atp6e1 (E1 cDNA),and Atp6e2 (E2 cDNA) obtained on polymerase chain reaction wereligated into pKT10-N-myc to construct recombinant plasmids formouse/yeast chimeric E genes. The recombinant plasmids weretransformed into S. cerevisiae Δvma4 strain RH403 (MATα, Δvma4:TRP1, leu2-Δ1, ura3-52, trp1-Δ63, his3-Δ200, lys2-801, ade2-101) [12].
Yeast cells were grown to the middle of the logarithmic phase(∼107 cells/ml), and then subjected to further studies. YEPD (yeastextract, peptone, dextrose) medium or SD (synthetic dextrose)medium was buffered to pH 5.0 or 7.5 with 50 mM succinate and/or50 mM K2HPO4 [18]. Other procedures for yeast studies weredescribed previously [19].
2.2. Preparation of vacuolar membrane vesicles and measurement oftheir activities
Δvma4 cells expressing different subunit Es (Vma4p, E1, orE1/VMA4 chimera) were converted into spheroplasts with zymolyase100 T (Seikagaku Kogyo) in Medium I (YEP medium containing 0.1 MMES-Tris, pH 7.5, 2% glucose, 0.7 M sorbitol and 2 mM dithiothreitol).The spheroplasts were washed with Medium II (YEP mediumcontaining 0.8 M sorbitol and 2% glucose), and then incubated at30 °C for 20 min. They were then disrupted with a Douncehomogenizer, followed by preparation of vacuolar vesicles using Ficolldensity gradient centrifugation [20].
The formation of an electrochemical proton gradient in the vesicles(10 μg protein/1.5 ml assay solution) was examined at 30 °C or 37 °C in10 mM MES-Tris, pH 6.9, containing 25 mM KCl, 5 mM MgCl2, and5 μMquinacrine [21]. Proton transport was initiated by the addition of0.5 mM ATP. When necessary, 1.5 μM concanamycin A1 in 0.1%dimethyl sulfoxide was added to the assay mixture. For detection ofthe effect of glucose on V-ATPase assembly, spheroplasts obtained inMedium I containing 0.5% glucose were washed and incubated inMedium II (adjusted to pH 5.0) with or without 2% glucose at 30 °C for15 min, and then used for the preparation of vacuolar vesicles.
2.3. Immunochemical procedures
Cells (1×108) were incubated at 30 °C or 37 °C for 15 min, or at37 °C for 15 min followed by a further 15 min at 30 °C. They were fixedwith 3.7% formaldehyde at room temperature overnight [22]. Fixedcells were incubated with a monoclonal antibody, 10D7, againstsubunit a (Vph1p), and then stained with fluorescein isothiocyanate-conjugated anti-mouse IgG.
An immunoprecipitate was obtained from the solubilized celllysate or vacuolar vesicles as previously described with minormodifications [23–25]. Briefly, spheroplasts were incubated for20 min in YEPD medium (pH 5.0 or pH 7.5) at indicated temperatures(30 °C or 37 °C), and then lysed and cross-linked in phosphate-
buffered saline (pH 7.3) containing 1 mM EDTA, 1% C12E9, 1 mMdithiobis(succinimidylpropionate), 1 mM phenymethane-sulfonylfluoride and a protease inhibitor cocktail (added following themanufacturer's recommendation). After elimination of the non-spe-cific binding to protein A-Sepharose beads (Pharmacia Biosciences),the supernatant was added to 500 μl of the above solution containingthe monoclonal antibody and 200 μg/ml bovine serum albumin butwithout C12E9 and dithiobis. After 1 hour incubation, ProteinA-Sepharose was added to the solution, followed by incubation for1 h at the respective temperature and extensive washing. Proteinfractions eluted from the beads were subjected to gel electrophoresis,and V-ATPase subunits were detected by Western blotting. Vacuolarvesicles were also incubated with 1% C12E9 for 15 min forimmunochemical analysis. V-ATPase complexes were precipitatedwith the monoclonal antibody against subunit A, and subunits a, A, Band E were detected as described below. To prevent re-association ofVo and V1, samples were kept at indicated temperature during theimmunochemical procedure.
2.4. Other procedures and materials
Protein concentrations were estimated by means of BCA ProteinAssay from Pierce. Samples dissolved in cracking buffer [26] weresubjected to polyacrylamide gel electrophoresis (10% acrylamide) andWestern blotting with several antibodies, as described below. Cellsstained with fluorescent probes were observed with a Zeiss LSM510Control Imaging System. Monoclonal antibodies against V-ATPasesubunits (anti-subunit a, 10D7; anti-subunit A, 8B1; and anti-subunitB, 13D11) and myc epitope (9E10) were obtained from MolecularProbes and Sigma, respectively. The protease inhibitor cocktail wasfrom Roche. The FITC conjugated anti-mouse IgG was from VectorLaboratory. All other chemicals used were of the highest gradecommercially available.
3. Results
3.1. Temperature-sensitive defect of mouse E1/yeast hybrid V-ATPase
As one of the V-ATPase subunit mutants [2,5], S. cerevisiae cellslacking the VMA4 gene for subunit E (Δvma4 cells) could grow at pH 5.0,but not at pH 7.5 [12]. Obviously, thewild-type could grow at either pH.We have previously observed that Δvma4 cells harboring a plasmidencoding testis-specific isoform E1 [Δvma4 (E1) cells] exhibited growthin pH 7.5 medium at 30 °C, but not at 37 °C. On the other hand, Δvma4cells with E2 [Δvma4 (E2)] or authentic yeast E [Δvma4 (VMA4)] did notshow such a phenotype. Thus, Δvma4 (E1) cells were similar towild-type and Δvma4 cells at 30 °C and 37 °C, respectively. The growthphenotype of Δvma4 (E1) may be due to the temperature-sensitiveenergy coupling between ATPase and proton transport of the mouseE1/yeast hybrid V-ATPase [12], although other possibilities remain tobe studied. Studies on the alteredproperties of the hybrid V-ATPasemaycontribute to understanding of the role of the E subunit.
We first addressed to the kinetics and reversibility of the tempera-ture-sensitive pH change of vacuoles having hybrid V-ATPases, andexamined their acidification in vivo using quinacrine, a lipophilicfluorescent amine [27]. This fluorescent dye can cross membranes inits uncharged form, but is retained once protonated in acidic compart-ments. The dye was accumulated into the vacuoles of Δvma4 (VMA4)cells expressing Vma4pat 30 °C or 37 °C, and those ofΔvma4 (E1) cells at30 °C (Fig.1). The dyewasnot visible inΔvma4 (E1) vacuoles at 37 °C, butaccumulatedwithin 15min after shifting of themedium temperature to30 °C (Fig. 1, 37 °C→30 °C). These results suggested that the protonpumping of E1/yeast hybrid V-ATPase was apparently defective at 37°and became reversibly active at 30 °C in vivo.
Next, we studied the time course of the loss of proton pumping bythe hybrid V-ATPase. When vacuole vesicles from Δvma4 (E1) cells
1373K. Hayashi et al. / Biochimica et Biophysica Acta 1777 (2008) 1370–1377
were incubated at 37 °C, the electrochemical proton gradientdecreased significantly within 5 min, and became virtually undetect-able in 15 min (Fig. 2, left). However, the proton gradient wasrecovered to more than half of the original level upon shifting to 30 °C(Fig. 2, left, 37 °C→30 °C). These results indicate that the effect of non-permissive temperature is reversible, and assembly was recoveredrapidly after temperature shift to 30 °C. Loss of the proton gradient
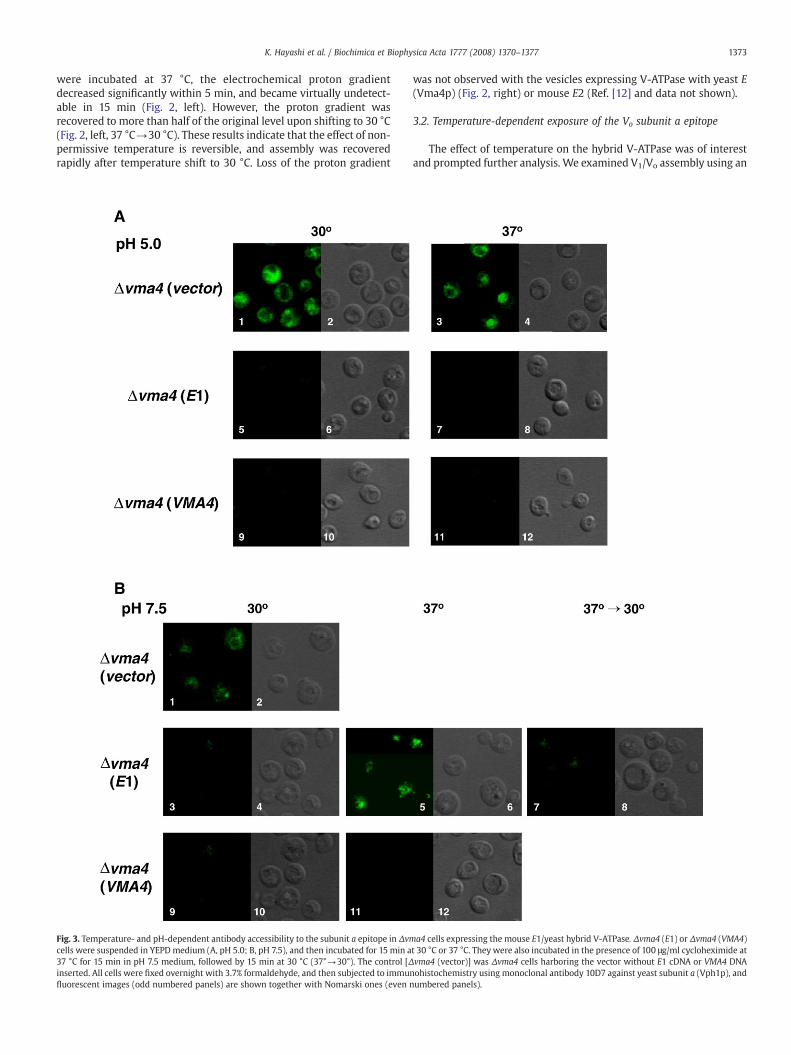
Fig. 3. Temperature- and pH-dependent antibody accessibility to the subunit a epitope in Δvmcells were suspended in YEPDmedium (A, pH 5.0; B, pH 7.5), and then incubated for 15 min a37 °C for 15 min in pH 7.5 medium, followed by 15 min at 30 °C (37°→30°). The control [Δinserted. All cells were fixed overnight with 3.7% formaldehyde, and then subjected to immunfluorescent images (odd numbered panels) are shown together with Nomarski ones (even n
was not observed with the vesicles expressing V-ATPase with yeast E(Vma4p) (Fig. 2, right) or mouse E2 (Ref. [12] and data not shown).
3.2. Temperature-dependent exposure of the Vo subunit a epitope
The effect of temperature on the hybrid V-ATPase was of interestand prompted further analysis. We examined V1/Vo assembly using an
a4 cells expressing the mouse E1/yeast hybrid V-ATPase. Δvma4 (E1) or Δvma4 (VMA4)t 30 °C or 37 °C. They were also incubated in the presence of 100 μg/ml cycloheximide atvma4 (vector)] was Δvma4 cells harboring the vector without E1 cDNA or VMA4 DNAohistochemistry using monoclonal antibody 10D7 against yeast subunit a (Vph1p), andumbered panels).

Fig. 4. Effects of temperature and pH on mouse E1/yeast hybrid V-ATPase assembly. (A)Presence of V-ATPase in a detergent solubilized cell lysate. Spheroplasts were incubatedfor 20min at indicated temperatures, and lysed in phosphate-buffered saline containing1% C12E9, 1 mM dithiobis(succinimidylpropionate), and a protease inhibitor mixture.The V-ATPase solubilized was immunoprecipitated with the antibody against subunit A.After gel electrophoresis of the precipitate, subunits a, A, and B were detected with thecorresponding antibodies. (B) Presence of V-ATPase in vacuolar vesicles. Vacuolarvesicles were prepared from Δvma4 (E1) cells and Δvma4 (VMA4) cells incubated for15 min at 30 °C or 37 °C, or fromΔvma4 (E1) cells incubated for 15 min at 37 °C and thenfurther for 15 min at 30 °C (37°→30°). V-ATPases solubilized from the vacuoles wereincubated with the antibody against subunit A. Immunoprecipitates were subjected togel electrophoresis, followed byWestern blotting with antibodies against subunits A, B,a, and E1 or Vma4p (anti-myc), respectively. The corresponding amounts of vacuolevesicles before immunoprecipitation are shown as controls (I).
1374 K. Hayashi et al. / Biochimica et Biophysica Acta 1777 (2008) 1370–1377
anti-subunit a antibody, whose epitope in Vph1p (yeast subunit a) isexposed when V1 is dissociated from the Vo sector [17]. Naturally, theantibody did not stain vacuoles of control normal yeast cells. However,it recognized the epitope in Δvma4 cells lacking subunit E [Δvma4(vector)] because the V1 sector was not assembled with Vo (Fig. 3A,panels 1–4, and Fig. 3B, panels 1 and 2).
The same antibody did not stain vacuoles of Δvma4 (VMA4) cellsgrown at either ≤30 °C or 37 °C in pH 5.0 or 7.5 medium (Fig. 3A and B)because the authentic yeast E subunit (Vma4p) was expressed, and V1
was assembled with Vo. Essentially the same results were obtainedwith Δvma4 (E1) cells grown at pH 5.0 (37 °C or 30 °C) (Fig. 3A) or pH7.5 (30 °C) (Fig. 3B), indicating that the V-ATPase was assemblednormally, and epitopes were not exposed under these conditions.
However, the antibody clearly stained vacuoles of Δvma4 (E1) cellsincubated at 37 °C in the pH 7.5 medium (Fig. 3B, panels 5 and 6), butcould not stain the same cells upon shifting of the temperature to30 °C in the presence or absence of 100 μg/ml cycloheximide (Fig. 3B,panels 7 and 8). Thus, the subunit a epitope, once exposed at 37 °C,became inaccessible at 30 °C without de novo protein synthesis. Theseresults suggest that VoV1 altered its assembly by exposing subunit aepitope at 37 °C in the pH 7.5 medium.
3.3. Association of Vo and V1 in mouse E1/yeast hybrid V-ATPase
To examine whether V1 sector completely dissociates from Vo inhybrid V-ATPase at 37 °C, the hybrid was studied immunochemicallyafter solubilization from a Δvma4 (E1) cell lysate in a buffer containinga detergent. The enzyme precipitatedwith the anti-subunit A antibodycontained both Vo and V1 subunits (a and B, respectively), regardless ofthe incubation temperature (30 °C or 37 °C) or the medium pH (pH 5.0or 7.5) (Fig. 4A). Similar results were obtained with the enzymesolubilized from vacuolar vesicles (Fig. 4B); the E1/yeast hybrid andcontrol yeast V-ATPases contained both V1 (A, B, E1 or Vma4p) and Vo
(a) subunits. These results suggested that the V1 and Vo sectors of thehybrid V-ATPase were associated, even when the subunit a epitopewas exposed to the antibody at 37 °C in the pH 7.5 medium (Fig. 3).
As a control, Δvma4 (VMA4) cells were cultured in pH 7.5 mediumwithout glucose, and a lysate of themwas incubated with a detergent.Subunit a was not found in the immunoprecipitate with the anti-subunit A antibody (Fig. 4A, lanes 7 and 8), indicating that V1 wasdissociated from Vo upon glucose depletion [7]. Thus, the assemblychange of the mouse E1/yeast hybrid V-ATPase is clearly differentfrom the dissociation of the two sectors.
3.4. Domain of E1 affecting the assembly of V-ATPase
To determine the domain of E1 isoforms affecting the assembly,genes encoding a series of chimeric Vma4p and E1 subunits wereconstructed, and transformed into Δvma4 cells. The Δvma4 cellsharboring the plasmid for VMA4, E1/VMA4-1, E1/VMA4-3, or E1/VMA4-6 could grow at 37 °C in the pH 7.5 medium. On the other hand,those harboring the plasmid for E1/VMA4-2, E1/VMA4-4, orE1/VMA4-5 could not grow, similar to the Δvma4 (E1) cells at 37 °Cin the pH 7.5 medium (Fig. 5A). Consistent with their growthphenotypes, vacuolar vesicles from these cells could not form anelectrochemical proton gradient at 37 °C (Fig. 5B). These cells had anE1 cDNA fragment between Lys26 and Val83 (E1 numbering) (Fig. 5C),indicating that this domain is responsible for the negative growth and
Fig. 5. Region of subunit E responsible for the temperature-sensitive assembly changes in Δplasmids for chimeric E subunits were introduced into Δvma4 cells: mouse E1, dark region;serially diluted 5-fold, and the diluted cultures were spotted on to a YPED (pH 5.0 or 7.5) plateand E2/VMA4-2) encoding mouse E2 (gray region) and VMA4 (light region) were also consfollow the E1 numbering. (B) Formation of an electrochemical proton gradient in vesicles fronomenclature for the chimeric E subunits being shown in A. Other conditions were as descsequences of mouse isoforms E1 and E2 were aligned together with that of yeast Vma4p usiboxed region (Lys26–Val83) is that responsible for the assembly property of the E1 isoform
defective proton transport due to the assembly change. The sameregion of the yeast E subunit (Vma4p) did not give such a phenotype(Fig. 5A and B). Furthermore, the Δvma4 cells with E2/VMA4-2encoding the chimeric subunit of Vma4p and E2 exhibited positivegrowth (Fig. 5A, bottom), and formed an electrochemical protongradient (data not shown). These results confirmed that the E2domain (in E2/VMA4-2 chimera) corresponding to between Lys26 andVal83 of E1 has no effect on the assembly of the V-ATPase.
3.5. Effects of glucose on the hybrid V-ATPase
As described above, the subunit a epitope of the hybrid V-ATPasewas exposed at 37 °C in the pH 7.5 but not in pH 5.0 medium. Thus, itbecame of interest to determine whether the glucose-dependent V1
dissociation from the hybrid is dependent on the medium pH. Whencontrol Δvma4 (VMA4) cells were incubated in the medium withoutglucose, V-ATPase became dissociated, as shown immunochemically(Fig. 6A, VMA4). Furthermore, the amounts of V1 subunits A, B, and E
vma4 cells. (A) Effects of chimeric E subunits on Δvma4 cell growth. The recombinantVMA4, light region. Individual Δvma4 cells (2×104 cells) grown to the log phase were. Growth was recorded after 48 h at 30 °C or 37 °C. Similarly, chimeric genes (E2/VMA4-1tructed, and introduced into Δvma4 cells. Residue numbers indicated (see closed box)m Δvma4 cells carrying chimeric E subunits. The assay temperatures are indicated, theribed in Fig. 2. (C) Alignment of the amino acid sequences of different E subunits. Theng the program ClustalW. Identical residues are highlighted (shaded residues), and the.

1375K. Hayashi et al. / Biochimica et Biophysica Acta 1777 (2008) 1370–1377
(Vma4p) in their vacuoles were significantly reduced, although that ofVo subunit awas maintained (Fig. 6B, VMA4). These results confirmedthat V1 became dissociated from Vo upon glucose depletion [17],regardless of the medium pH.
Essentially the same results were obtained with the hybridV-ATPase in Δvma4 (E1) cells grown in the pH 7.5 medium(Fig. 6A, E1). However, when the Δvma4 (E1) cells were incubatedat pH 5.0 with or without glucose, subunit a was detectable in the

Fig. 6. Glucose-independent assembly of the mouse E1/yeast hybrid ATPase. (A)Presence of V-ATPase in vacuoles derived from glucose-depleted cells. Spheroplasts ofΔvma4 (VMA4) or Δvma4 (E1) cells were incubated at 30 °C for 20 min in YEP medium(pH 5.0 or 7.5) with (+) or without (-) glucose, and then the V-ATPase was solubilized.The V-ATPase precipitated with an antibody against subunit A was subjected to gelelectrophoresis. Subunit a was detected by immunoblotting with the correspondingantibody. (B) Presence of V-ATPase in vacuoles derived from glucose-depleted cells.Spheroplasts of Δvma4 (VMA4) cells or Δvma4 (E1) cells were incubated at 30 °C for20min in YEP (pH 5.0) containing 0.8M sorbitol with (+) or without (−) glucose. Vacuolevesicles (V) were prepared, and then subjected to gel electrophoresis together with alysate (L). V-ATPase subunits were detected using antibodies against subunits a, A, B,and E (anti-myc). (C) Formation of an electrochemical proton gradient in vacuoles.Vacuole vesicles (10 μg protein/1.5 ml) were prepared from Δvma4 (VMA4) or Δvma4(E1) cells incubated with or without glucose (gray line, −glucose; dense line, +glucose).The formation of an electrochemical proton gradient was assayed at 30° as described inFig. 2. ATP and NH4Cl were added when indicated.
1376 K. Hayashi et al. / Biochimica et Biophysica Acta 1777 (2008) 1370–1377
immunoprecipitate (Fig. 6A, E1), and subunits A, B and E1 werefound in their vacuoles (Fig. 6B, E1). These vacuoles could form anelectrochemical proton gradient depending on ATP hydrolysis,whereas those from Δvma4 (VMA4) cells showed a significantlyreduced level (Fig. 6C). These results suggest that most of themouse E1/yeast hybrid V-ATPase was not dissociated into V1 andVo after glucose depletion in the pH 5.0 medium.
4. Discussion
Organelle or cell-specific isoforms have been identified for subunitsB, C, E, G, a, and d, consistent with diverse physiological roles ofmammalian V-ATPase [1–5,7]. The isoformsmay affect energy couplingbetween catalysis and proton transport, and also may determine the
localization to specific lumenal acidic compartments. Of these isoforms,subunit a may determine the localization of V-ATPase, since a2 wasfound in early endosomes [28], a3 in osteoclast plasmamembranes [29],and a3 in endocrine cells [30,31]. Subunit a2 was shown to recruitGDP/GTP exchange factor ARNO (ADP-ribosylation factor nucleotide siteopener) for transport vesicle formation [28]. The subunits in the stalkdomain may also affect V-ATPase localization. It should be emphasizedthat the properties of the isoforms could not be easily analyzed becausethe V-ATPasewith different isoforms is present in the samemammaliancells. However, their roles could be elucidated by analyzing hybridV-ATPases comprising mammalian and yeast subunits, examples beingthe low proton transport and high Km of yeast hybrid V-ATPasesassembled with mouse C2-a or C2-b isoforms [13].
The mouse E1/yeast hybrid V-ATPase showed apparent uncouplingbetween ATPase and proton transport at 37 °C, although it was activeat lower temperature [12]. We observed in this study that thetemperature-dependent proton transport activities and subunitinteractions were reversible, and V1 was associated with Vo even at37 °C. The subunit a (Vph1p) epitope became exposed to thecorresponding antibody when Δvma4 (E1) cells were incubated at37 °C in pH 7.5 medium, but not in pH 5.0 medium. Upon atemperature shift to b30 °C, the epitope became inaccessible almostimmediately, and the V-ATPase regained the ability to form anelectrochemical proton gradient. Immunoprecipitation study revealedthat V1 and Vo were associated even at non-permissive temperature,consistent with rapid recovery of proton pumping upon shift to lowertemperature. These results suggest that the hybrid enzyme showedaltered subunit assembly in response to the higher temperature, butstill remained assembled at 37 °C, in contrast to the dissociation of V1
and Vo caused by glucose depletion [17]. As described above, exposureof subunit a epitope was clearly correlated with defective protontransport activity at 37 °C, and quick recovery at 30 °C. Effects of pHwere also consistent with these results, suggesting that the defect inproton pumping of hybrid V-ATPase is due to altered assembly.
Analysis of the E1/Vma4p chimeric subunit indicated that the E1domain between Lys26 and Val83 is responsible for the alteredassembly of the hybrid V-ATPase. Eight residues are different betweenthe E1 and E2 domains, suggesting that they are pertinent as to theinteractions with other subunit(s) in the stalk and membranedomains. The importance of the amino terminus of Vma4p was alsosuggested for binding of the G subunit [35,36], and for interactionwith subunits C [37] and H [38].
The role of subunit E in assembly is supported by the interactionswith multiple subunits; subunit E is an elongated protein extendingfrom the top of V1 including the subunit B to Vo domains [33,34], andin the close proximity to stalk subunits C, H and G [35]. Subunit E mayplay a regulatory role, by interacting with the glycolytic enzymealdolase [39,40]. The RAVE (regulator of the H+-ATPase of vacuolar andendosomal membranes) complex was shown to associate withV1 released from V-ATPase [41]. This interaction was inhibited for V1
lacking E or G. The present study clearly supported the role ofsubunit E in V-ATPase assembly.
As described above, the accessibility of the subunit a epitopeand glucose-sensitive dissociation of the hybrid V-ATPase weredependent on the medium pH. In this regard, the effect of themedium pH on the vacuolar lumen was observed previously [32].The vacuolar lumen of Δvma4 (E1) was less acidic than that ofΔvma4 (VMA4), as shown above by staining with quinacrine.Consistently, the E1/yeast hybrid V-ATPase formed a slightly lowerelectrochemical gradient than the yeast V-ATPase. Effects of pH onV-ATPase assembly were also shown: functional association of V1
and Vo in vitro is strongly dependent on pH (optimum, 5.5) [24],and their glucose-dependent dissociation in vivo is inhibited by theincrease in lumenal pH caused by chloroquine [23]. Thus, thepresent results suggest that the medium affected the vacuolar pH,which regulated the assembly of the E1/yeast hybrid ATPase.

1377K. Hayashi et al. / Biochimica et Biophysica Acta 1777 (2008) 1370–1377
Acknowledgments
We are grateful for the support of CREST, the Japan Science andTechnology Agency, the Ministry of Education, Science and Culture ofJapan, and the Japan Foundation for Applied Enzymology. We alsothank Daiichi Pharmace:utical Co., Ltd., Shionogi & Co., Ltd., and EisaiCo., Ltd for their support.
References
[1] M. Futai, T. Oka, G.-H. Sun-Wada, Y. Moriyama, H. Kanazawa, Y. Wada, Luminalacidification of diverse organelles by V-ATPase in animal cells, J. Exp. Biol. 203(2000) 107–116.
[2] W. Harvey, N. Nelson, Vacuolar and plasma membrane proton-adenosinetripho-sphatases, Physiol. Rev. 79 (1999) 361–385.
[3] T. Nishi, M. Forgac, The vacuolar H+-ATPases—nature's most versatile protonpumps, Nat. Rev. Mol. Biol. 3 (2002) 94–103.
[4] G.-H. Sun-Wada, Y. Wada, M. Futai, Diverse and essential roles of mammalianvacuolar-type proton pump ATPase: toward the physiological understanding ofinside acidic compartments, Biochim. Biophys. Acta 1658 (2004) 106–114.
[5] P.M. Kane, The where, when, and how of organelle acidification by the yeastvacuolar H+-ATPase, Microbiol. Mol. Biol. Rev. 70 (2006) 177–191.
[6] G.-H. Sun-Wada, Y. Murata, A. Yamamoto, H. Kanazawa, Y. Wada, M. Futai, Acidicendomembrane organelles are required for mouse post-implantation develop-ment, Dev. Biol. 228 (2000) 315–325.
[7] G.-H. Sun-Wada, Y.Wada, M. Futai, Vacuolar H+ pumping ATPases in luminal acidicorganelles and extracellular compartments: common rotational mechanism anddiverse physiological roles, J. Bioenerg. Biomembr. 35 (2003) 347–358.
[8] T. Toyomura, T. Oka, C. Yamaguchi, Y. Wada, M. Futai, Three subunit a isoforms ofmouse vacuolar H+-ATPase. Preferential expression of the a3 isoform duringosteoclast differentiation, J. Biol. Chem. 275 (2000) 8760–8765.
[9] T. Oka, Y. Murata, M. Namba, T. Yoshimizu, T. Toyomura, A. Yamamoto, G.-H. Sun-Wada, N. Hamasaki, Y. Wada, M. Futai, a4, a unique kidney-specific isoformof mouse vacuolar H+-ATPase subunit a, J. Biol. Chem. 276 (2001) 40050–40054.
[10] T. Nishi, M. Forgac, Molecular cloning and expression of three isoforms of the100-kDa a subunit of the mouse vacuolar proton-translocating ATPase, J. Biol.Chem. 275 (2000) 6824–6830.
[11] A.N. Smith, K.E. Finberg, C.A. Wagner, R.P. Lifton, M.A. Devonald, Y. Su, F.E. Karet,Molecular cloning and characterization of Atp6n1b: a novel fourth murinevacuolar H+-ATPase a-subunit gene, J. Biol. Chem. 276 (2001) 42382–42388.
[12] G.-H. Sun-Wada, Y. Imai-Senga, A. Yamamoto, Y. Murata, T. Hirata, Y. Wada, M.Futai, A proton pump ATPase with testis-specific E1-subunit isoform required foracrosome acidification, J. Biol. Chem. 277 (2002) 18098–18105.
[13] G.-H. Sun-Wada, Y. Murata, M. Namba, A. Yamamoto, Y. Wada, M. Futai, Mouseproton pump ATPase C subunit isoforms (C2-a and C2-b) specifically expressed inkidney and lung, J. Biol. Chem. 278 (2003) 44843–44851.
[14] G.-H. Sun-Wada, T. Yoshimizu, Y. Imai-Senga, Y. Wada, M. Futai, Diversity of mouseproton-translocating ATPase: presence of multiple isoforms of the C, d and Gsubunits, Gene 302 (2003) 147–153.
[15] Y. Murata, G.-H. Sun-Wada, T. Yoshimizu, A. Yamamoto, Y. Wada, M. Futai,Differential localization of the vacuolar H+ pump with G subunit isoforms (G1 andG2) in mouse neurons, J. Biol. Chem. 277 (2002) 36296–36303.
[16] T. Hirata, A. Iwamoto-Kihara, G.-H. Sun-Wada, T. Okajima, Y. Wada, M. Futai,Subunit rotation of vacuolar-type proton pumping ATPase: relative rotation of theG and C subunits, J. Biol. Chem. 278 (2003) 23714–23719.
[17] P.M. Kane, Disassembly and reassembly of the yeast vacuolar H+-ATPase in vivo,J. Biol. Chem. 270 (1995) 17025–17032.
[18] R. Hirata, L.A. Graham, A. Takatsuki, T.H. Stevens, Y. Anraku, VMA11 and VMA16encode second and third proteolipid subunits of the Saccharomyces cerevisiaevacuolar membrane H+-ATPase, J. Biol. Chem. 272 (1997) 4795–4803.
[19] D. Burke, D. Dawson, T. Stearns, Methods in Yeast Genetics, Cold Spring HarborLaboratory Press, 2000.
[20] K. Takeshige, M. Baba, S. Tsuboi, T. Noda, Y. Ohsumi, Autophagy in yeastdemonstrated with proteinase-deficient mutants and conditions for its induction,J. Cell. Biol. 119 (1992) 301–311.
[21] T. Hirata, N. Nakamura, H. Omote, Y. Wada, M. Futai, Regulation and reversibility ofvacuolar H+-ATPase, J. Biol. Chem. 275 (2000) 386–389.
[22] C.J. Roberts, C.K. Raymond, C.T. Yamashiro, T.H. Stevens, Methods for studying theyeast vacuole, Methods Enzymol. 194 (1991) 644–661.
[23] E. Shao, M. Forgac, Involvement of the nonhomologous region of subunit A of theyeast V-ATPase in coupling and in vivo dissociation, J. Biol. Chem. 279 (2004)48663–48670.
[24] K.J. Parra, P.M. Kane, Wild-type and mutant vacuolar membranes supportpH-dependent reassembly of the yeast vacuolar H+-ATPase in vitro, J. Biol.Chem. 271 (1996) 19592–19598.
[25] P.M. Kane, C.T. Yamashiro, T.H. Stevens, Biochemical characterization of the yeastvacuolar H+-ATPase, J. Biol. Chem. 264 (1989) 19236–19244.
[26] P.M. Kane, M.C. Kuehn, I. Howald-Stevenson, T.H. Stevens, Assembly and targetingof peripheral and integral membrane subunits of the yeast vacuolar H+-ATPase,J. Biol. Chem. 267 (1992) 447–454.
[27] L.S. Weisman, R. Bacallao, W. Wickner, Multiple methods of visualizing the yeastvacuole permit evaluation of its morphology and inheritance during the cell cycle,J. Cell. Biol. 105 (1987) 1539–1547.
[28] A. Hurtado-Lorenzo, M. Skinner, J.E. Annan, M. Futai, G.-H. Sun-Wada, S. Bourgoin,J. Casanova, A. Wildeman, S. Bechoua, D.A. Ausiello, D. Brown, V. Marshansky,V-ATPase interacts with ARNO and Arf6 in early endosomes and regulates theprotein degradative pathway, Nat. Cell Biol. 8 (2006) 124–136.
[29] T. Toyomura, Y. Murata, A. Yamamoto, T. Oka, G.-H. Sun-Wada, Y. Wada, M.Futai, From lysosomes to the plasma membrane: localization of vacuolar-typeH+-ATPase with the a3 isoform during osteoclast differentiation, J. Biol. Chem.278 (2003) 22023–22030.
[30] G.-H. Sun-Wada, T. Toyomura, Y. Murata, A. Yamamoto, M. Futai, Y. Wada, The a3isoform of V-ATPase regulates insulin secretion from pancreatic beta-cells, J. Cell.Sci. 119 (2006) 4531–4540.
[31] G.-H. Sun-Wada, H. Tabata, N. Kawamura, M. Futai, Y. Wada, Differentialexpression of a subunit isoforms of the vacuolar-type proton pump ATPase inmouse endocrine tissues, Cell Tissue Res. 329 (2007) 239–248.
[32] P.J. Plant, M.F. Manolson, S. Grinstein, N. Demaurex, Alternative mechanisms ofvacuolar acidification in H+-ATPase-deficient yeast, J. Biol. Chem. 274 (1999)37270–37279.
[33] Y. Arata, J.D. Baleja, M. Forgac, Cysteine-directed cross-linking to subunit B suggeststhat subunit E forms part of the peripheral stalk of the vacuolar H+-ATPase, J. Biol.Chem. 277 (2002) 3357–3363.
[34] Y. Arata, J.D. Baleja, M. Forgac, Localization of subunits D, E, and G in the yeastV-ATPase complex using cysteine-mediated cross-linking to subunit B. Biochem-istry (Washington) 41 (2002) 11301–11307.
[35] T. Xu, E. Vasilyeva, M. Forgac, Subunit interactions in the clathrin-coated vesiclevacuolar H+-ATPase complex, J. Biol. Chem. 274 (1999) 28909–28915.
[36] M. Ohira, A.M. Smardon, C.M. Charsky, J. Liu, M. Tarsio, P.M. Kane, The E and Gsubunits of the yeast V-ATPase interact tightly and are both present at more thanone copy per V1 complex, J. Biol. Chem. 281 (2006) 22752–22760.
[37] R.P.O. Jones, L.J. Durose, J.B.C. Findlay, M.A. Harrison, Defined sites of interactionbetween subunits E (Vma4p), C (Vma5p), and G (Vma10p) within the statorstructure of the vacuolar H+-ATPase, Biochemistry (Washington) 44 (2005)3933–3941.
[38] M. Lu, S. Vergara, L. Zhang, L.S. Holliday, J. Aris, S.L. Gluck, The amino-terminaldomain of the E subunit of vacuolar H+-ATPase (V-ATPase) interacts with theH subunit and is required for V-ATPase function, J. Biol. Chem. 277 (2002)38409–38415.
[39] M. Lu, L.S. Holliday, L. Zhang, W.A. Dunn Jr., S.L. Gluck, Interaction betweenaldolase and vacuolar H+-ATPase: evidence for direct coupling of glycolysis to theATP-hydrolyzing proton pump, J. Biol. Chem. 276 (2001) 30407–30413.
[40] M. Lu, Y.Y. Sautin, L.S. Holliday, S.L. Gluck, The glycolytic enzyme aldolase mediatesassembly, expression, and activity of vacuolar H+-ATPase, J. Biol. Chem. 279 (2004)8732–8739.
[41] A.M. Smardon, M. Tarsio, P.M. Kane, The RAVE complex is essential for stableassembly of the yeast V-ATPase, J. Biol. Chem. 277 (2002) 13831–13839.

![AtHMA3, a P1B-ATPase Allowing Cd/Zn/Co/Pb … › content › plantphysiol › 149 › 2 › 894.full.pdfAtHMA3, a P 1B-ATPase Allowing Cd/Zn/Co/Pb Vacuolar Storage in Arabidopsis1[W]](https://img.pdfslide.us/doc/110x75/5f0eb3fb7e708231d44085a9/athma3-a-p1b-atpase-allowing-cdzncopb-a-content-a-plantphysiol-a-149-a.jpg)



























