Embed Size (px)
Citation preview

De-ubiquitination: a new player in Golgi to ER retrograde transport
1Mickaël Cohen, 2Françoise Stutz and 1Catherine Dargemont*
1Nucleocytoplasmic transport group. Institut Jacques Monod. Unité Mixte de Recherche
7592, CNRS, Universités Paris VI and VII, 2 Place Jussieu. Tour 43. 75251 Paris Cedex 05.
France.
2Dept. of Cell Biology. Sciences III. 30 Quai E. Ansermet. 1211 Genève 4. Switzerland.
Running title: De-ubiquitination of β’COP by the Ubp3p/Bre5p complex
*To whom correspondence should be addressed: Institut Jacques Monod. Unité Mixte de
Recherche 7592, CNRS, Universités Paris VI and VII, 2 Place Jussieu. Tour 43. 75251 Paris
Cedex 05. France.
Tel/Fax: 0033 1 44276956
E-mail : [email protected]
Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on October 30, 2003 as Manuscript C300451200 by guest on Septem
ber 27, 2018http://w
ww
.jbc.org/D
ownloaded from

Abstract
Modification by ubiquitin plays a major role in a broad array of cellular functions.
Although reversal of this process, deubiquitination, likely represents an important regulatory
step contributing to cellular homeostasis, functions of deubiquitination enzymes still remain
poorly characterized. We have previously shown that the ubiquitin protease Ubp3p requires a
co-factor, Bre5p, to specifically deubiquitinate the COPII subunit Sec23p, which is involved
in anterograde transport between ER and Golgi compartiments. In the present report, we show
that disruption of BRE5 gene also led to a defect in the retrograde transport from the Golgi to
the Endoplasmic reticulum. Further analysis indicate that the COPI subunit β'-COP represents
another substrate of the Ubp3p/Bre5p complex. All together, our results indicate that the
Ubp3p/Bre5p de-ubiquitination complex co-regulates anterograde and retrograde transports
between ER and Golgi compartments.
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Introduction
Modification of target proteins by ubiquitin participates in a wide array of biological
functions. Proteins destined for degradation or processing via the 26S proteasome are coupled
to multiple copies of ubiquitin. However, attachment of ubiquitin or ubiquitin-related
molecules may also result in changes in subcellular distribution or modification of protein
activity (1,2).
Understanding of mechanisms and regulation of ubiquitin conjugation considerably
improved over the past ten years and recent studies indicate that reversal of this modification,
namely deubiquitination, represents an additional level of regulation. The de-ubiquitination
process is catalyzed by proteases called deubiquitinating enzymes (DUB) which fall into four
distinct families (3), ubiquitin C-terminal hydrolases (UCHs), ubiquitin-specific processing
proteases (USPs or UBPs), OTU-domain ubiquitin-aldehyde-binding proteins (Otubains) and
Jab1/Pad1/MPN-domain-containing metallo-enzymes (JAMMs). Among these four families,
UBPs represent the most widespread and represented DUBs across evolution. In particular,
the Saccharomyces cerevisiae genome encodes for 16 UBPs and only one UCH but none of
these enzymes is essential for yeast viability. UBPs tend to release ubiquitin from a
conjugated protein. They display similar catalytic domains containing conserved Cys and His
boxes but divergent N-terminal and occasionally C-terminal extensions (4,5) which are
thought to function in substrate recognition, subcellular localization and protein-protein
interactions. Molecular basis for substrate recognition by UBP has been poorly described.
Indeed human HAUSP has been shown to be necessary and sufficient to de-ubiquitinate its
specific substrate, the p53 tumor suppressor (6) but no additional substrate for this Ubp has
been identified so far. The tumour suppressor CYLD has been shown to negatively regulate
NF-κB signalling by deubiquitinating NEMO and TRAF2 (7,8,9).
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

We recently reported that the yeast Ubp3p forms a complex with Bre5p which
specifically de-ubiquitinates Sec23p, a component of the COPII complex essential for
anterograde transport between the endoplasmic reticulum (ER) and the Golgi apparatus (10).
Ubp3p is the only yeast Ubp able to catalyze Sec23p deubiquitination indicating that Ubps
can exert their activity on specific substrates and are probably not redundant. Ubp3p directly
interacts with Sec23p and its catalytic cysteine residue is essential for the deubiquitinating
activity. In contrast Bre5p does not participate to Sec23p recognition nor complements a
catalytically inactive Ubp3p but rather acts as an essential positive regulator of Ubp3p. Bre5p
displays an N-terminal domain related to the Nuclear Transport Factor 2 (NTF2) and
responsible for the interaction with Ubp3p and and a C-terminal domain presenting putative
RNA-binding sites. Mammalian cells display two proteins homologous to Bre5p, G3BP1 and
G3BP2. Interestingly, G3BP1 has been shown to interact with USP10 (11), a human ubiquitin
protease sharing 46% similarity and 27% identity with Ubp3p. The interaction between
USP10 and G3BP1 suggests that the Ubp3p/Bre5p complex might have been conserved
during evolution. However there is no clue whether these yeast and human complexes are
implicated in similar functions.
In the present report, we analyzed whether the yeast Ubp3p/Bre5p complex could have
other substrates than Sec23p and whether the human USP10/G3BP1 complex could recognize
these substrates.
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Experimental procedures
Yeast strains and antibodies
The S. cerevisiae strains and plasmids used in this study are listed in the Supplementary Table
. When indicated, the BRE5 and UBP3 genes were disrupted as described (12). Rabbit
polyclonal antibodies to yeast Gcs1p (1:1000 dilution) and Glo3p (1:1000) were kindly
provided by G. Johnston, αCOP (1:800), β'COP (1:200) and δCOP (1:500) by F. Letourneur
and antibodies to βCOP#9381 (1:600), β'COP#9562 (1:1000), γCOP (1:1000), and εCOP
(1:800) were generous gifts from R. Duden.
ER retrieval defect assay of dilysine-tagged Ste2p
The in vivo assay to monitor ER retrieval of dilysine-tagged Ste2p in yeast was performed
essentially as previously described (13). Briefly, MATa cells (wt, ret1.1 and bre5∆) deleted of
the endogenous STE2 gene and expressing the Ste2-WBP1 fusion protein, were mated with
MATα cells (RH311-3D). Resulting diploids were selected on growth medium lacking uracil
and histidine.
Yeast extracts and GST Pull-Down Assay
Cells grown in YPD were collected during the exponential growth phase (A600 2). Total
protein extracts were prepared by the NaOH-TCA lysis method (14). Alternatively cells were
lysed at 4°C with glass beads in IP buffer (50 mM Hepes pH7.5, 150 mM NaCl, 1 mM DTT,
0,5% Triton, 10% Glycerol, supplemented with a mixture of protease inhibitors, 10µg/ml
aprotinine, pepstatine, leupeptine, 1 mM PMSF). The resulting lysate was centrifuged for 30
min at 13000 g. 3µg of GST-WBP1 or GST-WBP1-SS fusion proteins expressed from
plasmids generously provided by F. Letourneur. were incubated with lysates of BRE5 or
bre5∆ cells for 1h at 4°C and an additional hour at 4°C in the presence of glutathione-
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Sepharose beads. Beads were washed in IP buffer and heated in sample buffer before SDS-
PAGE and immunoblotting.
Two hybrid assay
A bait fusion protein between USP10 and the LexA DNA binding domain was expressed
from the pBTM116 plasmid (11). COPI bovine subunits fused to the Gal4 activation domain
were expressed from the pAD-Gal4-2.1 and pACTII plasmids (15). The two-hybrid assay was
performed as previously described (10).
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Results and Discussion
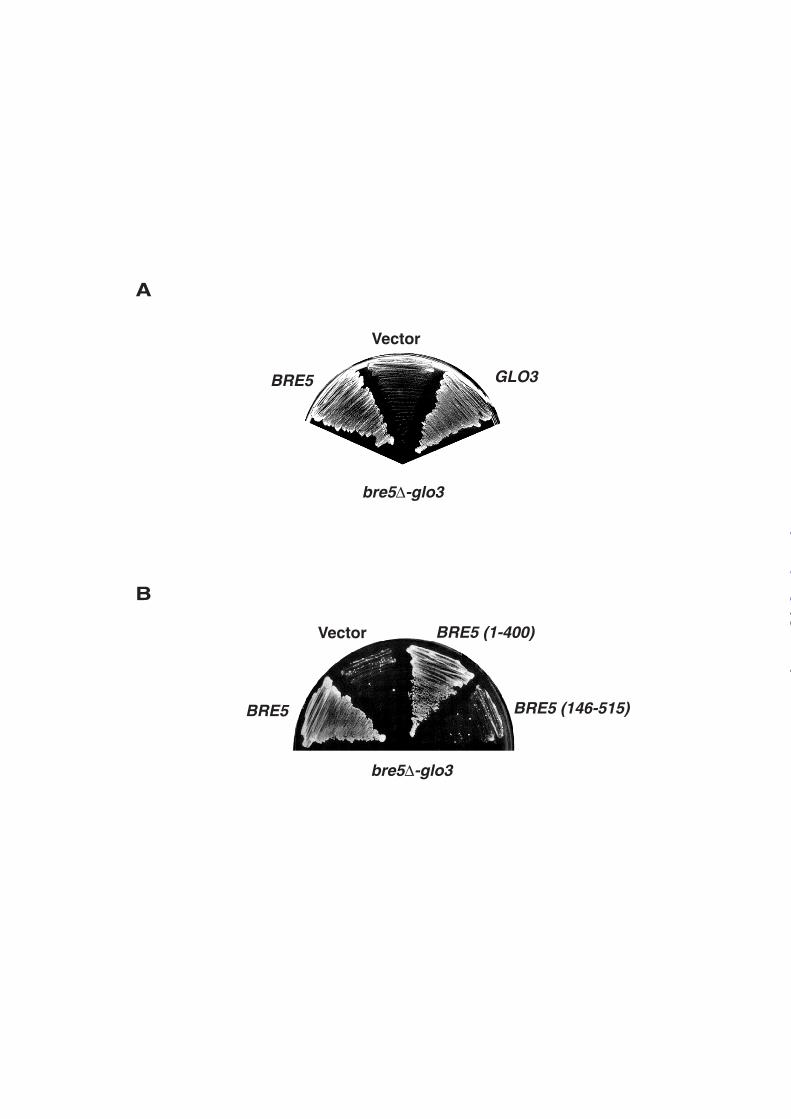
BRE5 genetically interacts with the GLO3 gene. Screening for mutations that induce
synthetic lethality upon disruption of the BRE5 gene recently led to the identification of a
genetic interaction between BRE5 and SFB3, a gene encoding for a Sec24-like component of
the COPII complex. Indeed Bre5p forms a complex with the ubiquitin protease Ubp3p that
de-ubiquitinates the COPII protein Sec23p (10). In order to identify additional substrates of
the Ubp3p/Bre5p complex, we characterized another mutant strain from this screen and found
that the viability of these cells could be rescued by the GLO3 gene. (Figure 1A). Both BRE5
and a truncated version of BRE5 encoding a protein deleted of its RNA-binding domain
(BRE5 (1-400)) could complement the bre5∆-glo3 mutant. In contrast, deletion of the Nuclear
Transport Factor 2 (NTF2)-like domain (BRE5 (146-515)) responsible for the interaction with
Ubp3p abolished the ability of Bre5p to rescue the viability of bre5∆-glo3 mutant (Figure
1B).
ER retrieval defect in cells disrupted for BRE5. Glo3p is one of the GTPase Activating
Proteins (GAP) for the small GTPase ADP-ribosylation factor 1 (Arf1p). Arf1p and the
heptameric coatomer complex (α, β, β’, γ, δ, ε, ζ-COP) form the Coat Protein Complex I
(COPI) involved in retrograde transport within the Golgi apparatus (16) and from the Golgi to
the ER (13,17). The GDP-bound form of Arf1p is likely recruited on Golgi membranes by the
transmembrane protein p23 prior nucleotide exchange. Arf1p-GTP as well as p23/p24
proteins then triggers coatomer binding and coat polymerization (18,19,20). Following
budding, Arf1p-dependent GTP hydrolysis initiates vesicle uncoating (21,22), a prerequisite
for vesicle docking to the acceptor compartment. The three yeast Arf1-GAPs share overall
similarity but appear specialized in specific transport routes. Gcs1p and Age2p have been
shown to be implicated in transport from the Trans Golgi Network (23), whereas Glo3p is the
major GAP involved in ER retrieval (24).
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

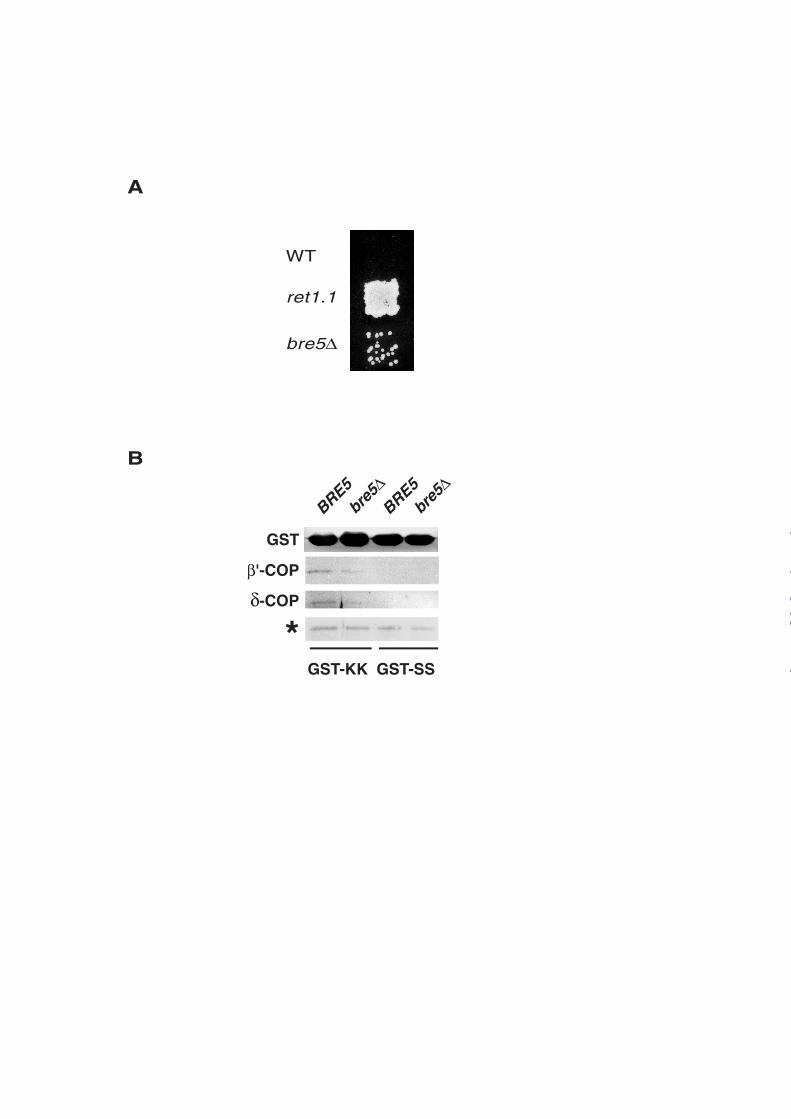
The genetic interaction between GLO3 and BRE5 led us to analyze whether deletion of
BRE5 could induce defects in retrograde transport from the Golgi to the ER. For this purpose,
we used a previously described in vivo assay to monitor ER retrieval of dilysine-tagged
proteins in yeast (13). Indeed, it has been clearly shown that dilysine signals confer a COPI-
mediated localization to the ER (13). This assay is based on the use of MATa cells deleted
from the endogenous α-factor receptor STE2 gene but expressing a fusion protein between
Ste2p and WBP1. The dilysine retrieval motif of WBP1 confers an ER localization to the
chimeric protein resulting in the inability of these ste2∆ Ste2-WBP1 (WT) cells to mate
(Figure 2A). However, when retrograde transport from Golgi to ER is defective, Ste2-WBP1
is mislocalized to the cell surface, allowing mating to occur. In particular, a ts mutation in the
α subunit of the coatomer (ret1.1) was able to restore mating (Figure 2A) as previously
described (13). Deletion of BRE5 led to a partial rescue of the ability to mate indicating that
lack of Bre5p causes a defect in ER retrieval (Figure 2A).
Mutations affecting ER retrieval of dilysine-tagged proteins fall into two classes
distinguished by their ability to alter, or not, binding of coatomer to dilysine motifs in vitro
(13). To better characterize the effects of BRE5 deletion on retrograde transport, lysates from
BRE5 and bre5∆ cells were incubated with purified recombinant fusion proteins between
GST and WBP1 peptides presenting dilysine (GST-WBP1) or diserine motifs (GST-WBP1-
SS) and pulled-down with glutathione beads as previously described (17). Bound proteins
were revealed by immunoblotting with antibodies to β’-COP and δ−COP, two subunits of
COPI coatomer (Figure 2B). Neither β’-COP nor δ−COP were retained on GST-WBP1-SS
beads. Although both subunits were bound to GST-WBP1 beads, the efficiency of recruitment
was affected upon BRE5 deletion. A precise quantification performed on three independent
experiments indicated that the amount of bound β’-COP and δ−COP from bre5∆ cells
extracts corresponded to 50 and 65% respectively of material bound using wt extracts. This
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

result suggests that coatomer assembly is likely altered in bre5∆ cells thus inducing an ER
retrieval defect. Consistently, cells disrupted for BRE5 encoding gene have been shown to be
hypersensitive to Brefeldine A (25), an inhibitor of nucleotide exchange on ARF which
blocks coatomer binding to membranes (26).
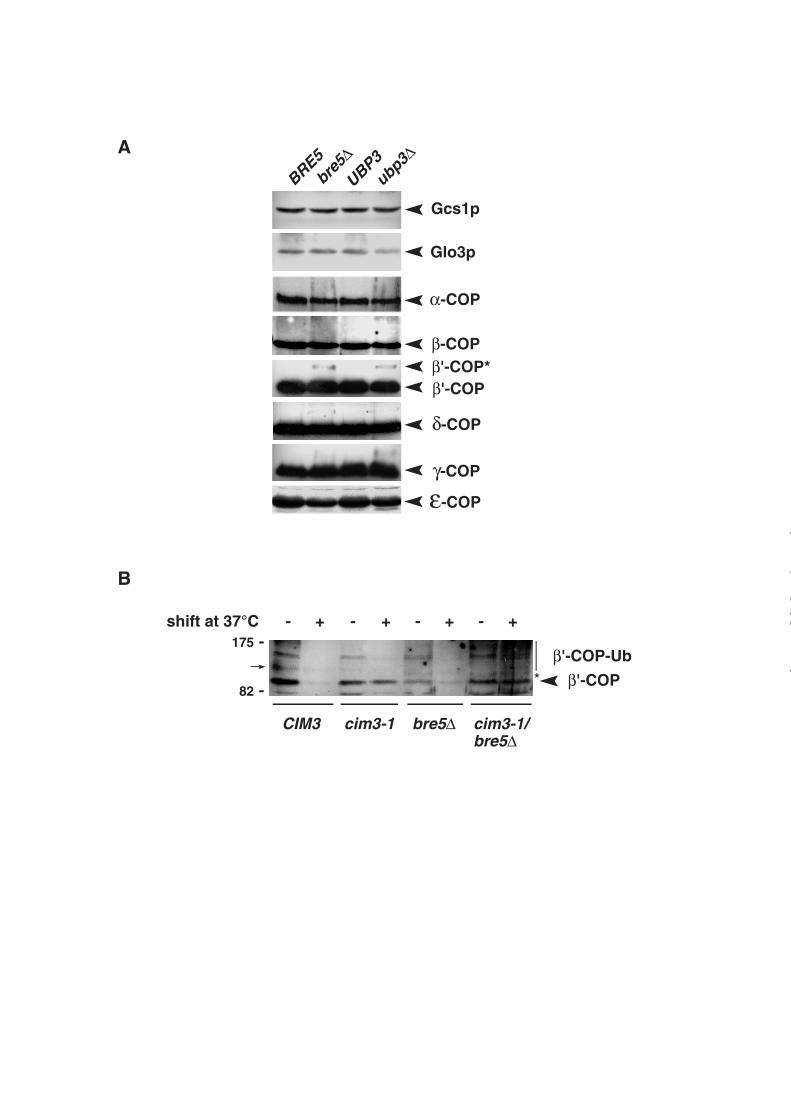
Bre5p/Ubp3p complex de-ubiquitinates the COPI subunit β’-COP/Sec27p. To analyze
the molecular basis of the ER retrieval defect in bre5∆ cells, expression levels of the different
COPI components were analyzed in wild-type, bre5∆ and ubp3∆ cells. No significant
difference could be observed in the steady-state level of the Arf-GAPs Gcs1p or Glo3p nor
coatomer subunits α−COP, β−COP, δ−COP, γ−COP or ε−COP in the various strains (Figure
3A). In contrast, a modified form of β’-COP with slower mobility than the β’-COP main
signal was reproducibly detected in bre5∆ and ubp3∆ cells but not in wild type cells (Figure
3A). The modified form of β’-COP not only displayed a molecular mass of ~10kD greater
than unmodified β’-COP and accumulated in the absence of the Ubp3p/Bre5p de-
ubiquitination complex but β’-COP has also been shown to be ubiquitinated (27). Thus the
modified form of β’-COP most likely corresponds to mono-ubiquitinated β’-COP that could
not be incorporated into the COPI complex and therefore affects coatomer assembly and ER
retrieval.
We recently reported that absence of the Bre5p/Ubp3p complex induces an
accumulation of mono-ubiquitinated Sec23p that facilitates its subsequent poly-ubiquitination
and degradation by the 26S proteasome(10). To address a possible role of the Bre5p/Ubp3p
complex in β’-COP turn-over, we first measured β’-COP stability in the presence of
cycloheximide and found that the half-life of this protein is shorter in bre5∆ cells, than in WT
cells (data not shown). Since polyubiquitination rather than modification with a single
ubiquitin triggers proteins for recognition and degradation by the 26S proteasome,
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

modification of β’-COP was analyzed in cim3-1 thermosensitive mutants defective for the
proteasome activity (28). A two hours shift from 23 to 37°C in the presence of cycloheximide,
led to a rapid degradation of β’-COP in wild-type cells (CIM3) and bre5∆ cells whereas it was
stabilized in cim3-1 strains (Figure 3B) indicating that degradation of β’-COP depends on the
proteasome activity. Interestingly, deletion of BRE5 in cim3-1 cells led to an accumulation at
37°C of both unmodified and high molecular weight forms of β’-COP likely corresponding to
polyubiquitinated species of β’-COP (Figure 3B). These results indicate that absence of a
functional Ubp3p/Bre5p complex led to a faster polyubiquitination and degradation of β’-
COP by the proteasome.
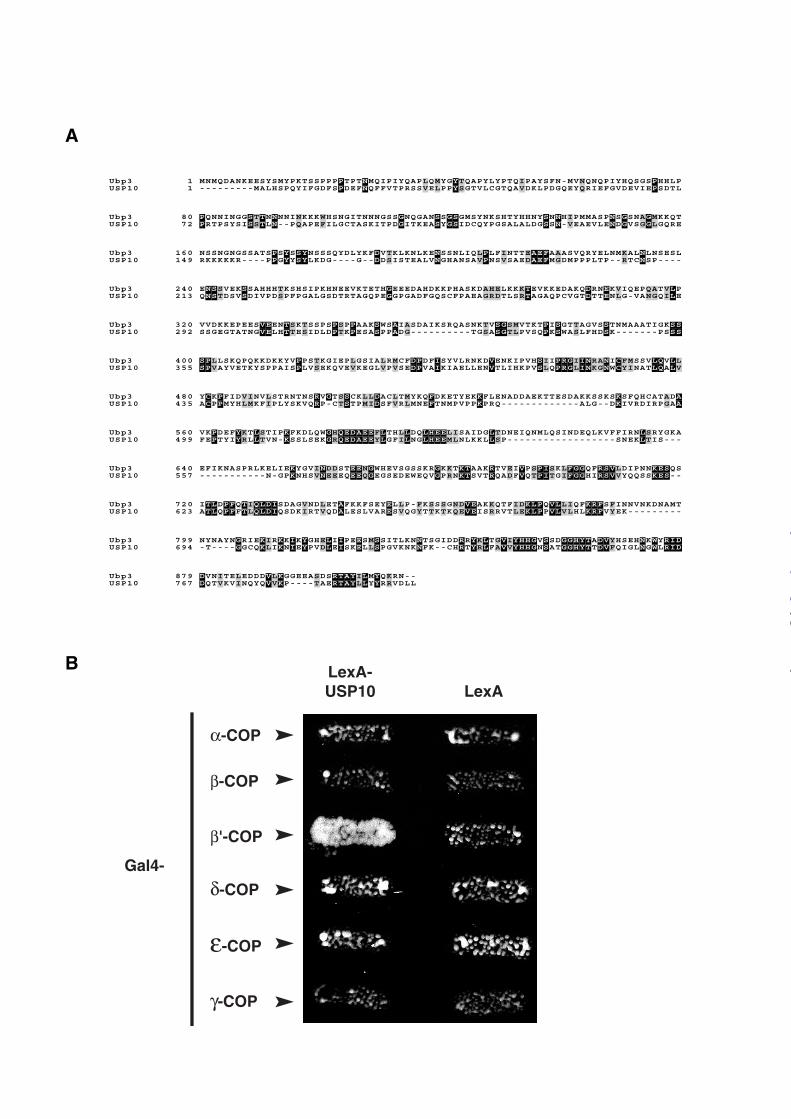
Mammalian β’-COP interacts with USP10. G3BP1, one of the two human proteins similar
to Bre5p, has been previously shown to interact with USP10 (11), a human ubiquitin protease
sharing 46% similarity and 27% identity with Ubp3p mainly concentrated in the catalytic
domain (Figure 4A). Using a 2-hybrid assay, we found that USP10 not only interacts with
G3BP1 but also with G3BP2, the second human homologue of Bre5p (data not shown).
Moreover, similarly to the Ubp3p/Bre5p complex, the interaction between USP10 and G3BP1
or G3BP2 is mediated by the NTF2-like domain of G3BPs (data not shown).
Yeast β’-COP is very similar to bovine β’-COP (43% identity, 64% similarity). To test
whether Ubp3p/Bre5p and USP10/G3BP complexes could exert their de-ubiquitination
activity on similar substrates, we tested whether human USP10 could interact with COPI
subunits. For this purpose, we used a two hybrid approach where the LexA DNA binding
domain alone or fused to USP10 was co-expressed with bovine COPI coatamer subunits fused
to the Gal4 activating domain (Figure 4B). We found that Gal4-β’-COP strongly interacted
with LexA-USP10 whereas the other fusion proteins did not display any significant
interaction using this assay. This result suggests that the yeast Ubp3p/Bre5p and the
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

mammalian USP10/G3BP complexes could be functionally homologous and could regulate
Golgi to ER transport by acting on the function and stability of β’-COP substrate.
In this study, we show that inactivation of the Ubp3p/Bre5p deubiquitination complex
induces an ER retrieval defect likely due to a defect of COPI coat assembly. This partial
defect led to cell lethality when combined with mutation on the ArfGAP Glo3p. Further
analysis indicates that the COPI subunit β’-COP is a substrate of the Ubp3p/Bre5p complex.
Indeed, inactivation of the deubiquitination complex leads to an accumulation of ubiquitinated
β’-COP and a faster proteasome-dependent degradation of the protein. The ability of USP10
to interact with mammalian β’-COP suggest that regulation of Golgi to ER transport by
ubiquitination could also occur in mammals. All together, these data therefore indicate that
the Ubp3p/Bre5p complex is able to regulate retrograde transport from the Golgi apparatus to
the ER by acting on the essential COPI subunit β’-COP. We have previously shown that the
Ubp3p/Bre5p complex can act on anterograde transport between ER and Golgi compartments
by deubiquitinating the COPII protein Sec23p (10). Therefore, the Ubp3p/Bre5p complex
appears to co-regulate anterograde and retrograde transports between ER and Golgi
compartments by deubiquitinating and controlling expression levels of COPII Sec23p and
COPI β’-COP respectively.
Interestingly, Ubp3p has been implicated in pheromone signaling (29), G3BP1 in the
Ras signaling pathway (30) and Bre5p as well as G3BP1 can be phosphorylated (31,32). It is
therefore tempting to speculate that Bre5p/G3BP phosphorylation is regulated by signaling
cascades and controls the activity of the ubiquitination complex. Ubp3/Bre5 complex might
therefore contribute to coordinate and adapt anterograde and retrograde transport routes
between ER and Golgi compartments to the cellular environment and physiological
conditions.
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Acknowledgments
We thank members of the lab as well as N. Belgareh and R. Tsapis-Haguenauer for
stimulatory discussions and critical reading of the manuscript. We are grateful to F.
Letourneur, R. Duden, G. Johnston, F. Wieland and G. Draetta for their generous gift of
antibodies, strains and expression plasmids. This study was funded by grants from the
Association de Recherche contre le Cancer and the Ligue contre le Cancer. M.C. is a fellow
from the Association de Recherche contre le Cancer.
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

References
1. Hershko, A., and Ciechanover, A. (1998) Annu. Rev. Biochem. 67, 425-479
2. Glickman, M. H., and Ciechanover, A. (2002) Physiol. Rev. 82, 373-428.
3. Kim, J. H., Park, K. C., Chung, S. S., Bang, O., and Chung, C. H. (2003) J Biochem
(Tokyo) 134, 9-18
4. Chung, C. H., and Baek, S. H. (1999) Biochem. Biophys. Res. Commun. 266, 633-640.
5. Wilkinson, K. D. (2000) Semin. Cell. Dev. Biol. 11, 141-148.
6. Li, M., Chen, D., Shiloh, A., Luo, J., Nikolaev, A. Y., Qin, J., and Gu, W. (2002)
Nature 416, 648-653.
7. Trompouki, E., Hatzivassiliou, E., Tsichritzis, T., Farmer, H., Ashworth, A., and
Mosialos, G. (2003) Nature 424, 793-796
8. Brummelkamp, T. R., Nijman, S. M., Dirac, A. M., and Bernards, R. (2003) Nature
424, 797-801
9. Kovalenko, A., Chable-Bessia, C., Cantarella, G., Israel, A., Wallach, D., and
Courtois, G. (2003) Nature 424, 801-805
10. Cohen, M., Stutz, F., Belgareh, N., Haguenauer-Tsapis, R., and Dargemont, C. (2003)
Nature Cell Biol. 5, 661-667
11. Soncini, C., Berdo, I., and Draetta, G. (2001) Oncogene 20, 3869-3879.
12. Longtine, M. S., McKenzie, A., 3rd, Demarini, D. J., Shah, N. G., Wach, A., Brachat,
A., Philippsen, P., and Pringle, J. R. (1998) Yeast 14, 953-961.
13. Letourneur, F., Gaynor, E. C., Hennecke, S., Demolliere, C., Duden, R., Emr, S. D.,
Riezman, H., and Cosson, P. (1994) Cell 79, 1199-1207
14. Avaro, S., Belgareh-Touze, N., Sibella-Arguelles, C., Volland, C., and Haguenauer-
Tsapis, R. (2002) Yeast 19, 351-371.
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

15. Faulstich, D., Auerbach, S., Orci, L., Ravazzola, M., Wegchingel, S., Lottspeich, F.,
Stenbeck, G., Harter, C., Wieland, F. T., and Tschochner, H. (1996) J. Cell. Biol. 135,
53-61
16. Orci, L., Stamnes, M., Ravazzola, M., Amherdt, M., Perrelet, A., Sollner, T. H., and
Rothman, J. E. (1997) Cell 90, 335-349
17. Cosson, P., and Letourneur, F. (1994) Science 263, 1629-1631
18. Nickel, W., Brugger, B., and Wieland, F. T. (2002) J Cell Sci 115, 3235-3240
19. Palmer, D. J., Helms, J. B., Beckers, C. J., Orci, L., and Rothman, J. E. (1993) J. Biol.
Chem. 268, 12083-12089
20. Donaldson, J. G., Cassel, D., Kahn, R. A., and Klausner, R. D. (1992) Proc. Natl.
Acad. Sci. 89, 6408-6412
21. Tanigawa, G., Orci, L., Amherdt, M., Ravazzola, M., Helms, J. B., and Rothman, J. E.
(1993) J. Cell. Biol. 123, 1365-1371
22. Reinhard, C., Schweikert, M., Wieland, F. T., and Nickel, W. (2003) Proc. Natl. Acad.
Sci. 100, 8253-8257
23. Poon, P. P., Nothwehr, S. F., Singer, R. A., and Johnston, G. C. (2001) J. Cell. Biol.
155, 1239-1250
24. Dogic, D., de Chassey, B., Pick, E., Cassel, D., Lefkir, Y., Hennecke, S., Cosson, P.,
and Letourneur, F. (1999) Eur J Cell Biol 78, 305-310.
25. Muren, E., Oyen, M., Barmark, G., and Ronne, H. (2001) Yeast 18, 163-172.
26. Lowe, M., and Kreis, T. E. (1996) J. Biol. Chem. 271, 30725-30730
27. Peng, J., Schwartz, D., Elias, J. E., Thoreen, C. C., Cheng, D., Marsischky, G.,
Roelofs, J., Finley, D., and Gygi, S. P. (2003) Nat Biotechnol 21, 921-926
28. Ghislain, M., Udvardy, A., and Mann, C. (1993) Nature 366, 358-362.
29. Wang, Y., and Dohlman, H. G. (2002) J. Biol. Chem. 277, 15766-15772.
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

30. Parker, F., Maurier, F., Delumeau, I., Duchesne, M., Faucher, D., Debussche, L.,
Dugue, A., Schweighoffer, F., and Tocque, B. (1996) Mol. Cell Biol. 16, 2561-2569.
31. Ficarro, S. B., McCleland, M. L., Stukenberg, P. T., Burke, D. J., Ross, M. M.,
Shabanowitz, J., Hunt, D. F., and White, F. M. (2002) Nat Biotechnol 20, 301-305
32. Gallouzi, I. E., Parker, F., Chebli, K., Maurier, F., Labourier, E., Barlat, I., Capony, J.
P., Tocque, B., and Tazi, J. (1998) Mol. Cell Biol. 18, 3956-3965.
33. Holm, C. (1993.) Methods Enzymol. 5, 102-109
34. Vojtek, A. B., Hollenberg, S. M., and Cooper, J. A. (1993) Cell 74, 205-214.
35. Gietz, R. D., and Sugino, A. (1988) Gene 74, 527-534.
36. Mumberg, D., Muller, R., and Funk, M. (1995) Gene 156, 119-122.
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Figure legends
Figure 1: Genetic interaction between BRE5 and GLO3 involves the NTF2-like domain of
Bre5p. bre5∆-glo3 strain transformed with the pCH1122-BRE5 plasmid was transformed A,
with pLAC111 empty vector (Vector), pBRE5 (BRE5) or pGLO3 (GLO3) B, with p415-ADH
empty vector (Vector), with pBRE5 (BRE5), pBRE5-1.400 (BRE5(1-400)), or pBRE5-
146.515 (BRE5 (146-515)). Transformants were subsequently streaked on 5-FOA medium to
induce loss of pCH1122-BRE5.
Figure 2: Bre5p regulates retrograde transport between ER and Golgi compartments. A,
Cells disrupted for BRE5 display an ER retrieval defect. MATa WT (FLY264), ret 1.1
(FLY306) or bre5∆ (FLY264 bre5∆) cells were mated with ΜΑΤα RH311-3D cells and
diploids were selected on appropriate minimal media. B, Purified recombinant fusion proteins
between GST and WBP1 peptides presenting dilysine (GST-WBP1) or diserine motifs (GST-
WBP1-SS) were incubated with lysates from wild type cells (BRE5) or cells disrupted for
BRE5 (bre5∆) and pulled-down with glutathione beads. Bound proteins were revealed by
immunoblotting with antibodies to β’-COP and δ−COP. Loading was controlled using a non
specific band (*).
Figure 3: β’-COP is a substrate of the Ubp3p/Bre5p complex. A, Yeast extracts were
prepared from wild-type cells, bre5∆ and ubp3∆ deletion mutants, and analyzed by SDS-
PAGE and by Western blotting using specific antibodies against the indicated proteins. B, β’-
COP degradation is proteasome-dependent. Wild type CIM3 (YPH 499), cim3-1 (CMY 763),
bre5∆ and cim3-1 bre5∆ strains were grown in YPD at 23°C (A600 0.5) and then shifted to
37°C for 120 minutes (+) in the presence of 100µg /ml cycloheximide. Yeast extracts were
prepared and analyzed by SDS-PAGE and immunoblotting using an anti-β’-COP antibody.
Loading was controlled using a non specific band (not shown). (arrow) non specific. (*)
mono-ubiquitinated β’-COP.
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Figure 4: USP10 interacts with mammalian β’-COP. A, Alignment of S. cerevisiae Ubp3p
and human USP10. Identical or similar amino acids are shown in black or grey background
respectively B, Two hybrid analysis of LexA-USP10 bait versus Gal4-COPI subunit preys.
Preys consisting of Gal4 activation domain fused to bovine α, β, β’,δ , ε and γ-COP subunits
were tested for interaction with USP10 fused to LexA DNA binding domain (LexA-USP10)
or with LexA DNA binding domain (LexA) used as a negative control. Interactions between
proteins were measured by cell growth on media without histidine or by activation of the
LacZ reporter gene in β-galactosidase activity assay (data not shown).
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from
BRE5 GLO3
Vector
bre5∆-glo3
A
B
BRE5
Vector BRE5 (1-400)
BRE5 (146-515)
bre5∆-glo3
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from
WT
ret1.1
bre5∆
BRE5bre
5∆BRE5
bre5∆
GST-KK GST-SS
β'-COP
δ-COP
GST
*
A
B
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from
BRE5bre
5∆
UBP3ubp3∆
Gcs1p
Glo3p
β'-COP*β'-COP
ε-COP
α-COP
δ-COP
γ-COP
β-COP
CIM3 cim3-1 bre5∆bre5∆cim3-1/
β'-COP
β'-COP-Ub
shift at 37°C +- +- +- +-
-82
-175
A
B
*
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from
80 PQNNINGGSTTNNNNINKKKWHSNGITNNNGSSGNQGANSSGSGMSYNKSHTYHHNYSNNHIPMMASPNSGSNAGMKKQT 72 PRTPSYSISSTLN--PQAPEFILGCTASKITPDGITKEASYGSIDCQYPGSALALDGSSN-VEAEVLENDGVSGGLGQRE
160 NSSNGNG SPSYSSYNSSS YSSAT QYDL KFDVTKLKNLKENSSNLIQLPLFINTTEAEFAAASVQRYELNMKALNLNSESL 149 RKKKKKR----PPGYYSYLKDG----G--DDSISTEALVNGHANSAVPNSVSAEDAEFMGDMPPPLTP--RTCNSP----
240 ENSSVEKSSAHHHTKSHSIPKHNEEVKTETHGEEEDAHDKKPHASKDAHELKKKTEVKKEDAKQDRNEKVIQEPQATVLP 213 QNSTDSVSDIVPDSPFPGALGSDTRTAGQPEGGPGADFGQSCFPAEAGRDTLSRTAGAQPCVGTDTTENLG-VANGQILE
320 VVDKKEPEESVEENTSKTSSPSPSPPAAKSWSAIASDAIKSRQASNKTVSGSMVTKTPISGTTAGVSSTNMAAATIGKSS 292 SSGEGTATNGVELHTTESIDLDPTKPESASPPADG----------TGSASGTLPVSQPKSWASLFHDSK-------PSSS
400 SPLLSKQPQKKDKKYVPPSTKGIEPLGSIALRMCFDPDFISYVLRNKDVENKIPVHSIIPRGIINRANICFMSSVLQVLL 355 SPVAYVETKYSPPAISPLVSEKQVEVKEGLVPVSEDPVAIKIAELLENVTLIHKPVSLQPRGLINKGNWCYINATLQALV
480 YCKPFIDVINVLSTRNTNSRVGTSSCKLLDACLTMYKQFDKETYEKKFLE KKSNADDAEKTTESDA SKSKSFQHCATADA 435 ACPPMYHLMKFIPLYSKVQRP-CTSTPMIDSFVRLMNEFTNMPVPPKPRQ-------------ALG--DKIVRDIRPGAA
560 VKPDEFYKTLSTI KFKDLQWGHQEDAEEFLTHLLDQLHEELISAIDGLTD FIRNLSRYNEIQNMLQSINDEQLKVF GKA 499 FEPTYIYRLLTVN-KSSLSEKGRQEDAEEYLGFILNGLHEEMLNLKKLLSP------------------SNEKLTIS---
640 E
P
EFIKNASPRLK LIEKYGVINDDSTEENGWHEVSGSSKRGKKTKTAAKRTVEIVPSPISKLFGGQFRSVLDIPNNKESQS 557 -----------N-GPKNHSVNEEEQEEQGEGSEDEWEQVGPRNKTSVTRQADFVQTPITGIFGGHIRSVVYQQSSKES--
720 ITLDPFQTIQLDISDAGVNDLETAFKKFSEYELLP-FKSSSGNDVEAKKQTFIDKLPQVLLIQFKRFSFIN MNVNKDNA T 623 ATLQPFFTLQLDIQSDKIRTVQDALESLVARESVQGYTTKTKQEVEISRRVTLEKLPPVLVLHLKRFVYEK---------
YPKTSSPPPPTPTNMQIPIYQAPLQMYGYTQAPYLYPTQIPAYSFN-MVNQNQPIYHQSGSPHHLPPQYIFGDFSPDEFNQFFVTPRSSVELPPYSGTVLCGTQAVDKLPDGQEYQRIEFGVDEVIEPSDTL
Ubp3 1 MNMQDANKEESYSMUSP10 1 ---------MALHS
Ubp3 USP10
Ubp3 USP10
Ubp3 USP10
Ubp3 USP10
Ubp3 USP10
Ubp3 USP10
Ubp3 USP10
Ubp3 USP10
Ubp3 USP10
Ubp3 USP10
Ubp3 USP10
799 YN NAYNGRIEKIRKKIKYGHELIIPEESMSSITLKNNTS DDRRYKLTGVIYHHGVSSDGGHYTADVYHSEHNKWYRID 694 -T----GGCQKLIKNIEYPVDLEISKELLSPGVKNKNFK--CHRTYRLFAVVYHHGNSATGGHYTTDVFQIGLNGWLRID
879 DVNITELEDDDVLKG SDSRTAYILMYQKRN-- 767 DQTVKVINQYQVVKP----TAERTAYLLYYRRVD
GI
GEEALL
β'-COP
ε-COP
α-COP
δ-COP
γ-COP
β-COP
LexA-USP10 LexA
Gal4-
B
A
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Strain Genotype Reference
L40FY1679-10AFY10305B (bre5∆)BY10000BY16148 (ubp3∆)RH311-3DFLY264 (wt)
FLY264 (bre5∆)
FLY306 (Ret 1.1)
CH1305CH1462CH1305-bre5∆CH1462-bre5∆bre5∆↑glo3YPH 499CMY 763Cim wt bre5∆Cim 3-1 bre5∆
MATaade2 his3 leu2 trp1 LYS::lexA-HIS3 URA3::lexA-lacZ MATα ura3 trp1 his3 MATα ura3 trp1 his3 YNR051c::kanMX4 MATα ura3 leu2 his3 lys2 MATα ura3 leu2 his3 lys2 YER151c::kanMX4 MATα ura3 leu2 trp1MATa ura3 leu2 his3 ade2/3 trp2 ste2::LEU2 STE2-WBP1::URA3MATa ura3 leu2 his3 ade2/3 trp2 ste2::LEU2 STE2-WBP1::URA3 YNR051c::kanMX6MATa ura3 leu2 his3 ade2/3 trp2 ste2::LEU2 STE2-WBP1::URA3 ret1.1MATa ade2 ade3 leu2 ura3 lys2 can1 MATα ade2 ade3 leu2 ura3 his3 MATa ade2 ade3 leu2 ura3 lys2 can1 YNR051c::kanMX4 MATα ade2 ade3 leu2 ura3 his3 YNR051c::kanMX4 MATα ade2 ade3 leu2 ura3 his3 YNR051c::kanMX4 glo3MATa ura3 leu2 his3 trp1 lys2 ade2 MATα ura3 leu2 his3 cim3-1 MATa ura3 leu2 his3 trp1 lys2 ade2 YNR051c::kanMX6MATα ura3 leu2 his3 cim3-1 YNR051c::kanMX6
Gift from J. CamonisEurofanEurofanEuroscarfEuroscarfGift from F. Letourneur(24)
This study
(24)
(33)(33)(10)(10)This study(28)(28)(10)(10)
Plasmid
pBTM-116pLexA-USP10pAD-Gal4-2.1pGal4-αCOPpGal4-βCOPpGal4-βCOPpGal4-δCOPpGal4-εCOPpGal4-γCOPpCH1122-BRE5YCpLAC111pGLO3p415-ADHpBRE5pBRE5-146.515pBRE5-1.400
2µm TRP1 ADH-LEXA (DNA Binding Domain)2µm TRP1 ADH-LEXA-USP102µm LEU2 ADH-GAL4 (Activation Domain)2µm LEU2 ADH-GAL4-αCOP2µm LEU2 ADH-GAL4-βCOP2µm LEU2 ADH-GAL4-βCOP2µm LEU2 ADH-GAL4-δCOP2µm LEU2 ADH-GAL4-εCOP2µm LEU2 ADH-GAL4-γCOPCEN URA3 ADE3 BRE5(0.5kb 5 and 3flanking sequences)CEN LEU2CEN LEU2 GLO3 (+0.5kb 5 and 3 flanking sequences)CEN LEU2 ADH promCEN LEU2 ADH-BRE5 (codon 1 to 515)CEN LEU2 ADH-BRE5 (codon 146 to 515)CEN LEU2 ADH-BRE5 (codon 1 to 400)
(34)(11)Stratagene(15)(15)(15)(15)(15)(15)(10)(35)This study(36)(10)(10)(10)
Supplementary table: Strains and plasmids used in this study
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Mickael Cohen, Francoise Stutz and Catherine DargemontDe-ubiquitination: A new player in Golgi to ER retrograde transport
published online October 30, 2003J. Biol. Chem.
10.1074/jbc.C300451200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2003/11/12/C300451200.DC1
by guest on September 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from





























