Embed Size (px)
Citation preview

11. RADIOLARIANS FROM THE MIDDLE AMERICA TRENCH OFF GUATEMALA,DEEP SEA DRILLING PROJECT LEG 671
M. J. Westberg and W. R. Riedel, Geological Research Division, Scripps Institution of Oceanography,University of California, San Diego, La Jolla, California
ABSTRACT
Radiolarians were found at all Leg 67 sites. At oceanic reference Site 495 and Trench floor Sites 499 and 500, radio-larian sequences from the lower Miocene Stichocorys delmontensis Zone through the Quaternary were recovered. Foursites were located on the inner Trench slope: at Sites 496 and 498, drilling penetrated to the Calocycletta costata Zone ofthe early Miocene, at Site 497 only Quaternary and Pliocene material was recovered, and at Site 494, the Cretaceous wasreached. An index of reliability is described and used to resolve conflicts in radiolarian correlations of the seven sites.The composition of assemblages is described in terms of relative abundances of radiolarian families, and these data areused as a basis for comparing margin and open-ocean sites.
INTRODUCTION
Radiolarians were recovered at all seven Leg 67 siteslocated on a transect of the Middle America Trench offGuatemala. Oceanic reference Site 495 is on the CocosPlate, 22 km seaward of the Trench axis; Sites 499 and500 are on the Trench floor; and Sites 494, 496, 497, and498 are on the inner Trench wall. The localities andwater depths of the sites are:
Site 494, 12°43.01'N, 90°55.97'W, 5529 metersSite 495, 12°29.78'N, 91°02.26'W, 4150 metersSite 496, 13°O3.82'N, 90°47.71'W, 2064 metersSite 497, 12°59.23'N, 90°49.68'W, 2358 metersSite 498, 12°42.68'N, 90°54.94'W, 5497 metersSite 499, 12°40.23'N, 90°56.65'W, 6127 metersSite 500, 12°41.16'N, 90°56.58'W, 6127 metersRadiolarian assemblages range from lower Miocene
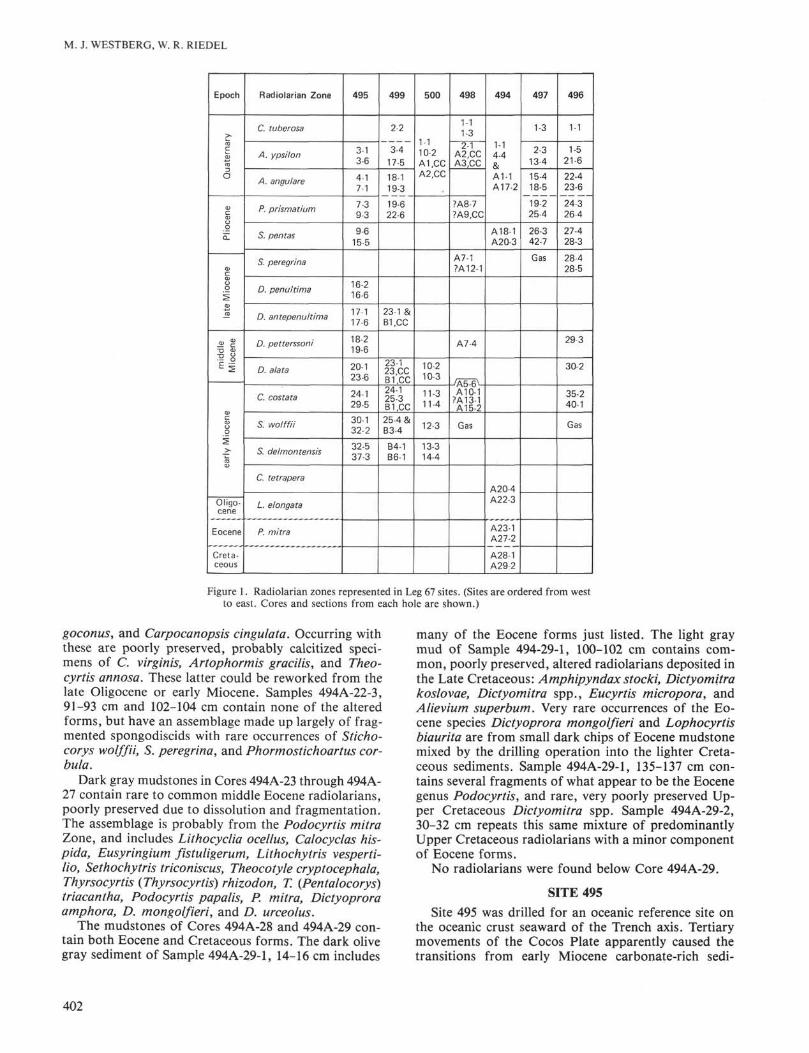
to Quaternary at Sites 495, 496, 498, 499, and 500 andfrom Pliocene to Quaternary at Site 497. Site 494 con-tains a Pliocene to Quaternary sequence from Cores 1 to20, rare lower Miocene forms in Cores 20 to 22, a mid-dle Eocene assemblage in Cores 23 to 27, and rare Creta-ceous and Eocene forms in Cores 28 and 29. Figure 1 isa synopsis of radiolarian zones represented in Leg 67sites.
In many of the Quaternary and Pliocene samples,and in Miocene slope sediments, radiolarian assem-blages are diluted by clay aggregates, ash, diatoms, orterrigenous minerals.
PROCEDURESGenerally, we took two or three samples per core, but rapidly ac-
cumulated sediments were sampled at longer intervals. Sediments weresieved at 44 µm and strewn slides prepared in our standard manner.
Relative abundances of species are recorded in two ways. First, inthe main part of ranges and at most morphotypic limits, five grades ofabundance indicate the percentage that a taxon represents of the totalassemblage on the strewn slide. These abundance grades are explained
Aubouin, J., von Huene, R., et al., Init. Repts. DSDP, 67: Washington (U.S. Govt.Printing Office).
in the note to Table 1. Second, near evolutionary transitions, "con-stant-numerator" percentage estimates were used. The density of ra-diolarians on a slide having been estimated, this density is then used toestimate the number of radiolarians in a counted number of fields thatmust be searched to find 10 specimens of a taxon (Riedel and San-filippo, 1978a).
Relative abundance data for radiolarian families are recorded in athird way. At least 300 radiolarians per slide were categorized in 15families, and percentages calculated and rounded to the nearest wholenumber. The family abundance tables also include an estimate of theproportion of each field covered by diluting components such as dia-toms, volcanic ash, or terrigenous debris.
The radiolarian zones used in this paper for the Tertiary are thechronozones defined by Riedel and Sanfilippo (1978b). The Quater-nary zones are those defined by Nigrini (1971).
SITE 494
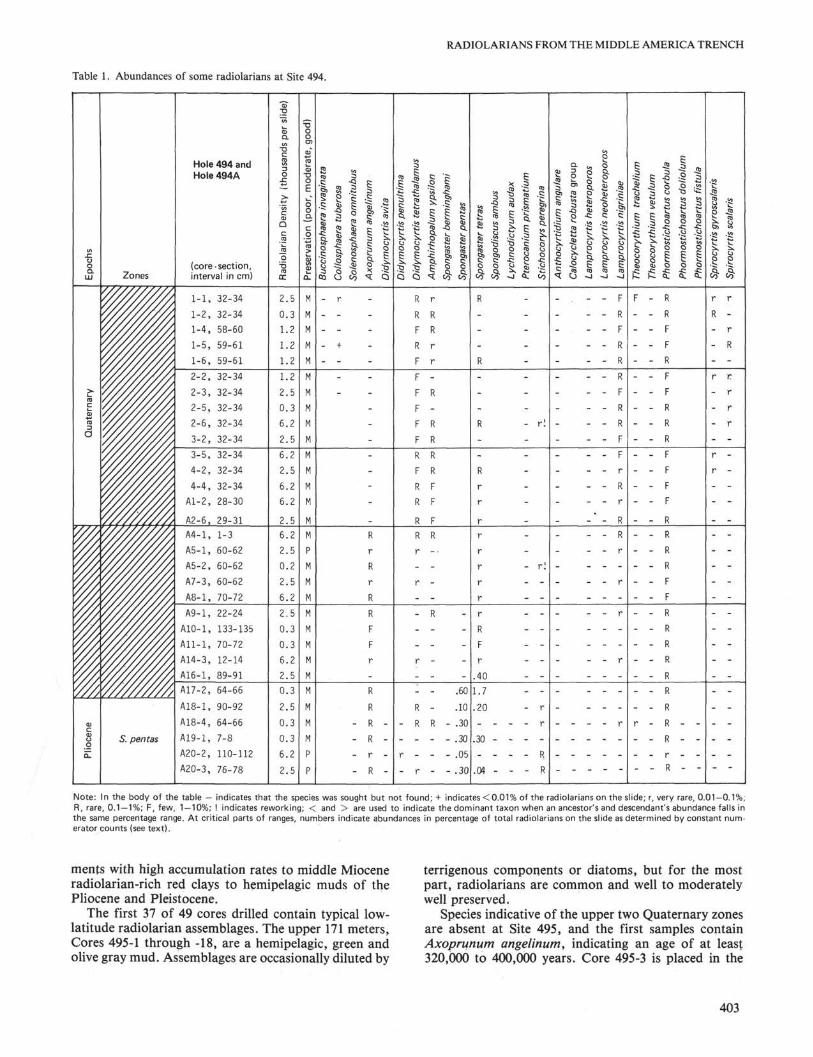
Site 494 was drilled on a small terrace of the lowerTrench slope. The Holocene to Pliocene, dark gray, dia-tomaceous mud of Cores 494-1 through 494A-20 con-tains radiolarian assemblages of moderate preservation,but often highly diluted by clay aggregates, diatoms, orash. Because of the sparseness of the fauna and missingindex species, the Quaternary zones cannot be deter-mined. The latest occurrence of Axoprunum angelinumis between Samples 494A-2-6, 29-31 cm and 494A-4-1,1-3 cm. According to Hays et al. {Stylatractus sp., 1969)and Johnson and Knoll (1975), this species became ex-tinct about 320,000 to 400,000 years ago. The top of theSpongaster pentas Zone, indicated by the morphotypictop of Stichocorys peregrina, is between Samples 494A-17-2, 64-66 cm and 494A-18-1, 90-92 cm. Blue clastswith middle Miocene species occur throughout the sec-tion. Occasionally, Eocene species are reworked intothese younger sediments. Relative abundances of Plio-cene and younger taxa found in these cores are recordedin Table 1.
The blue gray hemipelagic clays of Sections 494A-20-4 to 494A-22-3 contain radiolarians in two differentpreservational states. There are moderately well-pre-served specimens of Calocycletta virginis, Eucyrtidiumdiaphanes, Stichocorys delmontensis, Theocorys spon-
401
M. J. WESTBERG, W. R. RIEDEL
Epoch
Qua
tern
ary
Plio
cene
late
Mio
cene
mid
dle
Mio
cene
earl
y M
ioce
ne
Oligo-cene
Eocene
Creta-ceous
Radiolarian Zone
C. tuberosa
A. ypsilon
A. angulare
P. prismatium
S. pentas
S. peregrina
D. penultima
D. antepenultima
D. petterssoni
D. alata
C. costata
S. wolffii
S. delmontensis
C. tetrapera
L. elongate
P. mitra
495
3-13-6
4-17-1
7-39-3
9-615-5
16-216-6
17-117-6
18-219-6
20-123-6
24-129-5
30-132-2
32-537-3
499
2-2
3-4
17-5
18-119-3
19-622-6
23-1 &B1,CC
23-123,CCB1 CC24-125-3B1.CC25-4 &B3-4
B4-1B6-1
5 0 0
1-110-2A1,CCA2,CC
10-210-3
11-311-4
12-3
13-314-4
498
1-11-32-1
A2,CCA3,CC
7A8-7?A9,CC
A7-17A12-1
A7-4
A10-17A13-1A15-2
Gas
494
1-14-4&A MA17-2
A18-1A20-3
A20-4A22-3
A23-1A27-2
A28-1A29-2
497
1-3
2-313-4
15-418-5
19-225-4
26-342-7
Gas
496
1-1
1-521-6
22-423-6
24-326-4
27-428-3
28-428-5
29-3
30-2
35-240-1
Gas
Figure 1. Radiolarian zones represented in Leg 67 sites. (Sites are ordered from westto east. Cores and sections from each hole are shown.)
goconus, and Cαrpocαnopsis cingulαtα. Occurring withthese are poorly preserved, probably calcitized speci-mens of C. virginis, Artophormis grαcilis, and Theo-cyrtis αnnosα. These latter could be reworked from thelate Oligocene or early Miocene. Samples 494A-22-3,91-93 cm and 102-104 cm contain none of the alteredforms, but have an assemblage made up largely of frag-mented spongodiscids with rare occurrences of Sticho-corys wolffii, S. peregrina, and Phormostichoartus cor-bula.
Dark gray mudstones in Cores 494A-23 through 494A-27 contain rare to common middle Eocene radiolarians,poorly preserved due to dissolution and fragmentation.The assemblage is probably from the Podocyrtis mitraZone, and includes Lithocyclia ocellus, Calocyclas his-pida, Eusyringium fistuligerum, Lithochytris vesperti-lio, Sethochytris triconiscus, Theocotyle cryptocephala,Thyrsocyrtis {Thyrsocyrtis) rhizodon, T. (Pentalocorys)triacantha, Podocyrtis papa I is, P. mitra, Dictyoproraamphora, D. mongolfieri, and D. urceolus.
The mudstones of Cores 494A-28 and 494A-29 con-tain both Eocene and Cretaceous forms. The dark olivegray sediment of Sample 494A-29-1, 14-16 cm includes
many of the Eocene forms just listed. The light graymud of Sample 494-29-1, 100-102 cm contains com-mon, poorly preserved, altered radiolarians deposited inthe Late Cretaceous: Amphipyndax stocki, Dictyomitrakoslovae, Dictyomitra spp., Eucyrtis micropora, andAlievium superbum. Very rare occurrences of the Eo-cene species Dictyoprora mongolfieri and Lophocyrtisbiaurita are from small dark chips of Eocene mudstonemixed by the drilling operation into the lighter Creta-ceous sediments. Sample 494A-29-1, 135-137 cm con-tains several fragments of what appear to be the Eocenegenus Podocyrtis, and rare, very poorly preserved Up-per Cretaceous Dictyomitra spp. Sample 494A-29-2,30-32 cm repeats this same mixture of predominantlyUpper Cretaceous radiolarians with a minor componentof Eocene forms.
No radiolarians were found below Core 494A-29.
SITE 495
Site 495 was drilled for an oceanic reference site onthe oceanic crust seaward of the Trench axis. Tertiarymovements of the Cocos Plate apparently caused thetransitions from early Miocene carbonate-rich sedi-
402
RADIOLARIANS FROM THE MIDDLE AMERICA TRENCH
Table 1. Abundances of some radiolarians at Site 494.E
po
chs
>
<o
σ
V/λ
Y//λ
VÁv/λi
<u<uuo
Zones
yyyyyyyyyy/vyyyyyyy///
yyyyyyyyy/'yyyyyyyyyhyyyyy//y///
/yyyyyyy////yyyyyyyyys>'yyyyyyyy//'
(Vf/f/fY///
v//0/Zv///
S. pentas
Hole 494 andHole 494A
(core-section,interval in cm)
1-1,
1-2,
1-4,
1-5,
1-6,
2-2,
2-3,
2-5,
2-6,
3-2,
3-5,
4-2,
4-4,
A l - 2 ,
A2-6,
A4-1,
A5-1,
A5-2,
A7-3,
A8-1,
A9-1,
A10-1,
A l l - 1 ,
A14-3,
A16-1,
A17-2,
A18-1,
A18-4,
A19-1,
A20-2
A20-3,
32-34
32-34
58-60
59-61
59-61
32-34
32-34
32-34
32-34
32-34
32-34
32-34
32-34
28-30
29-31
1-3
60-62
60-62
60-62
70-72
22-24
133-135
70-72
12-14
89-91
64-66
90-92
64-66
7-8
110-112
76-78
ide)
Ra
dio
lari
an
De
nsi
ty
(th
ou
san
ds
per
s
2.5
0.3
1.2
1.2
1.2
1.2
2.5
0.3
6.2
2.5
6.2
2.5
6.2
6.2
2.5
6.2
2.5
0.2
2.5
6.2
2,5
0.3
0.3
6.2
2.5
0.3
2.5
0.3
0.3
6 2
2.5
Pre
serv
ati
on
(p
oo
r, m
od
era
te,
go
od
)M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
MP
MM
M
M
M
MM
M
M
M
M
Mp
P
Bu
ccin
osp
ha
era
in
vag
ina
ta
-
-
-
-
-
Co
llosp
ha
era
tu
be
rosa
r
-
-
+
-
-
-
So
len
osp
ha
era
om
nit
ub
us
-
-
-
Axo
pru
nu
m a
ng
elin
um
-
-
-
-
-
-
-
-
-
-
-
-
-
_
R
r
R
r
R
R
F
F
r
-
R
R
R
R
R
Did
ymo
cyr
tis
avi
ta
-
-
-
Did
ymo
cyr
tis
pe
nu
ltim
a
-
-
Did
ymo
cyr
tis
tetr
ath
ala
mu
s
R
R
F
R
F
F
F
F
F
F
R
F
R
R
R
R
r
-
r
-
-
-
-
r
-
-
R
R
-
r
Am
ph
irh
op
alu
m
ypsi
lon
r
R
R
r
r
-
R
-
R
R
R
R
F
F
F
R
- .
-
-
-
R
-
-
-
-
-
-
R
-
-
Sp
on
gas
ter
be
rmin
gh
am
i
-
-
-
Sp
on
gas
ter
pe
nta
s
-
-
-
-
-
.60
.10
.30
.30
OS
.30
Sp
on
gas
ter
tetr
as
R
-
-
-
R
-
-
-
R
-
-
R
rr
r
r
r
r
r
r
r
R
F
r
.40
1.7
.20
-
.30
.04
Sp
on
go
dis
cus
am
bu
s
-
Lyc
hn
od
icty
um
au
da
x
-
Pte
roc
an
ium
p
rism
ati
um
-
-
-
-
-
-
-
-
-
-
-
-
-
-
_
-
-
-
-
-
-
-
-
-
-
-
-
-
Sti
ch
oc
ory
s p
ere
gri
na
r!
rl
-
-
-
-
-
-
-
-
r
r
D
R
An
tho
cy
rtid
ium
an
gu
iare
-
-
-
-
-
-
-
-
-
-
-
-
-
-
_
-
-
-
-
-
-
-
-
-
-
-
-
-
Ca
loc
ycle
tta
ro
bu
sta
gro
up
-
Lam
pro
cyr
tis
he
tero
po
ros
-
-
-
-
-
-
-
-
-
-
-
-
-
-
_
-
-
-
-
-
-
-
-
-
-
-
-
La
mp
roc
yrti
s n
eo
he
tero
po
ros
-
-
-
-
-
-
-
-
-
-
-
-
-
-
_
-
-
-
-
-
-
-
-
-
-
-
-
Lam
pro
cyr
tis
nig
rin
iae
F
R
F
R
R
R
F
R
R
F
F
r
R
r
R
R
r
r
-
r
-
-
r
-
-
-
r
Th
eo
co
ryth
ium
tr
ac
he
lium
F
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
r
Th
eo
co
ryth
ium
ve
tulu
m
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
Ph
orm
ost
ich
oa
rtu
s c
orb
ula
R
R
F
F
R
F
F
R
R
R
F
F
F
F
R
R
R
R
F
F
R
R
R
R
R
R
R
R
R
R
Ph
orm
ost
ich
oa
rtu
s d
olio
lum
-
-
Ph
orm
ost
ich
oa
rtu
s fi
stu
la
-
-
Sp
iro
cyr
tis
gyr
osc
ala
ris
r
R
-
-
-
r
-
-
-
-
r
r
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
Sp
iro
cyr
tis
scai
aris
r
-
r
R
-
r
r
r
-
-
-
-
-
_
-
-
-
-
-
-
-
-
-
-
-
-
-
-
Note: In the body of the table — indicates that the species was sought but not found; + indicates < 0.01% of the radiolarians on the slide; r, very rare, 0.01-0.1%,R, rare, 0.1 — 1%; F, few, 1 — 10%; ! indicates reworking; < and > are used to indicate the dominant ta×on when an ancestors and descendants abundance falls inthe same percentage range. At critical parts of ranges, numbers indicate abundances in percentage of total radiolarians on the slide as determined by constant num-erator counts (see text).
ments with high accumulation rates to middle Mioceneradiolarian-rich red clays to hemipelagic muds of thePliocene and Pleistocene.
The first 37 of 49 cores drilled contain typical low-latitude radiolarian assemblages. The upper 171 meters,Cores 495-1 through -18, are a hemipelagic, green andolive gray mud. Assemblages are occasionally diluted by
terrigenous components or diatoms, but for the mostpart, radiolarians are common and well to moderatelywell preserved.
Species indicative of the upper two Quaternary zonesare absent at Site 495, and the first samples containAxoprunum angelinum, indicating an age of at least320,000 to 400,000 years. Gore 495-3 is placed in the
403

M. J. WESTBERG, W. R. RIEDEL
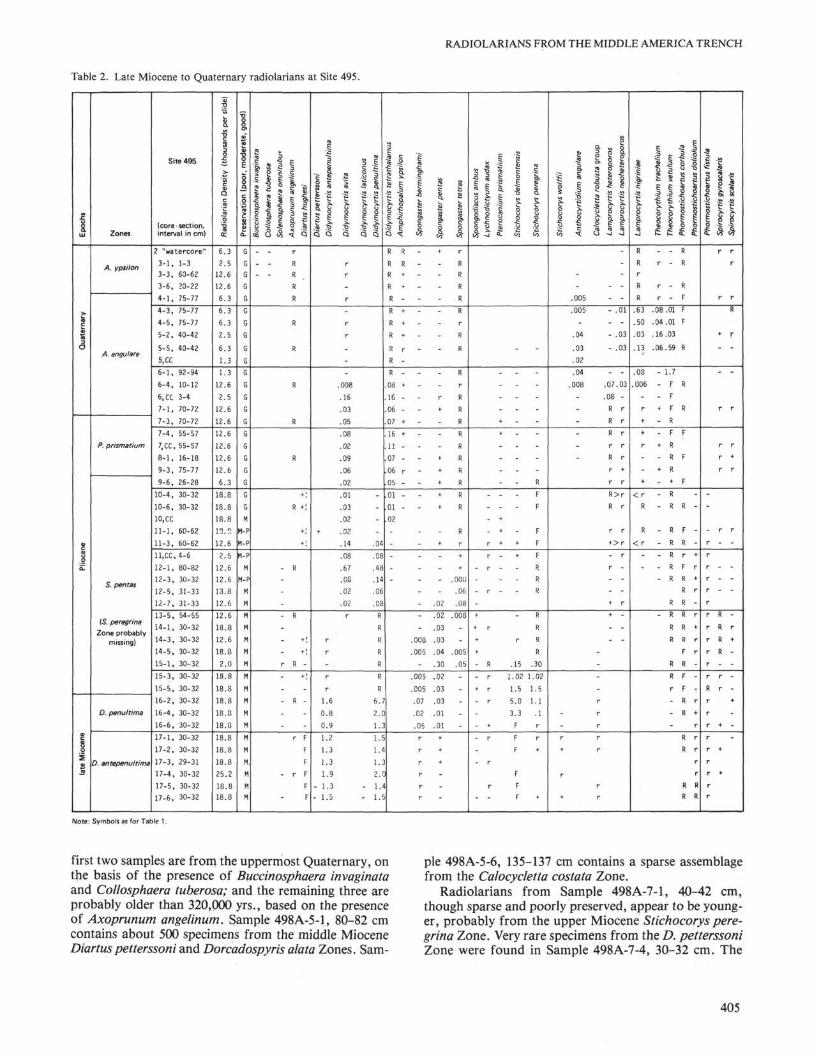
Amphirhopalum ypsilon Zone by the absence of Antho-cyrtidium angulare. Very rare occurrences of A. angu-lare in Cores 495-4 through -6 indicate the lowest Qua-ternary A. angulare Zone. The Pliocene/Pleistoceneboundary, and the top of the Pterocanium prismatiumZone, is placed between Samples 495-7-1, 70-72 cm and495-7-3, 70-72 cm by the latest occurrence of P. pris-matium. This designation, however, is tenuous becauseof the extreme rarity of P. prismatium and A. angulare.
The latest occurrence of Stichocorys peregrina be-tween Samples 495-9-3, 75-77 cm and 495-9-6, 26-28 cmmarks the bottom of the P. prismatium Zone.
The events indicating the bottoms of the Spongasterpentas and Stichocorys peregrina Zones, namely theevolutionary transitions between Spongaster berming-hami and S. pentas and between Stichocorys delmont-ensis and S. peregrina, respectively, occur in the sameinterval between Samples 495-16-2, 30-32 cm and 495-15-5, 30-32 cm, suggesting that the S. peregrina Zone iscompressed and not sampled. This conclusion is sup-ported by the latest occurrence of the Calocycletta ro-busta group in that same interval, an event known to oc-cur in the S. peregrina Zone.
The bottom of the Didymocyrtis penultima Zone isindicated by the latest occurrence of Diartus hughesibetween Samples 495-16-6, 30-32 cm and 495-17-1, 30-32 cm. The Didymocyrtis antepenultima Zone extendsfrom that interval down to the transition of Diartus pet-terssoni to D. hughesi between Samples 495-18-2, 90-92cm and 495-17-6, 30-32 cm. Relative abundances of lateMiocene to Quaternary radiolarians at Site 495 are re-corded in Table 2.
The muds of Core 495-18 and the carbonate-poor redclay of Core 495-19 belong to the middle Miocene D.petterssoni Zone. The earliest occurrence of D. petters-soni marks the bottom of this zone, between Samples495-19-6, 8-10 cm and 495-20-1, 60-62 cm.
Cores 495-20 through -37 are chalky carbonate oozewith chert in the lower section, and they continue a com-plete record of middle and lower Miocene radiolarianzones. The transition of Dorcadospyris dentata to D.alata, between Samples 495-23-6, 40-42 cm and 495-24-1, 15-17 cm, indicates the bottom of the D. alata Zone.The earliest occurrence of Calocycletta costata, betweenSamples 495-29-5, 19-21 cm and 495-30-1, 30-32 cm,marks the bottom of the C. costata Zone. The morpho-typic bottom of Stichocorys wolffii marks the bottomof the S. wolffii Zone, between Samples 495-32-2, 39-41 cm and 495-32-5, 39-41 cm. The remaining coresthrough 495-37-3 belong to the Stichocorys delmont-ensis Zone. Unusually low occurrences of Lithoperarenzae, S. wolfii, Phormostichoartus corbula, and Si-phostichoartus corona in abundances of < 0.1 °7o suggestdownworking between Cores 495-26 through -36. Sam-ples taken from Cores 495-38 through -43 and -46 wereall barren of radiolarians. Table 3 records abundancesof early to middle Miocene radiolarians at Site 495.
SITE 496At midslope Site 496, the objective was to penetrate a
landward-dipping seismic reflector, but drilling was ter-
minated short of this goal because of recovery of gase-ous hydrocarbons. The upper 226 meters of Hole 496are biogenic mud, but the well-preserved Quaternary ra-diolarians are diluted by 50 to 90% terrigenous debris inthe sand-size fraction.
Sample 496-1-1, 50-52 cm is assigned to the Collo-sphaera tuberosa Zone on the basis of the presence ofthat species. Between Sections 496-1-5 and 496-21-6,radiolarian assemblages belong to the Amphirhopalumypsilon Zone. Samples 496-22-4, 65-67 cm and 496-23-6, 32-34 cm are assigned to the Anthocyrtidium angu-lare Zone on the basis of very rare occurrences of A. an-gulare. The top of the Spongaster pentas Zone is placedbetween Samples 496-27-1, 70-72 cm and 496-27-4,29-31 cm by the latest occurrence of Stichocorys pere-grina. All of these zonal assignments are tenuous be-cause of the very low numbers of index species. Relativeabundances of Quaternary and Pliocene radiolarians atSite 496 are recorded in Table 4.
Samples 496-28-4, 87-89 cm and 496-28-5, 140-142cm are assigned to the 5. peregrina Zone because S. del-montensis is present, but in smaller numbers than S.peregrina. The upper Miocene Didymocyrtis penultimaand D. antepenultima Zones were not observed at thissite.
Samples 496-29-1, 90-92 cm and 496-29-3, 40-42 cmbelong to the middle Miocene Diartus petterssoni Zone.The Dorcadospyris alata Zone is indicated in Sample496-30-2, 45-47 cm by the presence of D. alata withoutD. dentata. Equal numbers of those two species in Core31, and their absence from samples from Cores 33 and34, prevent zonal assignments. However, all the samplestaken between Sections 496-35-2 and 496-40-1 appearto be in the Calocycletta costata Zone. Relative abun-dances of middle Miocene radiolarians at Site 496 arerecorded in Table 5.
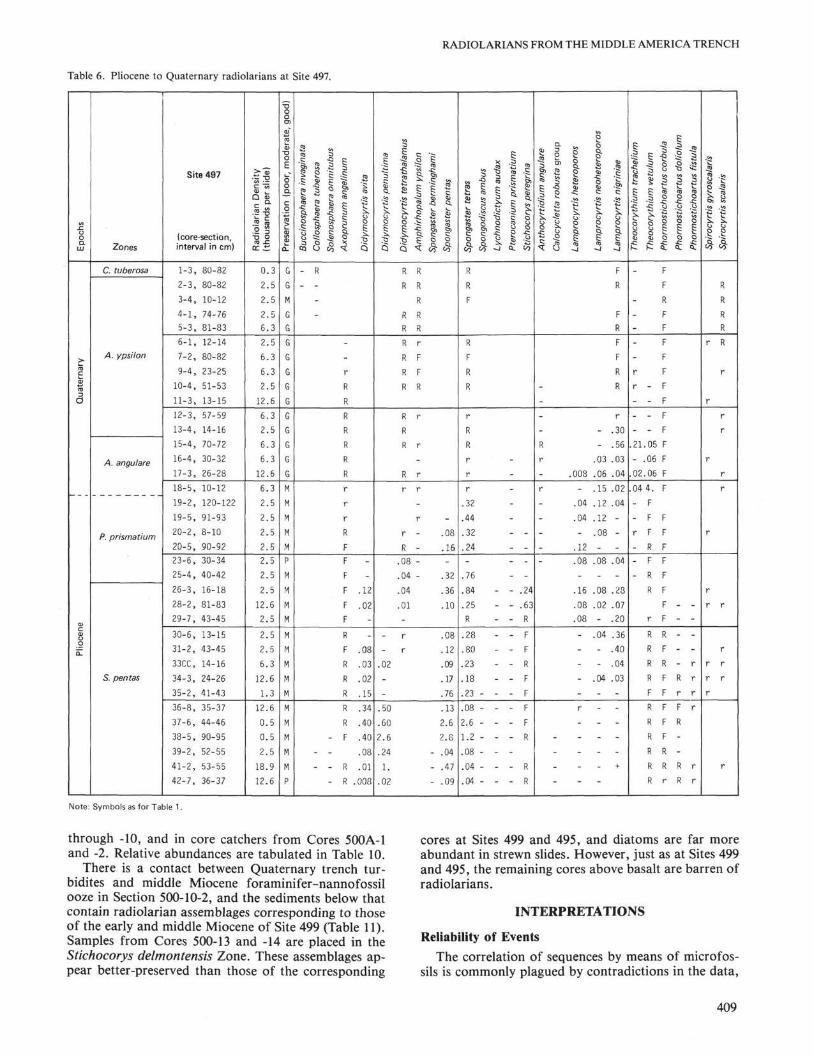
SITE 497Site 497 was drilled on the midslope 8 km seaward of
Site 496 and was also terminated early because of re-covery of gaseous hydrocarbons. The sediments are pre-dominantly dark olive gray muds containing moderatelywell-preserved Quaternary to Pliocene radiolarian as-semblages highly diluted with terrigenous components.Relative abundances of radiolarians and zonations areshown in Table 6.
SITE 498Site 498 was drilled 1 km east of Site 494 on the lower
slope to avoid a transverse fault and improve corerecovery. Again, the hole was abandoned because of gasrecovery. Most of the poorly recovered cores are drillingbreccia composed of dark olive gray mudstone, withchips of blue gray Miocene mudstone. Many of the sam-ples from Site 498 yielded too few radiolarians for strati-graphic interpretation. Reworking of Miocene and some-times Eocene species into younger sediments is frequentthroughout the entire section.
Samples 498-1-1, 28-30 cm, 498-1-3, 128-130 cm,498-2-1, 30-32 cm, 498A-2,CC (14-16 cm), and 498A-3,CC (25-27 cm) contain Quaternary radiolarians. The
404
RADIOLARIANS FROM THE MIDDLE AMERICA TRENCH
Table 2. Late Miocene to Quaternary radiolarians at Site 495.
Zones
Site 495
(core section,interval in cm)
c S £ •So ε S S
.5 ^
I Ilia ε 1 8 I i 1
1118 S S
3 S
i i
I ! 11 ! !oo ^ O
i, S
A. ypsilon
2 "watercore"
3-1, 1-3
3-3, 60-62
3-6, >O-22
4-1, 75-77
6.3
2.5
12.6
12.6
6.3 .005
- R- R
- R
- F
A. angulare
4-3, 75-77
4-5, 75-77
5-2, 40-42
5-5, 40-42
5,CC
6 . 3
6 . 3
2 . 5
6 . 3
1.3
.04
.03
.02
- .01
- .03
- .03
.63 .08 .01 F
.50 .04.01 F
,03 .16.03
.13 .06.59 R
6-1, 92-94
6-4, 10-12
6,CC 3-4
7-1, 70-72
7-3, 70-72
1.3
12.6
2.5
12.6
12.6
.008
.16
.03
.05
.04 - -
.008 .07.03
. 0 3 - 1.7
.006 - F R
- F
+ F R
- R
P. prismatium
7-4, 55-57
7,CC, 55-57
8-1, 16-18
9-3, 75-77
9-6, 26-28
12.6
12.6
12.6
12.6
6.3
F F
R
R F
R
+ F
10-4, 30-32
10-6, 30-32
10.CC
11-1, 60-62
11-3, 60-62
18.8
18.8
18.8
13.3
12.6
R>r
R r
< rR
R
R R
F -
R -
S. pentas
{S. peregrins
Zone probablymissing)
ll,CC,4-6
12-1, 80-82
12-3, 30-32
12-5, 31-33
12-7, 31-33
12.6
13.8
12.6
- .001!
- .06
.02 .08
R r
R F
R R
R
R R
13-5, 54-55
14-1, 30-32
14-3, 30-32
14-5, 30-32
15-1, 30-32
12.6
18.8
12.6
18.8
2.0
.02 .008
.03
.03
.04 .005
.30 .05 R .15
R R
R R
R R
F
R R
D. penultima
15-3, 30-32
15-5, 30-32
16-2, 30-32
16-4, 30-32
16-6, 30-32
18.8
18.8
18.8
18.8
18.8
1.6
0.8
0.9
.005
.005
.07
.02
.05
- r 1 . 0 2 1 . 0 2
+ r 1 . 5 1 . 5
- r 5 . 0 1 . 1
3 . 3 . 1
- + F r
r r
r +
r r
r -
r - -
r - -
R -
R r
R +
R -
r -
r
D. antepenultima
17-1, 30-32
17-2, 30-32
17-3, 29-31
17-4, 30-32
17-5, 30-32
17-6, 30-32
18.8
18.8
18.8
25.2
18.8
18.8
r F
F
F
r F
1.2
1.3
1.3
1.9
1.3
1.5
r
R R
R R
Note: Symbols as for Table 1.
first two samples are from the uppermost Quaternary, onthe basis of the presence of Buccinosphaera invaginataand Collosphaera tuberosa; and the remaining three areprobably older than 320,000 yrs., based on the presenceof Axoprunum angelinum. Sample 498A-5-1, 80-82 cmcontains about 500 specimens from the middle MioceneDiartuspetterssoni and Dorcadospyris alata Zones. Sam-
ple 498A-5-6, 135-137 cm contains a sparse assemblagefrom the Calocycletta costata Zone.
Radiolarians from Sample 498A-7-1, 40-42 cm,though sparse and poorly preserved, appear to be young-er, probably from the upper Miocene Stichocorys pere-grina Zone. Very rare specimens from the D. petterssoniZone were found in Sample 498A-7-4, 30-32 cm. The
405
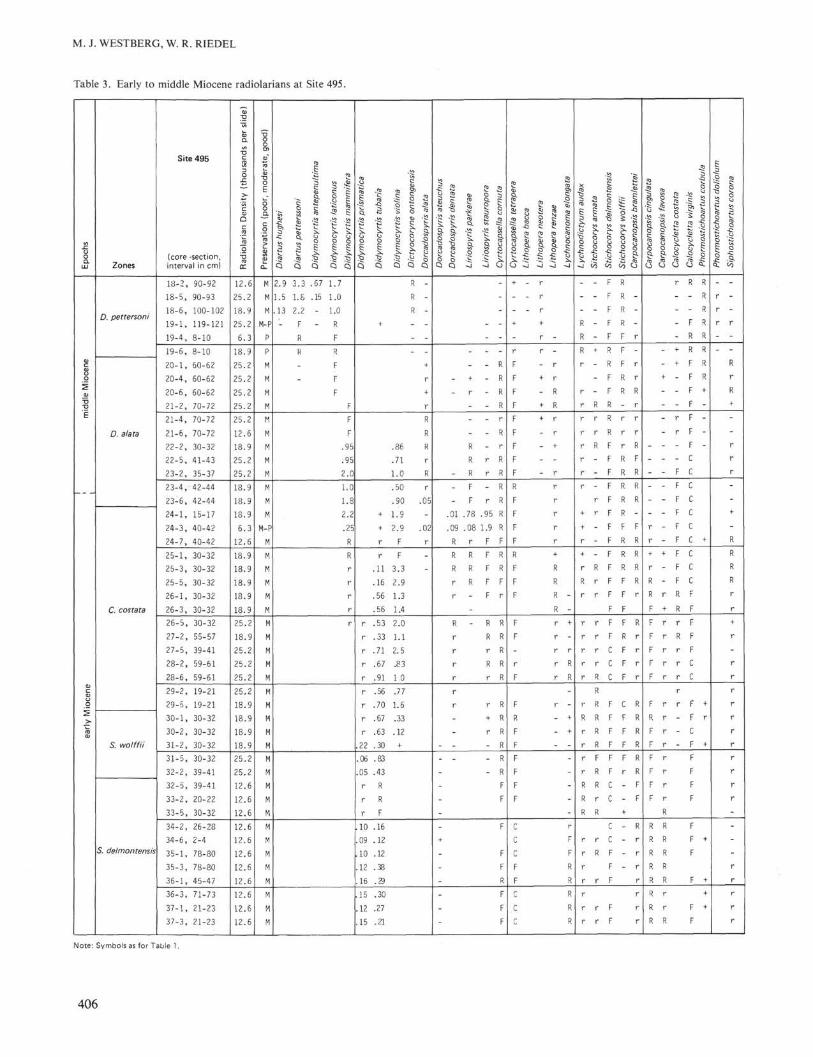
Table 3. Early to middle Miocene radiolarians at Site 495.jo
ch
s
LU
α>cα>u0
üα>
idd
ε
ene
ioc
>
ea
rl
Zones
D. pettersoni
D. alata
C. costata
S. wolffii
S. delmontensis
Site 495
(core -section,
interval in cm)
18-2,
18-5,
18-6,
19-1,
19-4,
19-6,
20-1,
20-4,
20-6,
21-2,
21-4,
21-6,
22-2,
22-5,
23-2,
23-4,
23-6,
24-1,
24-3,
24-7,
25-1,
25-3,
25-5,
26-1,
26-3,
26-5,
27-2,
27-5,
28-2,
28-6,
29-2,
29-5,
30-1,
30-2,
31-2,
31-5,
32-2,
32-5,
33-2,
33-5,
34-2,
34-6,
35-1,
35-3,
36-1,
36-3,
37-1,
37-3,
90-92
90-93
100-102
119-121
8-10
8-10
60-62
60-62
60-62
70-72
70-72
70-72
30-32
41-43
35-37
42-44
42-44
15-17
40-42
40-42
30-32
30-32
30-32
30-32
30-32
30-32
55-57
39-41
59-61
59-61
19-21
19-21
30-32
30-32
30-32
30-32
39-41
39-41
20-22
30-32
26-28
2-4
78-80
78-80
45-47
71-73
21-23
21-23
slid
e)
en
sity
(t
ho
usa
nd
s p
er:
ad
iola
ria
n D
cr
12.6
25.2
18.9
25.2
6.3
18.9
25.2
25.2
25.2
25.2
25.2
12.6
18.9
25.2
25.2
18.9
18.9
18.9
6.3
12.6
18.9
18.9
18.9
18.9
18.9
25.2
18.9
25.2
25.2
25.2
25.2
18.9
18.9
18.9
18.9
25.2
25.2
12.6
12.6
12.6
12.6
12.6
12.6
12.6
12.6
12.6
12.6
12.6
ao
or,
mo
de
rate
, g
oo
d)
ese
rva
tio
n (
α.
M
M
M
M-P
P
P
M
M
M
M
M
M
M
M
M
M
M
M
M-P
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
iart
us
hu
gh
e:
Q
2.9
1.5
13
-
\son
iia
rtu
s p
ett
en
Q
3.3
l.S
2.2
F
R
R
-
-
an
tep
en
ult
ima
idym
oc
yrti
s
Q
.67
.15
-la
tic
on
us
idym
oc
yrti
s
Q
1.7
1.0
1.0
R
F
R
F
F
F
to
>£
1
1
•d
ymo
cyrt
is
Q
F
F
F
.95
.95
2.0
1.0
1.8
2.2
.25
R
R
r
r
r
r
r
pri
sma
tic
aid
ymo
cyr
tis
Q
r
r
r
r
r
r
r
r
r
.22
.06
.05
r
r
r
.10
.09
,10
.12
.16
.15
.12
.15
tub
ari
aid
ymo
cyr
tis
Q
-t-
+
+
r
r
.11
.16
.56
.56
.53
.33
.71
.67
.91
.56
.70
.67
.63
.30
.83
.43
R
R
F
.16
.12
.12
.38
.29
.30
.27
.21
vio
lina
idym
oc
yrti
s
Q
.86
.71
1.0
.50
.90
1.9
2.9
F
F
3.3
2.9
1.3
1.4
2.0
1.1
2.5
.83
1.0
.77
1.6
.33
.12
+
on
ton
ge
nsi
sic
tyo
co
ryn
e
Q
R
R
R
-
-
-
ala
tao
rca
do
spyr
is
Q
-
-
-
-
+
r
+
r
R
R
R
r
R
r
.05
-
.02
r
-
-
i ate
uc
hu
so
rca
do
spyr
ii
Q
-
-
-
-
-
-
-
+
-
-
-
-
-
-
de
nta
tao
rca
do
spyr
is
Q
-
-
-
-
-
.01
.09
R
R
R
r
r
R
r
r
r
r
r
r
-
-
-
-
~ker
aeir
iosp
yris
pai
-
-
+
r
-
-
-
R
R
R
F
F
.78
.08
r
R
R
R
-
-
-
uro
po
rair
iosp
yris
sta
•~j
-
-
-
-
-
-
-
-
-
-
r
r
-
r
.95
1.9
F
F
F
F
F
R
R
r
R
r
r
+
r
-
-
-
co
rnu
tayr
toc
ap
seila
ü
-
-_
-
-
-
R
R
R
R
r
R
r
R
R
R
R
R
R
F
R
R
F
r
R
R
R
R
R
R
R
R
R
R
R
F
F
F
F
F
R
F
F
F
tetr
aper
ayr
toca
psei
la
+
-
_
+
-
r
F
F
F
F
F
F
F
F
F
R
F
F
F
F
R
F
F
F
F
F
-
r
F
F
R
F
F
F
F
F
F
C
ccF
F
C
C
C
'tho
pe
ra b
aα
-
-
tera
ith
op
era
ne
o
r
r
r
+
r
r
-
+
-
+
+
-
-
-
-
tbto
'th
op
era
re
n.
-
-
r
r
R
R
r
r
+
-
r
r
r
r
r
r
+
R
R
R
R
r
r
r
r
r
r
-
-
-
a e
lon
ga
tayc
hn
oc
an
om
-
-
+
-
r
R
R
-
-
+
+
-
-
-
-
-
-
r
F
F
R
R
R
R
R
m a
uda×
vch
no
dic
tyu
-
-
RR
R
r
r
r
r
r
r
r
r
r
+
+
r
+
r
R
r
r
r
r
r
r
r
R
r
r
r
r
R
R
R
r
r
r
r
r
r
r
2to
|
CO
oo
ito-
-
_
-
-
+
-
-
-
R
r
r
R
-
-
-
r
r
-
-
-
R
r
r
r
r
r
r
R
R
R
R
R
R
F
R
R
r
R
r
R
r
r
r
i/mon
tens
is'ic
hoco
rys
di
to
F
F
F
FF
R
R
F
F
R
R
R
F
F
F
F
F
F
F
F
F
F
F
F
F
F
F
C
cc
F
F
F
F
F
F
C
C
ccF
F
F
F
F
oiff
ii'ic
hoco
rys
w
CO
R
R
R
RF
F
F
R
R
-
r
r
r
R
R
R
R
R
F
R
R
R
F
F
F
F
R
F
F
F
C
F
F
F
F
r
-
-
+
-
-
-
-
bra
mle
ttei
irpoc
anop
sis
-
-r
-
r
r
R
r
r
r
R
F
R
R
R
-
F
R
R
R
R
r
R
r
r
r
r
R
R
R
R
R
R
F
F
R
r
r
r
r
r
r
r
cinguia
ta
11g.
------r
r
+
r
R
R
F
F
F
F
F
F
F
R
F
F
F
F
F
F
R
R
R
R
R
R
R
R
: fa
vosa
arpo
cano
psis
o
-
-
+
-
-
-
-
-
-
-
-
-
-
-
-
+
-
-
r
+
r
r
r
r
r
r
r
r
r
r
r
r
r
R
R
R
R
R
R
r
r
R
osta
taC
alo
cycl
etta c
r
-
-
-+
+
-
-
-
r
r
-
-
F
F
F
F
F
F
F
F
F
R
R
r
R
r
r
r
r
r
-
-
-
•irgi
nis
alo
cycl
ett
a u
R
-_
FR
R
F
F
F
F
F
F
F
C
C
ccccccccF
F
F
F
F
C
c
F
F
C
F
F
F
F
F
F
F
F
F
F
F
artu
s co
rbula
o
εa.
R
R
R
RR
R
R
R
+
-
-
-
-
+
+
r
+
+
+
+
+
art
us
do
liolu
mh
orm
ost
ich
o
-
r
r
r
-
-
tus
co
ron
aip
ho
stic
ho
ar
-
-
r
-
-
R
r
R
+
-
-
r
r
r
-
-
-
R
R
R
R
r
r
+
r
-
r
r
r
r
r
r
r
r
r
r
r
-
-
-
-
r
r
r
r
r
Note: Symbols as for Table 1.
406
M. J. WESTBERG, W. R. RIEDEL
RADIOLARIANS FROM THE MIDDLE AMERICA TRENCH
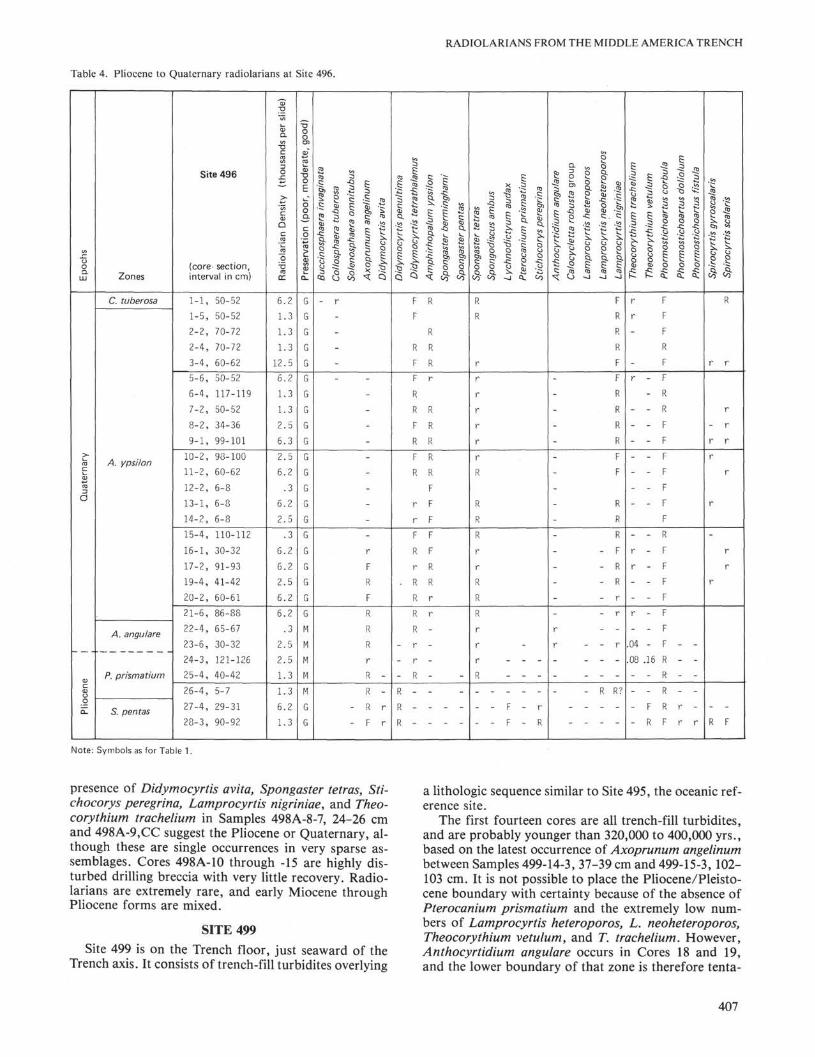
Table 4. Pliocene to Quaternary radiolarians at Site 496.E
po
chs
>roc
α>
ua
t
σ
cen
Plio
Zones
C. tuberosa
A. ypsiion
A. angulare
P. prismatium
S. pentas
Site 496
(core section,interval in cm)
1-1,
1-5,
2-2,
2-4,
3-4,
5-6,
6-4,
7-2,
8-2,
9 - 1 ,
10-2,
11-2,
12-2,
13-1,
14-2,
15-4,
1 6 - 1 ,
17-2,
19-4,
20-2,
21-6,
22-4,
23-6,
24-3,
25-4,
26-4,
27-4,
28-3,
50-52
50-52
70-72
70-72
60-62
50-52
117-119
50-52
34-36
99-101
98-100
60-62
6-8
6-8
6-8
110-112
30-32
91-93
41-42
60-61
86-88
65-67
30-32
121-126
40-42
5-7
29-31
90-92
Iide)
ds p
er s
c
Ra
dio
lari
an
De
ns
ity
(th
ou
sa
6.2
1.3
1.3
1.3
12.5
6.2
1.3
1.3
2.5
6.3
2.5
6.2
.3
6.2
2.5
.3
6.2
6.2
2.5
6.2
6.2
.3
2.5
2.5
1.3
1.3
6.2
1.3
, g
oo
d)
O)
Pre
serv
ati
on
(p
oo
r, m
od
era
tG
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
M
M
M
M
M
G
G
Bu
ccin
osp
ha
era
inva
gin
ata
-
Co
llosp
ha
era
tu
be
rosa
r
-
-
-
-
-
So
len
osp
ha
era
om
nit
ub
us
-
-
Axo
pru
nu
m a
ng
elin
um
-
-
-
-
-
_
-
-
-
-
-
r
F
R
F
R
R
R
r
R
R
R
F
Did
ymo
cyr
tis
avi
ta
-
-
r
r
Did
ymo
cyr
tis
pe
nu
ltim
a
-
-
-
R
R
R
Did
ymo
cyr
tis
tetr
ath
aia
mu
s
F
F
R
F
F
R
R
F
R
F
R
r
r
F
R
r
R
R
R
R
r
r
R
-
-
-
Am
ph
irh
op
aiu
m
ypsi
ion
R
R
R
R
r
R
R
R
R
R
F
F
F
F
F
R
R
r
r
-
-
-
-
-
-
-
Sp
on
gas
ter b
erm
ing
ha
mi
-
-
Sp
on
gas
ter
pe
nta
s
-
-
-
-
Sp
on
gas
ter
tetr
as
R
R
r
r
r
r
r
r
r
R
R
R
R
r
r
R
R
R
r
r
r
R
-
-
Sp
on
go
dis
cus
am
bu
s
-
-
Lyc
hn
od
icty
um
a
ud
ax
-
-
F
F
Pte
roc
an
ium
pri
sma
tiu
m
-
-
-
-
-
Sti
ch
oc
ory
s p
ere
gri
na
-
-
V
R
An
tho
cy
rtid
ium
an
gu
lare
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
r
r
-
-
Ca
loc
ycle
tta
ro
bu
sta
gro
up
-
-
La
mp
roc
yrti
s h
ete
rop
oro
s
-
-
-
La
mp
roc
yrti
s n
eo
he
tero
po
ro
-
-
-
-
-
-
-
-
R
-
-
La
mp
roc
yrti
s n
igri
nia
e
F
R
R
R
F
F
R
R
R
R
F
F
R
R
R
F
R
R
r
r
-
r
-
R?
-
-
Th
eo
co
ryth
ium
tra
ch
eliu
m
r
r
-
-
r
-
-
-
_
-
-
-
-
r
r
-
-
r
-
.04
.08
-
-
-
Th
eo
co
ryth
ium
ve
tulu
m
-
-
-
-
-
_
-
-
-
-
-
-
-
-
-
-
-
.15
-
F
R
Ph
orm
ost
ich
oa
rtu
s c
orb
uia
F
F
F
R
F
F
R
R
F
F
F
F
F
F
F
R
F
F
F
F
F
F
F
R
R
R
R
F
Ph
orm
ost
ich
oa
rtu
s d
olio
lum
-
-
-
r
r
Ph
orm
ost
ich
oa
rtu
s fi
stu
la
-
-
-
-
rS
pir
oc
yrti
s g
yro
sca
lari
s
r
-
r
r
r
-
r
-
RS
pir
oc
yrti
s sc
alar
is
R
r
r
r
r
r
r
r
-
F
Note: Symbols as for Table 1.
presence of Didymocyrtis αvitα, Spongaster tetras, Sti-chocorys peregrina, Lamprocyrtis nigriniae, and Theo-corythium trachelium in Samples 498A-8-7, 24-26 cmand 498A-9,CC suggest the Pliocene or Quaternary, al-though these are single occurrences in very sparse as-semblages. Cores 498A-10 through -15 are highly dis-turbed drilling breccia with very little recovery. Radio-larians are extremely rare, and early Miocene throughPliocene forms are mixed.
SITE 499
Site 499 is on the Trench floor, just seaward of theTrench axis. It consists of trench-fill turbidites overlying
a lithologic sequence similar to Site 495, the oceanic ref-erence site.
The first fourteen cores are all trench-fill turbidites,and are probably younger than 320,000 to 400,000 yrs.,based on the latest occurrence of Axoprunum angelinumbetween Samples 499-14-3, 37-39 cm and 499-15-3, 102-103 cm. It is not possible to place the Pliocene/Pleisto-cene boundary with certainty because of the absence ofPterocanium prismatium and the extremely low num-bers of Lamprocyrtis heteroporos, L. neoheteroporos,Theocorythium vetulum, and T. trachelium. However,Anthocyrtidium angulare occurs in Cores 18 and 19,and the lower boundary of that zone is therefore tenta-
407
M. J. WESTBERG, W. R. RIEDEL
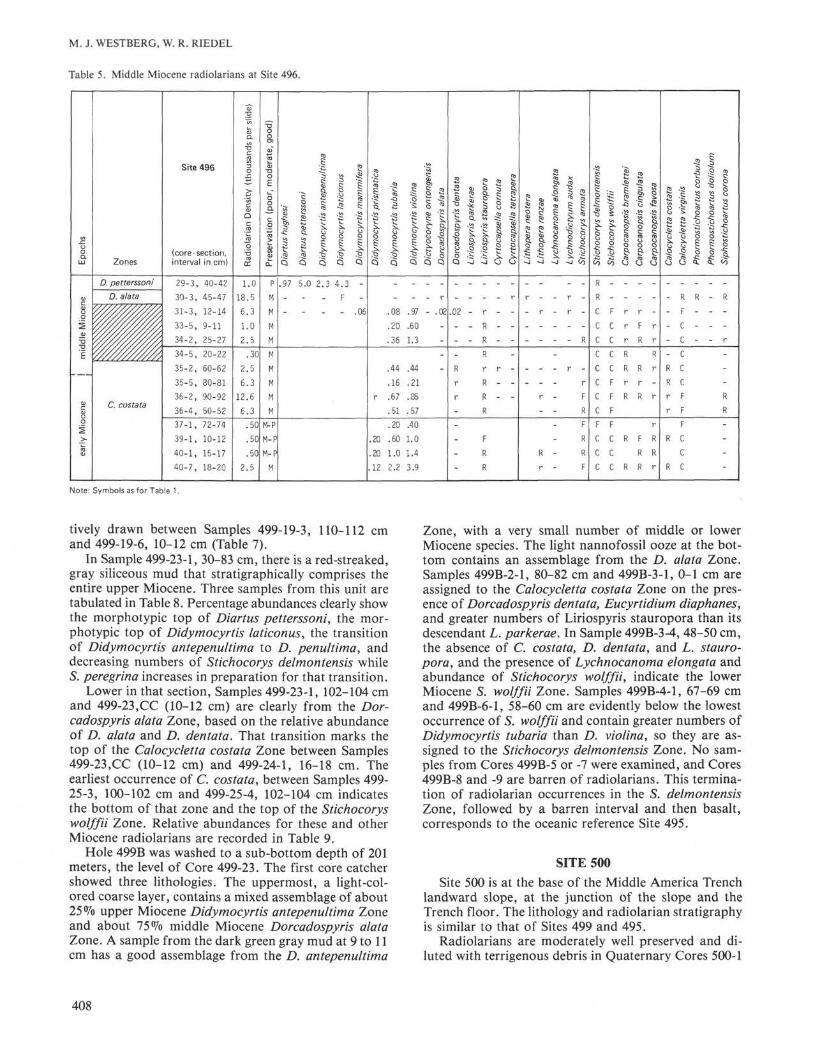
Table 5. Middle Miocene radiolarians at Site 496.
uo
LU
cuoi
idle
ε
i/l ioc
ene
>α>
Zones
D. petterssoni
D. aiata
^ ^ ^ ^ ^ ^
C. costata
Site 496
(core-section,interval in cm)
29-3,
30-3
31-3,
33-5,
34-2,
34-5,
35-2,
35-5,
36-2,
36-4,
37-1,
39-1,
40-1,
40-7,
40-42
45-47
12-14
9-11
25-27
20-22
60-62
80-81
90-92
50-52
72-74
10-12
15-17
18-20
slid
e)d
iola
rian
Den
sity
(th
ousa
nd
oc
1.0
18 5
6.3
1.0
2.5
.30
2.5
6.3
12.6
6.3
.50
.50
.50
2.5
8oσ>
eser
vatio
n (p
oor,
mod
erat
e,α_
P
M
M
M
M
M
M
M
M
M
M-P
M-P
M-P
M
artu
s hu
ghes
i
Q
.97
_ar
tus
pett
erss
oni
Q
5.0
-dy
moc
yrtis
an
tepe
nulti
ma
Q
2.3
_
dym
ocyr
tis
Iatic
onus
Q
4.3p
idym
ocyr
tis
mam
mife
ra
Q
-
.06
dym
ocyr
tis
pris
mat
ica
Q
r
.20
.20
.12
dym
ocyr
tis
tuba
ria
Q
-
.08
.20
.36
.44
.16
.67
.51
.20
.60
1.0
2.2
idym
ocyr
tis
vio
lina
Q
.97
.60
1.3
.44
.21
.85
.57
.40
1.0
1.4
3.9
ctyo
cory
ne o
nton
gens
is
Q
_
orca
dosp
yris
aia
ta
Q
.02
-
-
-
-
orca
dosp
yris
den
tata
Q
.02
-
-
-
R
r
r
-
-
-
-
-rio
spyr
is p
arke
rae
_
-
-
riosp
yris
sta
urop
ora
- j
r
R
R
R
r
R
R
R
F
R
R
/rto
caps
eiia
cor
nuta
-
-
r
-_
Cyr
toca
psel
la t
etra
pera
-
-
-
-
-_
thop
era
neot
era
-
-
-
-
thop
era
renz
ae
- j
r
-
-
-
-
r
-
R
r
/chn
ocan
oma
eion
gata
-
-
-
-
-
-_
-
-
-
-
-
ychn
odic
tyum
aud
a×
-1
r-
-
r
icho
cory
s ar
mat
a
CO
-
-
R
-rFRFR
R
F
icho
cory
s de
lmon
tens
is
CO
Rp
C
C
ccccccF
ccc
icho
cory
s w
oiffi
i
CO
F
CC
ccFFFF
C
cc
irpoc
anop
sis
bram
lette
i
cj
rrr
RRrR
R
R
irpoc
anop
sis
cing
ulat
a
0
rFR
R
rR
F
RR
irpoc
anop
sis
favo
sa
cj
-
rr
Rr
-r
rR
Rr
ilocy
clet
ta v
irgin
is
Cj G
-
-
-
-
RP
rr
P
R
R
F
CCC
C
cFFF
CC
c
^orm
ostic
hoar
tus
corb
ula
p
-
-
norm
ostic
hoar
tus
dolio
lum
-
-
-
phos
ticho
artu
s co
rona
CO
R
-r
-
--RR
---
-
Note: Symbols as for Table 1.
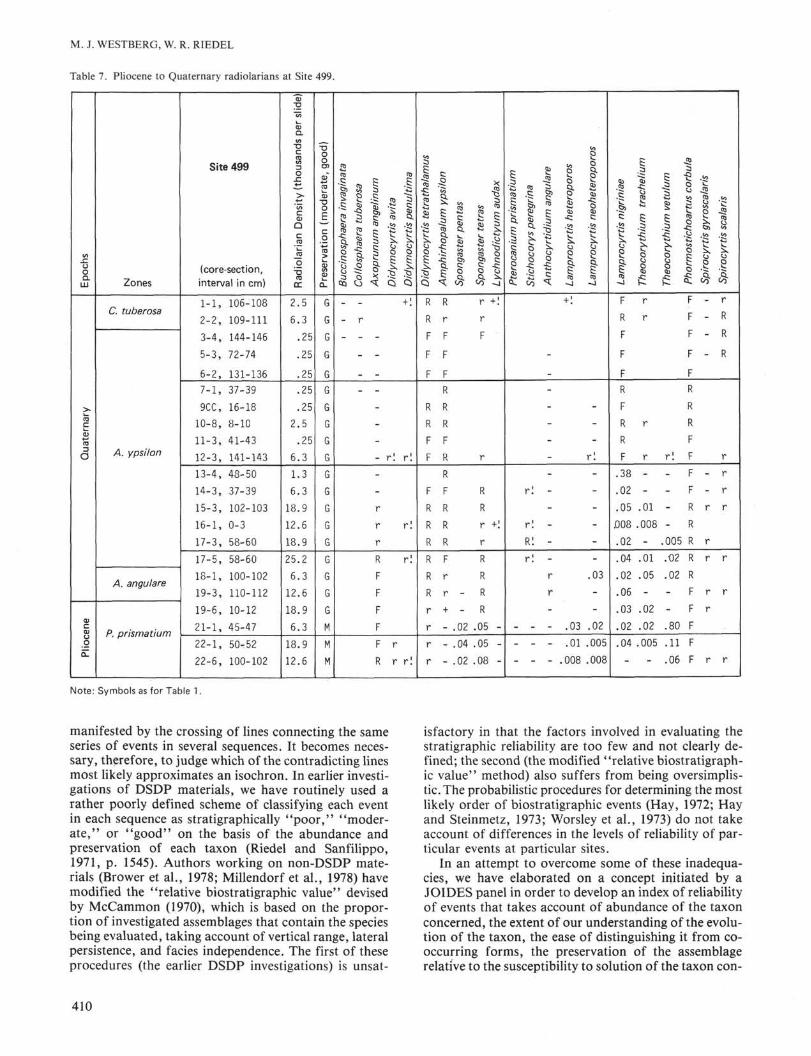
tively drawn between Samples 499-19-3, 110-112 cmand 499-19-6, 10-12 cm (Table 7).
In Sample 499-23-1, 30-83 cm, there is a red-streaked,gray siliceous mud that stratigraphically comprises theentire upper Miocene. Three samples from this unit aretabulated in Table 8. Percentage abundances clearly showthe morphotypic top of Diαrtus petterssoni, the mor-photypic top of Didymocyrtis Iaticonus, the transitionof Didymocyrtis antepenultima to D. penultima, anddecreasing numbers of Stichocorys delmontensis whileS. peregrina increases in preparation for that transition.
Lower in that section, Samples 499-23-1, 102-104 cmand 499-23,CC (10-12 cm) are clearly from the Dor-cadospyris aiata Zone, based on the relative abundanceof D. aiata and D. dentata. That transition marks thetop of the Calocycletta costata Zone between Samples499-23,CC (10-12 cm) and 499-24-1, 16-18 cm. Theearliest occurrence of C. costata, between Samples 499-25-3, 100-102 cm and 499-25-4, 102-104 cm indicatesthe bottom of that zone and the top of the Stichocoryswoiffii Zone. Relative abundances for these and otherMiocene radiolarians are recorded in Table 9.
Hole 499B was washed to a sub-bottom depth of 201meters, the level of Core 499-23. The first core catchershowed three lithologies. The uppermost, a light-col-ored coarse layer, contains a mixed assemblage of about25% upper Miocene Didymocyrtis antepenultima Zoneand about 75% middle Miocene Dorcadospyris aiataZone. A sample from the dark green gray mud at 9 to 11cm has a good assemblage from the D. antepenultima
Zone, with a very small number of middle or lowerMiocene species. The light nannofossil ooze at the bot-tom contains an assemblage from the D. aiata Zone.Samples 499B-2-1, 80-82 cm and 499B-3-1, 0-1 cm areassigned to the Calocycletta costata Zone on the pres-ence of Dorcadospyris dentata, Eucyrtidium diaphanes,and greater numbers of Liriospyris stauropora than itsdescendant L. parkerae. In Sample 499B-3-4, 48-50 cm,the absence of C. costata, D. dentata, and L. stauro-pora, and the presence of Lychnocanoma eiongata andabundance of Stichocorys woiffii, indicate the lowerMiocene S. woiffii Zone. Samples 499B-4-1, 67-69 cmand 499B-6-1, 58-60 cm are evidently below the lowestoccurrence of S. woiffii and contain greater numbers ofDidymocyrtis tubaria than D. violina, so they are as-signed to the Stichocorys delmontensis Zone. No sam-ples from Cores 499B-5 or -7 were examined, and Cores499B-8 and -9 are barren of radiolarians. This termina-tion of radiolarian occurrences in the S. delmontensisZone, followed by a barren interval and then basalt,corresponds to the oceanic reference Site 495.
SITE 500
Site 500 is at the base of the Middle America Trenchlandward slope, at the junction of the slope and theTrench floor. The lithology and radiolarian stratigraphyis similar to that of Sites 499 and 495.
Radiolarians are moderately well preserved and di-luted with terrigenous debris in Quaternary Cores 500-1
408
RADIOLARIANS FROM THE MIDDLE AMERICA TRENCH
Table 6. Pliocene to Quaternary radiolarians at Site 497.E
po
chs
> •
<oc
atei
α
<L>
Plio
cen
Zones
C. tuberosa
A. ypsilon
A. anguiare
P. prismatium
S. pentas
Site 497
(core-section,interval in cm)
1-3,
2-3,
3-4,
4 - 1 ,
5-3,
6 - 1 ,
7-2,
9-4,
10-4,
11-3,
12-3,
13-4,
15-4,
16-4,
17-3,
18-5,
19-2,
19-5,
20-2,
20-5,
23-6,
25-4,
26-3,
28-2,
29-7,
30-6,
31-2,
33CC,
34-3,
35-2,
36-8,
37-6,
38-5,
39-2,
41-2,
42-7,
80-82
80-82
10-12
74-76
81-83
12-14
80-82
23-25
51-53
13-15
57-59
14-16
70-72
30-32
26-28
10-12
120-122
91-93
8-10
90-92
30-34
40-42
16-18
81-83
43-45
13-15
43-45
14-16
24-26
41-43
35-37
44-46
90-95
52-55
53-55
36-37
sity
ide)
Ra
dio
lari
an
De
n(t
hous
and
s p
er
si0.3
2.5
2.5
2.5
6.3
2.5
6.3
6.3
2.5
12.6
6.3
2.5
6.3
6.3
12.6
6.3
2.5
2.5
2.5
2.5
2.5
2.5
2.5
12.6
2.5
2.5
2.5
6.3
12.6
1.3
12.6
0.5
0.5
2.5
18.9
12.6
| (P
oo
22CO
or,
mo
de
rP
rese
rvat
ion
(p
o
G
G
M
G
G
G
G
G
G
G
G
G
G
G
G
M
M
M
M
MP
M
M
M
M
M
M
M
M
M
M
M
M
M
M
P
nvag
inat
aB
ucci
nosp
haer
a /
-
-
eros
a
1to
-c
S•o
δ
R
-
-
-
-
-
3•o
a1
Soi
enos
pha
era
oi
----
?lin
umA
×op
runu
m a
ngt
--r
R
R
R
R
R
R
R
r
r
r
R
FF
F
F
F
F
R
F
R
R
R
R
R
F
R
R
itaD
idym
ocyr
tis a
v,
-
-
.12
.02
-
-
.08
.03
.02
.15
.34
.40
.40
.08
.01
.008
nulti
ma
Did
ymoc
yrtis
pe
-
-
.02
-
-
.50
.60
2.6
.24
1.
.02
rath
alam
uD
idym
ocyr
tis t
et
R
R
R
R
R
R
R
R
R
R
R
R
r
r
R.08
.04
.04
.01
-
r
r
ypsi
lon
Am
phirh
opal
um
R
R
R
R
R
r
F
F
R
r
r
-
r
r
-
r
-
-
-
-
<ngh
ami
Spo
ngas
ter
berm
,
-
-
-
is
Spo
ngas
ter
pent
i
-
.08
.16-
.32
.36
.10
.08
.12
.09
.17
.76
.13
2.6
2.8
.04
.47
.09
Spo
ngas
ter
tetr
a
R
R
F
R
F
R
R
r
R
R
r
r
r
.32
.44
.32
.24-
.76
.84
.25
R
.28
.80
.23
.18
.23
.08
2.6
1.2
.08
.04
.04
3
Spo
ngod
iscu
s a
n
-
-
-
-
-
-
-
auda
×Ly
chno
dict
yum
-
-
-
-
-
-
-
-
-
-
-
-
-
-
'<m
atiu
m
Q.
.3
11
-----------------------
grin
a1
Stic
hoco
rys
pere
----.24
.63
R
F
F
R
F
F
F
F
R
R
R
angu
iare
An th
ocy
rtid
ium
-
-
-
-
R
r
-
r
-
-
-
-
-us
ta g
roup
Cal
ocyc
letta
rob
-
-
-
-
lero
poro
sLa
mpr
ocyr
tis h
e,
.008
-
.04
.04
-
.12
.08
-
.16
.08
.08
-
-
-
-
-
r
-
-
-
-
-
o
ohet
erop
oiLa
mpr
ocyr
tis n
e
-
-
.03
.06
.15
.12
.12
.08
-.08
-
.08
.02
-
.04
-
-
.04
-
-
-
-
-
-
-
irini
aeLa
mpr
ocyr
tis n
ic,
F
R
FR
F
F
R
R
r
.30
.56
.03
.04
.02
.04
-
-
-.04
-
.28
.07
.20
.36
.40
.04
.03
-
-
-
-
-
+
trac
heliu
m1
The
ocor
ythi
um ;
-
-
-
-
-
-
r
r
-
-
-
.21
-.02
.04
-
-
r
--
-
ε.3
Th
eo
cory
thiu
m i
-
-
-
-
.05
.06
.06
4.
F
F
F
R
F
R
R
r
R
R
R
R
F
R
R
R
R
R
R
"us
corb
ula
| P
ho
rmo
stic
ho
an
F
F
R
F
F
F
F
F
F
F
F
F
F
F
F
F
F
F
F
F
F
F
F
F
R
F
R
F
F
F
F
F
R
R
r
ε
'us
do
liolu
Ph
orm
ost
ich
oan
-----R
r
F
R
-
-
R
R
'us
fistu
laP
ho
rmo
stic
ho
an
-
-
-
-
r
r
r
r
r
r
scal
aris
Sp
iro
cyrt
is g
yro,
r
r
r
r
r
r
r
r
r
ris
Sp
iro
cyrt
is s
cala
R
R
R
R
R
r
r
r
r
r
r
r
r
r
r
Note: Symbols as for Table 1.
through -10, and in core catchers from Cores 500A-1and -2. Relative abundances are tabulated in Table 10.
There is a contact between Quaternary trench tur-bidites and middle Miocene foraminifer-nannofossilooze in Section 500-10-2, and the sediments below thatcontain radiolarian assemblages corresponding to thoseof the early and middle Miocene of Site 499 (Table 11).Samples from Cores 500-13 and -14 are placed in theStichocorys delmontensis Zone. These assemblages ap-pear better-preserved than those of the corresponding
cores at Sites 499 and 495, and diatoms are far moreabundant in strewn slides. However, just as at Sites 499and 495, the remaining cores above basalt are barren ofradiolarians.
INTERPRETATIONS
Reliability of Events
The correlation of sequences by means of microfos-sils is commonly plagued by contradictions in the data,
409
M. J. WESTBERG, W. R. RIEDEL
lableE
poc
hsQ
uat
ern
ary
Plio
cene
7. Pliocene to Quaternary radiolaπans
Zones
C. tuberosa
A. ypsilon
A. angulare
P. prismatium
Site 499
(core-section,interval in cm)
1-1, 106-108
2-2, 109-111
3_4, 144-146
5-3, 72-74
6-2, 131-136
7-1, 37-39
9CC, 16-18
10-8, 8-10
11-3, 41-43
12-3, 141-143
13-4, 48-50
14-3, 37-39
15-3, 102-103
16-1, 0-3
17-3, 58-60
17-5, 58-60
18-1, 100-102
19-3, 110-112
19-6, 10-12
21-1, 45-47
22-1, 50-52
22-6, 100-102
at Site 499
Ra
dio
lari
an
De
nsi
ty (
thou
sand
s p
er s
lide)
2.5
6.3
.25
.25
.25
.25
.25
2.5
.25
6.3
1.3
6.3
18.9
12.6
18.9
25.2
6.3
12.6
18.9
6.3
18.9
12.6
Pre
serv
atio
n (m
od
era
te, g
oo
d)
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
M
M
M
Buc
cino
spha
era
in v
agin
a ta
Col
losp
haer
a tu
bero
sa
A×
opru
num
ang
eiin
um
Did
ymoc
yrtis
a vi
ta
Did
ymoc
yrtis
pen
ultim
a
i i
i
1 1
1 -s
1
1 1
1
- r\ r\
rr r!rR r!
F
F
F
F
F r
R r r !
Did
ymoc
yrtis
te
trat
hala
mus
Am
phirh
opal
um
ypsi
lon
Spo
ngas
ter
pent
as
Spo
ngas
ter
tetra
s
Lych
nodi
ctyu
m a
uda×
R R r +!
R r r
F F F •
F F
F F
R
R R
R R
F F
F R r
R
F F R
R R R
R R r +:
R R r
R F R
R r R
R r - R
r + - R
r - . 02 .05 -
r - .04 .05 -
r - .02 .08 -
Pte
roca
nium
pris
mat
ium
Stic
hoco
rys
pere
grin
a
Anth
ocy
rtid
ium
ang
ular
e
Lam
proc
yrtis
het
erop
oros
Lam
proc
yrtis
neo
hete
ropo
ros
-
r!
rl -
rl -
R : -rl -
r . 0 3
r
- - - . 03 .02
- - - .01 .005
- - - .008 .008
Lam
proc
yrtis
nig
rinia
e
The
ocor
ythi
um
trac
heiiu
m
The
ocor
ythi
um v
etul
um
Pho
rmos
ticho
artu
s co
rbui
a
Spi
rocy
rtis
gyr
osca
laris
Spi
rocy
rtis
sca
iaris
F r F - r
R r F - R
F F - R
F F - R
F F
R R
F R
R r R
R F
F r r ! F r
.38 - - F - r
.02 - - F - r
.05 .01 - R r r
.008 .008 - R
.02 - .005 R r
.04 .01 .02 R r r
.02 .05 .02 R
.06 - - F r r
.03 .02 - F r
.02 .02 .80 F
.04 .005 .11 F
- .06 F r r
Note: Symbols as for Table 1.
manifested by the crossing of lines connecting the sameseries of events in several sequences. It becomes neces-sary, therefore, to judge which of the contradicting linesmost likely approximates an isochron. In earlier investi-gations of DSDP materials, we have routinely used arather poorly defined scheme of classifying each eventin each sequence as stratigraphically "poor," "moder-ate," or "good" on the basis of the abundance andpreservation of each taxon (Riedel and Sanfilippo,1971, p. 1545). Authors working on non-DSDP mate-rials (Brower et al., 1978; Millendorf et al., 1978) havemodified the "relative biostratigraphic value" devisedby McCammon (1970), which is based on the propor-tion of investigated assemblages that contain the speciesbeing evaluated, taking account of vertical range, lateralpersistence, and facies independence. The first of theseprocedures (the earlier DSDP investigations) is unsat-
isfactory in that the factors involved in evaluating thestratigraphic reliability are too few and not clearly de-fined; the second (the modified "relative biostratigraph-ic value" method) also suffers from being oversimplis-tic. The probabilistic procedures for determining the mostlikely order of biostratigraphic events (Hay, 1972; Hayand Steinmetz, 1973; Worsley et al., 1973) do not takeaccount of differences in the levels of reliability of par-ticular events at particular sites.
In an attempt to overcome some of these inadequa-cies, we have elaborated on a concept initiated by aJOIDES panel in order to develop an index of reliabilityof events that takes account of abundance of the taxonconcerned, the extent of our understanding of the evolu-tion of the taxon, the ease of distinguishing it from co-occurring forms, the preservation of the assemblagerelative to the susceptibility to solution of the taxon con-
410
RADIOLARIANS FROM THE MIDDLE AMERICA TRENCH
Table 8. Late Miocene radiolarians at Site 499.
en.CooQ.
LLJ
Φ
2 8to O
Zones
D. antepen-ultima
Site 499
(core-section.interval in cm)
23-1, 20-22
23-1, 40-42
23-1, 69-71
α>
lids
pe
r s
sand
3O
4-1
>
an D
en
sit
CO
O
CO
25.2
18.9
18.9
goo
d)
rate
,
α>^ 3O
E
ion
(p
oor
CO
0 )
α_
M
N
M
ughe
si
|."3
Q
.67
.76
2.9
ette
rsso
ni
|CO
Q
-
.06
.19
11&
1Q
.29
.70
1.4
o
yrti
s la
tic
O
ε
-.09
.63
CO
timyr
tis p
ern
1O
O
4.4
1.5
.19
a ba
cca
1
thi
.25
.25
.03
3 ne
ote
ra
1•c
-
-
.02
ten:
co
1QGO
"SCO
.95
1.0
1.4
CD
rys
pere
gr
o6
"SCO
.80
.38
.19
rys
wo
lffi
oCJ
O,O
CO
-
-
.01
Note: Symbols as for Table 1.
Table 9. Early to middle radiolarians at Site 499.
Zones
Site 499
(core-section,interval in cm) cc Q Q Q Q. Q
O D O
ε ε εQ Q Q Q Q
!° !« !C >; £ £ &11
IIIco Co ò
δ •S
+rJ •K. 4-J " S
II 1
D. alata
C. costata
S. wolffii
23-1, 102-10423,CC,10-1224-1, 16-1824-3, 84-8625-3, 100-10225-4, 102-104
18.9
12.6
25.2
25.2
18.9
12.6
.05!* - .05* ?/ ?/
- - - ?/ n2.2
.59
.25
r 1.1
r .41
.18 .63
.36 .42
- R
- R
R .02
r -
.50 R
F F
F R
R R
r R -
R R -
- r R- R R
R R R
F R R -
C F R R
C R R
C F r
F C - -
F C - -
F F + -
R C -
R C -
R R - F
Note: Symbols as for Table 1. * = probable downworking (this chalky sample had some dark in i t ) , •f• = relative abundances indeterminate, only cortical shells are preserved.
cerned, low abundances due to dilution, intermittenceor constancy of occurrence of the taxon in the sequence,the relation of the locality to the periphery of the area ofdistribution of the taxon, and the effects of reworking.Some of these factors are numerical, and all are placedarbitrarily on a numerical scale of values 0.1, 0.25, 0.5,0.75, and 1 (Table 12).
In applying this procedure here for the first time, wefind the following explanations of the factors necessary.
A) Abundance of taxσn is divided into three ranges,as indicated in Table 12. This refers not to relative abun-dance but to the absolute number of specimens of thetaxon found in the samples near the limits of the strati-graphic range. The purpose is to reflect the greater like-lihood of having found the true stratigraphic limit if
more than five specimens have been observed in eachsample.
B) Average relative abundances of ancestor and de-scendant above and below evolutionary transitions. Forexample, if the descendant is 6 times as abundant as theancestor above the event, and the ancestor is 1.5 timesas abundant as the descendant below the event, theaverage is slightly less than 4, and would score 0.75.
C) Type of event, reflecting whether or not the an-cestor or descendant of the taxon concerned is known,and the type of evolutionary interpretation. "Crypto-genetic origin" refers to the earliest occurrence of a tax-on for which there are several or no candidate ancestors.An extinction is regarded as "cryptogenetic" when thetaxon left no known evolutionary descendant, and "evo-
411
M. J. WESTBERG, W. R. RIEDEL
Table 10. Quaternary radiolarians at Site 500.
Epo
chs
Qua
tern
ary
Hole 500 andHole 500A
(core-section,interval in cm)
1-3, 4-6
2 - 1 , 50-52
3-4, 70-72
4-2, 6-8
5-2, 34-36
7-7, 90-92
8-2, 116-118
9-2, 130-132
9-4, 77_79
10-2, 21-23
A 1, CC, 11-13
A 2,CC
Rad
iola
rian
Den
sity
(tho
usan
ds p
er s
lide)
6.3
6.3
1.3
1.3
1.3
1.3
0.5
1.3
0.3
0.3
1.3
1.3
Pre
serv
atio
n (m
od
era
te,
good
)
G
G
G
G
G
G
G
M
M
M
M
M
Buc
cino
spha
era
inva
gina
ta
Col
losp
haer
a tu
bero
sa
A×
op
run
um
ang
elin
um
Did
ymocy
rtis
avi
ta
Did
ymoc
yrtis
penulti
ma
- r -
- r -
r R -
R R:
R R:
R R
R R
Did
ymocy
rtis
te
trat
haia
mus
Am
ph
irh
op
aiu
m y
psilo
n
Spo
ngas
ter p
enta
s
Spo
ngas
ter
tetr
as
Lyc
hn
od
icty
um
aud
a×
R R r
F R r
F R R
R F R
R R
R R R
F - R
F R R
F R R
F R R
R R R
R R R
Pte
roca
nium
pr
ism
atiu
m
Stic
hoco
rys
pere
grin
a
Anth
ocy
rtid
ium
ang
uiar
e
Lam
proc
yrtis
het
erop
oros
Lam
proc
yrtis
neo
hete
ropo
ros
- + '. -
Rl - .33
R.' -
R
R
Lam
proc
yrtis
ni
grin
iae
The
ocor
ythi
um
trac
heliu
m
The
ocor
ythi
um
vetu
lum
Pho
rmos
ticho
artu
s co
rbia
Spi
rocy
rtis
gy
rosc
alar
is
Spi
rocy
rtis
sc
alar
is
F .03 F r r
F .02 F - R
R - F - -
R - F - -
R - F - -
R - F - -
.20 - - F - -
.38 - - F R -
.67 - - F - -
1.0 .33 - F - -
.07 - - F - -
. . . F - -
Note: Symbols as for Table 1.
Table 11. Early to middle Miocene radiolarians at Site 500.
Epo
chs
early
Mio
cene
j m
iddl
eM
ioce
ne
Zones
D. alata
C. costata
S. woiffii
S. deimontensis
Site 500
(core-section,interval in cm)
10-2, 127-129
10-3, 127-129
11-3, 73-75
11-4, 32-34
12-3, 127-129
13-2, 50-52
13-3, 60-62
14-2, 10-12
14-4, 30-32
Rad
iola
rian
Den
sity
(tho
usan
ds
per
slid
e)
6.3
12.6
2.5
6.3
18.9
18.9
18.9
18.9
6.3
Pre
serv
atio
n (p
oor,
mod
erat
e)
P
M
M
M
M
M
M
M
M
Dia
rtus
hug
hesi
Dia
rtus
pet
ters
soni
Did
ymoc
yrtis
ant
epen
ultim
a
Did
ymoc
yrtis
lat
icon
us
Did
ymoc
yrtis
mam
mife
ra
- - - - 1.9
- - - - 1.7
1.3
.95
Did
ymoc
yrtis
pris
mat
ica
Did
ymoc
yrtis
tub
aria
Did
ymoc
yrtis
vio
iina
Dic
tyoc
oryn
e on
tong
ensi
s
Dor
cado
spyr
is a
lata
r .91 - r
r .57 - r
r .80
r r 1.1
r 4.4 8.3
r .52 .35
r .25 .25
.11 .07 -
.13 .06
Dor
cado
spyr
is d
enta
ta
Lirio
spyr
is p
arke
rae
Lirio
spyr
is s
taur
opor
a
Cyr
toca
psei
ia c
ornu
ta
Cyr
toca
psei
ia t
etra
pera
- R r r F
- R - R F
r .30 .70 F
r .02 .13 F F
- - - F C
- - - R C
F C
R F
R F
Lith
oper
a ne
oter
a
Lith
oper
a re
nzae
Lych
noca
nom
a el
onga
ta
Lych
nodi
ctyu
m a
uda×
Sitc
hoco
rys
arm
ata
- r R -
r -
R - R R
r - F
- R r F
- R R -
R
R
F
Stic
hoco
rys
deim
onte
nsis
Stic
hoco
rys
woi
ffii
Car
poca
nops
is b
ram
iette
i
Car
poca
nops
is c
ingu
iata
Car
poca
nops
is
favo
sa
C R r r
F F R r -
F F r • -
C C r R -
C C r F R
C - - R R
F - - R R
C R R
F R R
Cal
ocyc
letta
cos
tata
Cal
ocyc
letta
virg
inis
Pho
rmos
ticho
artu
s co
rbui
a
Pho
rmos
ticho
artu
s do
lioiu
m
Sip
host
icho
artu
s co
rona
F C r - -
F F - - -
R F r - r
F C -
- C -
- F r r
R -
C -
R -
Note: Symbols as for Table 1.
lutionary" when it is confidently interpreted as havingterminating its lineage. As with evolutionary extinc-tions, a score of 0.75 is assigned to morphotypic limitsthat bracket known evolutionary transitions, as well asto earliest morphotypic occurrences of an evolutionaryoffshoot from a persistent lineage, because in both ofthese cases the limit would be extended by searching
through much larger samples (Fig. 2). By "evolutionarytransition" is meant the level at which the numericaldominance changes from ancestor to descendant.
D) Ease of distinguishing the taxon from co-occur-ring fossils. The reliability of species distinctions is wellknown to vary widely, and the scoring of this factortherefore ranges from the minimum value to the max-
412

RADIOLARIANS FROM THE MIDDLE AMERICA TRENCH
Table 12. Scoring of the value of a particular biostratigraphic event at a particular locality.
______̂ MultiplierFactor " —^__^^^
A) Abundance of taxon (numbers of speci-mens observed in each sample)
B) Average relative abundances ofancestor and descendant above andbelow the event (applies only to evolu-tionary transitions)
C) Type of event
D) Ease of distinguishing the taxon fromco-occurring fossils
E) Is the assemblage well preserved,relative to the preservation threshold ofthe taxon concerned, or are samplesimmediately above or below the eventpoorly preserved or highly diluted?
F) Constancy of occurrence in thesequence, above or below the event(not applicable to evolutionarytransitions)
G) Relation of locality to periphery ofarea of distribution of taxon
H) Effects of reworking
0.10
Difficult
No
0.25
1
Near periphery
0.50
2-5
< 2 ×
Cryptogenetic originor extinction
Moderately easy
Uncertain, or somedifficulty dueto preservation
Unknown
The limit as here interpreted may be unreliabledue to reworking
0.75
2-5 ×
Evolutionary extinction, ormorphotypic limit, orevolutionary offshoot
Interrupted
Occurrences outside thisrange are certain to bedue to reworking
1.00
>5
> 5 ×
Evolutionary transition
Very easy
Yes
Constant
Well within
No reworking
Morphotypicupper limitof ancestor
Morphotypiclower limit of -an evolutionaryoffshoot
Evolutionary transition
Morphotypic lower limitof descendant
MORPHOLOGY
Figure 2. Diagram illustrating the use of terms describing stratigraphicevents associated with lineages and evolutionary offshoots.
imum. This is a less objective factor than some otherslisted.
E) Assemblage preservation relative to the preserva-tion threshold of the taxon concerned. This is clearly amore pertinent factor than an evaluation of the overallpreservation of the assemblage. A median score is givenwhen one is uncertain whether an absence or low abun-dance is due to solution or when solution has removedfeatures necessary for identification from some speci-mens in the sample.
This category is also used to downgrade the score ofan upper morphotypic limit above which (or a lowermorphotypic limit below which) absence of the taxon issuspected to be due to dilution or dissolution.
F) Constancy of occurrence in the sequence, withinthe stratigraphic range of the taxon. This is not appli-cable to evolutionary transitions because these are lo-cated by the changing proportions of ancestor and de-scendant, not by the limit of range of a morphotype.The need to consider intermittence of occurrence is clear-ly demonstrated by the highly variable abundances ofsome rather common species throughout their ranges, asdescribed by Westberg and Riedel (1978).
G) Relation of locality to area of distribution of thetaxon. It seems obvious that the earliest and latest rec-ords of a taxon near the periphery of its area of distribu-tion will be less reliable stratigraphically than those wellwithin the boundaries. Also, for localities whose rela-tion to biogeographic distributions have changed throughtime (because of seafloor spreading or changes in oce-anic conditions), this factor may be evaluated differ-ently for the upper and lower limits of the same taxon.
H) Effects of reworking. The index of reliability ofan event is downgraded slightly if the investigator isconfident of having recognized that an extension ofstratigraphic range was due to reworking, and is down-graded more if he is less confident. This downgrading isdone on the assumption that the reworking is taken intoconsideration in interpreting the upper or lower limit,and the reworked occurrences are excluded from therange. This is clearly an area of subjective judgment, inwhich the investigation must take account of sedimento-logical evidences of disturbance, the abruptness of ap-pearances and disappearances of the taxon in other se-quences, evidence of reworking of other taxa into theassemblage, and so on.
As an example of the application of this index, theupper limit of Anthocyrtidium angulare scored 0.5 oneach factor A, C, D, E, and G, 0.75 on factor F, and
413

M. J. WESTBERG, W. R. RIEDEL
1.0 on factor H. Multiplying these together yields 0.02,which converts to 2°7o (the first entry for Site 495 inTable 13).
In designing this index of stratigraphic reliability, wehave attempted to formulate a procedure that will be ap-plicable not only to radiolarians but to all microfossilgroups. Thus it should be as useful for resolving con-flicts between various groups used for stratigraphic cor-relation, as it is for the various taxa of a single group(see the following section and Riedel and Westberg, inpress).
Correlations
Figure 3 shows the correlations among all of the siteson this Leg. Most of the correlation lines present noproblem, but there are two conflicts that the reliabilityindex scores shown in Table 13 help to resolve.
The order of the evolutionary transition from Didy-mocyrtis violina to D. mammifera, and the morpho-typic top of the Carpocanopsis favosa (Events 37 and38) is reversed at Site 499 as compared to Sites 495 and500, and although the reliability index for Event 37 isequally low at all three sites, the index for Event 38 atSite 500 is higher than that at Site 499, indicating thatthe order at Site 500 is more likely correct.
The morphotypic top of Carpocanopsis cingulata(Event 36) at Site 500 occurs above the evolutionarytransition from Dorcadospyris dentata to D. alata (Event34); however, the reliability index for Event 36 at thatsite is lower than the indices at Sites 495, 499, and 496.Therefore the sequence at these sites is considered thecorrect one.
Relative Abundances of Families
Because Sites 494 and 496 are located on the Guate-malan margin and Site 495 is on the Cocos Plate, it is ofinterest to investigate whether the radiolarian assem-blages on the continental side of the Trench differ sig-nificantly from those on the oceanic side.
The radiolarian species treated in the main body ofthis paper were selected (from the much larger totalpresent) because of their known stratigraphic useful-ness. It seems appropriate, however, to approach thegeographic-environmental question by examining dif-ferences in the assemblages at the family level, becausethis provides a practical means of recording the entireassemblages.
We selected approximately time-equivalent portionsof the sequences at the three sites (S. pentas Zonethrough Quaternary), and assigned 300 specimens fromeach sample to 14 family categories and the suborderSpyrida. Because we were working at the same time onmaterial from DSDP Leg 68, we gathered the same dataon samples from Hole 503A (4°03'N, 95°38'W) for anadditional oceanic reference site. Relative percentagesare shown in Tables 14 through 17, together with indica-tions of the preservations and the degree of dilution byvolcanic ash, diatoms, or other terrigenous materials.
We also compared a data set from a previous publica-tion (Riedel and Sanfilippo, 1978a) as a third exampleof a non-open-ocean environment. These data are con-
stant-numerator percentage estimates of the same 15family categories in samples from the Pliocene CapoRossello section in southern Sicily.
These constant-numerator percentage estimates showmore details of the contributions of rare taxa to theassemblages but are too time-consuming to apply rou-tinely in preliminary studies such as most of those forDSDP Initial Reports.
After eliminating two families whose abundanceswere almost always less than 1%, the data from eachsite were transformed to an M × M matrix of correla-tion coefficients between pairs of families, as a first stepin the cluster analysis program of Davis (1973, p. 467).The dendrograms of the margin sites show a cluster offour families, spyrids, plagoniids, theoperids, and pter-ocorythids (Fig. 4). The same families are clustered inthe samples from the Sicilian Pliocene, with the additionof cannobotryids and artostrobiids, but no such struc-ture is seen in the dendrograms of the oceanic sites.
Table 18 shows correlation coefficients for some fam-ily pairs in all of the sites examined. Nine of the four-teen pairs (rows 1-9, Table 18) have high positive ornegative correlation in the margin sites and at CapoRossello, but insignificant correlations at the open oceansites. Three pairs (rows 10-12, Table 18) correlate sig-nificantly at the margin sites, Capo Rossello, and at Site495, but not at Site 503. Two family pairs (rows 13 and14, Table 18) have significant negative correlations at allsites investigated.
These findings represent a substantial advance be-yond the tentative indications of covariances resultingfrom previous work on the Capo Rossello assemblages(Riedel and Sanfilippo, 1978a) and augur well for thepossibility of using the relative proportions of high-lev-el radiolarian taxa in order to recognize restrictedpaleoenvironments.
SPECIES LIST
The purpose of this list is to provide bibliographic references to thetaxa mentioned in this chapter. The only literature references given areto the original description, and to our present concept of the species, ifdifferent from or more detailed than the original one.
Alievium supβrbum (Squinabol)Theodiscus superbus Squinabol, 1914, p. 271, pi. XX(I), fig. 4.Alievium superbus (Squinabol), Pessagno, 1972, p. 302, pi. 24,figs. 5, 6, pi. 25, fig. 1.
Amphipyndax stocki (Campbell and Clark)StichocapsaiP.) stocki Campbell and Clark, 1944b, p. 44, pi. 8,figs. 31-33.Amphipyndaxstocki (Campbell and Clark), Foreman, 1968, p. 78,pi. 8, figs. 12a-c.
Amphirhopalum ypsilon HaeckelAmphirhopalum ypsilon Haeckel, 1887, p. 522; Nigrini, 1971, p.447, pi. 34.1, figs. 7a-c.
Anthocyrtidium angulare NigriniAnthocyrtidium angulare Nigrini, 1971, p. 445, pi. 34.1, figs.3a, b.
Artophormis gracilis RiedelArtophormis gracilis Riedel, 1959, p. 300, pi. 2, figs. 12, 13.
Axoprunum angelinum (Campbell and Clark)Stylosphaera angelina Campbell and Clark, 1944a, p. 12, pi. 1,figs. 14-20.Stylatractus universus Hays, 1970, p. 215, pi. 1, figs. 1, 2.Axoprunum angelinum (Campbell and Clark), Kling, 1973, p. 634,pi. 1, figs. 13-16, pi. 6, figs. 14-18.
414
RADIOLARIANS FROM THE MIDDLE AMERICA TRENCH
Table 13. Radiolarian events at Leg 67 sites.
Zones
C. tuberosa
A. ypsilon
A. angulare
P.prismatium
S. pent as
S. peregrina
D.penultima
D. ante-penultima
D.petterssoni
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
Events
Bm C. tuberosa
Tm A. angelinum
Tm A. angulare
Tm L. neoheteroporos
T. vetulum —T. trachelium
Bm A. angulare
Tm P. prismatium
Tm S. peregrina
D. avita —D. tetrathalamus
Tm. L. audax
Tm P. doliolum
Bm P. prismatium
D. penultima —D. avita
S. pent as —S. tetras
Tm S. ambus
S. berminghami —S. pentas
Tm C. robusta grp.
Bm S. ambus
S. delmontensis -S. peregrina
Tm D. hughesi
Tm D. ontongensis
D. petterssoni —D. hughesi
Bm D. hughesi
Bm D. ontongensis
495
—
3-6, 20-224-1, 75-77
24-1, 75-774-3, 75-77
145-2, 40-425-5, 40-42
256-4, 10-126,CC (3-4)
27-1, 70-727-3, 70-72
99-3, 75-779-6, 26-28
756-1, 92-9411-1, 60-62
311-1, 60-6211-3, 60-62
3711-3, 60-62ll,CC(4-6)
1911-3, 60-62ll,CC(4-6)
1411-3,60-6212-3, 30-32
412-7, 31-3313-5, 54-55
1812-7, 31-3313-5, 54-55
y15-5, 30-3216-2, 30-32
3815-5, 30-3216-2, 30-32
5015-5, 30-3216-2, 30-32
Q
y15-1, 30-3216-2, 30-32
3716-6, 30-3217-1, 30-32
7517-6, 30-3218-2, 90-92
5017-6, 30-3218-2, 90-92
2518-6, 101-10219-1, 119-121
3818-6, 101-10219-1, 119-121
25
2-2,3-4,
14-315-3
17-518-1
17-518-1
16-121-1
19-319-6
499
109-111144 146
7, 37-39, 102-103
75, 58-60, 100-102
6, 58-60, 100-102
7,0-3, 45-47
38, 110-112, 10-12
6
Holes
500 494
3-4, 70-72 1-5, 59-614-2, 6 8 1-6, 59 61
59-2, 130-132 A2-6,9-4,77-79 A4-1,
7510-2, 21-23Al.CC (11-13)
3
A17-2A18-1
A18-1A20-2
729-311-375—
—
—
—
—
, 64-66, 90-92
7_
—
—
—
, 90-92, 110-11212
497
1-3, 80-822-3, 80-82
127-2, 80-829-4, 23-25
7513-4, 14-1615-4, 70-72
516-4, 30-3217-3, 26-28
1415-4, 70-7217-3, 26-28
618-5, 10-1219-2, 120-122
5—
25-4, 40-4226-3, 16-18
7525-4, 40-4226-3, 16-18
3—
33,CC (14-16)34-3, 24-26
56—
35-2, 41-4336-8, 35-37
934-3, 24-2635-2, 41-43
9—
496
1-1, 50-521-5, 50-52
1915-4, 110-11216-1, 30-32
7521-6, 86-8822-4, 65-67
3—
23-6, 30-3224-3, 121-126
923-6, 30-3224-3, 121-126
3
26-4, 5-727-4, 29-31
7525-4, 40-4227-4, 29-31
326-4, 5-727-4, 29-31
5026-4, 5-727-4, 29-31
19—
—
—
415
M. J. WESTBERG, W. R. RIEDEL
Table 13. (Continued).
Zones
D.petterssoni
D. alata
C. costata
S. wolffii
S. delmont-ensis
Events
25 Bm P. doliolum
26 Tm C. bramlettei
27 Bm D. petterssoni
28 Tm D. alata
29 Tm C. cornuta
30 Tm L. renzae
31 L. renzae —L. neotera
32 Bm P. corbula
33 Tm C. costata
34 D. dentata —D. alata
35 L. stauropora -~L. parkerae
36 Tm C. cingulata
37 D. violina —D. mammifera
38 Tm C. favosa
39 Tm L. elongata
40 Bm C. costata
41 Bm D. dentata
42 £>. tubaria —Zλ violina
43 Bm L. stauropora
44 Bm 5. wo/#i7
45 Zλ prismatka —D. tubarius
495
19-1, 119-12119-4, 8-10
319-1, 119-12110-4 fl-if)l7"*T, O I v
3819-6, 8-1020-1, 60-62
7519-6, 8-1020-1, 60-62
3819-6, 8-1020-1, 60-62
5019-6, 8-1020-1, 60-62
3819-6, 8-1020-1, 60-62
5020-6, 60-6221-2, 70-72
3822-5, 30-3223-2, 41-43
3823-6, 41-4324-1, 15-17
5023-6, 41-4324-1, 15-17
5024-1, 15-1724-3, 40-42
5024-1, 15-1724-3, 40-42
524-7, 40-4225-1, 30-32
1926-3, 30-3226-5, 30-32
1929-5, 19-2130-1, 30-32
7529-5, 19-2130-1, 30-32
3829-5, 19-2130-1, 30-32
3830-2, 30-3231-2, 30-32
1333-5, 30-3234-2, 26-28
56—
499
23.CC (10-12)24-1, 16-18
10023,CC (10-12)24-1, 16-18
10024-1, 16-1824-3, 84-86
5024-3, 84-8625-3, 100-102
524-1, 16-1824-3, 84-86
2524-3, 84-8625-3, 100-102
2524-3, 84-8625-3, 100-102
50B3-1.0-1B3-4, 48-50
75B3-1, 0-1B3-4, 48-50
38B3-1, 0-1B3-4, 48-50
75B3-4, 48-50B4-1, 67-69
75
Holes
500 494
10-3, 127-12911-3, 73-75
5010-3, 127-12911-3, 73-75
10010-2, 21-2310-2, 127-129
2511-3,73-7511-4, 32-34
511-4, 32-3412-3, 127-129
5011-4, 32-3412-3, 127-129
5011-4, 32-3412-3, 127-129
5011-4, 32-3412-3, 127-129
3812-3, 127-12913-2, 50-52
3811-4, 32-3412-3, 127-129
1912-3, 127-12913-2, 50-52
7513-3, 60-6214-2, 10-12
50
497 496
29-3, 40-4230-3, 45-47
7529-3, 40-4230-3, 45-47
9—
30-3, 45-4731-3, 12-14
730-3, 45-4731-3, 12-14
1330-3, 45-4731-3, 12-14
2834-5, 20-2235-2, 60-62
2530-3, 45-4735-2, 60-62
13—
30-3, 45-4731-3, 12-14
50—
31-3, 12-1433-5, 9-11
25
Note: Presented are core-section and intervals in cm of the samples bracketing the event. The number below the sample numbers indicates the index ofreliability (see text). Tm and Bm indicate the morphotypic top or bottom of a species; arrows indicate evolutionary transitions.
416

RADIOLARIANS FROM THE MIDDLE AMERICA TRENCH
Epoch Zone
C. tu-beroseA. yp•silonA. an-
oulare
P. pris•matium
Events495 499 500
Holes
494 497 496
,Bm C. tuberosa 1
Tm A. angelinum 2
Tm A. angulare 3 ,
T m L neoheteroporos 4,
T. vetulum —*-T. trachelium
Bm A. angulare
~Tm P. prismatium
.TmS. peregrina
D. avita — * -D. tetrathalamus
Tm L. audax
Tm A. doliolum
Bm P. prismatium
D. penultima — * •D. avita
S. pentas—*~S. tetras
Tm S. ambus
S. berminghami—S. pentas
Tm C. robusta grp.
Bm S. ambus
- S. delmontensis — *S. peregrina
. hughesi
Tm D. ontongensis
s D. petterssoni — » -D. hughesi
Bm D. hughesi
BmD, ontongensis
Bm P. doliolum
Tm C. bramlettei
Bm D. petterssoni
Tm D. alata
Tm C. cornuta
Tm L. renzae
L. renzae — * -L. neotera
Bm P. corbula
Tm C. costata
^ D. dentata — * -D. alata
L. stauropora — » -L. parkerae
Tm C. cingulata
D. violina •-
D. mammifera
Tm C. favosa
Tm L. elongate
Bm C. costata
BmD. dentata
D. tubaria *•
D. violina
Bm L. stauropora
Bm S. wolffii
43
44
25
100
125
150 ?
175 I
200
225
250
275
300
325
350
Figure 3. Correlation of radiolarian events in Leg 67 sites. (Each rectangular box represents a core with its numbershown inside. Numbers labeling correlation lines correspond to the radiolarian events listed on the left. Tm in-dicates the morphotypic top of a species and Bm the morphotypic bottom of a species; an arrow denotes evolu-tionary transitions. Sample intervals bracketing events, and a reliability index for each event, are given in Table13.)
417

M. J. WESTBERG, W. R. RIEDEL
Table 14. Percent estimate of radiolarian families and the suborder Spyrida at Site 494.
Site 494
(core-section.interval in cm)
1-1, 32-34
2-3, 32-34
3-2, 32-34
4-4, 32-34
A
A
A
A
A
A
A
A
A
A
A
A
A
A
A
A
2-6, 29-31
4-1, 1-3
5-1, 60-62
7-3, 60-62
8-1, 70-72
9-1, 20-24
10-1, 133-135
11-1, 70-72
14-3, 12-14
16-1, 87-91
17-2, 64-68
18-1, 90-92
18-4, 64-66
19-1, 7-9
20-2, 110-112
20-3, 76-78
1Io§3
1
3
2
5
2
1
1
1
1
1
1
1
0
0
0
2
2
0
0
1εso
5
10
6
8
6
8
9
3
6
7
21
10
4
9
8
12
10
13
6
15
oto
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1§a5
1
1
2
1
1
0
1
0
1
0
2
0
0
0
0
1
0
0
1
1I&33
36
33
35
52
47
36
41
46
52
56
42
31
59
47
52
42
42
62
65
1C
8
5
8
14
13
12
9
9
11
9
9
15
11
7
13
10
9
11
15
4
1£.o0fS
0
0
0
0
0
0
0
0
0
0
0
1
0
0
0
0
0
0
0
0
1
11
12
11
11
11
11
7
5
13
11
5
11
13
9
16
16
9
8
14
10
«
5
3
5
4
4
5
3
4
3
1
0
2
6
0
0
1
2
2
0
0
I1Q ,
5
6
4
2
2
2
6
6
5
3
0
4
7
1
1
0
4
3
0
0
11110
9
10
10
3
5
13
13
5
3
1
4
8
4
1
1
6
6
1
1
cu
11s•<3
l
l
l
i
0
i
3
2
3
2
1
1
1
2
2
1
2
1
0
0
8
19
8
9
2
2
1
3
8
4
3
2
2
7
1
2
0
3
3
0
0
1
|6
5
9
8
2
4
8
6
2
4
5
4
10
7
9
5
10
8
2
3
o
0
1
1
0
1
0
2
2
1
1
0
2
1
0
1
0
1
1
1
1
gcα>
£
M
M
M
M
M
P
P
P
P
P
P
P
M
M
M
M
M
M
M
M
CΛ
Eou.αi
CO
o
Q
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
IOu_co
m
r
r
r
r
R
R
R
R
R
R
R
R
R
R
R
R
F
R
R
R
co
Q
50X
60A
50X
-
75X
75X
75X
50X
80X
90X
99X
95X
60DX
50X
95A
50DA
99X
99X
95X
95X
Note: In the Preservation column G indicates good, M moderate and P poor. In the next two columns,— indicates none, r, 0.01—0.1%, and R, 0.1 — 1%. The Dilution column indicates the percentage of non-radiolarian components in an average field: D for diatoms, A for volcanic ash, and X for other terrigenousmaterial.
Buccinosphaera invaginata HaeckelBuccinosphaera invaginata Haeckel, 1887, p. 99, pi. 5, fig. 11;Strelkov and Reshetnyak, 1962, p. 129 and 137, fig. 12; Nigrini,1971, p. 445, pi. 34.1, fig. 2; Knoll and Johnson, 1975, p. 63, pi. 1,figs. 3-6.
Calocyclas hispida (Ehrenberg)Anthocyrtis hispida Ehrenberg, 1873, p. 216.Calocyclas hispida (Ehrenberg), Riedel and Sanfilippo, 1978b, p.65, pi. 3, fig. 6.
Calocycletta costàta (Riedel)Calocyclas costata Riedel, 1959, p. 296, pi. 2, fig. 9; Riedel andSanfilippo, 1978b, p. 66, pi. 3, fig. 9.
Calocycletta robusta Moore groupCalocycletta robusta Moore, 1971, p. 743, pi. 10, figs. 5, 6.Calocycletta caepa Moore, 1972, p. 150, pi. 2, figs. 4-7.Calocycletta robusta Moore group Riedel and Sanfilippo, 1978b,p. 66, pi. 3, figs. 10, 11.
Calocycletta virginis HaeckelCalocyclas {Calocycletta) virginis Haeckel, 1887, p. 1381, pi. 74,fig. 4.Calocycletta virginis (Haeckel), Riedel and Sanfilippo, 1978b, p.66, pi. 3, figs. 13, 14.
Carpocanopsis bramlettei Riedel and SanfilippoCarpocanopsis bramlettei Riedel and Sanfilippo, 1971, p. 1597, pi.2G, figs. 8-14, pi. 8, fig. 7; 1978b, p. 67, pi. 4, fig. 6.
Carpocanopsis cingulata Riedel and SanfilippoCarpocanopsis cingulata Riedel and Sanfilippo, 1971, p. 1597, pi.2G, figs. 17-21, pi. 8, fig. 8; 1978b, p. 67, pi. 4, fig. 4.
Carpocanopsis favosa (Haeckel)Cycladophora favosa Haeckel, 1887, p. 1380, pi. 62, figs. 5, 6.Carpocanopsis favosa (Haeckel), Sanfilippo and Riedel, 1973, p.531.
Collosphaera tuberosa HaeckelCollosphaera tuberosa Haeckel, 1887, p. 97; Nigrini, 1971, p. 445,pi. 34.1, fig. 1; Strelkov and Reshetnyak, 1971, p. 336-337, pi. 4,figs. 24, 25; Knoll and Johnson, 1975, p. 63, pi. 2, figs. 1-3.
Cyrtocapsella cornuta HaeckeiCyrtocapsa {Cyrtocapsella) cornuta Haeckel, 1887, p. 1513, pi. 78;fig. 9.Cyrtocapsella cornuta Haeckel, Riedel and Sanfilippo, 1978b, p.68, pi. 4, fig. 17.
Cyrtocapsella tetrapera HaeckelCyrtocapsa {Cyrtocapsella) tetrapera Haeckel, 1887, p. 1512, pi.78, fig. 5.Cyrtocapsella tetrapera Haeckel, Riedel and Sanfilippo, 1978b, p.68, pi. 4, fig. 18.
Diartus hughesi (Campbell and Clark)Ommatocampe hughesi Campbell and Clark, 1944a, p. 23, pi. 3,fig. 12.Ommatartus hughesi (Campbell and Clark), Riedel and San-filippo, 1978b, p. 71, pi. 7, fig. 7.Diartus hughesi (Campbell and Clark), Sanfilippo and Riedel,1980, p. 1010, text-fig. 1, i.
Diartus petterssoni (Riedel and Sanfilippo)Cannartus{1) petterssoni Riedel and Sanfilippo, 1970, p. 520, pi.14, fig. 3; 1978b, p . 67, pi. 4, fig. 2.
418

RADIOLARIANS FROM THE MIDDLE AMERICA TRENCH
Table 15. Percent estimates of radiolarian families and the suborder Spyrida at Site 496.
Site 496
(core-section,interval in cm)
1-1,
3-4,
5-6,
7-2,
9-1,
11-2,
13-1,
14-2,
15-4,
16-1,
19-4,
21-6,
22-4,
23-6,
24-3,
25-4,
26-4,
27-4,
28-3,
50-52
60-62
117-119
50-52
99-101
60-62
6-8
6-8
110-112
30-32
41-42
86-88
65-67
30-32
124-126
40-42
5-7
29-31
90-92
<llo
spha
erid
ae
<S
7
1
0
3
0
2
6
3
3
6
2
0
1
1
1
3
7
2
4
^.tin
omm
idae
8
7
5
6
5
5
13
16
7
6
11
8
10
11
16
24
48
17
17
11
0
0
0
0
0
0
0
0
0
0
0
1
0
0
0
0
0
1
0
>cco
disc
idae
ö5
3
1
3
0
1
0
0
3
1
1
3
1
1
0
0
4
3
3
G
1£22
18
37
33
20
35
25
45
46
29
34
50
50
43
48
48
19
44
18
'loni
idae
<c8
6
7
18
14
13
7
4
10
11
8
8
8
6
3
7
5
5
1
<olo
niid
ae
fc0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
0
1
thel
iidae
•Z
12
20
16
11
14
12
11
11
16
12
10
15
12
10
9
6
11
14
>yrid
a
5
7
3
5
7
6
6
1
2
7
6
3
2
3
3
1
1
1
3
agon
iidae
S:
7
7
4
L.
1
4
5
0
1
6
5
1
1
2
1
1
0
0
6
iβo
pe
rid
ae
9
10
6
5
12
7
8
4
3
7
10
4
3
6
4
L.
4
8
14
'rp
oca
niid
ae
<3
i
0
0
2
1
0
0
1
0
1
0
2
0
1
1
1
0
1
0
tero
co
ry th
ida
e
12
12
o
9
8
6
5
6
4
7
3
3
1
3
3
1
2
4
4-t
ost
rob
iida
e
7
9
11
4
11
7
13
8
6
6
7
6
4
11
9
4
4
5
14
inn
ob
otr
yid
ae
Q
0
0
1
0
1
2
0
0
0
1
1
1
3
0
0
0
0
0
0
ese
rva
tio
n
G
G
G
M
M
M
M
M
M
M
M
M
M
M
M
M
P
M
P
jlic
ate
Fo
rms
α
-------------------
oke
n F
orm
s
CO
r
r
r
r
r
r
r
r
r
r
r
r
r
r
r
r
R
r
R
iluti
on
Q
80X
60XD
50X
50X
50X
SOX
50X
50X
70X
40X
70X
80X
90X
90X
90X
95X
99X
99X
85X
Note: See Table 14 for explanations of symbols.
Diαrtuspetterssoni (Riedel and Sanfilippo), Sanfilippo and Riedel,1980, p. 1010, text-fig. 1, h.
Dictyocoryne ontongensis Riedel and SanfilippoDictyocoryne ontongensis Riedel and Sanfilippo, 1971, p. 1588,pi. IE, figs. 1, 2, pi. 4, figs. 9-11; 1978b, p. 68, pi. 5, fig. 1.
Dictyomitrα koslovαe ForemanDictyomitrα koslovαe Foreman, 1975, p. 614, pi. 7, fig. 4.
Dictyoprorα αmphorα (Haeckel) groupDictyocephαlus αmphorα Haeckel, 1887, p. 1305, pi. 62, fig. 4.Dictyoprorα αmphorα (Haeckel) group Nigrini, 1977, p. 250, pi. 4,figs. 1, 2.
Dictyoprorα mongolfieri (Ehrenberg)Eucyrtidium mongolfieri Ehrenberg, 1854, pi. 36, fig. 18, B lower.Dictyoprorα mongolfieri (Ehrenberg), Nigrini, 1977, p. 250, pi. 4,fig. 7.
Dictyoprorα urceolus (Haeckel)Dictyocephαlus urceolus Haeckel, 1887, p. 1305.Dictyoprorα urceolus (Haeckel), Nigrini, 1977, p. 251, pi. 4, figs.9, 10.
Didymocyrtis antepenultima (Riedel and Sanfilippo)Ommatartus antepenultimus Riedel and Sanfilippo, 1970, p. 521,pi. 14, fig. 4; Westberg and Riedel, 1978, p. 22, pi. 2, figs. 4, 5.Didymocyrtis antepenultima (Riedel and Sanfilippo), Sanfilippoand Riedel, 1980, p. 1010.
Didymocyrtis avita (Riedel)Panartus avitus Riedel, 1953, p. 808, pi. 84, fig. 7.Didymocyrtis avita (Riedel), Sanfilippo and Riedel, 1980, p. 1010.
Didymocyrtis laticonus (Riedel)
Cannartus laticonus Riedel, 1959, p. 291, pi. 1, fig. 5; Westbergand Riedel, 1978, p. 20, pi. 2, figs. 1-3.Didymocyrtis laticonus (Riedel), Sanfilippo and Riedel, 1980, p.1010, text-fig. 1, e.
Didymocyrtis mammifera (Haeckel)Cannartidium mammiferum Haeckel, 1887, p. 375, pi. 39, fig. 16.
Cannartus mammiferus (Haeckel), Riedel, 1959, p. 291, pi. 1,
fig. 4.Didymocyrtis mammifera (Haeckel), Sanfilippo and Riedel, 1980,p. 1010.
Didymocyrtis penultima (Riedel)Panarium penultimum Riedel, 1957, p. 76, pi. 1, fig. 1.Ommatartus penuttimus (Riedel), Westberg and Riedel, 1978, p.22, pi. 2, figs. 6-8.Didymocyrtis penultima (Riedel), Sanfilippo and Riedel, 1980, p.1010, text-fig. 1, f.
Didymocyrtis prismatica (Haeckel)Pipettella prismatica Haeckel, 1887, p. 305, pi. 39, fig. 6.Didymocyrtis prismatica (Haeckel), Sanfilippo and Riedel, 1980,p. 1010, text-fig. 1, c.
Didymocyrtis tetrathalamus (Haeckel)Panartus tetrathalamus Haeckel, 1887, p. 378, pi. 40, fig. 3.Didymocyrtis tetrathalamus (Haeckel), Sanfilippo and Riedel,1980, p. 1010, text-fig. 1, g.
Didymocyrtis tubaria (Haeckel)Pipettaria tubaria Haeekel, 1887, p. 339, pi. 39, fig. 15; Riedel,1959, p. 289, pi. 1, fig. 2.Didymocyrtis tubaria (Haeckel), Sanfilippo and Riedel, 1980, p.1010.
Didymocyrtis violina (Haeckel)Cannartus violina Haeckel, 1887, p. 358, pi. 39, fig. 10; Riedel,1959, p. 290, pi. 1, fig. 3.Didymocyrtis violina (Haeckel), Sanfilippo and Riedel, 1980, p.1010, text-fig. 1, d.
Dorcadospyris alata (Riedel)Brachiospyris alata Riedel, 1959, p. 293, pi. 1, figs. 11, 12.Dorcadospyris alata (Riedel), Riedel and Sanfilippo, 1978b, p. 68,pi. 5, fig. 2.
Dorcadospyris ateuchus (Ehrenberg)Ceratospyris ateuchus Ehrenberg, 1873, p. 218.
419

M. J. WESTBERG, W. R. RIEDEL
Table 16. Percent estimates of radiolarian families and the suborder Spyrida at Site 495.
Site 495
(core-section.interval in cm)
3-1,
3-3,
4 -1 ,
4-3,
5-2,
5-5,
6-4,
7-1,
7-3,
8-1,
9-3,
9-6,
10-4,
10-6,
11-3,
12-1,
12-5,
13-5,
14-3,
14-5,
15-3,
15-5,
16-4,
1-3
60-62
75-77
75-77
40-42
40-42
10-12
70-72
70-72
16-18
75-77
26-28
30-32
30-32
60-62
30-32
30-32
54-55
30-32
30-32
30-32
30-32
30-32
llosp
haer
idae
2
5
4
2
1
0
8
1
1
0
5
2
0
0
3
0
0
2
6
2
1
0
8
-.tin
omm
idae
-̂
15
10
10
6
12
13
29
17
26
17
15
11
19
22
25
29
32
26
18
18
20
19
10
^aco
disc
idae
*-
0
0
0
0
0
0
0
3
0
0
0
0
0
0
1
0
0
0
0
1
1
1
2
ccod
isci
dae
<35
2
0
2
2
1
1
1
0
0
1
1
1
1
2
5
1
3
1
3
3
2
6
ongo
disc
idae
%
33
26
35
34
37
55
33
23
34
40
27
27
30
24
32
16
27
29
31
25
30
21
18
'loni
idae
<C8
8
11
4
7
4
4
10
8
7
6
6
10
9
9
8
4
6
3
6
7
4
4
<olo
niid
ae
fc0
0
1
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
{hel
iidae
6
7
5
10
11
8
4
7
5
7
6
9
6
14
7
11
5
4
4
9
10
14
11
<yrid
a
6
5
5
7
6
3
2
7
4
5
6
8
3
2
5
1
1
3
4
8
3
2
7
agon
iidae
S:
5
3
8
10
5
3
1
5
1
1
3
5
1
3
1
1
0
1
3
3
1
3
6
1Iß5
7
10
8
6
4
5
6
6
4
8
7
8
8
6
9
7
8
8
8
16
14
15
rpoc
aniid
ae
a2
1
2
2
2
1
1
3
1
3
2
3
1
1
1
0
0
0
0
0
0
1
0
ero
cory
tbid
ae
7
14
4
6
1
3
3
4
3
4
9
4
5
2
3
5
3
6
5
4
4
4
3
tost
robi
idae
**>
5
12
4
7
8
6
8
13
10
11
8
15
14
13
6
15
19
11
16
13
4
16
9
'nno
botr
yida
e
c3l
1
l
2
1
0
1
1
0
2
5
2
1
0
1
0
0
0
0
0
0
0
1es
erva
tion
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
Mili
cate
For
ms
IDQ
-
-
r
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
oke
n F
orm
s
m
R
r
r
r
r
R
R
R
r
R
R
R
R
R
R
R
r
R
R
r
r
r
r
lutio
n
b50X
-
-
25D
50X
50X
-
25X
-
-
-
90A
-
-
SOX
25A
-
-
50DX
-
-
-
-
Note: See Table 14 for explanations of symbols.
Dorcαdospyris αteuchus (Ehrenberg), Riedel and Sanfilippo,1978b, p. 68, pi. 5, fig. 3.
Dorcαdospyris dentαtα HaeckelDorcαdospyris dentαtα Haeckel, 1887, p. 1040, pi. 85, fig. 6;Riedel and Sanfilippo, 1978b, p. 68, pi. 5, fig. 4.
Eucyrtidium diαphαnes Sanfilippo and RiedelEucyrtidium diαphαnes Sanfilippo and Riedel, in Sanfilippo et al.,1973, p. 221, pi. 5, figs. 12-14.
Eucyrtis microporα (Squinabol)Archicαpsα microporα Squinabol, 1903, p. 129, pi. 9, fig. 14.Eucyrtis microporα (Squinabol), Foreman, 1975, p. 615, pi. 21,figs. 2-5.
Lαmprocyrtis heteroporos (Hays)Lαmprocyclαs heteroporos Hays, 1965, p. 179, pi. 3, fig. 1.Lαmprocyrtis heteroporos (Hays), Kling, 1973, p. 639, pi. 5, figs.19-21, pi. 15, fig. 6.
Lαmprocyrtis neoheteroporos KlingLαmprocyrtis neoheteroporos Kling, 1973, p. 639, pi. 5, figs. 17,18, pi. 15, figs. 4, 5.
Lαmprocyrtis nigriniαe (Caulet)Conαrαchnium nigriniαe Caulet, 1971, p. 3, pi. 3, figs. 1-4, pi. 4,figs. 1-4.Lαmprocyrtis hαysi Kling, 1973, p. 639, pi. 5, figs. 15, 16, pi. 15,figs. 1-3.Lαmprocyrtis nigriniαe (Caulet), Kling, 1977, p. 217, pi. 1, fig. 17.
Eusyringium fistuligerum (Ehrenberg)Eucyrtidium fistuligerum Ehrenberg, 1873, p. 229.Eusyringium fistuligerum (Ehrenberg), Riedel and Sanfilippo,1978b, p. 68, pi. 5, figs. 6, 7.
Liriospyhs pαrkerαe Riedel and SanfilippoLiriospyris pαrkerαe Riedel and Sanfilippo, 1971, p. 1590, pi. 2C,fig. 15, pi. 5, fig. 4.
Liriospyris stαuroporα (Haeckel)Trissocyclus stαuroporus Haeckel, 1887, p. 987, pi. 83, fig. 5.Liriospyris stαuroporα (Haeckel), Riedel and Sanfilippo, 1971, p.1590, pi. 2C, figs. 16-19.
Lithochytris vespertilio EhrenbergLithochytris vespertilio Ehrenberg, 1873, p. 239; Riedel and San-filippo, 1978b, p. 69, pi. 6, fig. 4.
Lithocyclia ocellus Ehrenberg groupLithocyclia ocellus Ehrenberg, 1854, pi. 36, fig. 30.Lithocyclia ocellus Ehrenberg group Riedel and Sanfilippo, 1978b,p. 70, pi. 6, fig. 8.
Lithopera bacca EhrenbergLithopera bacca Ehrenberg, 1872, p. 314; Riedei and Sanfilippo,1978b, p. 70, pi. 6, fig. 9.
Lithopera neotera Sanfilippo and RiedelLithopera neotera Sanfilippo and Riedel, 1970, p. 454, pi. 1, figs.24-26, 28; Riedel and Sanfilippo, 1978b, p. 70, pi. 6, fig. 10.
Lithopera renzae Sanfilippo and RiedelLithopera renzae Sanfilippo and Riedel, 1970, p. 454, pi. 1, figs.21-23, 27; Riedel and Sanfilippo, 1978b, p. 70, pi. 6, fig. 11.
Lophocyrtis biaurita (Ehrenberg)Eucyrtidium biauritum Ehrenberg, 1873, p. 226.Lophocyrtis biaurita (Ehrenberg), Haeckel, 1887, p. 1411; Cita etal., 1970, p. 404, pi. 2, I, J, K.
Lychnocanoma elongata (Vinassa)Tetrahedrina elongata Vinassa, 1900, p. 243, pi. 2, fig. 31.
420

RADIOLARIANS FROM THE MIDDLE AMERICA TRENCH
Table 17. Percent estimates of radiolarian families and the suborder Spyrida at Site 500.
Hole 503A
(core-section,interval in cm)
1-1, 50-54
4-1, 50-54
5-2, 50-54
7-1, 50-54
9-1, 50-54
10-1, 50-54
12-3, 50-54
15-2, 66-70
19-2, 60-64
21-2, 50-54
24-3, 50-54
29-2, 50-54
31-2, 50-54
34-3, 50-54
36-1, 81-84
37-2, 50-54
40-1, 50-54
42-1, 50-54
43-2, 50-54
48-2, 50-54
49-1, 50-54
50-2, 50-54
51-1, 50-54
52-1, 50-54
54-2, 50-54
1
Is4
2
0
1
4
3
3
6
3
7
2
4
2
5
9
8
17
5
14
7
7
5
16
8
1
1εε
o
<*11
13
14
10
30
18
14
12
9
15
10
16
7
9
10
6
4
9
4
6
5
9
8
10
10
1-50to
11
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
1
2
1
1
§
3
3
1
2
0
1
2
1
3
3
3
2
1
1
2
2
1
0
2
2
1
0
0
4
3
1G
,S2
1a17
21
30
45
24
21
21
23
20
24
15
17
25
23
24
20
16
14
20
21
19
21
15
14
24
11J14
11
5
8
9
11
12
12
14
7
10
6
6
3
6
11
10
3
4
8
7
5
4
5
7
15
o
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
7
4
8
9
9
9
11
12
8
6
9
5
8
5
6
9
3
4
10
9
6
7
8
5
8
114
6
6
4
2
5
4
7
6
4
6
4
3
6
4
7
3
4
5
5
6
7
7
5
5
δ
1Q.
9
7
8
4
2
6
5
4
7
4
11
8
13
16
11
8
9
22
12
7
17
13
11
13
6
1119
4
4
1
3
4
5
6
11
14
13
19
13
13
15
12
17
19
12
15
13
13
17
15
20
10&
0
3
I
2
2
4
4
2
2
2
4
2
1
0
0
0
0
1
1
2
0
1
0
1
1
11IQ,
5
7
5
4
4
6
6
3
6
4
4
4
2
5
4
6
4
3
2
2
4
4
3
4
2
1!S
13
18
17
9
13
10
11
10
8
10
12
12
16
13
9
9
9
11
11
13
12
13
9
11
9
11Q
-QO
12
1
1
1
1
2
2
2
4
0
2
2
1
2
0
3
2
4
4
1
4
1
2
3
3
cg
>CD
CD
αZ
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
c/>
Eou.
CD
roo
"5JQ
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
EO
LLC
oQQ
r
R
R
R
R
r
r
r
r
r
r
r
r
r
r
r
r
r
r
r
r
r
r
r
r
"O
c
c o.2 S
oè
80
<l<l
<l
<l
10
10
10
10
10
10
20
40
80
60
40
60
80
80
10
50
50
20
10
<l
Note: See Table 14 for explanations of symbols.
Lychnocanoma elongata (Vinassa), Riedel and Sanfilippo, 1978b,p. 70, pi. 7, fig. 4.
Lychnodictyum audax RiedelLychnodictyum audax Riedel, 1953, p. 810, pi. 85, fig. 9.
Phormostichoartus corbula (Harting)Lithocampe corbula Harting, 1863, p. 12, pi. 1, fig. 21.Phormostichoartus corbula (Harting), Nigrini, 1977, p. 252, pi. 1,fig. 10.
Phormostichoartus doliolum (Riedel and Sanfilippo)Artostrobium doliolum Riedel and Sanfilippo, 1971, p. 1599, pi.1H, figs. 1-3, pi. 8, figs. 14, 15; Westberg and Riedel, 1978, p. 20,pi. 3, figs. 10, 11.Phormostichoartus doliolum (Riedel and Sanfilippo), Nigrini,1977, p. 252, 253, pi. 1, fig. 14.
Phormostichoartus fistula NigriniPhormostichoartus fistula Nigrini, 1977, p. 253, pi. 1, figs. 11-13.
Podocyrtis mitra EhrenbergPodocyrtis mitra Ehrenberg, 1854, pi. 36, fig. B20; Riedel andSanfilippo, 1978b, p. 72, pi. 8, fig. 7.
Podocyrtis papalis EhrenbergPodocyrtis paplis Ehrenberg, 1847, fig. 2; Riedel and Sanfilippo,1970, p. 532, pi. 11, fig. 1.
Pterocanium prismatium RiedelPterocanium prismatium Riedel, 1957, p. 87, pi. 3, figs. 4, 5;Riedel and Sanfilippo, 1978b, p. 72, pi. 9, fig. 1.
Sethochytris triconiscus HaeckelSethochytris triconiscus Haeckel, 1887, p. 1239, pi. 57, fig. 13;Riedel and Sanfilippo, 1978b, p. 73, pi. 9, fig. 6.
Siphostichoartus corona (Haeckel)Cyrtophormis (Acanthocyrtis) corona Haeckel, 1887, p. 1462, pi.77, fig. 15.Phormostichoartus corona (Haeckel), Riedel and Sanfilippo, 1978b,p. 71, pi. 7, fig. 12.Siphostichoartus corona (Haeckel), Nigrini, 1977, p. 257, pi. 2,figs. 5-7.
Solenosphaera omnitubus Riedel and SanfilippoSolenosphaera omnitubus omnitubus Riedel and Sanfilippo, 1971,p. 1586, pi. 1A, fig. 24, pi. 4, figs. 1, 2; 1978b, p. 73, pi. 9, figs.8, 9.
Spirocyrtis gyroscalaris NigriniSpirocyrtis gyroscalaris Nigrini, 1977, p. 258, pi. 2, figs. 10, 11.
Spirocyrtis scalaris HaeckelSpirocyrtisscalaris Haeckel, 1887, p. 1509, pi. 76, fig. 14; Nigrini,1977, p. 259, pi. 2, figs. 12, 13.
421

M. J. WESTBERG, W. R. RIEDEL
Site 494COLLOSCOCCODSPYRIDPLAGONPTEROCTHEOPECARPOCCANNOBARTOSTPYLONILITHELACTINOSPONGO
.30 -.20 .11 .01 .08 .18 .27 .36 .46 .55 .65 .74 .84
Site 496COLLOSACTINOCOCCODLITHELSPYRIDPLAGONTHEOPEPTEROCARTOSTSPONGOCARPOCPYLONICANNOB
-.25 -.15 -.06 .03 .13 .22 .32 .41 .50 .60 .69 .79
Capo Rossello
r
COLLOSACTINOSPONGOLITHELPYLONICOCCODSPYRIDCANNOBPLAGONARTOSTTHEOPEPTEROCCARPOC
-.28-.17 -.07-.04 .15 .25 .36 .47 .58 .68 .79 .90 1.00
Site 495COLLOSPTEROCSPYRIDPLAGONCARPOCCANNOBPYLONISPONGOACTINOARTOSTCOCCODLITHELTHEOPE
.30.22- .14 -.06 .02 .10 .18 .27 .35 .43 .51 .59 .68
Site 503COLLOSPLAGONTHEOPECANNOBSPYRIDACTINOCARPOCPYLONIPTEROCCOCCODARTOSTSPONGOLITHEL
.20.14 - .07.01 .06 .12 .19 .25 .32 .38 .45 .51 .58
Figure 4. Dendrograms showing family clusters at margin Sites 494 and 496, Capo Rossello, and at oceanic Sites 495 and 503. (Values onthe horizontal axis are similarities.)
Spongaster berminghami (Campbell and Clark)Spongasteriscus berminghami Campbell and Clark, 1944a, p. 30,pi. 5, figs. 1, 2.Spongaster berminghami (Campbell and Clark), Riedel and San-filippo, 1978b, p. 73, pi. 2, figs. 14-16.
Spongaster pentas Riedel and SanfilippoSpongasterpentas Riedel and Sanfilippo, 1970, p. 523, pi. 15, fig.3; 1978b, p. 74, pi. 2, figs. 5-8.
Spongaster tetras EhrenbergSpongaster tetras Ehrenberg, 1860, p. 833; Riedel and Sanfilippo,1978b, p. 74, pi. 2, figs. 2, 3.
Spongodiscus ambus Sanfilippo and RiedelSpongodiscus ambus Sanfilippo and Riedel, 1974, p. 1024, pi. 1,figs. 12-14.
Stichocorys armata (Haeckel)Cyrtophormis armata Haeckel, 1887, p. 1460, pi. 78, fig. 17.Stichocorys armata (Haeckel), Riedel and Sanfilippo, 1971, p.1595, pi. 2E, figs. 13-15.
Stichocorys delmontensis (Campbell and Clark)Eucyrtidium delmontense Campbell and Clark, 1944a, p. 56, pi. 7,figs. 19, 20.Stichocorys delmontensis (Campbell and Clark), Westberg andRiedel, 1978, p. 22, pi. 3, figs. 1-5.
Stichocorys peregrina (Riedel)Eucyrtidium elongatum peregrinum Riedel, 1953, p. 812, pi. 85,fig. 2.Stichocorys peregrina (Riedel), Westberg and Riedel, 1978, p. 22,pi. 3, figs. 6-9.
Stichocorys wolffii HaeckelStichocorys wolffii Haeckel, 1887, p. 1479, pi. 80, fig. 10; Riedeland Sanfilippo, 1978b, p. 74, pi. 9, fig. 12.
Theocorys spongoconus KlingTheocorys spongoconus Kling, 1971, p. 1087, pi. 5, fig. 6.
Theocorythium trachelium (Ehrenberg)Eucyrtidium trachelius Ehrenberg, 1872, p. 312.Theocorythium trachelium (Ehrenberg), Nigrini, 1967, pp. 77-81,
422

RADIOLARIANS FROM THE MIDDLE AMERICA TRENCH
Table 18. Correlation coefficients of some family pairs in the Neogeneof the eastern tropical Pacific and Sicily.
1. Plagoniids/theoperids2. Plagoniids/pterocorythids3. Theoperids/pterocorythids4. Theoperids/spyrids5. Spyrids/pterocorythids6. Theoperids/artostrobiids7. Spongodiscids/spyrids8. Spongodiscids/plagoniids9. Spongodiscids/pterocorythids
10. Plagioniids/spyrids11. Spongodiscids/artostrobiids12. Actinommids/spyrids13. Spongodiscids/theoperids14. Actinommids/plagoniids
GuatemalanMargin
494
+ 0.8+ 0.8+ 0.8+ 0.7+ 0.7+ 0.5-0.8-0.8-0.8+ 0.7-0.6-0.6-0.8-0.5
496
+ 0.8+ 0.7+ 0.5+ 0.6+ 0.6+ 0.6-0.6-0.7-0.6+ 0.8-0.4-0.5-0.7-0.5
CapoRossello
+ 0.6+ 0.6+ 0.5+ 0.5+ 0.5+ 0.7-0.8-0.9-0.6+ 0.8-0.8-0.4-0.9-0.5
Open Ocean
CocosPlate495
+ 0.2+ 0.06-0.04-0.06+ 0.1+ 0.02-0.03+ 0.02-0.2+ 0.6-0.5-0.7-0.5-0.8
East PacificRise503
+ 0.5-0.2-0.5+ 0.09+ 0.2-0.3-0.2-0.5+ 0.04+ 0.2+ 0.06-0.4-0.6-0.6
pi. 8, figs. 1, 2, pi. 9, figs. 1, 2 [including subspecies T. t. dianaβ];Riedel and Sanfilippo, 1978b, p. 76, pi. 9, fig. 17.
Theocorythium vβtulum NigriniTheocorythium vetulum Nigrini, 1971, p. 447, pi. 34.1, figs. 6a, b.
Theocotyle cryptocephala (Ehrenberg)Eucyrtidium cryptocephalum Ehrenberg, 1873, p. 227.Theocotyle cryptocephala cryptocephala (Ehrenberg), Riedel andSanfilippo, 1978b, p. 78, pi. 9, fig. 19.Theocotyle cryptocephala (Ehrenberg), Sanfilippo and Riedel, inpress.
Theocyrtis annosa (Riedel)Phormocyrtis annosa Riedel, 1959, p. 295, pi. 2, fig. 7.Theocyrtis annosa (Riedel), Riedel and Sanfilippo, 1978b, p. 78,pi. 10, fig. 3.
Thyrsocyrtis (Thyrsocyrtis) bromia EhrenbergThyrsocyrtis bromia Ehrenberg, 1873, p. 260; Riedel and San-filippo, 1978b, p. 78, pi. 10, figs. 4, 5.Thyrsocyrtis {Thyrsocyrtis) bromia Ehrenberg, Sanfilippo andRiedel, in press.
Thyrsocyrtis (Thyrsocyrtis) rhizodon EhrenbergThyrsocyrtis rhizodon Ehrenberg, 1873, p. 262.Thyrsocyrtis (Thyrsocyrtis) rhizodon Ehrenberg, Sanfilippo andRiedel, in press.
Thyrsocyrtis (Pentalacorys) triacantha (Ehrenberg)Podocyrtis triacantha Ehrenberg, 1873, p. 254.Thyrsocyrtis triacantha (Ehrenberg), Riedel and Sanfilippo, 1978b,p. 82, pi. 10, figs. 10, 11.Thyrsocyrtis (Pentalacorys) triacantha (Ehrenberg), Sanfilippoand Riedel, in press.
ACKNOWLEDGMENTS
This study was supported by National Science Foundation GrantNo. OCE79-20265. We are grateful to K. J. McMillen and AnnikaSanfilippo for reviewing the manuscript. M. A. Hanger competentlytyped the manuscript and assisted with the proofreading. Evelyn Por-tillo drafted the figures. Alexis Budai and W. Coulbourn advised us onthe statistical analysis of the data on abundances of families.
REFERENCES
Brower, J. C , Millendorf, S. A., and Dyman, T. S., 1978. Methodsfor the quantification of assemblage zones based on multivariateanalysis of weighted and unweighted data. Comput. Geosci., 4:221-227.
Campbell, A. S., and Clark, B. L., 1944a. Miocene radiolarian faunasfrom southern California. Geol. Soc. Am. Spec. Pap., 51:1-76.
, 1944b. Radiolaria from Upper Cretaceous of Middle Cali-fornia. Geol. Soc. Am. Spec. Pap., 57:1-61.
Caulet, J. P., 1971. Contribution a l'étude de quelques radiolairesnassellaires des boues de la Méditerranée et du Pacifique. Cah. Mi-cropaléontol., Sér. 2(10): 1-10.
Cita, M. B., Nigrini, C , and Gartner, S., 1970. Biostratigraphy. InPeterson, M. N. A., Edgar, N. T., et al., Init. Repts. DSDP, 2:Washington (U.S. Govt. Printing Office), 391-411.
Davis, J. C , 1973. Statistics and Data Analysis in Geology: New York(Wiley).
Ehrenberg, C. G., 1847. über die mikroskopischen kieselschaligenPolycystinen als màchtige Gebirgsmasse von Barbados ünd uberdas Verhàltnis der aus mehr als 300 neuen Arten bestehenden ganzeigenthümlichen Formengruppe jener Felsmasse zu den lebendenThieren und zur Kreidebildung. Eine neue Anregung zur Erfor-schung der Erdlebens. Monatsber. Kg I. Preuss. Akad. Wiss. BerlinJahrg., pp. 40-60.
, 1854. Mikrogeologie: Leipzig (Leopold Voss)., 1860. Uber den Tiefgrund des stillen Oceans zwischen Cali-
fornien und den Sandwich-Inseln aus bis 15600' Tiefe nach Lieut.Brooke. Monatsber. Kgl. Preuss. Akad. Wiss. Berlin Jahrg., pp.819-833.
., 1872. Mikrogeologischen Studien als Zusammenfassungseiner Beobachtungen des kleinsten Lebens der Meeres-Tiefgrundealler Zonen und dessen geologischen Einfluss. Monatsber. Kgl.Preuss. Akad. Wiss. Berlin Jahrg., pp. 265-322.
., 1873. Gróssere Felsproben des Polycystinen-Mergels vonBarbados mit weiteren Erlàuterungen. Monatsber. Kgl. Preuss.Akad. Wiss. Berlin Jahrg., pp. 213-263.
Foreman, H. P., 1968. Upper Maestrichtian Radiolaria of California:Special Papers in Palaeontology (No. 3): Palaeontological Associ-ation, London, 1-82.
, 1975. Radiolaria from the North Pacific, Deep Sea DrillingProject, Leg 32. In Larson, R. L., Moberly, R., et al., Init. Repts.DSDP, 32: Washington (U.S. Govt. Printing Office), 579-676.
Haeckel, E., 1887. Report on the Radiolaria collected by H.M.S. Chal-lenger during the years 1873-76. Report on the Scientific Resultsof the Voyage of the H.M.S. Challenger, Zoology, 18:1-1303.
Harting, P., 1863. Bijdrage tot de kennis der mikroskopische faune enflora van de Banda-Zee, naar aanleiding van een onderzoek vaneenige door diepzeeloodingen van 990 tot 4000 vademen uit die zeeopgebragte gronden. K. Acad. Wet. Amsterdam Verh., 10:1-34.
Hay, W. W., 1972. Probabilistic stratigraphy. Eclogae Geol. Helv.,65(2):255-266.
Hay, W. W., and Steinmetz, J. C , 1973. Probabilistic analysis of dis-tribution of Late Paleocene-Early Eocene calcareous nanno-fossils. In Smith, L. A., and Hardenbol, J. (Eds.), Proceedings ofSymposium on Calcareous Nannofossils: Houston (Gulf CoastSection, Soc. Econ. Paleontol. Mineral.), pp. 58-70.
Hays, J. D., 1965. Radiolaria and late Tertiary and Quaternary his-tory of Antarctic seas: Biology of the Antarctic Seas. II. Antarct.Res. Ser., 5:125-184.
, 1970. Stratigraphy and evolutionary trends of radiolariain North Pacific deep sea sediments. In Hays, J. D. (Ed.),Geological Investigations of the North Pacific. Geol. Soc. Am.Mem., 126:185-218.
Hays, J. D., Saito, T., Opdyke, N. D., and Burckle, L. H., 1969. Plio-Pleistocene sediments of the equatorial Pacific: their paleomag-netic, biostratigraphic, and climatic record. Geol. Soc. Am. Bull.,80:1481-1514.
Johnson, D. A., and Knoll, A. H., 1975, Absolute ages of Quaternaryradiolarian datum levels in the equatorial Pacific. Quat. Res.,5:99-110.
Kling, S. A., 1971. Radiolaria: Leg 6 of the Deep Sea Drilling Project.In Fischer, A. G., Heezen, B. C , et al., Init. Repts. DSDP, 6:Washington (U.S. Govt. Printing Office), 1069-1117.
, 1973. Radiolaria from the eastern North Pacific, Deep SeaDrilling Project, Leg 18. In Kulm, L. D., von Huene, R., et al.,Init. Repts. DSDP, 18: Washington (U.S. Govt. Printing Office),617-672.
., 1977. Local and regional imprints on radiolarian assem-blages from California coastal basin sediments. Mar. Micropale-ontol, 2(3):207-221.
Knoll, A. H., and Johnson, D. A., 1975. Late Pleistocene evolutionof the collosphaerid radiolarian Buccinosphaera invaginata Hae-ckel. Micropaleontology, 21(l):60-68.
McCammon, R. B., 1970. On estimating the relative biostratigraphicvalue of fossils. Bull. Geol. Inst. Univ. Uppsala (new series), 2:49-57.
Millendorf, S. A., Brower, J. C , and Dyman, T. S., 1978. A compari-son of methods for the quantification of assemblage zones. Com-put. Geosci., 4:229-242.
423

M. J. WESTBERG, W. R. RIEDEL
Moore, T. C , Jr., 1971. Radiolaria. In Tracey, J. I., Jr., Sutton, G.H., et al., Init. Repts. DSDP, 8: Washington (U.S. Govt. PrintingOffice), 727-775.
, 1972. Mid-Tertiary evolution of the radiolarian genus Calo-cycletta. Micropaleontology, 18:144-152.
Nigrini, C , 1967. Radiolaria in pelagic sediments from the Indian andAtlantic Ocean. Bull Scripps Inst. Oceanogr., 11:1-125.
, 1971. Radiolarian zones in the Quaternary of the equatorialPacific Ocean. In Funnell, B. M., and Riedel, W. R. (Eds.), TheMi-cropalaeontology of the Oceans: Cambridge (Cambridge UniversityPress), pp. 443-461.
., 1977. Tropical Cenozoic Artostrobiidae (Radiolaria). Micro-paleontology, 23(3):241-269.
Pessagno, E. A., 1972. Cretaceous Radiolaria. Part II. Pseudoaulopha-cidae Riedel from the Cretaceous of California and the Blake-Bahama Basin (JOIDES Leg I). Bull. Am. Paleontoi, 61(270):281-328.
Riedel, W. R., 1953. Mesozoic and late Tertiary Radiolaria of Rotti.J. Paleontoi., 27:805-813.
, 1957. Radiolaria: a preliminary stratigraphy. Rep. Swed.Deep-Sea Exped., 6:59-96.
., 1959. Oligocene and Lower Miocene Radiolaria in tropicalPacific sediments. Micropaleontology, 5:285-302.
Riedel, W. R., and Sanfilippo, A., 1970. Radiolaria, Leg 4, Deep SeaDrilling Project. In Bader, R. G., Gerard, R. D., et al., Init.Repts. DSDP, 4: Washington (U.S. Govt. Printing Office),503-575.
, 1971. Cenozoic Radiolaria from the western tropical Pa-cific, Leg 7. In Winterer, E. L., Riedel, W. R., et al., Init. Repts.DSDP, 7, Pt. 2: Washington (U.S. Govt. Printing Office),1529-1672.
., 1978a. Radiolaria. Utrecht Micropaleontol. Bull., 17:81-128.
., 1978b. Stratigraphy and evolution of tropical Cenozoic ra-diolarians. Micropaleontology, 24(l):61-96.
Riedel, W. R., and Westberg, M. J., in press. Neogene radiolariansfrom the eastern tropical Pacific and Caribbean, DSDP Leg 68. InPrell, W., and Gardner, J. V., et al., Init. Repts. DSDP, 68:Washington (U.S. Govt. Printing Office).
Sanfilippo, A., Burckle, L. H., Martini, E., and Riedel, W. R., 1973.Radiolarians, diatoms, silicoflagellates and calcareous nanno-
fossils in the Mediterranean Neogene. Micropaleontology, 19(2):209-234.
Sanfilippo, A., and Riedel, W. R., 1970. Post-Eocene "closed" the-operid radiolarians. Micropaleontology, 16(4):446-462.
, 1973. Cenozoic Radiolaria (exclusive of theoperids, arto-strobiids and amphipyndacids) from the Gulf of Mexico, DSDPLeg 10. In Worzel, J. L., Bryant, W., et al., Init. Repts. DSDP, 10:Washington (U.S. Govt. Printing Office), 475-611.
., 1974. Radiolaria from the West-Central Indian Ocean andGulf of Aden, DSDP Leg 24. In Fisher, R. L., Bunce, E. T., et al.,Init. Repts. DSDP, 24: Washington (U.S. Govt. Printing Office),997-1035.
., 1980. A revised generic and suprageneric classification ofthe Articins (Radiolaria). J. Paleontoi., 54(5):1008-1012.
., in press. Revision of the radiolarian genera Theocotyle,Theocotylissa and Thyrsocyrtis. Micropaleontology.
Squinabol, S., 1903. Le Radiolarie dei noduli selciosi nella Scagliadegli Euganei. Contribuzione I. Riv. Hal. Paleontoi., 9:105-151.
, 1914. Contributo alia conoscenza dei Radiolarii fossili delVeneto. Appendice—Di un genere di Radiolari caratteristico delsecondario. Mem. 1st. Geol. R. Univ. Padova, 2:249-306.
Strelkov, A. A., and Reshetnyak, V. V., 1962. Kolonialnye radiolyariiSpumellaria yuzhno-kitaiskogo morya (raion yuzhoy okonech-nosti ostrova Hainan). Stud. Mar. Sin., 1:121-139.
, 1971. Colonial Spumellarian radiolarians of the worldocean. In Strelkov, A. A. (Ed.), Exploration of the Fauna ofthe Seas: Moscow (Zoological Institute, Academy of Sciences,U.S.S.R.), IX(XVII):295-373.
Vinassa de Regny, P., 1900. Radiolari miocenici italiani. Mem. R.Accad. Sci. 1st. Bologna Ser. 5, 8:227-257.
Westberg, M. J., and Riedel, W. R., 1978. Accuracy of radiolariancorrelations in the Pacific Miocene. Micropaleontology, 24(1):1-23.
Worsley, T. R., Blechschmidt, G., Ralston, S., and Snow, B., 1973.Probability-based analysis of the area-time distribution of Oligo-cene calcareous nannofossils. In Smith, L. A., and Hardenbol, J.(Eds.), Proceedings of Symposium on Calcareous Nannofossils:Houston (Gulf Coast Section, Soc. Econ. Paleontoi. Mineral.),pp. 71-79.
424





























