Embed Size (px)
Citation preview

f u n g a l e c o l o g y 4 ( 2 0 1 1 ) 1 1 0e1 1 4
ava i lab le a t www.sc iencedi rec t .com
journa l homepage : www.e lsev ie r . com/ loca te / funeco
Short Communication
Decomposing capacity of fungi commonly detected inPinus sylvestris needle litter
Johanna B. BOBERG*, Katarina IHRMARK, Bjorn D. LINDAHL
Uppsala BioCenter, Department of Forest Mycology & Pathology, Swedish University of Agricultural Sciences, Box 7026,
SE-750 07 Uppsala, Sweden
a r t i c l e i n f o
Article history:
Received 25 May 2010
Revision received 20 August 2010
Accepted 3 September 2010
Available online 30 October 2010
Corresponding editor: Petr Baldrian
Keywords:
Cellulose
Decomposition
Dothideomycetes
Helotiales
Humus formation
Lignin
Litter fungi
Pinus sylvestris
* Corresponding author. Tel.: þ46 18 671806;E-mail address: Johanna.Boberg@mykopa
1754-5048/$ e see front matter ª 2010 Elsevdoi:10.1016/j.funeco.2010.09.002
a b s t r a c t
Amajor part of the fungal community in coniferous litter consists of fungi whose taxonomic
position and ecology are unknown. Here, nine isolates from within commonly occurring
phylogenetic groups were tested for their ability to decompose Pinus sylvestris needles. In a 1-
yr long incubation study, needle mass loss as well as changes in cellulose and lignin content
were determined and compared to those caused by two litter basidiomycetes (Marasmius
androsaceus and Mycena epipterygia) with recognized ability to decompose needles. A basid-
iomycetous Clavulina/Sistotrema strain appeared to be cellulolytic but not ligninolytic. Chalara
longipes and three other strains within Helotiales also decomposed cellulose but not lignin,
whereas Mollisia cinerea (also Helotiales) and two Dothideomycetes e Sydowia polyspora and
a Mytilinidion sp., seemed unable to cause significant mass loss of cellulose. Lophodermium
pinastri (Rhytismatales) readily decomposed cellulose, and also caused considerable loss of
lignin.
ª 2010 Elsevier Ltd and The British Mycological Society. All rights reserved.
Saprotrophic fungi are the main decomposers of coniferous
wood and litter (Rayner & Boddy 1988) and, thereby, play
important roles in nutrient and carbon circulation in the
boreal forest. The decomposing capacity of fungi associated
with broad leaf litter in temperate forests has been studied
extensively (reviewed by Osono 2007). Still, the ecophysio-
logical properties of many dominant litter fungi associated
with coniferous litter in the boreal zone remain uncertain.
Basidiomycetes are considered particularly important for the
decomposition of plant material, due to their production of
a wide range of lignocellulolytic enzymes (Osono & Takeda
fax: þ46 18 673599.t.slu.se (J.B. Boberg).ier Ltd and The British M
2002). However, recent studies using molecular methods
have indicated that a major part of the fungal community
present in decomposing litter consists of ascomycetes
(O’Brien et al. 2005; Lindahl et al. 2007). About 50e70 % of PCR
amplicons obtained from coniferous and deciduous litter were
attributed ascomycetous taxa, and the most abundant geno-
types were assigned either to the Leotiomycetes or to the
Dothideomycetes. Earlier observations have indicated that the
litter decomposing abilities of basidiomycetes to a large extent
exceed those of ascomycetes, and that most ascomycetes
have limited ability to decompose lignin, Xylariales
ycological Society. All rights reserved.

Decomposing capacity of fungi 111
(Sordariomycetes) being an exception (Osono & Takeda 2002).
In this study we investigated the capacity of some commonly
detected needle litter fungi to decompose different chemical
fractions of Pinus sylvestris needle litter.
The study included six Leotiomycete strains: the facultative
endophyte Lophodermium pinastri (Rhytismatales), Mollisia cin-
erea, Chalara longipes and three unidentified strains within the
Helotiales. In addition, two Dothideomycete strains e Sydowia
polyspora (Dothideales) and a Mytilinidion sp. (Pleosporales),
were included (Table 1). A strain within the Clavulina/Sistotrema
group (Cantharellales) was also tested together with the
recognized litter decomposing fungiMarasmius androsaceus and
Mycena epipterygia (Cox et al. 2001; Boberg et al. 2008, 2009).
M. androsaceus and M. epipterygia were isolated from sporo-
carps, and the ascomycetes together with the Clavulina/Sisto-
trema strain were isolated from Scots pine needles collected
from the forest floor at different localities in central Sweden
(Table 1). Stock cultures were maintained on Hagem agar
(Stenlid 1985), except for L. pinastri which was cultured on
vegetable juice agar (200ml Granini� vegetable juice, mainly
containing tomato, celery and carrot juice, 800 ml H2O, 20 g
agar). To confirm the identities ofM. androsaceus, M. epipterygia
and L. pinastri, and to identify the rest of the isolates, the ITS
region of the ribosomal DNA was sequenced following the
method described by Persson et al. (2009). The obtained
sequences were compared with identified reference sequences
at NCBI using the BLASTn algorithm. When no close matches
were found in the database, similar ITS sequences were
selected, aligned and analysed for similarity by neighbour
joining. All fungal strains are deposited at the Department of
Forest Mycology and Pathology, SLU, Uppsala, Sweden
(the isolate of L. pinastri is also deposited at CBS), and the
sequences are deposited at NCBI (Table 1).
Decomposing capacity was assayed in plastic Petri dishes
filled with 30 ml of 1.5 % water agar. Approximately 600 mg of
freeze dried, weighed and autoclaved (121 �C for 15 min)
brown, abscised P. sylvestris needles was added to the Petri
dishes. The dishes were inoculated with 4 mm diameter plugs
obtained from fungal stock cultures, sealed with Parafilm to
keep the systems axenic and restrict evaporation, and incu-
bated in darkness at 20 �C. Seven replicates for each strain, as
well as uninoculated control systems, were prepared. After
365 d the systems were harvested and the needles were dried
and weighed for estimation of mass loss. Lignin and cellulose
contents of the decomposed needles, pooled for each fungal
strain, were determined by proximate analysis using the acid
detergent fibre (ADF) sulphuric acid method (Rowland &
Roberts 1994). Briefly, a milled sample was boiled in acid
detergent to remove protein, lipids, pectin, starch, water
soluble carbohydrates and hemi-cellulose (ADF soluble frac-
tion), leavingafibre residue. Cellulosewas removedby treating
the fibre fraction with 72 % sulphuric acid, leaving ‘lignin’ and
ash. Thus, cellulose content is defined as the difference
between the weight before and after sulphuric acid treatment.
Lignin was determined by weight loss of the residue upon
ashing. Chitin, a constituent of fungal cell walls, will mainly
end up in the ADF soluble fraction (Fioretto et al. 2005).
All fungal strains were able to grow on the needles and
covered the Petri dishes at the end of the incubation
period. Needles colonized by M. androsaceus, M. epipterygia
and L. pinastri were clearly bleached, a feature associated
with lignin break down (Osono 2007), whereas needles
colonized by the other fungal strains appeared unaffected
or darker than control needles. M. cinerea, S. polyspora and
the Mytilinidion strain also produced dark coloured aerial
hyphae.
The five helotialean strains caused between 16 and 28 %
needle mass loss (Table 1). They lacked ligninolytic capacity,
but the decreases in the cellulose fraction (5e29 %) caused by
four of the five strains show that they exhibit cellulolytic
capacities, although to a lower extent than the basidiomycetes
(33e69 %). The order Helotiales encompasses a diverse group of
ecologically different fungi; plant endophytes and pathogens,
saprotrophs on wood and litter and mutualistic ericoid- or
ectomycorrhizal fungi (Vralstad et al. 2002; Allmer et al. 2006;
Korkama-Rajala et al. 2008). C. longipes caused the highest
mass loss of the helotialean strains. In earlier studies, decom-
position of spruce needles by C. longipes appeared limited
(Koukol et al. 2006b; Osono & Takeda 2006). Two helotialean
strains belonged to the subfamily Hyaloscyphoideae within the
familyHyaloscyphaceae (Cantrell &Hanlin 1997), and appeared
to be varieties of the same species. Although closely related,
they differed in cellulolytic capacity. Similar genotypes were
frequently detected in litter from Swedish pine forests
(Helotiales group A in Lindahl et al. 2007; J. Stenlid et al.
unpublished) and appeared to persist during later decomposi-
tion stages. The taxonomic placement of Helotiales 1 remains
uncertain. M. cinerea caused the lowest needle mass loss of the
helotialean strains and did not appear to alter the cellulose
fraction. The species M. cinerea includes a range of different
varieties, of which many are reported as endophytes, but are
also found frequently in decomposing litter (Kowalski & Rys
1996; Korkama-Rajala et al. 2008). M. cinerea f. minutella was
found in thewhole organic horizon in the study by Lindahl et al.
(2007) and in well decomposed litter by J. Stenlid et al.
(unpublished). It has been reported to decompose spruce nee-
dles (Koukol et al. 2006b) and compete well with other fungi in
a laboratory study (Koukol et al. 2006a).
TheDothideomycetes S. polyspora and theMytilinidion strain,
caused the lowest needle mass loss, 14 and 8 %, respectively
(Table 1). For the latter, needle mass loss equalled that of the
uninoculated control needles. These two strains caused no
decrease in the cellulose fraction, but instead a slight increase
with up to 4 % (Table 1). Thus, these fungi seem to lack explicit
cellulolytic capacity and mainly utilise soluble compounds in
needles. S. polyspora frequently associates with living, or
recently fallen, needles (Muller et al. 2001; Sinclair & Lyon 2005;
J. Stenlid et al. unpublished). In contrast to the results presented
here, S. polyspora has previously been shown to be capable of
spruce needle decomposition (Muller et al. 2001). Specieswithin
the genera Mytilinidion are typically associated with coniferous
substrata (e.g.Minter 2007). In the studies by Lindahl et al. (2007)
and J. Stenlid et al. (unpublished), Mytilinidion genotypes
(Dothideomycete group B) were mainly found in more decom-
posed needle litter. The apparent lack of litter decomposing
capacity might indicate that this group of fungi depends on the
activity of other decomposers, in order to persist during later
stages of decomposition.
TheClavulina/Sistotrema strain (Moncalvo et al. 2006) caused
33 % needle mass loss and appeared to be cellulolytic with
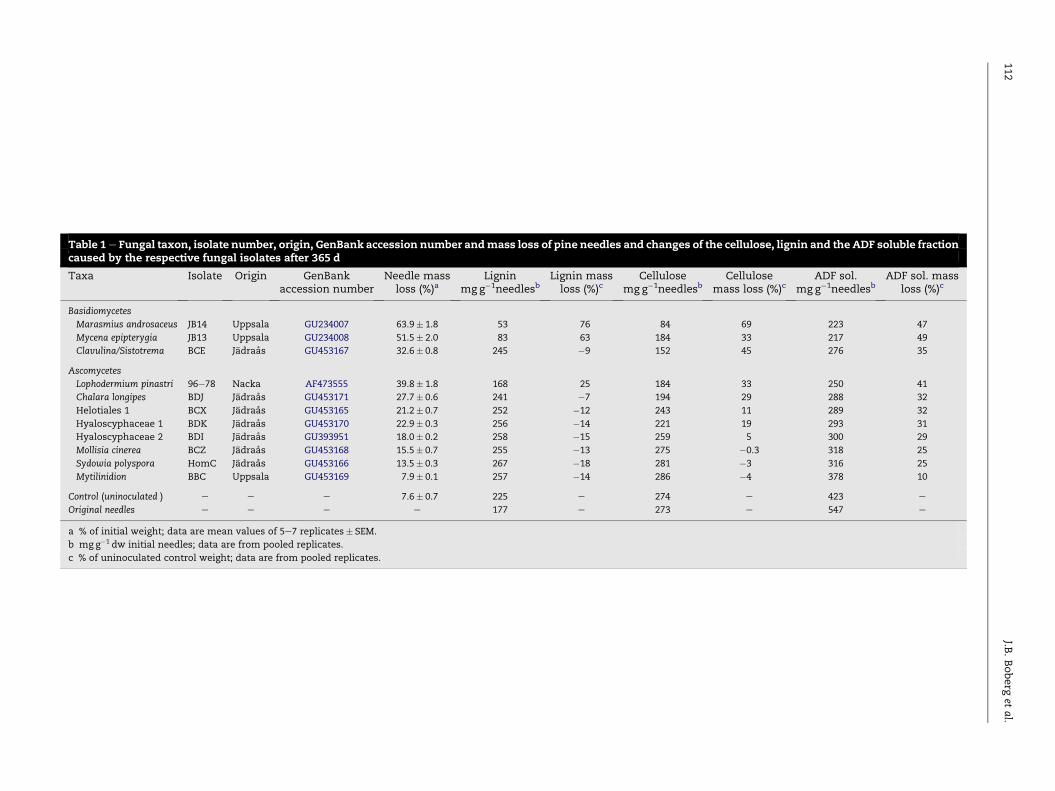
Table 1e Fungal taxon, isolate number, origin, GenBank accession number andmass loss of pine needles and changes of the cellulose, lignin and the ADF soluble fractioncaused by the respective fungal isolates after 365 d
Taxa Isolate Origin GenBankaccession number
Needle massloss (%)a
Ligninmg g�1needlesb
Lignin massloss (%)c
Cellulosemg g�1needlesb
Cellulosemass loss (%)c
ADF sol.mg g�1needlesb
ADF sol. massloss (%)c
Basidiomycetes
Marasmius androsaceus JB14 Uppsala GU234007 63.9� 1.8 53 76 84 69 223 47
Mycena epipterygia JB13 Uppsala GU234008 51.5� 2.0 83 63 184 33 217 49
Clavulina/Sistotrema BCE Jadraas GU453167 32.6� 0.8 245 �9 152 45 276 35
Ascomycetes
Lophodermium pinastri 96e78 Nacka AF473555 39.8� 1.8 168 25 184 33 250 41
Chalara longipes BDJ Jadraas GU453171 27.7� 0.6 241 �7 194 29 288 32
Helotiales 1 BCX Jadraas GU453165 21.2� 0.7 252 �12 243 11 289 32
Hyaloscyphaceae 1 BDK Jadraas GU453170 22.9� 0.3 256 �14 221 19 293 31
Hyaloscyphaceae 2 BDI Jadraas GU393951 18.0� 0.2 258 �15 259 5 300 29
Mollisia cinerea BCZ Jadraas GU453168 15.5� 0.7 255 �13 275 �0.3 318 25
Sydowia polyspora HomC Jadraas GU453166 13.5� 0.3 267 �18 281 �3 316 25
Mytilinidion BBC Uppsala GU453169 7.9� 0.1 257 �14 286 �4 378 10
Control (uninoculated ) e e e 7.6� 0.7 225 e 274 e 423 e
Original needles e e e e 177 e 273 e 547 e
a % of initial weight; data are mean values of 5e7 replicates� SEM.
b mg g�1 dw initial needles; data are from pooled replicates.
c % of uninoculated control weight; data are from pooled replicates.
112
J.B.Boberg
etal.

Decomposing capacity of fungi 113
limited ability to break down lignin (Table 1). Species within
this clade include both saprotrophic as well as ectomycor-
rhizal fungi (Di Marino et al. 2008). The strain tested here was
genetically similar (97 % ITS similarity) to a Cantharellales
taxon (Genbank accession AM999656.1) obtained from bryo-
phytes in a boreal forest in Norway and identical to a sequence
(C1z 3.5) from the moss dominated L horizon in the study by
Lindahl et al. (2007), suggesting thatmossesmay be its primary
substratum.
In systems with species not causing any lignin mass loss,
the ‘lignin’ fraction increased by 7e18 % compared to control
needles (Table 1). It appears as recalcitrant compounds, not
hydrolysable by sulphuric acid, formed due to the presence and
activities of fungi. Even in needles colonized by theMytilinidion
strain, where mass loss did not exceed the control, the acid
insoluble fraction increased during the incubation. A similar
net increase of the acid-unhydrolysable fraction has been
found for decomposing litter in earlier studies (Osono et al.
2006; Preston et al. 2009). �Snajdr et al. (2010) also found that
a significant part of a synthetic lignin analogue was trans-
formed into humic substances during litter decomposition. In
addition, autoclaving of the needles increased the acid insol-
uble fraction by 27 % (Table 1). The cellulose fraction remained
unaltered, suggesting that some compounds found in the ADF
soluble fraction of original needles were converted into more
recalcitrant compounds during autoclaving. As litter undergoes
decomposition, some of the organic material is gradually con-
verted to humus, which has a very long turnover time and
generally accumulates in soils. The exact process of humifica-
tion and the chemical nature of humus are to date not fully
understood (Kogel-Knabner 2002; Sutton & Sposito 2005). The
results presented here suggest that fungal colonisation of litter
could contribute to the formation of recalcitrant compounds,
even without significant decomposition.
L. pinastri caused 40 % needle mass loss and decomposed
both cellulose and lignin (Table 1). L. pinastri caused 25 % lignin
loss relative to autoclaved but uninoculated control needles,
but only a 5 % decrease relative to the original needles before
autoclaving (Table 1). It is therefore not fully certain whether
L. pinastri was able to break down the plant derived lignin or
merely recalcitrant compounds formed during the steriliza-
tion. Still, the lignin mass loss concurred with the observed
lignin break down caused by L. pinastri decomposing pine litter
in a study by Osono & Hirose (2010). L. pinastri is a well known
facultative endophyte, which colonises the needleswhen they
are still attached to the tree, but remains active as a sapro-
troph in litter on the ground. In spite of the extensive ability of
L. pinastri to decompose needles, the species has been found to
be restricted to relatively fresh litter components (Lindahl
et al. 2007; J. Stenlid et al. unpublished), and a low competi-
tive strengthmay limit the overall contribution of L. pinastri to
litter decomposition.
Species belonging to the genera Mycena and Marasmius are
very common in decomposer communities in forest litter
(Frankland 1998; O’Brien et al. 2005; Lindahl et al. 2007). The
isolates of M. androsaceus and M. epipterygia, as expected,
decomposed needles effectively with mass losses reaching 64
and 52 %, respectively (Table 1). Lignin decomposition was
also extensive with a reduction of 76 and 63 % for needles
inoculatedwithM. androsaceus andM. epipterygia, respectively.
These results confirm that these species are well adapted to
use needles as a resource and verify their importance in litter
decomposition. Interestingly, M. epipterygia decomposed
lignin at a higher rate than cellulose compared to M. andro-
saceus, which caused similar cellulose and lignin loss (Table 1).
Selective lignin decomposition has been observed for other
fungal species belonging to the Agaricales decomposing
different types of litter (Osono et al. 2003; Osono & Takeda
2006). Possibly this difference can be related to their ecolog-
ical strategies; M. androsaceus is mainly found in the newly
shed litter in the L horizon (Holmer & Stenlid 1991; Lindahl
et al. 2007), where cellulose is abundant. Mycena species, in
contrast, often colonize litter at slightly later stages of
decomposition (Frankland 1998; Lindahl et al. 2007) with lower
cellulose to lignin ratios.
In conclusion, four out of five tested helotialean strains
decomposed cellulose, whereas the Dothidiomycetes
seemed to lack cellulolytic capacity. Neither of these strains
appeared to have any lignin decomposing capacity.
However, the observed ‘ligninolytic’ capacity of L. pinastri
illustrates that other groups of ascomycetous fungi than the
Xylariales may exhibit significant decay capacity. The
detailed ecological strategies remain unclear for most of the
tested taxa. In the light of the intense focus on boreal litter
decomposition in relation to carbon cycling and global
climate change, it is striking that we know almost nothing
about a major part of the fungal community found in
decomposing needle litter.
Acknowledgements
Financial support from SLU is gratefully acknowledged. The
authors would also like to thank Ond�rej Koukol for identifying
Chalara longipes.
r e f e r e n c e s
Allmer J, Vasiliauskas R, Ihrmark K, Stenlid J, Dahlberg A, 2006.Wood-inhabiting fungal communities in woody debris ofNorway spruce (Picea abies (L.) karst.), as reflected bysporocarps, mycelial isolations and T-RFLP identification.FEMS Microbiology Ecology 55: 57e67.
Boberg JB, Finlay RD, Stenlid J, Lindahl BD, 2009. Fungal Ctranslocation restricts N-mineralization in heterogeneousenvironments. Functional Ecology 24: 454e459.
Boberg J, Finlay RD, Stenlid J, Nasholm T, Lindahl BD, 2008.Glucose and ammonium additions affect needledecomposition and carbon allocation by the litter degradingfungus Mycena epipterygia. Soil Biology and Biochemistry 40:995e999.
Cantrell SA, Hanlin RT, 1997. Phylogenetic relationships in thefamily Hyaloscyphaceae inferred from sequences of itsregions, 5.8s ribosomal DNA and morphological characters.Mycologia 89: 745e755.
Cox P, Wilkinson SP, Anderson JM, 2001. Effects of fungal inoculaon the decomposition of lignin and structuralpolysaccharides in Pinus sylvestris litter. Biology and Fertility ofSoils 33: 246e251.
Di Marino E, Scattolin L, Bodensteiner P, Agerer R, 2008. Sistotremais a genus with ectomycorrhizal species e confirmation of

114 J.B. Boberg et al.
what sequence studies already suggested. Mycological Progress7: 169e176.
Fioretto A, Di Nardo C, Papa S, Fuggi A, 2005. Lignin and cellulosedegradation and nitrogen dynamics during decomposition ofthree leaf litter species in a Mediterranean ecosystem. SoilBiology and Biochemistry 37: 1083e1091.
Frankland JC, 1998. Fungal succession-unravelling theunpredictable. Mycological Research 102: 1e15.
Holmer L, Stenlid J, 1991. Population structure and mating systemin Marasmius androsaceus. New Phytologist 119: 307e314.
Kogel-Knabner I, 2002. The macromolecular organic compositionof plant and microbial residues as inputs to soil organicmatter. Soil Biology and Biochemistry 34: 139e162.
Korkama-Rajala T, Mueller MM, Pennanen T, 2008. Decompositionand fungi of needle litter from slow- and fast-growing Norwayspruce (Picea abies) clones. Microbial Ecology 56: 76e89.
Koukol O, Mrnka L, Kulhankova A, Vosatka M, 2006a. Competitionof Scleroconidioma sphagnicola with fungi decomposing sprucelitter needles. Canadian Journal of Botany 84: 469e476.
Koukol O, Novak F, Hrabal R, Vosatka M, 2006b. Saprotrophicfungi transform organic phosphorus from spruce needle litter.Soil Biology and Biochemistry 38: 3372e3379.
Kowalski T, Rys J, 1996. Endophytic fungi inneedles ofAbies albaandPicea abies. Acta Agraria et Silvestria. Series Silvestris 34: 33e45.
Lindahl BD, Ihrmark K, Boberg J, Trumbore SE, Hogberg P,Stenlid J, Finlay RD, 2007. Spatial separation of litterdecomposition and mycorrhizal nitrogen uptake in a borealforest. New Phytologist 173: 611e620.
Minter DW, 2007. Mytilinidion mytilinellum. IMI Descriptions of Fungiand Bacteria 174: 1738.
Moncalvo JM, Nilsson RH, Koster B, Dunham SM, Bernauer T,Matheny PB, Porter TM, Margaritescu S, Weiss M, Garnica S,Danell E, Langer G, Langer E, Larsson E, Larsson KH, Vilgalys R,2006. The cantharelloid clade: dealing with incongruent genetrees and phylogenetic reconstruction methods. Mycologia 98:937e948.
Muller MM, Valjakka R, Suokko A, Hantula J, 2001. Diversity ofendophytic fungi of single Norway spruce needles and theirrole as pioneer decomposers. Molecular Ecology 10: 1801e1810.
O’Brien HE, Parrent JL, Jackson JA, Moncalvo JM, Vilgalys R, 2005.Fungal community analysis by large-scale sequencing ofenvironmental samples. Applied and Environmental Microbiology71: 5544e5550.
Osono T, Fukasawa Y, Takeda H, 2003. Roles of diverse fungi inlarch needle-litter decomposition. Mycologia 95: 820e826.
Osono T, 2007. Ecology of ligninolytic fungi associated with leaflitter decomposition. Ecological Research 22: 955e974.
Osono T, Hirose D, 2010. Colonization and lignin decompositionof pine needle litter by Lophodermium pinastri. Forest Pathology.doi:10.1111/j.1439-0329.2010.00648.x.
Osono T, Hobara S, Koba K, Kameda K, 2006. Reduction of fungalgrowth and lignin decomposition in needle litter by avianexcreta. Soil Biology and Biochemistry 38: 1623e1630.
Osono T, Takeda H, 2002. Comparison of litter decomposingability among diverse fungi in a cool temperate deciduousforest in Japan. Mycologia 94: 421e427.
Osono T, Takeda H, 2006. Fungal decomposition of Abies needleand Betula leaf litter. Mycologia 98: 172e179.
Persson Y, Vasaitis R, Langstrom B, Ohrn P, Ihrmark K, Stenlid J,2009. Fungi vectored by the bark beetle Ips typographusfollowing hibernation under the bark of standing trees and inthe forest litter. Microbial Ecology 58: 651e659.
Preston CM, Nault JR, Trofymow JA, Smyth C, Grp CW, 2009.Chemical changes during 6 years of decomposition of 11 littersin some Canadian forest sites. Part 1. Elemental composition,tannins, phenolics, and proximate fractions. Ecosystems 12:1053e1077.
Rayner ADM, Boddy L, 1988. Fungal Decomposition of Wood e itsbiology and ecology. John Wiley & Sons Ltd, Portsmouth.
Rowland AP, Roberts JD, 1994. Lignin and cellulose fractionationin decomposition studies using acid-detergent fibre methods.Communications in Soil Science and Plant Analysis 25: 269e277.
Sinclair WA, Lyon HH, 2005. Diseases of Tree and Shrubs. CornellUniversity Press, New York.
�Snajdr J, Steffen KT, Hofrichter M, Baldrian P, 2010.Transformation of 14C-labelled lignin and humic substances inforest soil by the saprobic basidiomycetes Gymnopus erythropusand Hypholoma fasiculare. Soil Biology and Biochemistry 42:1541e1548.
Stenlid J, 1985. Population structure of Heterobasidion annosum asdetermined by somatic incompatibility, sexualincompatibility, and isoenzyme patterns. Canadian Journal ofBotany 63: 2268e2273.
Sutton R, Sposito G, 2005. Molecular structure in soil humicsubstances: the new view. Environmental Science and Technology39: 9009e9015.
Vralstad T, Myhre E, Schumacher T, 2002. Molecular diversity andphylogenetic affinities of symbiotic root-associatedascomycetes of the Helotiales in burnt and metal pollutedhabitats. New Phytologist 155: 131e148.
![Artificial Regeneration of Pinus Sylvestris var. Mongolica ......mongolica [1], afforestation techniques, growth characteristics and ecological functions of Pinus sylvestris var. mongolica,](https://img.pdfslide.us/doc/110x75/60e65d5182ddcc0ef10d2146/artificial-regeneration-of-pinus-sylvestris-var-mongolica-mongolica-1.jpg)

















![Pine - Wikipedia · Pine - Wikipedia 6/20/2017 2:11:22 PM] Pinus sylvestris prepared for transport, Hungary Pine cone Tongue and groove solid pine](https://img.pdfslide.us/doc/110x75/5d62849d88c99309088b4588/pine-pine-wikipedia-6202017-21122-pm-pinus-sylvestris-prepared-for.jpg)










