Embed Size (px)
Citation preview

Archs oral Bid. Vol. 36, No. 10, pp. 747-758, 1991 0003-9969/91 $3.00 + 0.00 Printed in Great Britain. All rights reserved Copyright 0 1991 Pergamon Press plc
CYTOKERATIN PATTERNS OF HUMAN ORAL MUCOSAE IN HISTIOTYPIC CULTURE
A. H. M. SHABANA, J. P. OUHAYOUN, M. H. SAWAF and N. FOREST
Laboratoin: de Biologie-Odontologie, lnstitut Biomedical des Cordeliers, Escalier E, 2tme &age, 15 rue de 1’Ecole de Mbdecine, 75006 Paris, France
(Received 10 December 1990; accepted 4 April 1991)
Summary-In a three-dimensional culture model, oral epithelial differentiation was investigated ultra- structurally and biochemically for cytokeratin expression. Epithelia from the hard palate, gingiva and alveolar mucosa grown on freely floating collagen lattices populated with fibroblasts from homotypic origins, and fed with medium containing 10% delipidized fetal calf serum for 21 days before analysis, stratified and differentiated to basal cuboidal cells, polyhydral spinous cells and elongated superficial cells. The epithelium of palatal origin had non-nucleated superficial cells resembling orthokeratinized cells. The upper spinous cells had keratohyalin-like granules. The corresponding cells of gingival and alveolar mucosal origins retained their nuclei and had smaller numbers of keratohyalin-like granules. Basal cell keratins (CK 5 and 14) and those of hyperproliferation (CK 6 and 16) were consistently found in all epithelia. Furthermore, simple epithelial keratins (CK 18 and 19) were variably expressed by cells from different oral origins. In epithelial cells from the alveolar mucosa, CK 13 and 19 formed major bands, which correlates with their expression in uivo. In contrast, these polypeptides were either absent or formed minor bands in extracts of gingival and hard palatal cells. Although in small quantities, keratins of terminal differentiation (CK 1, 2, 10 and 11) were detected in gels prepared from palatal epithelia. This expression correlates with the higher morphological differentiation of these cells in this model. The model is of interest for studies of epithelial differentiation, as the differentiation markers of keratinized epithelia (CK I and 10) were expressed by cells from palatal origin, and those of non-keratinized epithelia (CK 4, 13 and 19) were prominent in cells from alveolar mucosal origin.
Key words: cq tokeratins, oral epithelia, oral mucosa, differentiation, culture.
INTRODUCTION
In man, the oral mucosa is lined by three distinct types of stratified squamous epithelia: a keratinized epithelium in the gingiva and hard palate, a non-ker- atinized epithelium in the alveolar mucosa, the cheeks and the sublingual region, and a complex epithelium made of both types on the dorsal surface of the tongue (Sawaf et al, 1990). These epithelia can be easily distinguished microscopically. However, subtle changes in epithelial differentiation during develop- ment and in pathological conditions may be detected only by more advanced techniques. Cytokeratins, the intermediate filamems of the epithelial cell, constitute a family of at least l!) different polypeptides (Cooper, Schermer and Sun, 1985; Franke et al., 1981; Moll, Franke and Schiller, 1982). Moll ef al. (1982) num- bered them according to their molecular weight and isoelectric mobility. Different cytokeratin subsets were shown to characterize different epithelia. In keratinized epithelia of the gingiva and hard palate, cytokeratin polypeptides (CK) 1,2,4, 5,6, 10, 11, 13, 14, 16 and 17 were expressed. In non-keratinized epithelia of the alveolar mucosa and cheeks, CK 4, 5, 6, 13, 14, 17 and 19 were found (Ouhayoun et al., 1985; Clausen et al.., 1986a, b; Morgan et al., 1987; Juhl, Reibel and Stoltze, 1989; Shabana et al., 1989).
Abbreviations: EMEM.. Eagle’s minimal essential medium; FCS, fetal calf serum; FITC, fluoroscein isothiocyanate.
Cytokeratins of simple epithelia (7, 8, 18 and 19) could be identified in salivary glands (Born et al., 1987) and odontogenic epithelia (Gao et al., 1988; Kasper et al., 1989; Pelissier et al., 1990).
The factors involved in modulating epithelial differentiation are not well understood. However, several mesenchymal elements are known to influence epithelial differentiation, particularly during embry- onic life (Hay, 1977; Slavkin, 1980). In vitro, several soluble elements such as calcium (Breitkreutz et al., 1984; Sacks et al., 1985; Watt, 1984), hydrocortisone, cholera toxin (Ponec et al., 1988; Southgate et al., 1987) and vitamin A (Green and Watt, 1982; Kopan, Traska and Fuchs, 1984; Brown, Gray and Bernstein, 1985) modulate epithelial differentiation. Most of these studies have been made on epithelial cells from skin. Epidermal cells in culture lacked the granular and keratinized layers of the normal epidermis; this was associated with loss of the terminal differen- tiation cytokeratins (CK 1, 2, 10 and 11) (Fuchs and Green, 1981; Kim, Schwartz and Fuchs, 1984; Sun et al., 1984; Tyner and Fuchs, 1986). However, these cytokeratins were re-expressed when vitamin A was removed from the serum and when the epithelium was grown at the air-liquid interface (Asselineau et al., 1985, 1989; Fuchs and Green, 1981; Regnier et al., 1986). In order to gain further insight into the different differentiation programmes and their regula- tors in keratinized and non-keratinized oral mucosal epithelia an in vitro model is needed. So far, detailed
747

748 A. H. M. SHABANA et al.
description of cytokeratin profiles of different human oral epithelia grown in culture is lacking. We now report on cytokeratin expression using biochemical and immunohistochemical methods and relate the findings to the morphological pattern of differen- tiation. The three-dimensional lattice made of colla- gen and populated with fibroblasts was chosen as a model for studying epithelial differentiation because there are several descriptions of better attachment, growth and differentiation of epithelial cells on colla- gen than on plastic (Karasek and Charlton, 1971; Clark, Folkvord and Mertz, 1985; Kubo et al., 1987). Furthermore, this model appears of interest for studies on cell-cell and cell-matrix interactions. Homotypic fibroblasts were incorporated in collagen lattices in order to provide, as far as possible, con- ditions of growth and differentiation resembling those in vivo. We have sought to characterize the cytokeratin profiles of three oral mucosal epithelial cells, hard palatal, gingival and alveolar mucosal, cultured on free-floating collagen lattices.
MATERIALS AND METHODS
Preparation of histiotypic cultures
Heathy tissues from gingiva, hard palate and alve- olar mucosa were collected from five young patients (20-40 yr) during the extraction of impacted canine and premolar teeth. All specimens were prepared for culture by being washed in EMEM with Earle’s salts (Boeringher, Meyland, France), 10% FCS (Flow, Puteaux, France), 2.5 pg/ml amphotericin, 100 iu/ml penicillin and 100 ng/ml streptomycin (GIBCO, Cergy, France). The issues were cut into 1 mm3 fragments with a punch. Primary fibroblast cultures were initiated in 60-mm tissue-culture dishes (Becton Dickinson, Grenoble, France) containing 5 ml of medium. The cultures were incubated in humid air containing 5% CO2 at 37°C and fed twice a week. The tissue explants were removed after 14 days, leaving primary growths of fibroblasts and epithelial cells. After confluency, fibroblasts were harvested and sub- cultured in a calcium- and magnesium-free solution of 0.02% EDTA and 0.25% trypsin (GIBCO) at 37°C for 10 min. Second-passage fibroblasts were exam- ined for mycoplasms before reconstitution of the mucosae in culture.
Collagen type 1 was prepared from rat’s tail tendon by 0.1% acetic acid extraction; the collagen lattices were prepared as described previously (Bell, Ivarsson and Merrill, 1979; Bell et al., 1981). The three-dimen- sional lattices were prepared in ice cold, 25-m] glass flasks. For each lattice, 2.3 ml of concentrated ( x 1.7) EMEM containing the above antibiotics, 0.45 ml FCS, 1.50 ml of 2 mg/ml collagen solution, 0.25 ml of 1 M NaOH and 0.50ml fibroblast suspension (5 x lo4 cells/ml) were mixed, and reversed in 60-mm Petri dishes. Gel polymerization was effected at 37°C for 2 h. Further biopsy fragments were prepared as described above and implanted, one fragment per gel, with the epithelial side at the surface of the collagen gel [Saiag, Coulomb and Lebreton. 1985). After 24 h, the exudated medium was exchanged for complete medium containing IO ng/ml epidermal growth factor (GIBCO), 10% delipidized FCS [filtered with 0.2 pm millipore filters to remove fat globules (Basset-SCguin
et al., 1990)] and the temperature was lowered to 36°C (Jensen and Therkelsen, 1981). The gels were then freed from the Petri dishes and left floating freely in the medium. The cultures were fed twice a week with this medium for 21 days. By this technique, hard palate1 gingival and alveolar mucosae were reconsti- tuted using homotypic epithelia and fibroblasts from each site.
Biochemistry and immunohistochemistry
Fibroblasts contracted the collagen gel from 53 mm in diameter to about 10 mm in 21 days. The tissues became dense and could be further treated as conventional biopsy material. The tissues were either frozen in liquid nitrogen-cooled isopentane for im- munohistochemistry and cytokeratin extraction or fixed for electron microscopy. Great care was taken to cut out the explant before biochemical and immunohistochemical analyses of cytokeratins. The degenerating explant was easy to identify by its yellowish colour, and was cut while frozen. Six-pm frozen sections were used for in situ localization of individual or small groups of cytokeratins using monoclonal antibodies: EE21.6 antiCK I, 2, 10, and 11 (gift from Dr Serre, Toulouse, France), K, 8.60 anti-CK 10 and 11 [Biomaker, Rehovot, Israel (Huszar et al., 1986)], K, 19.1 antiCK 19 [Progen, Heidelberg, Fed. Rep. Germany (Karsten et al., 1985)], 6BlO anti-CK 4 [gift from Professor Franke, Heidelberg, Fed. Rep. Germany (van Muijen et al., 1986)], KB-37 anti-basal cell cytokeratins (Sigma, La Verpilliire, France), lC7 anti-CK 13 [gift from Pro- fessor Franke (van Muijen et al., 1986)], K, 13.1 antiCK 13 and 14 [Progen (Moll et al., 1982)], K, 18.174 anti-CK 18 [Progen (Moll et al., 1988)]. The reaction site was detected by using goat anti-mouse IgG (L + H) FITC-labelled antibody (Kirkegaard & Perry Inc., Maryland, MD, U.S.A.). The technique was described thoroughly by Shabana et al. (1989).
Cytokeratin was extracted by the method of Acht- statter et al. (1986) to remove soluble proteins in high- and low-salt buffers. Fifty frozen thick (20 pm) sections were collected in Eppendorf tubes for cyto- keratin extraction. The material was homogenized and agitated in 1 ml of first, high-salt buffer, second, low-salt buffer, then washed in phosphate-buffered saline. The pH of the buffers were all adjusted to 7.4. The tissue was homogenized with a clean, stainless- steel spatula and mechanical agitation was for 5 min, with the exception of the high-salt buffer, which was agitated for 1 h. After each buffer, pellets were collected by centrifugation at 1200 g for 5 min. Two- dimensional gel electrophoresis was done with the non-equilibrium pH gradient for the first dimension as described by O’Farrell, Goodman and O’Farrell (1977) and modified by Franke et al. (1981) and for the second dimension with techniques described by Laemmli (1970). The internal markers, bovine serum albumin (68 kDa, pH 6.3) phosphoglycerokinase (42 kDa, pH 7.4) and rabbit actin (42 kDa, pH 5.4) were added. The gels were stained with Coomassie blue (Ouhayoun et al., 1985).
For transmission electron microscopy, the tissues were washed in 0.1 M sodium cacodylate buffer containing 0.1 M sucrose (pH 7.4), fixed for 30 min in 2% glutaraldehyde in the washing buffer, and

Cytokeratins of cultured oral epithelia 749
post-fixed for 15 min in 0.5% osmium tetroxide and 0.8% potassium ferrocyanide in cacodylate buffer. After washing, the tissues were incubated in 0.15% tannic acid for 1 min to enhance staining, then thoroughly washed. The tissues were stained with 2% uranyl acetate in 50% ethanol for 40 min, then dehydrated in graded ethanols and propylene oxide before embedding in Epon-Araldite. Semithin sec- tions were cut and stained with methylene blue and thin sections were st.sined with lead citrate solution (1.76 g lead citrate and 1.83 g lead nitrate in 50 ml H,O) for 30 min. A IPhilips 400 electron microscope was used to examin’? the sections at 80 kV. These methods were found to reduce artifacts and provide high contrast (Shabana et al., 1991).
RESULTS
Histology
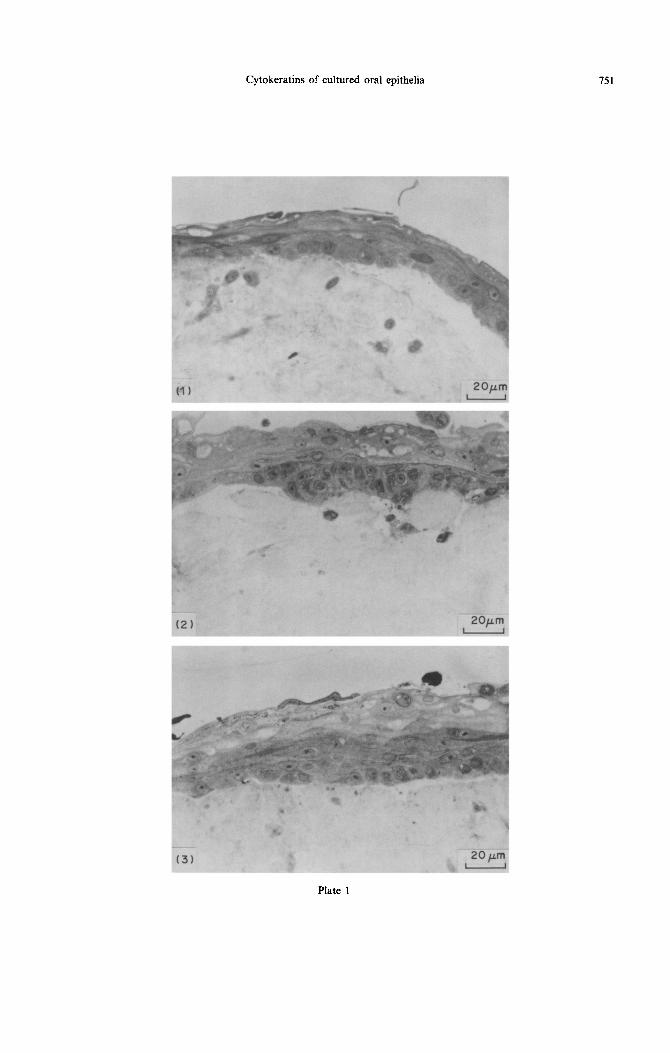
The epithelium covered the collagen surface with stratified squamous type epithelium and extended to the sides of the lattices but did not cover the lower aspect. It was lo-14 cells thick at the middle part near the degenerating explant, and gradually became thin- ner at the leading edge where it was l-2 cells thick, Of the three groups, the middle part of the reconsti- tuted alveolar mucosa had the thickest epithelium and that of the palatal mucosa the thinnest (Figs l-3). The junction between the epithelium and the collagen lattice was generally flattened, with excep- tionally short epithelial extensions into the collagen. In some lattices, epithelial cells penetrated the pores of the lattices. These appeared in sections as irregular strands and islands of variable depth and thickness. The basal epithelial cells were cuboidal in the middle part and flattened at the borders. The suprabasal cells were polygonal and the most superficial layers were flattened. The superficial cells of gingival and alveolar mucosal origins retained their nuclei, whereas those of palatal origin were non-nucleated and densely stained, resembling orthokeratinized cells.
Ultrastructurally, l.he epithelial cells maintained healthy features and had numerous intercellular bridges in the spinous and superficial cell layers. Tonofilaments were also more prominent as the cells matured in the suprabasal layers. Keratohyalin-like granules were detected in the superficial cell layers, principally in cells of palatal origin (Fig. 4), although a few could be found in cells of other origins. Intercellular attachment occurred through tight junc- tions, particularly in the basal cell layer, and through desmosomes, particularly in the spinous and super- ficial layers. Intercellular bridges from adjacent cells were attached side-to-side by desmosomes. The epithelial-collagen junction was flattened, with no evidence of either basement membrane or hemi- desmosome formation. However, electron-dense, filamentous structures and vesicles were noticed in the subepithelial collagen which could be extracellular epithelial products such as fibronectin and collagen. In the lattice, collager. bundles were almost 40 nm in diameter and did not have a specific orientation. Fibroblasts were stellate with long cytoplasmic pro- cesses extending in the three-dimensional space. The cells were rich in mitochondria, Golgi, rough endo- plasmic reticulum, free and attached ribosomes.
Two -dimensional gel eiectrophoresis
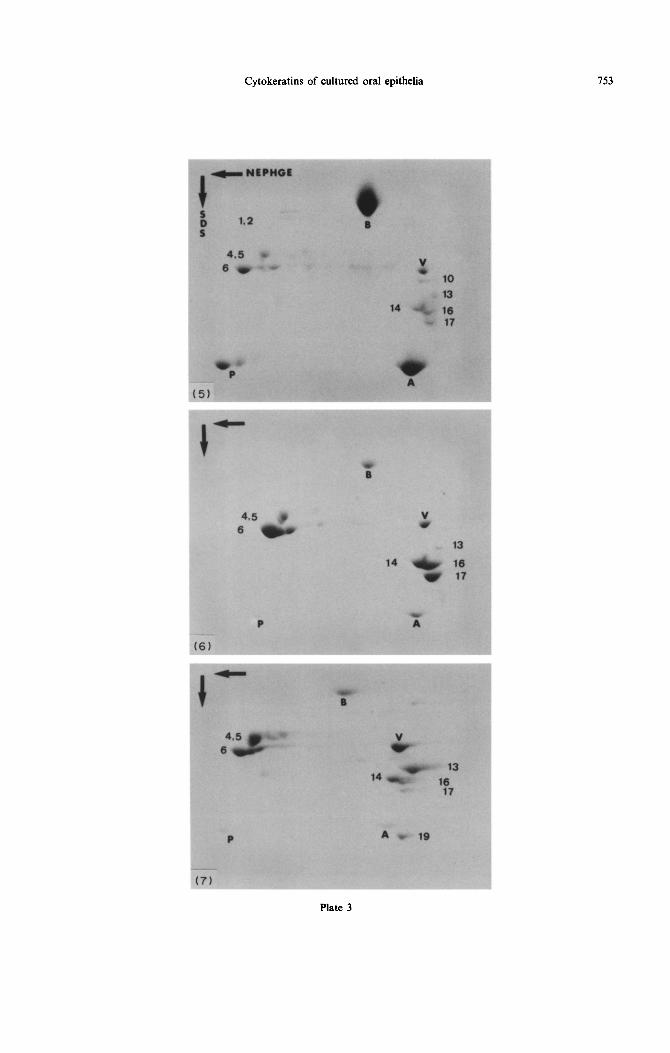
Cytokeratins isolated from the different groups are shown in Figs 5-7. Vimentin was identified in all gels prepared from whole tissue. When the collagen con- taining fibroblasts was removed by peeling the epi- thelium with fine tweezers, this intermediate filament protein was not detected (not shown). This suggested that vimentin was not expressed by these epithelial cells or was expressed in quantities that escaped detection in the gels. A constant set of polypeptides, CK 5, 6, 14, 16 and 17, was found in all groups. Polypeptides 13 and 19 were also found in all groups but with differences between them. In the alveolar mucosa, CK 13 and 19 formed major bands (Fig. 7), whereas in the other mucosae, they formed less prominent bands or were even absent, depending on the protein charge (Figs 5 and 6). Cytokeratin 4 appeared too close to CK 5; whether it was also a major band in the alveolar mucosa was difficult to ascertain. Terminal differentiation keratins were detected as minor bands in reconstituted palatal mucosa (Fig. 5). Cytokeratin 18 formed small spots in overloaded gels of different epithelia, not in normal or underloaded ones.
Immunohistochemistry
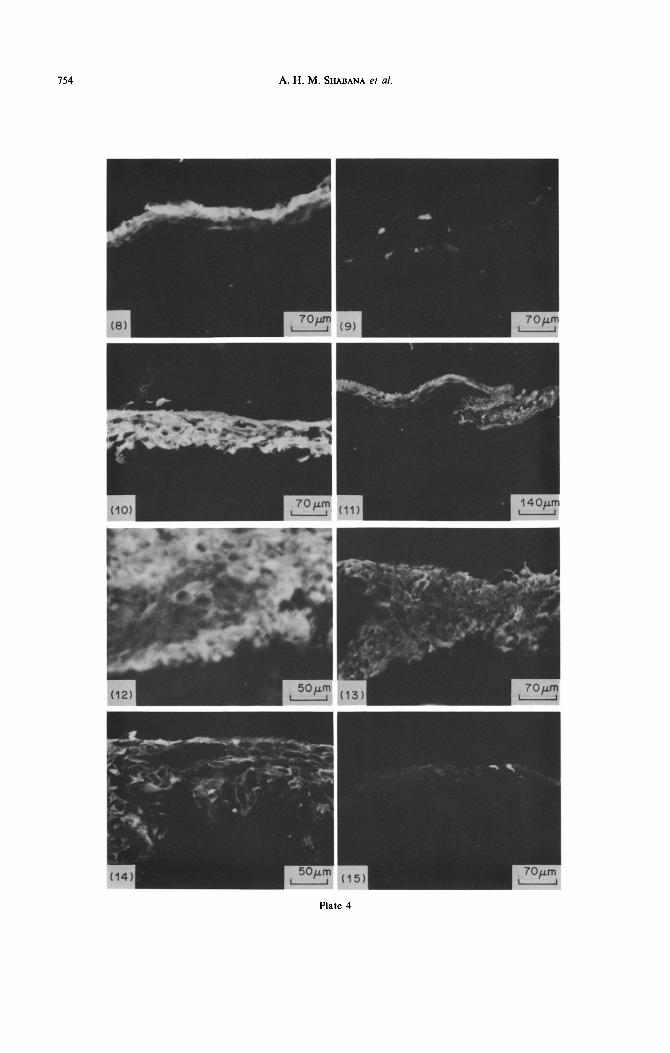
Terminal differentiation keratins were found in suprabasal palatal cells, as detected by monoclonal EE21.6 and K,8.60 (Fig. 8). The thin epithelium at the borders of the mucosae did not react with these antibodies. In the gingival and alveolar cells, these keratins were generally not detectable; however, weak labelling of individual, superficial, gingival cells was found (Fig. 9). The monoclonal antibodies K,l3.1 and lC7 labelled the suprabasal cells in all groups. Whilst K,l3.1 did not show differences between groups, lC7 labelling was stronger and more homo- geneous in alveolar epithelial cells than in cells of palatal and gingival origin (Figs 10 and 11). Cytoker- atin 4, whose presence in gels was uncertain, was clearly labelled by monoclonal 6BlO. The suprabasal cells were moderately labelled, with no differences between groups (Fig. 12).
Strong and homogeneous reaction was obtained with monoclonal KB-37, an antibody that labels basal-cell cytokeratins in stratified squamous epithe- lia. In culture, the reaction was not restricted to the basal-cell layer but extended to the suprabasal cells. This antibody also labelled the thin epithelium at the borders. The simple epithelial keratin CK 18 was detected with monoclonal antibody K, 18.174. Mod- erately strong reaction was observed in all epithelial layers of all epithelial groups (Fig. 13), In contrast, in situ localization of CK 19 varied between different epithelia and within one epithelial group. Generally the reaction was stronger and more homogeneous in cells from the alveolar mucosa (Fig. 14). In some areas, labelling occurred in the superficial cell layers rather than in the basal-cell layer (Fig. 15).
DISCUSSION
We here describe cytokeratin expression by human oral epithelia cultured in a three-dimensional model (see Table 1). This system provided a tissue bearing

750 A. H. M. SHABANA et al.
some resemblance to the oral mucosa in the presence of connective tissue (collagen lattice populated with living fibroblasts) and covering stratified squamous epithelium. As homotypic cells were used in the preparation, we named each culture according to the anatomical origin of the cells. This model provides a possibility of growing fibroblasts in three-dimen- sional collagen and of co-culturing epithelial cells on the surface with no risk of their being over- grown by the fibroblasts. This model is then poten- tially of use in studying cellcell and cell-matrix interactions as well as their influences on epithelial differentiation.
By using explant outgrowth on collagen lattices (Saiag et al., 1985) we avoided the extensive mechan- ical and chemical manipulations with enzymes and detergents that result in the death of an important proportion of cells. The explants were removed to avoid any ambiguity in the cytokeratin profile.
Epidermal cells in primary cultures stratify and present some aspects of normal epidermal differen- tiation in uivo; they form a stratum corneum-like superficial layer and keratin envelopes (Flaxman, Lutzner and van Scott, 1967; Fusenig and Worst, 1975; Jepsen, MacCallum and Lilhe, 1980; Jensen
and Bolund, 1988). However, they develop only a small number of desmosomes and rarely keratohyalin granules (Breitkreutz et al., 1984). We found that the oral epithelia similarly stratified and differentiated on collagen gels populated with fibroblasts. When com- paring epithelia from different oral sites, we detected differences between those from keratinized and non- keratinized origins which correlated with the mor- phological differentiation of these epithelia in uivo. The palatal epithelium was thin, with a flattened epithelialcollagen junction, and superficial cells were non-nucleated and had keratohyalin-like granules. In contrast, the alveolar mucosal epithelium was slightly thicker, with the superficial layers mostly lacking keratohyalin granules and retaining their nuclei.
Cytokeratin polypeptides provide markers for epi- thehal differentiation and are site-specific for different epitheha. We found that CK 5, 6, 14, 16 and 17 were consistently expressed by all epithelial cells. All these cytokeratins are expressed by normal oral epithelial cells in viva and, with the exception of CK 17, by cultured gingival and epidermal cells (Bowden et al., 1987); Oda, Dale and Bourekis, 1990). Cytokeratin 5 and 14 are basically expressed by the basal and lower spinous cells of stratified epithelia, and their
Plate 1
Fig. 1. Semithin Epon-Araldite sections of reconstituted mucosa stained with methylene blue. The figure shows randomly distributed fibroblasts in collagen matrix and the covering epithelium. Both cells originate from the mucosa of the hard palate (PE/PF). The covering epithelium is stratified squamous: the basal cells are cuboidal, the middle cells are polygonal spinous, and the superficial cells are flattened with no nuclei. The junction between the collagen lattice and the epithelium is weak and flattened, resulting in
artificial separation between the two tissues. x 500
Fig. 2. Reconstituted gingival mucosa also produced a stratified squamous epithelium which is slightly thicker than the palatal. The epithehalLcollagen junction has short epithelial processes. The superficial cells
retained their nuclei and the cytoplasm is lightly stained. x 500
Fig. 3. Stratified squamous epithelium of the reconstituted alveolar mucosa was slightly thicker than that of both reconstituted gingival and palatal mucosae, and this thickness was mostly in the spinous layer. The superficial cells stained lightly and retained their nuclei. The epithelial-collagen junction is weak and
flattened. x 500
Plate 2
Fig. 4. Thin section of reconstituted palatal mucosa. The superficial cell layer of the epithelium has desmosomal junctions between the lateral aspects of intercellular bridges from adjacent cells (short arrows). The cells cytoplasm is rich in intermediate filaments, which form bundles. In association with
these filaments, several keratohyaline-like granules (long arrows) were found. x 9524
Plate 3
Fig. 5-7. Two-dimensional gel electrophoresis of three reconstituted oral mucosae. Coomassie-blue staining revealed cytoskeletal proteins in these gels. The horizontal, large arrow denotes non-equilibrium- pH gradient-electrophoresis used in the first dimension; the vertical arrow indicates migration in sodium dodecyl sulphate used in the second dimension. The different polypeptides are numbered according to the catalogue of Moll et al., (1988). The internal reference markers are actin (A), bovine serum albumin (B) and phosphoglycerokinase (P). Note the presence of vimentin (V) in the intermediate filaments of fibroblasts. In all gels, keratin 5 overlapped with another band, possible keratin 4, which made interpretation of the existence of K 4 difficult in gels. The band of CK 6 migrates slow in these gels. Such
slow migration was also seen in gels from inflamed gingiva (Ouhayoun et al., 1990).
Fig. 5. Palatal epithelium expressed traces of terminal differentiation keratins (CK 1, 2, 10 and 1 l), basal cell keratins (CK 5 and 14), hyperproliferation keratins (CK 6 and 16), and traces of oesophageal keratin (CK 13) as well as keratin 17. Simple epithelial keratins (CK 7, 8, 18 and 19) were not detected in gels.
Fig. 6. Two-dimensional gel of reconstituted gingiva. The gingival epithelial extracts differed from the palatal by the absence of terminal differentiation keratins, and the variable expression of simple epithelial
keratins (CK 19 and 18), appearing as minor bands in overloaded gels.
Fig. 7. Two-dimensional gel of reconstituted alveolar mucosa. The alveolar epithehum expresses the oesophageal keratin 13 and the simple epithelial keratin 19 as major bands. Terminal differentiation keratins were not detected. In common with palatal and gingival epithelia, keratins 5, 6, 14, 16 and 17
were detected.
Cytokeratins of cultured oral epithelia 751
_-__.-
(3)
Plate I

752 A. H. M. SHABANA et al.
Plate 2
Cytokeratins of cultured oral epithelia 753
Plate 3
154

1 2
Cytokeratins of cultured oral epithelia
Table I. Cytokeratins of the reconstituted mucosae
Cytokeratin polypeptides
4 5 6 10 11 13 14
755
16 17 18 19
Palatal + + ++ ++ + ++ ++ + Gingival - - ++ ++ + Alveolar +* ++ ++ - - +++ ++ ++ +
-, Not detected; +, trace; + +, large quantity. *Results based on immunohistochemical detection using monoclonal antibodies.
expression in our model suggests the presence of this phenotype in the culture. Furthermore, the hyperpro- liferation markers, CK 6 and 16 (Weiss, Eichner and Sun, 1984) were of the more prominant polypeptides expressed by our cultured oral epithelial cells.
The greater part of our understanding of cytoker- atin expression and lepithelial differentiation comes from studies on epidennal cells. In conventional, submerged cultures ofepidermal cells, cytokeratins of terminal differentiation (CK 1, 2, 10 and 11) are not expressed. The expression of these polypeptides is inhibited by vitamin A in serum and by retinoids (see Introduction). We found that they are expressed by palatal epithelium in the presence of 10% filter- delipidized FCS when grown on freely floating colla- gen lattices. This is consistent with their expression by epidermal cells grown at the air-liquid interface (Asselineau et al., 1985), and when filter-delipidized serum was used (Basset-Seguin et al., 1990). Gingival and alveolar mucosal epithelial cells did not express these terminal differentiation markers. They ex- pressed CK 13 and 4, which are differentiation markers for non-keralinized stratified squamous epi- thelia such as in the alveolar mucosa, although they may also be detected in small quantities in the gingival and palatal epithelia. In our culture, CK 13 was weakly detected in cultured palatal and gingival cells and strongly in alveolar mucosal cells. Oda et al. (1990) did not detect CK 13 in adult gingival cells in a serum-free growth medium containing low Ca*+ but did detect it in moderate quantities at physiological levels of Ca*+.
Certain simple epithelial keratins were expressed in our culture. CK 19 was detected in an important
quantity in alveolar epithelial cells. Gingival and palatal cells expressed this polypeptide in quantities detectable by immunofluorescence, but it was very weak or absent in gels. In viva, CK 19 is detectable in the basal and parabasal cell layers of normal alveolar mucosa (Shabana et al., 1989), in inflamed gingiva (Ouhayoun et al., 1990; Bosch et al., 1990) and in dysplastic epithelia of different oral sites (Lindberg and Reinwald, 1989; Mori et al., 1985). Cultured epidermal and gingival epithelia may ex- press this polypeptide in culture. CK19 was expressed in large quantities by gingival epithelia when the media contained cholera toxin (Southgate et al., 1987) and by gingival cells cultured in serum-free medium con- taining epidermal growth factor (Oda et al., 1990). It is of interest that elevators of cyclic AMP reverse the keratinization of the trachea of vitamin A-deficient hamsters in organ culture and CK 19 is re-expressed (Schiff and Moore, 1985). These findings suggest that CK 19 is expressed by CAMP-enhanced epithelial cells. If this suggestion holds true, it can explain the in situ alteration of CK 19 distribution in culture.
The expression of CK 18 in our cultured gingival cells is in agreement with the findings of Bowden et al. (1987), who used foreskin keratinocytes, but contradict those of Southgate et al. (1987), who used gingival epitheha cells at low serum level. Retinoic acid was found to enhance the expression of simple epithelial cytokeratins (18 and 19), as shown in rabbit tracheal cells (Jetten et al., 1989).
There is some evidence that different cells respond differently to the modulators of epithelial differen- tiation: for example, fetal and not adult gingival cells express CK 13 in low Ca2+ and serum-free medium
Plate 4 Fig. 8. Indirect irnmunofluorescence for terminal differentiation keratins (CK I, 2, IO and 11). Monoclonal
EE21.6 labelled suprabasal cell layers of reconstituted palatal mucosa. x 142
Fig. 9. In reconstituted gingival mucosa, terminal differentiation keratins were detected in scattered suprabasal cells. The monoclonal antibody Kk8.60 labelled the cells expressing keratins 10 and 11. x 142
Fig. 10. Cytokeratin 13 was strongly labelled in the epithelium of reconstituted alveolar mucosa using monoclonal antibody lC7. The reaction was detected in the suprabasal cell layers. x 142
Fig. 11. The reconstituted gingiva expressed keratin 13 in suprabasal cell layers as detected by monoclonal antibody lC7. The reaction is, however, weak as compared with that found in the reconstituted alveolar
mucosa (see Fig. 10). x70
Fig. 12. Cytokeratin 4 was detected principally in suprabasal layers of reconstituted alveolar mucosa using monoclonal antibody 6BlO. x 142
Fig. 13 Simple epithelial keratin, CK 18, was detected in the stratified squamous epithelium of the reconstituted mucosae (in this case, reconstituted gingival mucosa). The monoclonal K, 18.174 showed
variation of labelling intensity for one area to another. x 142
Fig. 14. Simple epithelial keratin, CK 19, was expressed by epithelial cells of reconstituted alveolar mucosa. The monoclonal antibody K,19.1 strongly labelled the cells of the superficial and spinous cell layers. x 200
Fig. 15. Simple epithelial keratin, CK 19, was expressed by individual epithelial cells of the reconstituted palatal mucosa. Monoclonal K”l9.1 labelled weakly some cells in the spinous cell layer. x 142

756 A. H. M. SHABANA et al.
(Oda et al., 1990). Furthermore, in conventional, and in cystadenolymphomas demonstrated by mono- submerged culture at normal levels of vitamin A, clonal antibodies against selective cytokeratin polypep-
HACAT human keratinocytes express terminal tides. Virchows Arch. 411A, 583-589.
differentiation keratins (Ryle et al., 1989) whereas Bosch F. X., Ouhayoun J. P., Bader B. L., Collin C., Grund
SCC- 13 cells show marked reduction of these keratins C., Lee I. and Franke W. W. (1990) Extensive changes in
by retinoic acid (Kopan and Fuchs, 1989). Site of cytokeratin expression patterns in pathologically affected
origin of epidermal cells also influences cytokeratin human gingiva. Virchows Arch. 58B, 59-77.
expression in vitro; this was found when comparing Bowden P. E., Stark H. J., Breitkreutz D. and Fusenig
epidermal cells from foreskin with those from the N. E. (1987) Expression and modification of keratin during terminal differentiation of mammalian epidermis.
back, arm, leg and breast [Bowden et al., 19871. In Current Topics in Developmental Biology, Vol. 22, We found that the alveolar mucosal epithelial cells pp. 35-68. Academic Press, London.
(from non-keratinized oral mucosa) express import- Breitkreutz D., Bonhart A., Hertzmann E., Bowden P. E.
ant quantities of CK 13 and 19 as compared with Boukamp P. and Fusenig N. E. (1984) Differentiation
those of the gingival and palatal cells. CK 4, which specific functions in cultured and transplanted mouse
was difficult to identify in gels, was readily detected keratinocytes: environmental influences on ultrastructure
by a monoclonal antibody. In contrast, the palatal and keratin expression. Differentiation 26, 154-169.
cells expressed, though in small quantities, terminal Brown R., Gray R. H. and Bernstein I. A. (1985) Retinoids
differentiation keratins characteristic of keratinizing alter the direction of differentiation in primary cultures of cutaneous keratinocytes. Di&entiati& 28, i68-278.
stratified squamous epithelium. These findings corre- Clark R. A. F.. Folkvord J. M. and Mertz R. L. (1985) late well with cytokeratin profiles of these epithelia Fibronectin, as well as other extracellular matrix proteins;
in vivo (Clausen et al., 1986a, b; Morgan et al., 1987; mediate human keratinocyte adherence. J. invest. Derm.
Ouhayoun et al., 1985; Shabana et al., 1989). 84, 378-383.
Our model is of interest for studying epithelial Clausen H., Moe D., Buscard D. and Dabelsteen E. (1986a)
differentiation, as the differentiation markers of kera- Keratin proteins in human oral mucosa. J. oral Path. 15,
tinized epithelia were expressed by the palatal cells, 36-42.
and those of non-keratinized epithelia were promi- Clausen H., Vedtofte P., Moe D., Dabelsteen E., Sun T.-T.
nent in the alveolar mucosal cells. These findings and Dale B. (1986b) Differentiation-dependent expression
provide a baseline for further studies on the modu- of keratins in human oral epithelia. J. invest. Derm. 86, 249-254.
lation of oral epithelial differentiation. Cooper D., Schermer A. and Sun T.-T. (1985) Classification of human epitheha and their neoplasms using monoclonal
Acknowlednemenfs-Thanks are due to Dr B. Coulomb and antibodies to keratins: strategies, applications and limi-
Mrs C. Lebreton, Laboratoire de Dermatologie (Professor tations Lab. Invest. 52, 243-256.
L. Dubertret), INSERM U, 312, Creteil, France, for helping Flaxman B. A., Lutzner M. A. and van Scott E. J. (1967)
us in realizing the culture model. We are grateful to Cell maturation and tissue organization in epithelial
Professor W. W. Franke, German Cancer Research Centre. outgrowth from skin and buccal mucosa in vitro. J. invest.
Heidelbere. Fed. Reo. Germanv. and Dr G. Serres Labora- Derm. 49, 322-331.
toire d’E;plorations Biologiques Cehulaires, Toulouse, Franke W. W., Schiller D. L., Mall R., Winter S., Schmid
France, for providing us with valuable monoclonal anti- E., Engelbrecht I., Denke H., Krepler R. and Platzer B.
bodies. The technical assistance of Mme N. Martin and (1981) Diversity of cytokeratins: differentiation-specific
Mme. M. Oboeuf was much appreciated. We also thank the expression of cytokeratin polypeptides in epithelial cells
secretarial work of Mr E. Marie-Rose. This work was and tissues. J. Molec. Biol. 153, 933-959.
supported by INSERM Grant No. Cre, 877007. Fuchs E. and Green H. (1981) Regulation of terminal differentiation of cultured human keratinocytes by vita-
REFERENCES min A. Cell 25, 617-625.
Fusenig N. E. and Worst P. K. M. (1975) Mouse epidermal Achtstatter T., Hatzfeld M., Quinlan R. A., Parmelee D. C. cell cultures--II. Isolation, characterization and culti-
and Franke W. W. (1986) Seoaration of cvtokeratin polypeptides by gel electrophoreiic and chromatographic
vation of epidermal cells from perinatal mouse skin. Expt Cell Res. 93, 443-457.
techniques and their identification by immunoblotting. Gao Z., Mackenzie I. C., Williams D. M., Cruchley A. T. Merh. Enzym. 134, 355-371. and Lane E. B. (1988) Patterns of keratin expression in
Assehneau D., Bernhard B., Bailly C. and Darmon M. rests of Malassez and oeriaoical lesions. J. Oral Path. 17, . . (1985) Epidermal morphogenesis and induction of 67 kD 178-185. keratin polypeptide by culture of human keratinocytes at Green H. and Watt F. M. (1982) Regulation by vitamin A the liquid-air interface. Expl Cell Eiol. 159, 5366539. of envelope cross-linking in cultured keratinocytes de-
Asselineau D., Bernhard B., Bailly C. and Darmon M. rived from different human epithelia. Molec. Ce// Eiol. 2, (1989) Retinoic acid improves epidermal morphogenesis. 1115-1117. Deal Biol. 133, 322-335. Hay E. D. (1977) Cell-matrix interaction in embryonic
Basset-Stguin N., Culard J.-F., Kerai C., Bernard F., induction. In International Cell Biology (Ed. Brinkley Watrin A., Demaille J. and Guilhou J.-J. (1990) Reconsti- B. R. and Porter K. R.), p. 50. Rockefeller University tuted skin in culture: a simple method with optimal Press, New York. differentiation. Differenfiation 44, 232-238. Huszar M., Gigi-Leitner O., Mall R., Franke W. W. and
Bell E., Erlich H. P., Buttle D. and Nakatsuji T. (1981) Geiger B. (1986) Monoclonal antibodies to various acidic Living tissue formed in vitro and accepted as a skin- (type I) cytokeratins of stratified epithelia. Differentialion equivalent tissue of full thickness. Science 211, 1052-1054. 31, 141-153.
Bell E., Ivarsson B. and Merrill C. (1979) Production of a Jensen P. K. A. and Bolund L. (1988) Low Ca*+ stripping tissue-like structure by contraction of collagen lattices by of differentiating cell layers in human epidermal cultures: human fibroblasts of different proliferation potential an in oitro model of epidermal regeneration. Expl Cell in vitro. Proc. natn. Acad. Sci., U.S.A. 76, 1274-1278. Res. 175, 63-73.
Born I. A., Schwechheimer K., Maier H. and Otto H. F. Jensen P. K. A. and Therkelsen A. J. (1981) Cultivation at (1987) Cytokeratin expression in normal salivary glands low temperature as a measure to prevent contamination

Cytokeratins of cultured oral epithelia 157
with fibroblasts in epithelial cultures from human skin. J. invest. Derm. 11, 210-212.
Jepsen A., MacCallum. D. K. and Lillie J. H. (1980) Fine stucture of subcultivated stratified squamous epithelium. Expl Cell Res. 125, 141-152.
Jetten A. M., George M. A., Smits H. L. and Vollberg T. M. (1989) Keratin 13 expression is linked to squamous differentiation in rabbit tracheal epithelial cells and down-regulated by retinoic acid. Expl Cell Res. 182, 622-634.
Juhl M., Reibel J. and Stoltze K. (1989) Immunohisto- chemical distribution of keratin proteins in clinically healthy human gingival epithelium. Stand. J. dent. Res. 97, 159-170.
Karasek M. A. and {Charlton M. E. (1971) Growth of postembryonic skin epithelial cells on collagen gels. J. Invest. Dermat. 56, 205-210.
Karsten U., Papsdorf G., Roloff G. Stolley P., Abel H., Walther I and Weiss H. (1985) Monoclonal anti-cytoker- atin antibody from a hybridoma clone generated by electrophoresis. Eur. J. Cancer Clin. Oncol. 21, 733-740.
Kasper M., Karsten lJ., Stosiek P. and Moll R. (1989) Distribution of intermediate-filament proteins in the human enamel organ; unusually complex pattern of coexpression of cytokeratin polypeptides and vimentin. Differentiation 40, 207-214.
Kim K. H., Schwartz F:. and Fuchs E. (1984) Differences in keratin synthesis between normal epithelial cells and squamous cell carcinoma are mediated by vitamin A. Proc. natn. Acad. Sci. U.S.A. 81. 4280-4284.
Kopan R. and Fuchs 13. (1989) The use of retinoic acid to probe the relation between hyperproliferation-associated keratins and cell proliferation in normal and malignant epidermal cells. J. Cell Biol. 109, 295-307.
Kopan R., Traska G. and Fuchs E. (1984) Retinoids as important regulators of terminal differentiation: examin- ing keratin expression in individual epidermal cells at various stages of keratinization. J. Cell Biol. 105, 427-440.
Kubo M., Kan M., Isemura M., Yamane I. and Tagami H. (1987) Effects of extracellular matrices on human kera- tinocyte adhesion ar.d growth, and on its secretion and deposition of fibronectin in culture. J. invest. Derm. 88, 594-601.
Laemmli U.K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-685.
Lindberg E. K. and Relnwald J. G. (1989) Suprabasal40 kd keratin (K19) an immunohistologic marker of premalig- nancy in oral epitheiium. Am. J. Path. 134, 89-98.
Moll R., Achstatter ‘I’., Becht E., Barcarora-Stander J., Ittensohn M. and Franke W. W. (1988) Cytokeratins in normal and malignant transitional epithelium. Am. J. Path. 132, 123-144.
Mall R., Franke W. W. and Schiller D. L. (1982) The catalogue of human cytokeratins: patterns of expression in normal epithelial, tumor and cultured cells. Cell 31, 1 l-24.
Morgan P. R., Leigh I. M., Purkis P. E., Gardner I. D., van Muijen G. N. P. and Lane E. B. (1987) Site variation in keratin expression in human oral epithelia-an immuno- cytochemical study of individual keratins. Epithelia 1, 31-43.
Mori M., Nakai M., Hyun K.-H., Noda Y. and Kawamura K. (1985) Distribution of keratin proteins in neoplastic and tumor-like lesions of squamous epithelium, an im- munohistochemical study. Oral Surg. 59, 63-69.
van Muijen G. N. P., Rinter D., Franke W. W., Achtstltter T., Haasnoot W. H. B., Ponec M. and Warnaar S. 0. (1986) Cell type heterogeneity of cytokeratin expression in complex epithelia and carcinomas as demonstrated by monoclonal antibodies specific for cytokeratins no. 4 and 13. Expl Cell Res. 162, 97-l 13.
Oda D., Dale B. A. and Bourekis G. (1990) Human oral epithelial cell culture-II. Keratin expression in fetal and adult gingival cells. In Vitro 26, 596-603.
O’Farrell P. Z. Goodman H. H. and O’Farrell P. H. (1977) High resolution two dimensional electrophoresis of basic as well as acidic proteins. Cell 12, 1133-l 142.
Ouhayoun J. P., Goffaux J. C., Sawaf M. H., Shabana A. H. M., Collin C. and Forest N. (1990) Changes in cytokeratin expression in gingiva during inflammation. J. periodont. Res. 25, 283-292.
Ouhayoun J. P., Gosselin F., Forest N., Winter S. and Franke W. W. (1985) Cytokeratin patterns of human oral epithelia. Differences in cytokeratin synthesis in gingival epithelium and the adjacent alveolar mucosa. Di&en- tiation 30, 123-129.
Pelissier A., Ouhayoun J. P., Sawaf M. H. and Forest N. (1990) Evolution of cytokeratin expression in developing human tooth germ. J. Biol. Buccale. 18, 99-108.
Ponec M., Weerheim A., Kempenaar J. and Boonstra J. (1988) Proliferation and differentiation of human squamous carcinoma cell lines and normal keratinocytes: effects of epidermal growth factor, retinoids and hydro- cortisone. In Vitro 24, 764-770.
Regnier M., Schweize J., Michel S., Bailly C. and Prunieras M. (1986) Keratin in human keratinocytes cultured on dead de-epidermized dermis. Expl Cell Res. 165, 63-72.
Ryle C. M., Breitkruetz D., Stark H.-J., Leigh I. M., Steinert P. M., Roop D. and Fusenig N. E. (1989) Den- sity-dependent modulation of synthesis of keratin 1 and 10 in the human keratinocyte line HACAT and in ras-transfected tumorgenic clones. Differentiation 40, 42-54.
Sacks P. G., Parnes S. M., Price J. C., Risemberg H., Goldstein J. C., Marko M. and Parsons D. F. (1985) In vitro modulation of differentiation by calcium in organ cultures of human and murine epithelial tissue. In Vitro 21, 99-107.
Saiag P., Coulomb B. and Lebreton C. (1985) Psoriatic fibroblasts induce hyperproliferation of normal kerati- nocytes in a skin equivalent model in vitro. Science 230, 669-672.
Sawaf M. H., Ouhayoun J. P., Shabana A. H. M. and Forest N. (1990) Cytokeratin expression in human tongue epithelium. Am. J.. Anat. 189, i55-166.
Schiff L. J. and Moore S. J. (1985) Effects of cvclic adenosine 3’: 5’-monophosphate-elevating agents -and retinoic acid on differentiation in retinoid-deficient tra- cheal cultures. In Vitro 21, 688-692.
Shabana A. H. M., Ouhayoun J. P., Boulekbach H. and Forest N. (1991) Ultrastructural study of the effects of coral skeleton on cultured human gingival fibroblasts in three-dimensional collagen lattices. Mater. Med. In press.
Shabana A. H. M., Ouhayoun J. P., Sawaf M. H. and Forest N. (1989) A comparative biochemical and immunological analysis of cytokeratin patterns in the oral epithelium of the miniature Die. and man. Archs oral Biol. 34. 249-259.
Slavkin H. C. (1980) Oral Premalignancy (Eds MacKenzie I. C., Dabelsteen E. and Squire C. A.), Chap. 10, p. 139. University of Iowa Press, IA.
Southgate J., Williams H. K., Trejdosiewicz L. K. and Hodges G. M. (1987) Primary culture of human oral epithelial cells. Growth requirements and expression of differentiated characteristics. Lab. Invesf. 56, 21 l-223.
Sun T.-T., Eichner R., Schermer A., Cooper D., Nelson W. G. and Weiss R. A. (1984) Classification. exnression . , _ and possible mechanism of evolution of mammalian epithelial keratins: a unifying model. Cancer Cell 1, 169-176.
Tyner A. L. and Fuchs E. (1986) Evidence for posttranscrip- tional regulation of the keratins expressed during hyper- proliferation and malignant transformation in human epidermis. J. Cell Biol. 103, 1945-1955.
AOB 36,1&-c

758 A. H. M. SHABANA et al.
Watt F. M. (1984) Selective migration of terminally differen- antibody analysis of keratin expression in epidermal tiated cells from the basal layer of cultured human diseases: a 48- and 56-K dalton keratins as molecular epidermis J. Cell Biol. 98, 16-21. markers for hyperproliferative keratinocytes. J. Cell. Biol.
Weiss R. A., Eichner R. and Sun T.-T. (1984) Monoclonal 98, 1397-1406.