Embed Size (px)
Citation preview

DCM: Advanced issues
Klaas Enno Stephan
Laboratory for Social & Neural Systems Research Institute for Empirical Research in EconomicsUniversity of Zurich
Functional Imaging Laboratory (FIL)Wellcome Trust Centre for NeuroimagingUniversity College London
Methods & Models for fMRI data analysis17 December 2008
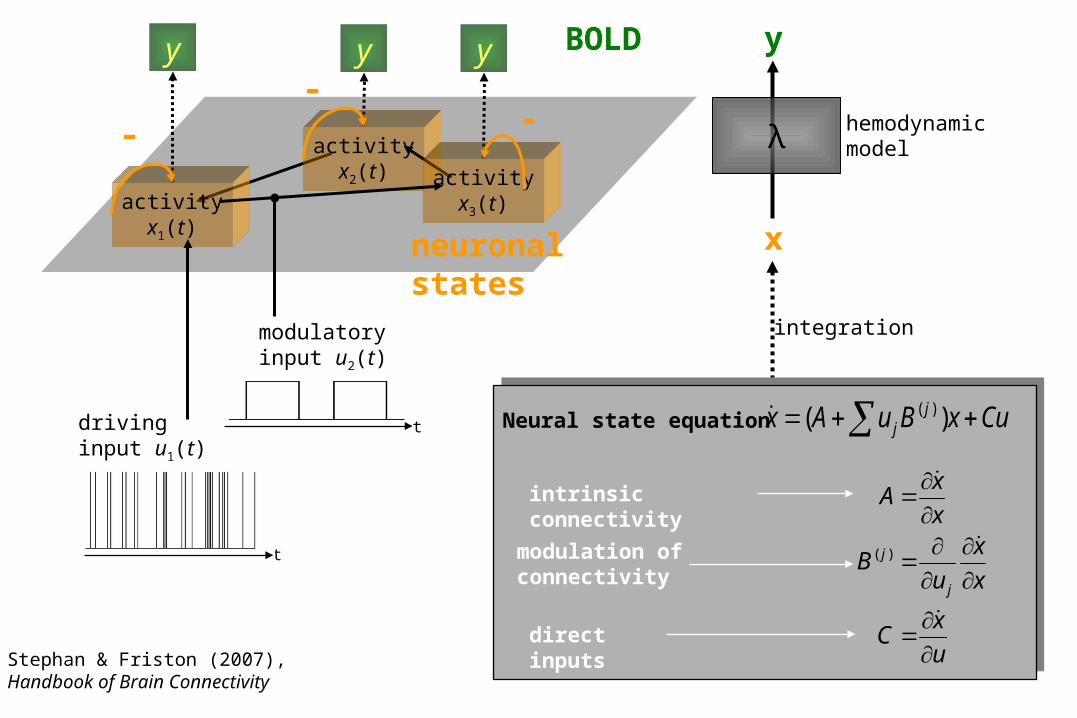
intrinsic connectivity
direct inputs
modulation ofconnectivity
Neural state equation CuxBuAx jj )( )(
u
xC
x
x
uB
x
xA
j
j
)(
hemodynamicmodelλ
x
y
integration
BOLDyyy
activityx1(t)
activityx2(t) activity
x3(t)
neuronalstates
t
drivinginput u1(t)
modulatoryinput u2(t)
t
Stephan & Friston (2007),Handbook of Brain Connectivity
Overview
• Nonlinear DCM for fMRI
• The hemodynamic model in DCM
• Timing errors & sampling accuracy
• Bayesian model selection (BMS)
• DCMs for electrophysiological data

bilinear DCM
CuxDxBuAdt
dx m
i
n
j
jj
ii
1 1
)()(CuxBuA
dt
dx m
i
ii
1
)(
Bilinear state equation:
driving input
modulation
non-linear DCM
driving input
modulation
...)0,(),(2
0
uxux
fu
u
fx
x
fxfuxf
dt
dx
Two-dimensional Taylor series (around x0=0, u0=0):
Nonlinear state equation:
...2
)0,(),(2
2
22
0
x
x
fux
ux
fu
u
fx
x
fxfuxf
dt
dx
0 10 20 30 40 50 60 70 80 90 100
0
0.1
0.2
0.3
0.4
0 10 20 30 40 50 60 70 80 90 100
0
0.2
0.4
0.6
0 10 20 30 40 50 60 70 80 90 100
0
0.1
0.2
0.3
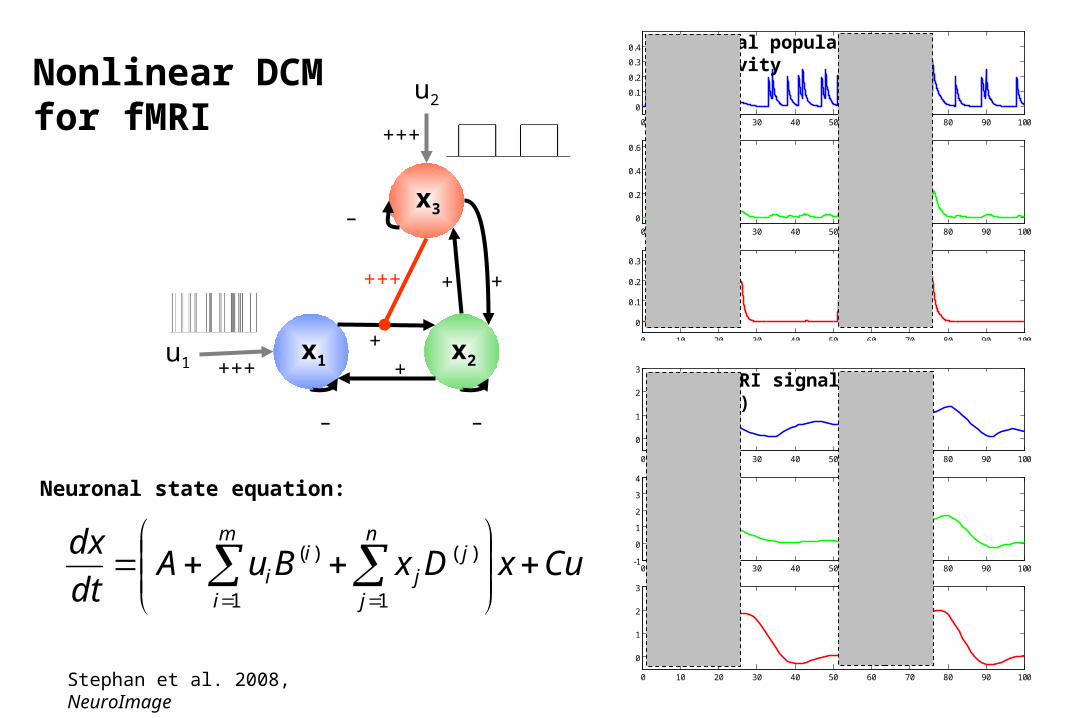
Neural population activity
0 10 20 30 40 50 60 70 80 90 100
0
1
2
3
0 10 20 30 40 50 60 70 80 90 100-1
0
1
2
3
4
0 10 20 30 40 50 60 70 80 90 100
0
1
2
3
fMRI signal change (%)
x1 x2u1
x3
u2
–
– –
++
++++
+++
+++
+
CuxDxBuAdt
dx n
j
jj
m
i
ii
1
)(
1
)(
Neuronal state equation:
Stephan et al. 2008, NeuroImage
Nonlinear DCMfor fMRI

Nonlinear DCM: Attention to motion
V1 IFG
V5
SPC
Motion
Photic
Attention
.82(100%)
.42(100%)
.37(90%)
.69 (100%).47
(100%)
.65 (100%)
.52 (98%)
.56(99%)
Stimuli + Task
250 radially moving dots (4.7 °/s)
Conditions:F – fixation onlyA – motion + attention (“detect changes”)N – motion without attentionS – stationary dots
Previous bilinear DCM
Friston et al. (2003)
Friston et al. (2003):attention modulates backward connections IFG→SPC and SPC→V5.
Q: Is a nonlinear mechanism (gain control) a better explanation of the data?
Büchel & Friston (1997)
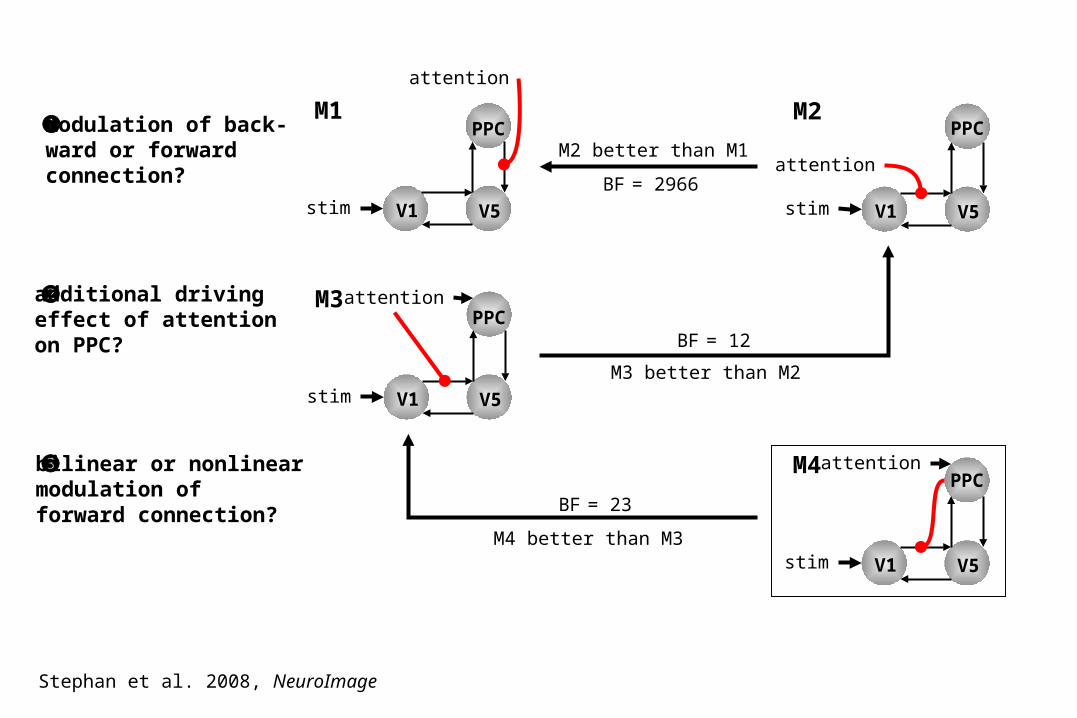
modulation of back-ward or forward connection?
additional drivingeffect of attentionon PPC?
bilinear or nonlinearmodulation offorward connection?
V1 V5stim
PPCM2
attention
V1 V5stim
PPCM1
attention
V1 V5stim
PPCM3attention
V1 V5stim
PPCM4attention
BF = 2966
M2 better than M1
M3 better than M2
BF = 12
M4 better than M3
BF = 23
Stephan et al. 2008, NeuroImage
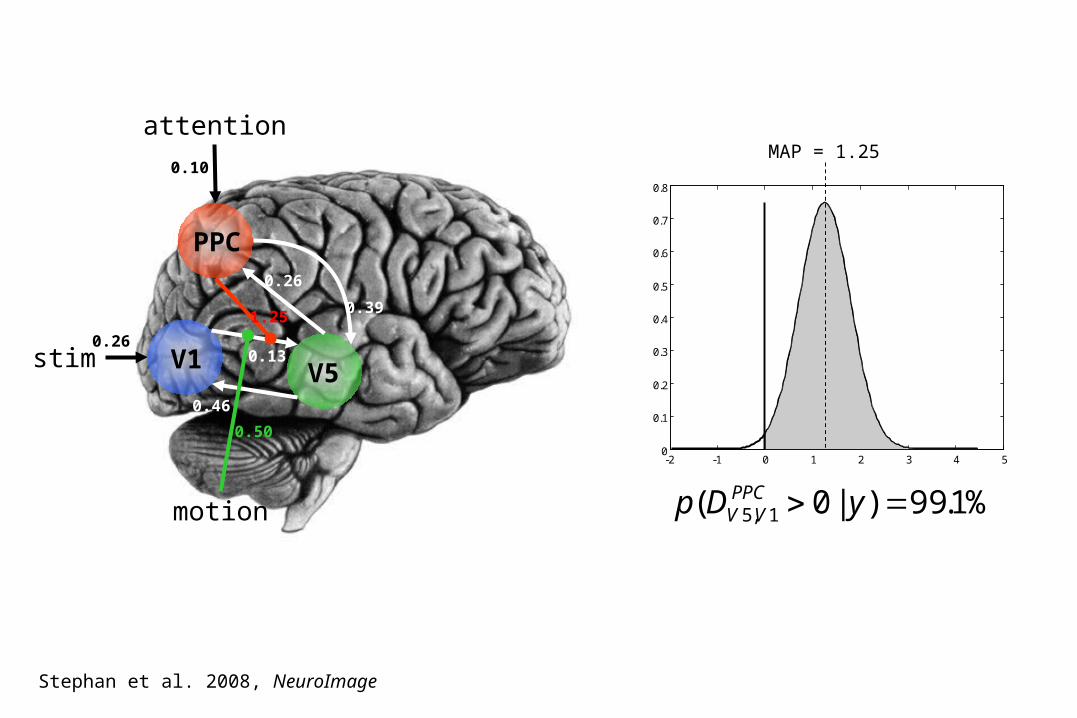
V1 V5stim
PPC
attention
motion
-2 -1 0 1 2 3 4 50
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
%1.99)|0( 1,5 yDp PPCVV
1.25
0.13
0.46
0.39
0.26
0.50
0.26
0.10MAP = 1.25
Stephan et al. 2008, NeuroImage
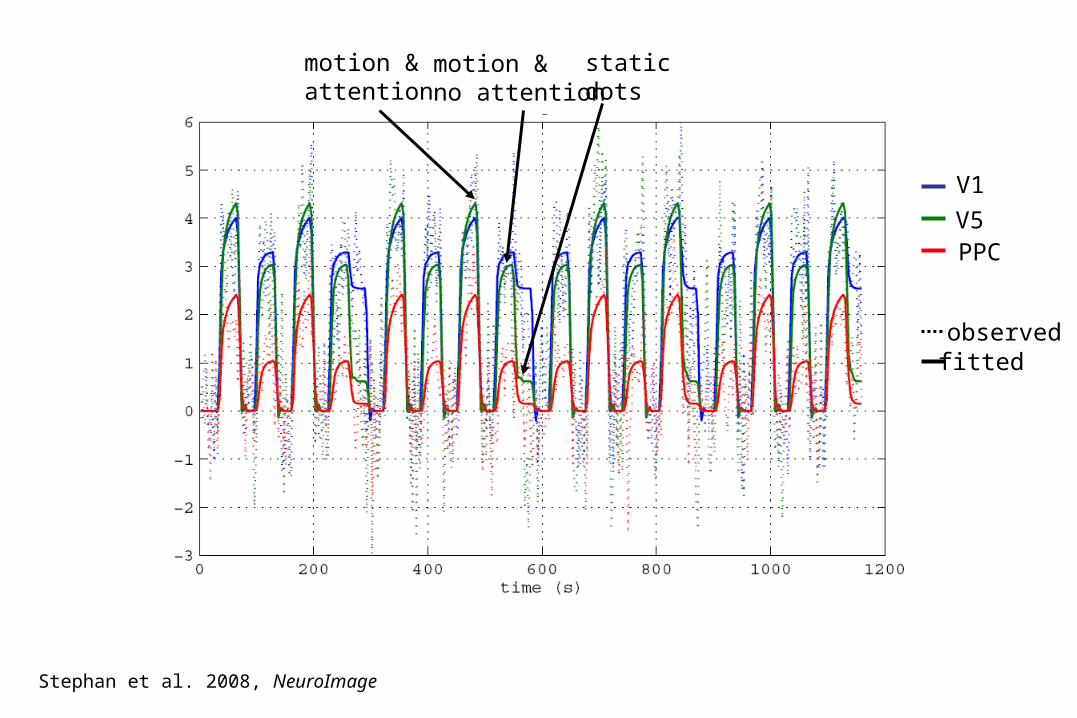
V1
V5PPC
observedfitted
motion &attention
motion &no attention
static dots
Stephan et al. 2008, NeuroImage
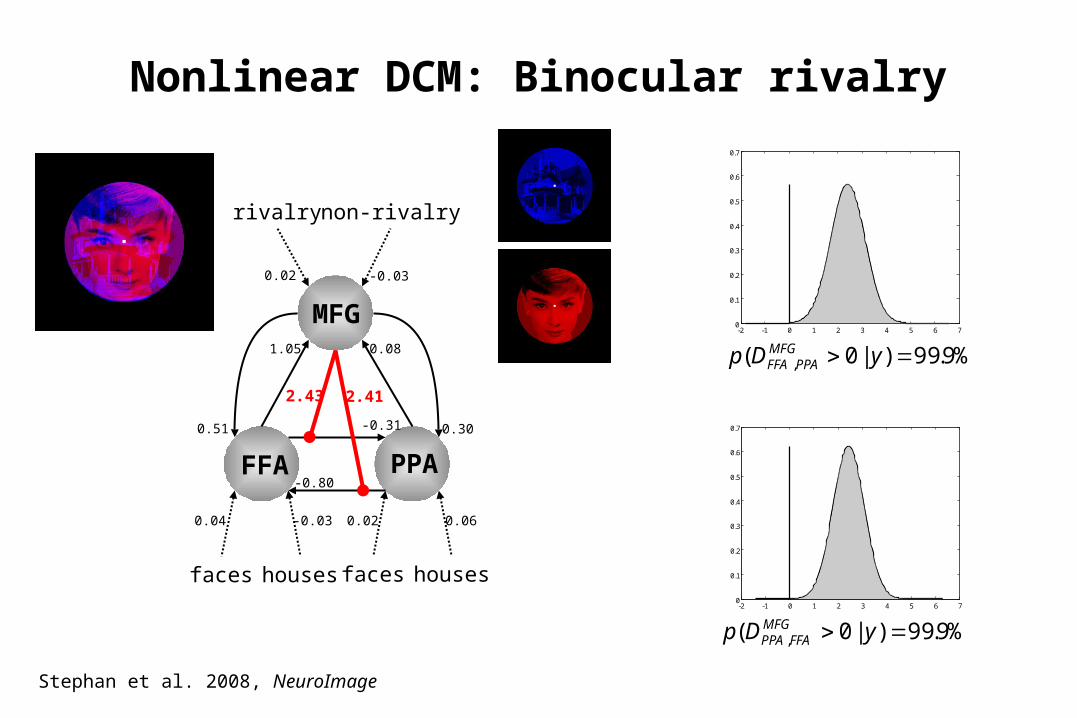
FFA PPA
MFG
-0.80
-0.31
faces houses faces houses
rivalry non-rivalry
1.05 0.08
0.300.51
2.43 2.41
0.04 -0.03 0.02 0.06
0.02 -0.03
-2 -1 0 1 2 3 4 5 6 70
0.1
0.2
0.3
0.4
0.5
0.6
0.7
-2 -1 0 1 2 3 4 5 6 70
0.1
0.2
0.3
0.4
0.5
0.6
0.7
%9.99)|0( , yDp MFGFFAPPA
%9.99)|0( , yDp MFGPPAFFA
Nonlinear DCM: Binocular rivalry
Stephan et al. 2008, NeuroImage
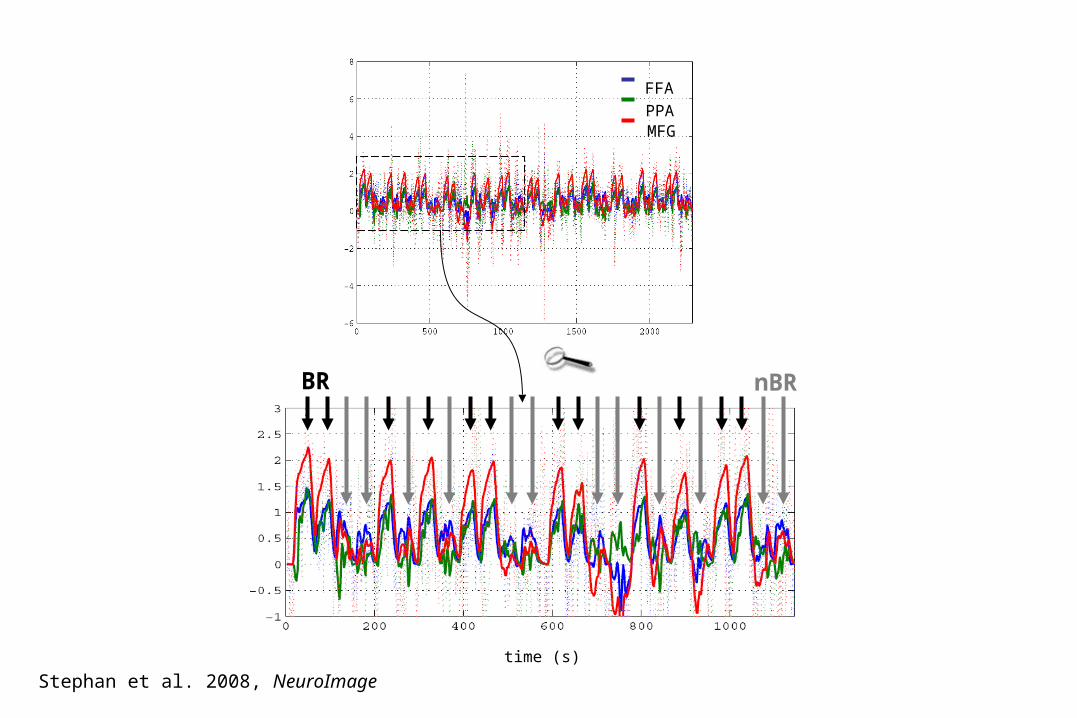
BR nBR
FFA
PPAMFG
time (s)
Stephan et al. 2008, NeuroImage

Overview
• Nonlinear DCM for fMRI
• The hemodynamic model in DCM
• Timing errors & sampling accuracy
• Bayesian model selection (BMS)
• DCMs for electrophysiological data
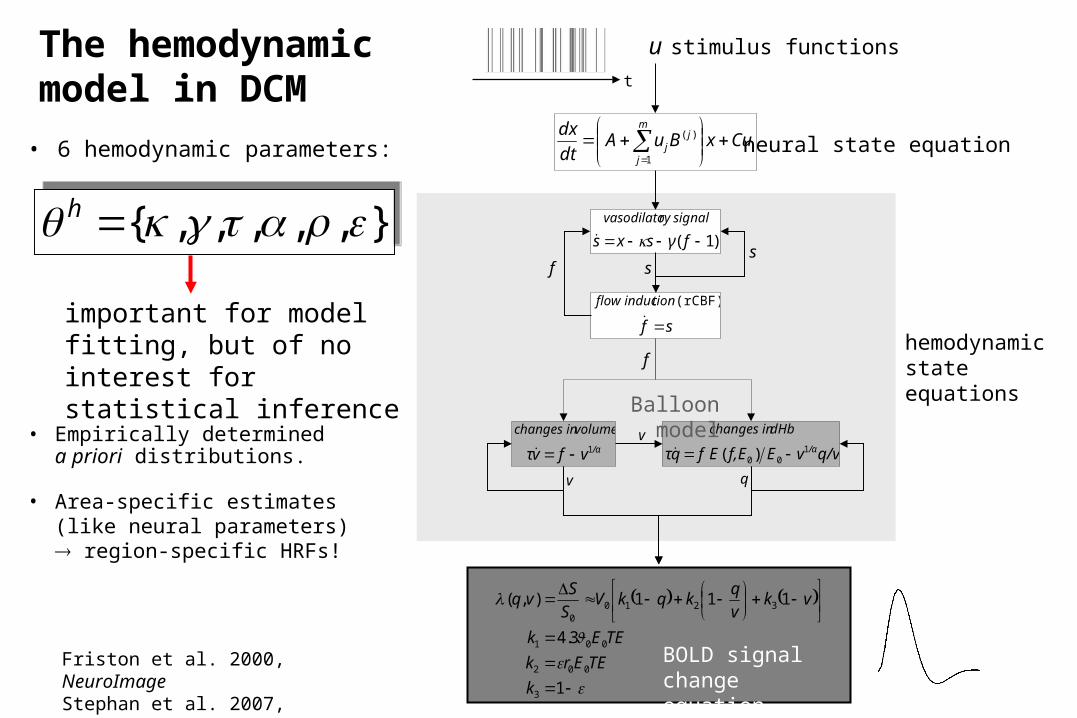
sf
tionflow induc
(rCBF)
s
v
stimulus functions
v
q q/vvEf,EEfqτ /α
dHbchanges in
100 )( /αvfvτ
volumechanges in
1
f
q
)1(
fγsxs
signalryvasodilato
u
s
CuxBuAdt
dx m
j
jj
1
)(
t
neural state equation
1
3.4
111),(
3
002
001
32100
k
TEErk
TEEk
vkv
qkqkV
S
Svq
hemodynamic state equationsf
Balloon model
BOLD signal change equation
},,,,,{ h},,,,,{ h
important for model fitting, but of no interest for statistical inference
• 6 hemodynamic parameters:
• Empirically determineda priori distributions.
• Area-specific estimates (like neural parameters) region-specific HRFs!
The hemodynamic model in DCM
Friston et al. 2000, NeuroImageStephan et al. 2007, NeuroImage
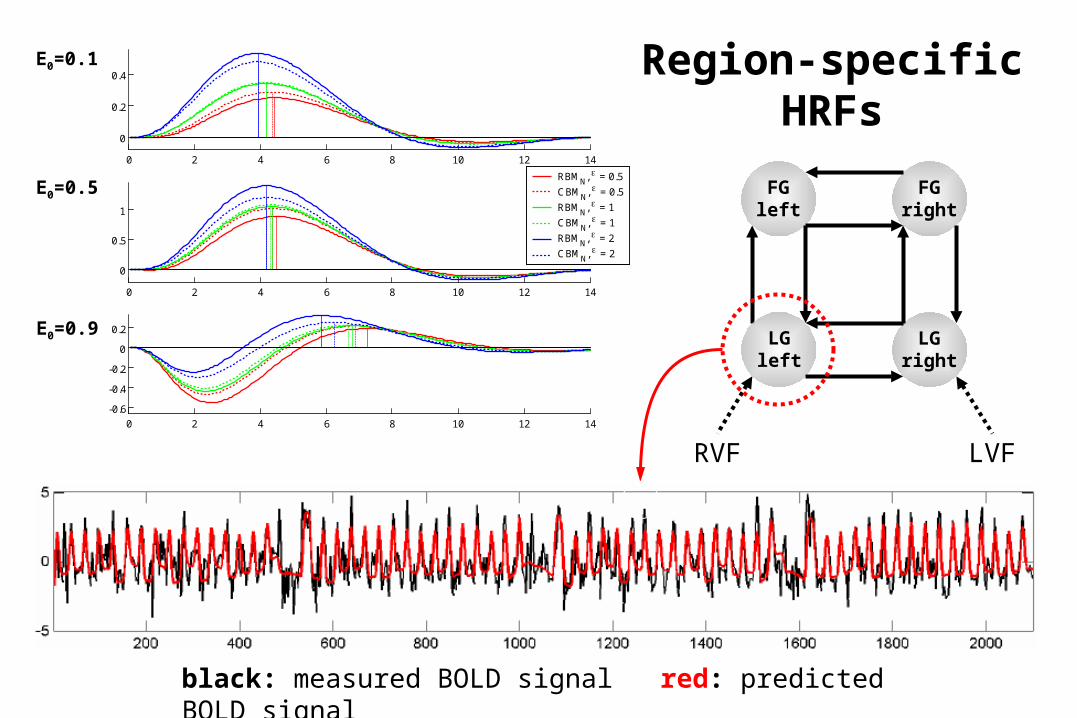
0 2 4 6 8 10 12 14
0
0.2
0.4
0 2 4 6 8 10 12 14
0
0.5
1
0 2 4 6 8 10 12 14
-0.6
-0.4
-0.2
0
0.2
RBMN
, = 0.5
CBMN
, = 0.5
RBMN
, = 1
CBMN
, = 1
RBMN
, = 2
CBMN
, = 2
LGleft
LGright
RVF LVF
FGright
FGleft
black: measured BOLD signal red: predicted BOLD signal
Region-specific HRFs
E0=0.1
E0=0.5
E0=0.9

Recent changes in the hemodynamic model
(Stephan et al. 2007, NeuroImage)
• new output non-linearity, based on new exp. data and mathematical derivations
less problematic to apply DCM to high-field fMRI data
• field-dependency of output coefficients is handled better, e.g. by estimating intra-/extravascular BOLD signal ratio
BMS indicates that new model performs better than original Buxton model
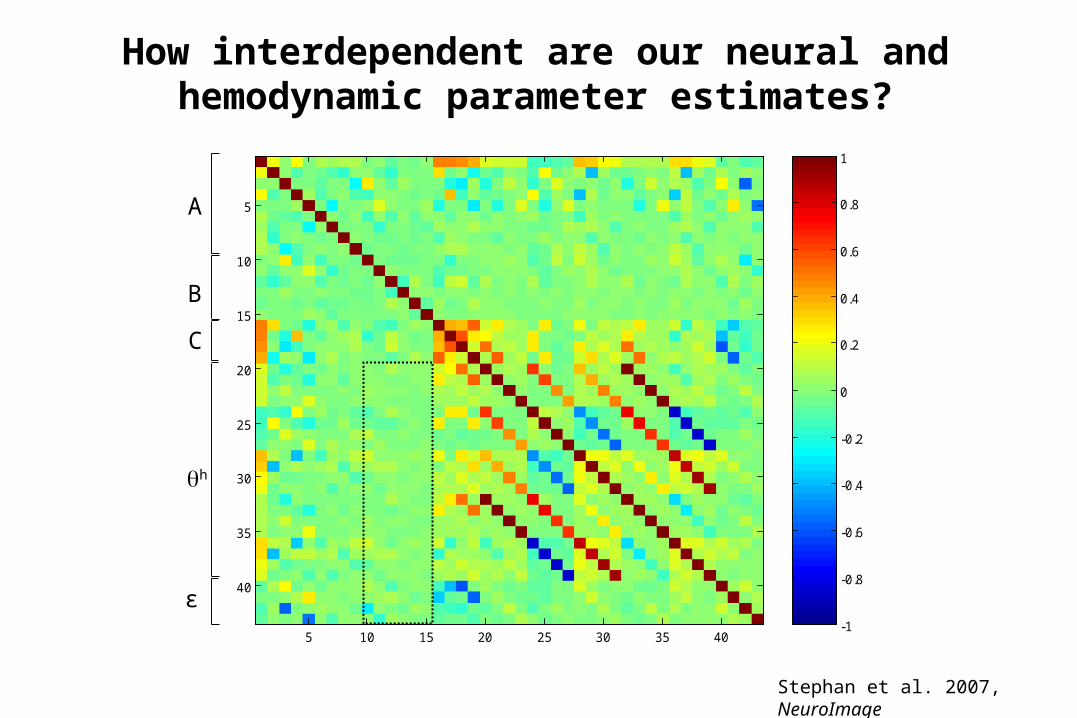
5 10 15 20 25 30 35 40
5
10
15
20
25
30
35
40
-1
-0.8
-0.6
-0.4
-0.2
0
0.2
0.4
0.6
0.8
1
A
B
C
h
ε
How interdependent are our neural and hemodynamic parameter estimates?
Stephan et al. 2007, NeuroImage

Overview
• Nonlinear DCM for fMRI
• The hemodynamic model in DCM
• Timing errors & sampling accuracy
• Bayesian model selection (BMS)
• DCMs for electrophysiological data

Timing problems at long TRs/TAs
• Two potential timing problems in DCM:
1. wrong timing of inputs2. temporal shift between
regional time series because of multi-slice acquisition
• DCM is robust against timing errors up to approx. ± 1 s – compensatory changes of σ and θh
• Possible corrections:– slice-timing in SPM (not for long TAs)– restriction of the model to neighbouring regions– in both cases: adjust temporal reference bin in SPM defaults
(defaults.stats.fmri.t0)• Best solution: Slice-specific sampling within DCM
1
2
slic
e a
cquis
itio
n
visualinput
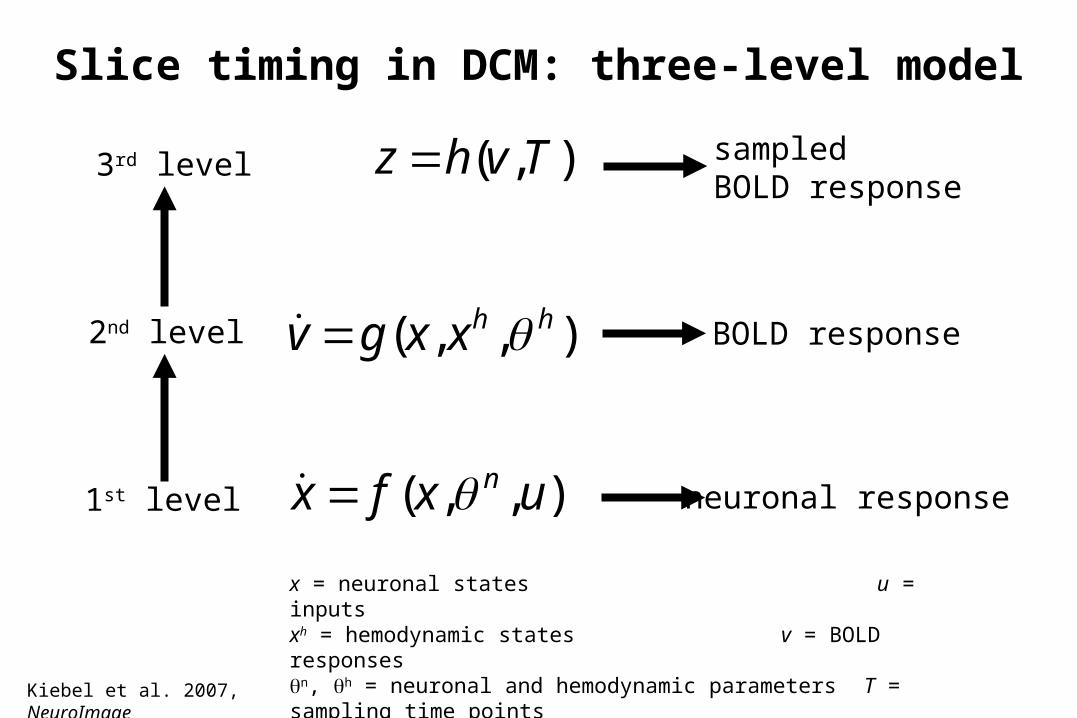
Slice timing in DCM: three-level model
),,( hhxxgv
),( Tvhz
),,( uxfx n
3rd level
2nd level
1st level
sampled BOLD response
BOLD response
neuronal response
x = neuronal states u = inputsxh = hemodynamic states v = BOLD responsesn, h = neuronal and hemodynamic parameters T = sampling time points
Kiebel et al. 2007, NeuroImage
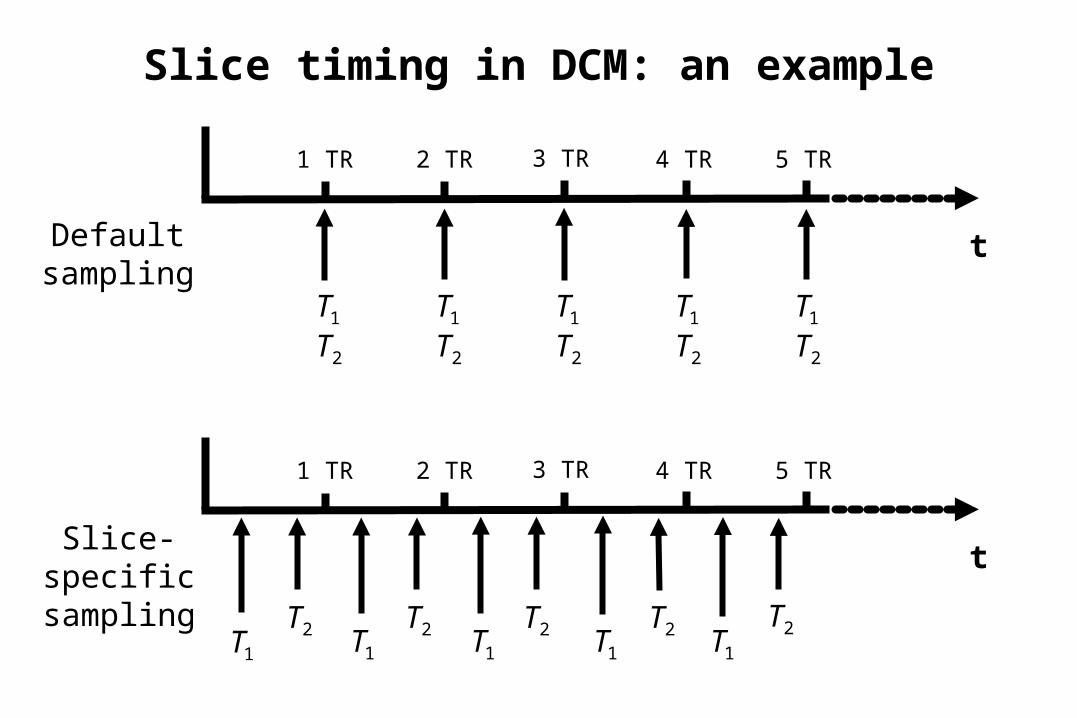
Slice timing in DCM: an example
t
1 TR 2 TR 3 TR 4 TR 5 TR
t
1 TR 2 TR 3 TR 4 TR 5 TR
Defaultsampling
Slice-specific sampling
1T
2T1T
2T1T
2T1T
2T1T
2T
1T 1T 1T 1T 1T2T 2T 2T 2T 2T

Overview
• Nonlinear DCM for fMRI
• The hemodynamic model in DCM
• Timing errors & sampling accuracy
• Bayesian model selection (BMS)
• DCMs for electrophysiological data
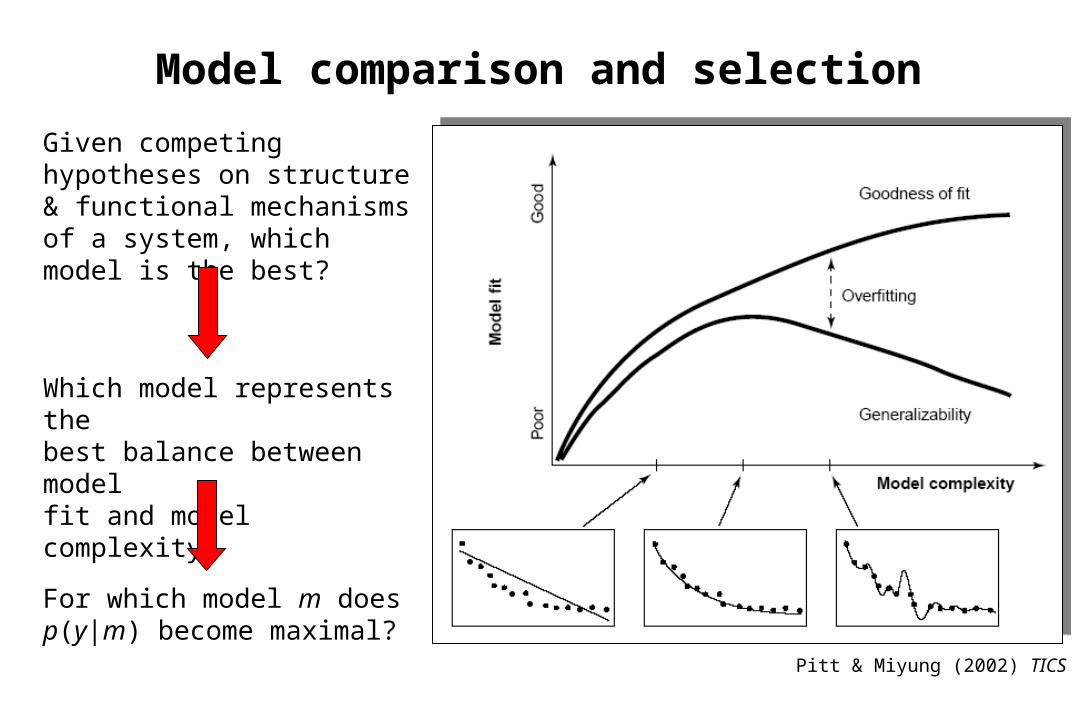
Model comparison and selection
Given competing hypotheses on structure & functional mechanisms of a system, which model is the best?
For which model m does p(y|m) become maximal?
Which model represents thebest balance between model fit and model complexity?
Pitt & Miyung (2002) TICS

dmpmypmyp )|(),|()|( Model evidence:
Bayesian model selection (BMS)
)|(
)|(),|(),|(
myp
mpmypmyp
Bayes’ rule:
accounts for both accuracy and complexity of the model
allows for inference about structure (generalisability)of the model
integral usually not analytically solvable, approximations necessary
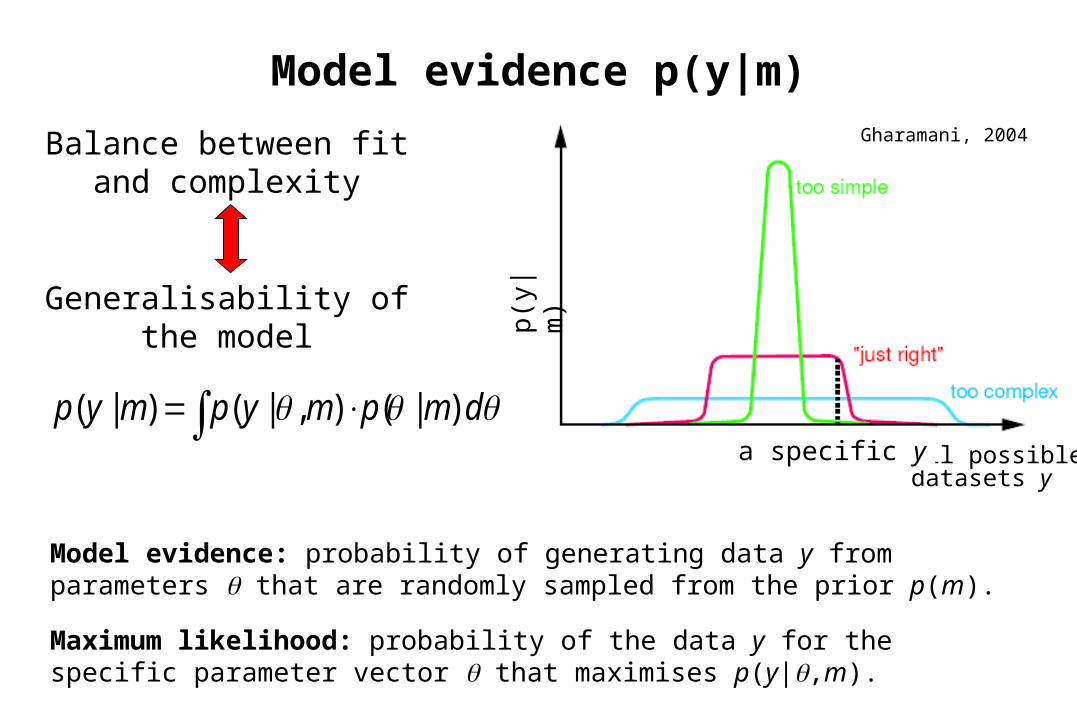
dmpmypmyp )|(),|()|(
Model evidence p(y|m)Gharamani, 2004
p(y
|m
)
all possible datasets y
a specific y
Balance between fit and complexity
Generalisability of the model
Model evidence: probability of generating data y from parameters that are randomly sampled from the prior p(m).
Maximum likelihood: probability of the data y for the specific parameter vector that maximises p(y|,m).

pmypAIC ),|(log
Logarithm is a monotonic function
Maximizing log model evidence= Maximizing model evidence
)(),|(log
)()( )|(log
mcomplexitymyp
mcomplexitymaccuracymyp
In SPM2 & SPM5, interface offers 2 approximations:
Np
mypBIC log2
),|(log
Akaike Information Criterion:
Bayesian Information Criterion:
Log model evidence = balance between fit and complexity
Penny et al. 2004, NeuroImage
Approximations to the model evidence in DCM
No. of parameters
No. ofdata points
AIC favours more complex models,BIC favours simpler models.

Bayes factors
)|(
)|(
2
112 myp
mypB
positive value, [0;[
But: the log evidence is just some number – not very intuitive!
A more intuitive interpretation of model comparisons is made possible by Bayes factors:
To compare two models, we can just compare their log evidences.
B12 p(m1|y) Evidence
1 to 3 50-75 weak
3 to 20 75-95 positive
20 to 150 95-99 strong
150 99 Very strong
Kass & Raftery classification:
Kass & Raftery 1995, J. Am. Stat. Assoc.
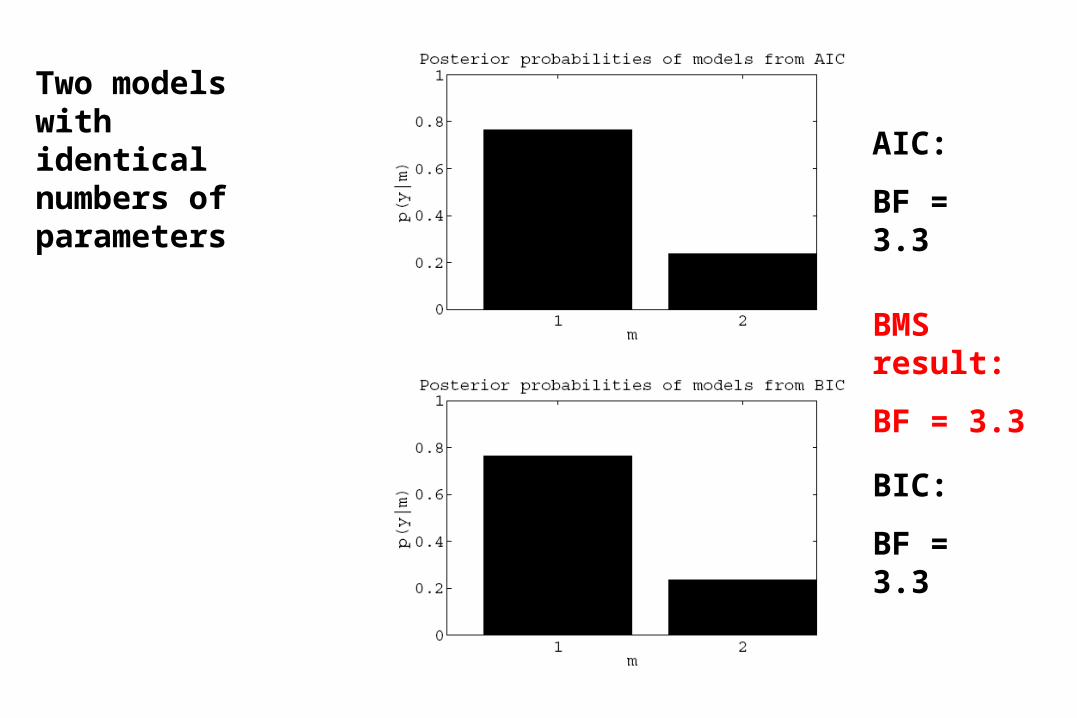
AIC:
BF = 3.3
BIC:
BF = 3.3
BMS result:
BF = 3.3
Two models with identical numbers of parameters
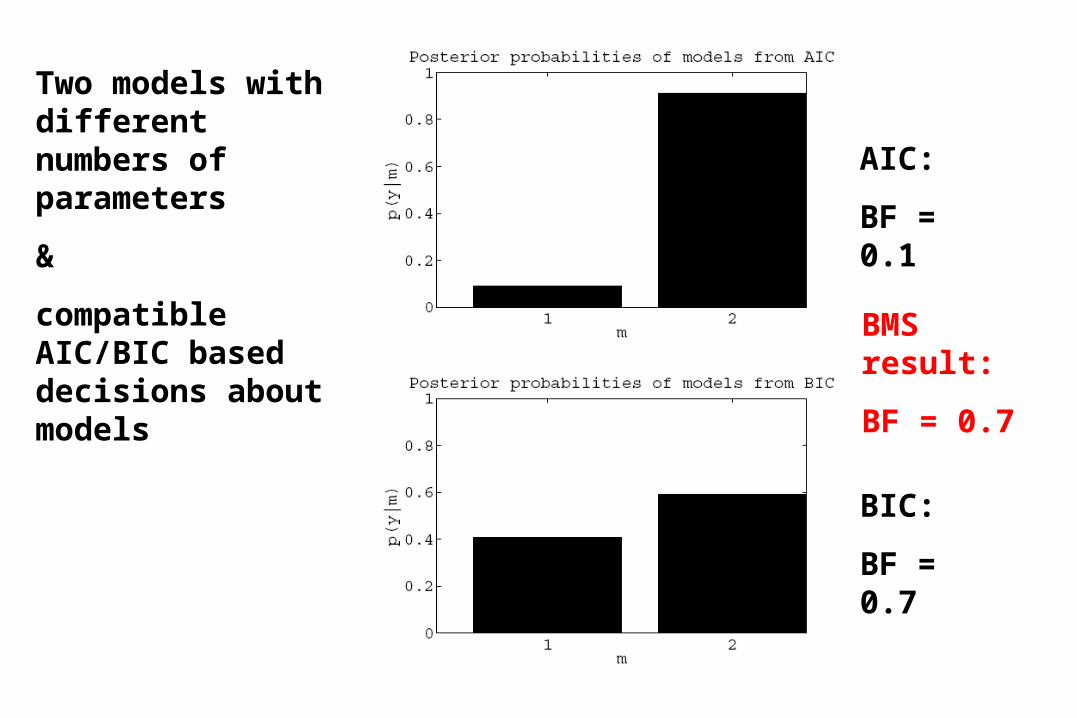
AIC:
BF = 0.1
BIC:
BF = 0.7
BMS result:
BF = 0.7
Two models with different numbers of parameters
&
compatible AIC/BIC based decisions about models
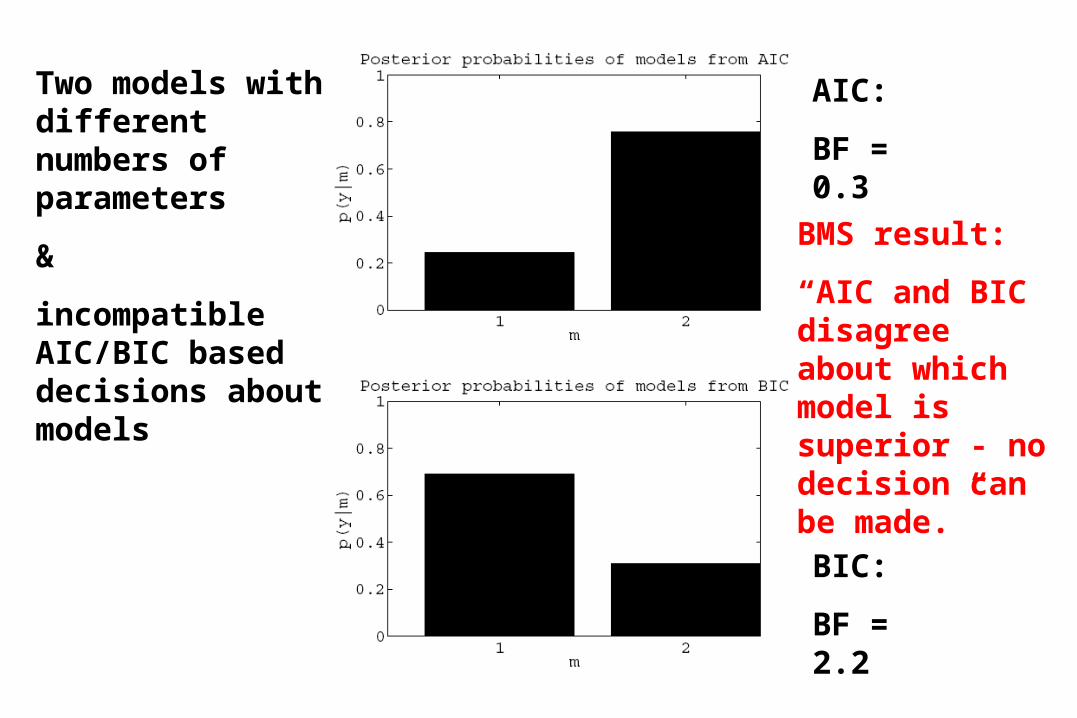
AIC:
BF = 0.3
BIC:
BF = 2.2
BMS result:
“AIC and BIC disagree about which model is superior - no decision can be made.”
Two models with different numbers of parameters
&
incompatible AIC/BIC based decisions about models

The negative free energy approximation
• Under Gaussian assumptions about the posterior (Laplace approximation), the negative free energy F is a lower bound on the log model evidence:
mypqKLF
mypqKLmpqKLmyp
dmpmypmyp
,|,
,|,|,),|(log
)|(),|()|(
mypqKLmypF ,|,)|(

The complexity term in F
• In contrast to AIC & BIC, the complexity term of the negative free energy F accounts for parameter interdependencies.
• The complexity term of F is higher– the more independent the prior parameters ( effective DFs)
– the more dependent the posterior parameters
– the more the posterior mean deviates from the prior mean
• NB: SPM8 only uses F for model selection !
y
Tyy CCC
mpqKL
|1
|| 2
1
2
1
2
1
)|(),(

Selected literature on BMS of DCMs
• Theoretical papers:– Penny et al. (2004) Comparing dynamic causal models. NeuroImage 22:
1157-1172.
– Stephan et al. (2007) Comparing hemodynamic models with DCM. NeuroImage 38: 387-401.
– Stephan et al. Bayesian model selection for group studies. NeuroImage, in revision.
• Application papers:– Grol et al. (2007) Parieto-frontal connectivity during visually-guided
grasping. J. Neurosci. 27: 11877-11887.
– Kumar et al. (2007) Hierarchical processing of auditory objects in humans. PLoS Computat. Biol. 3: e100.
– Smith et al. (2006) Task and content modulate amygdala-hippocampal connectivity in emotional retrieval. Neuron 49: 631-638.
– Stephan et al. (2007) Inter-hemispheric integration of visual processing during task-driven lateralization. J. Neurosci. 27: 3512-3522.

Overview
• Nonlinear DCM for fMRI
• The hemodynamic model in DCM
• Timing errors & sampling accuracy
• Bayesian model selection (BMS)
• DCMs for electrophysiological data
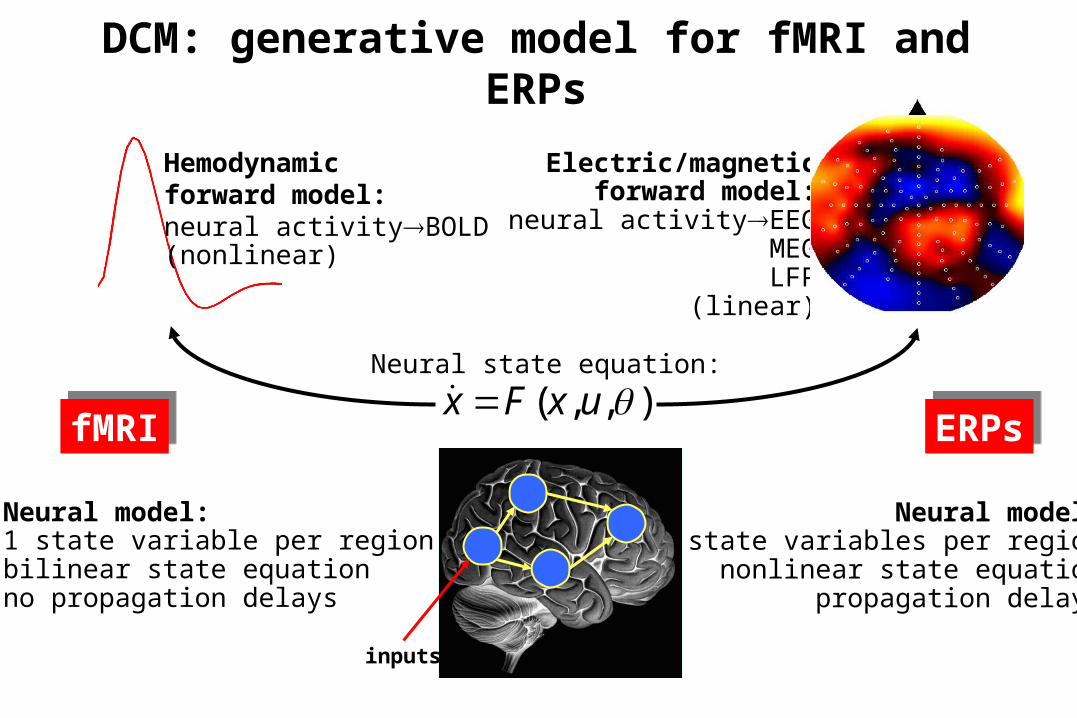
),,( uxFx Neural state equation:
Electric/magneticforward model:
neural activityEEGMEGLFP
(linear)
DCM: generative model for fMRI and ERPs
Neural model:1 state variable per regionbilinear state equationno propagation delays
Neural model:8 state variables per region
nonlinear state equationpropagation delays
fMRIfMRI ERPsERPs
inputs
Hemodynamicforward model:neural activityBOLD(nonlinear)
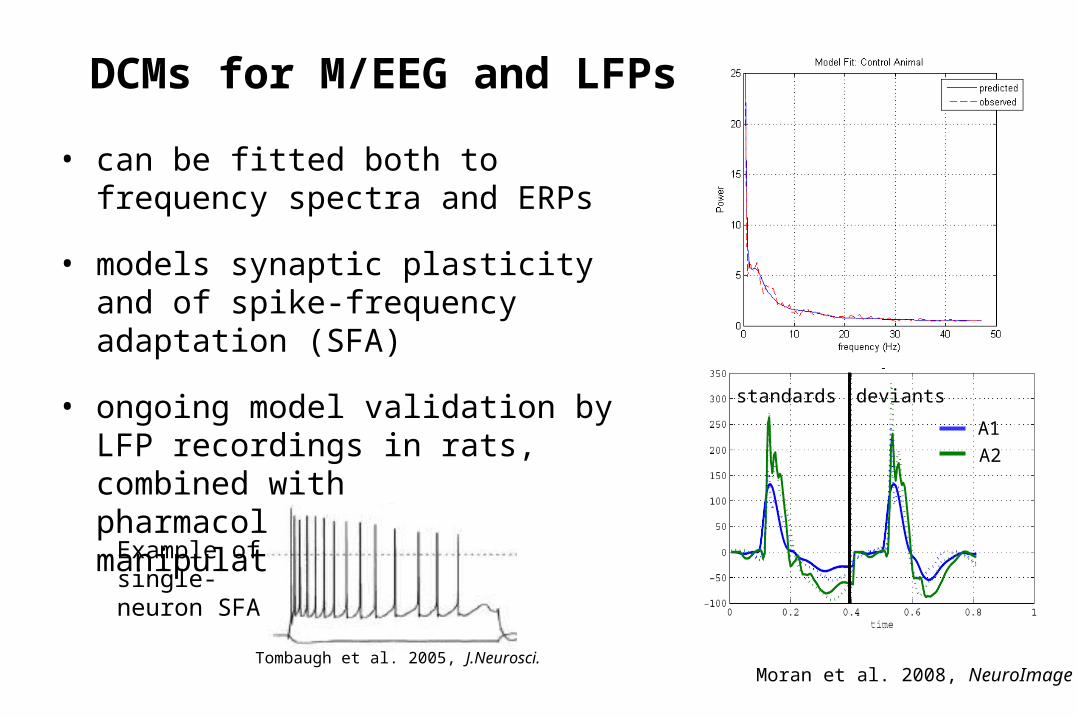
DCMs for M/EEG and LFPs
• can be fitted both to frequency spectra and ERPs
• models synaptic plasticity and of spike-frequency adaptation (SFA)
• ongoing model validation by LFP recordings in rats, combined with pharmacological manipulations
Moran et al. 2008, NeuroImage
standards deviants
A1
A2
Tombaugh et al. 2005, J.Neurosci.
Example of single-neuron SFA
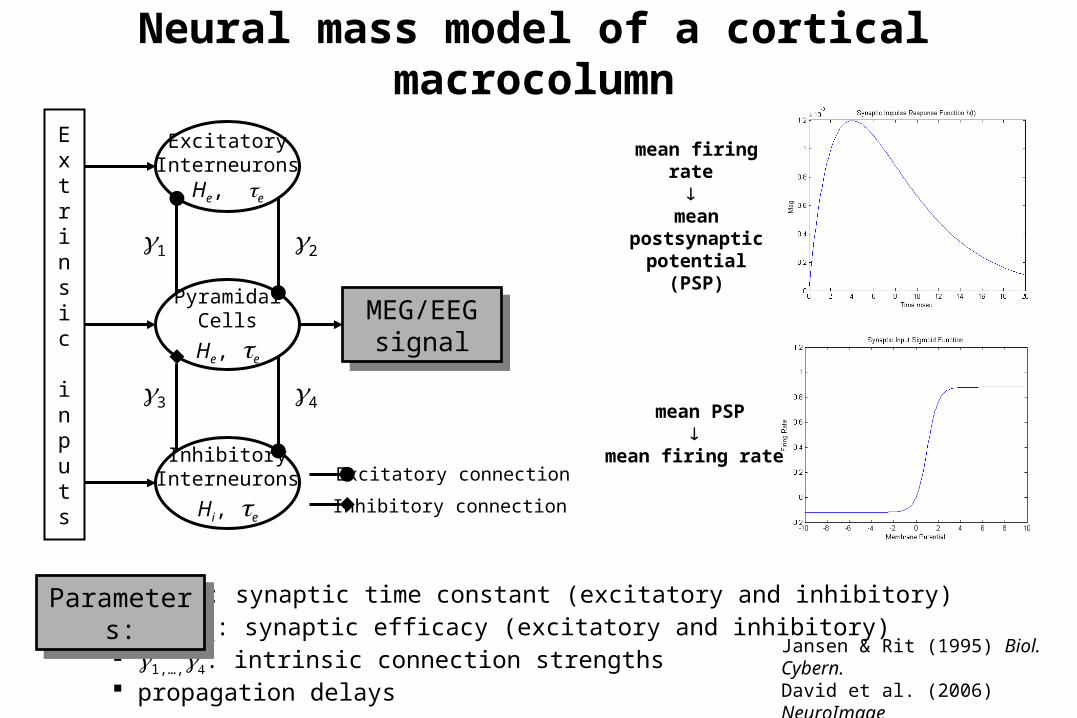
Neural mass model of a cortical macrocolumn
ExcitatoryInterneurons
He, e
PyramidalCells
He, e
InhibitoryInterneurons
Hi, e
Extrinsic inputs
Excitatory connection
Inhibitory connection
e, i : synaptic time constant (excitatory and inhibitory) He, Hi: synaptic efficacy (excitatory and inhibitory) 1,…,: intrinsic connection strengths propagation delays
21
43
MEG/EEGsignal
MEG/EEGsignal
Parameters:
Parameters:
Jansen & Rit (1995) Biol. Cybern.David et al. (2006) NeuroImage
mean firing rate
mean postsynaptic
potential (PSP)
mean PSP
mean firing rate
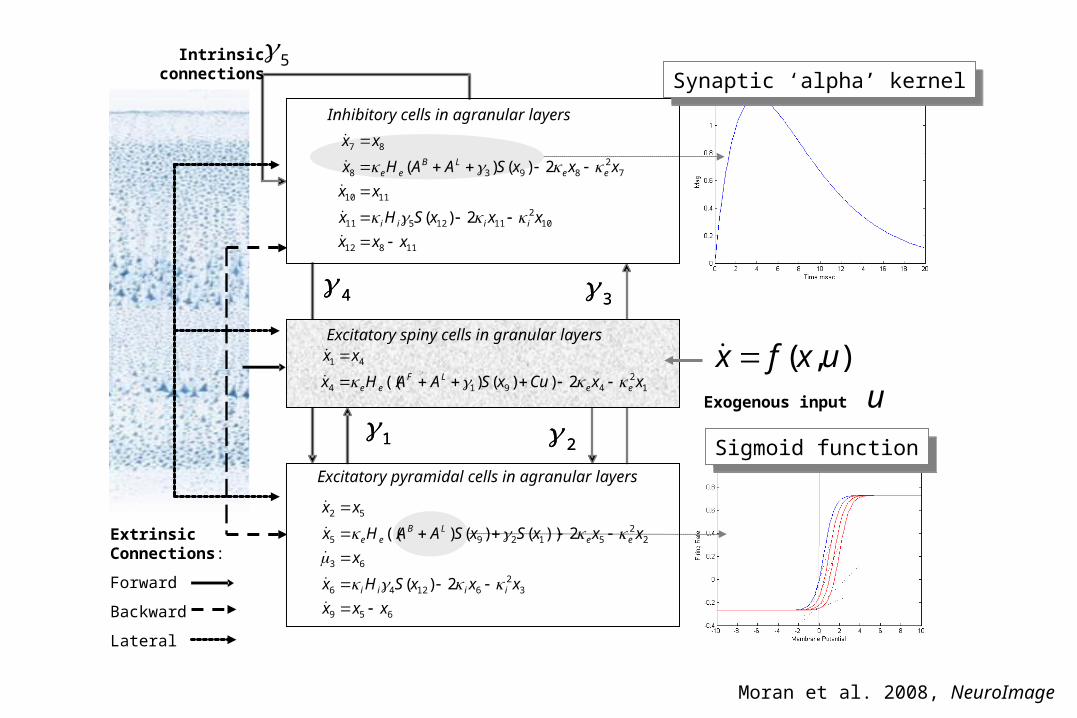
4 3
1 2
12
4914
41
2))(( xxuaxsHx
xx
eeee
Excitatory spiny cells in granular layers
Exogenous input u
4 3
1 2
Intrinsicconnections
5
Excitatory spiny cells in granular layers
Excitatory pyramidal cells in agranular layers
Inhibitory cells in agranular layers
),( uxfx
11812
102
1112511
1110
72
8938
87
2)(
2)()(
xxx
xxxSHx
xx
xxxSAAHx
xx
iiii
eeLB
ee
12
4914
41
2))()(( xxCuxSAAHx
xx
eeLF
ee
Synaptic ‘alpha’ kernelSynaptic ‘alpha’ kernel
Sigmoid functionSigmoid function
659
32
61246
63
22
51295
52
2)(
2))()()((
xxx
xxxSHx
x
xxxSxSAAHx
xx
iiii
eeLB
ee
Extrinsic
Connections:
Forward
Backward
Lateral
Moran et al. 2008, NeuroImage
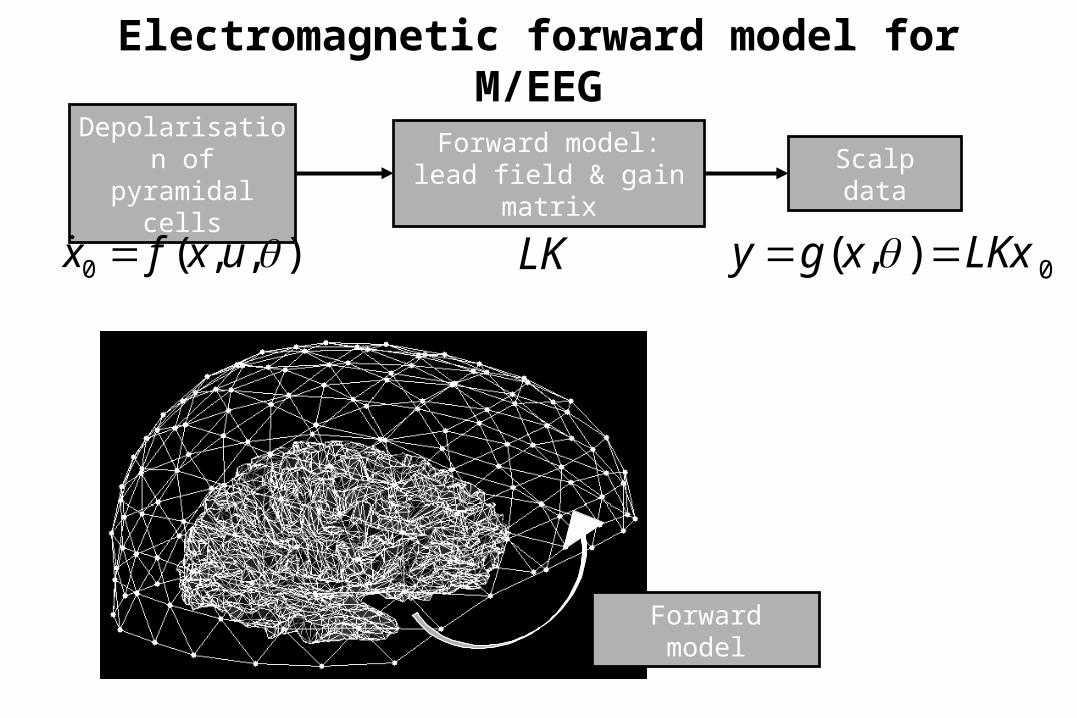
Electromagnetic forward model for M/EEG
Depolarisation of
pyramidal cells
Forward model:lead field & gain
matrixScalp data
),,(0 uxfx LK 0),( LKxxgy
Forward model

Thank you





























