Embed Size (px)
Citation preview

Journal of Experimental Botany, Vol. 59, No. 7, pp. 1935–1950, 2008
doi:10.1093/jxb/ern086
SPECIAL ISSUE RESEARCH PAPER
Cysteine proteinases regulate chloroplast protein contentand composition in tobacco leaves: a model for dynamicinteractions with ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) vesicular bodies
Anneke Prins1,2, Philippus D.R. van Heerden3, Enrique Olmos4, Karl J. Kunert2 and Christine H. Foyer1,*
1 School of Agriculture, Food and Rural Development, Agriculture Building, Newcastle University, Newcastle uponTyne NE1 7RU, UK2 Forestry and Agricultural Biotechnology Institute, Botany Department, University of Pretoria, Pretoria 0002,South Africa3 School of Environmental Sciences and Development, Section Botany, North-West University, Potchefstroom 2520,South Africa4 CEBAS-CSIC, Department of Plant Physiology, PO Box 164, E-30080 Murcia, Spain
Received 31 January 2008; Revised 26 February 2008; Accepted 27 February 2008
Abstract
The roles of cysteine proteinases (CP) in leaf protein
accumulation and composition were investigated in
transgenic tobacco (Nicotiana tabacum L.) plants
expressing the rice cystatin, OC-1. The OC-1 protein
was present in the cytosol, chloroplasts, and vacuole
of the leaves of OC-1 expressing (OCE) plants.
Changes in leaf protein composition and turnover
caused by OC-1-dependent inhibition of CP activity
were assessed in 8-week-old plants using proteomic
analysis. Seven hundred and sixty-five soluble pro-
teins were detected in the controls compared to 860
proteins in the OCE leaves. A cyclophilin, a histone,
a peptidyl-prolyl cis-trans isomerase, and two ribulose-
1,5-bisphosphate carboxylase/oxygenase (Rubisco)
activase isoforms were markedly altered in abundance
in the OCE leaves. The senescence-related decline in
photosynthesis and Rubisco activity was delayed in
the OCE leaves. Similarly, OCE leaves maintained
higher leaf Rubisco activities and protein than controls
following dark chilling. Immunogold labelling studies
with specific antibodies showed that Rubisco was
present in Rubisco vesicular bodies (RVB) as well as
in the chloroplasts of leaves from 8-week-old control
and OCE plants. Western blot analysis of plants at 14
weeks after both genotypes had flowered revealed
large increases in the amount of Rubisco protein in
the OCE leaves compared to controls. These results
demonstrate that CPs are involved in Rubisco turnover
in leaves under optimal and stress conditions and that
extra-plastidic RVB bodies are present even in young
source leaves. Furthermore, these data form the basis
for a new model of Rubisco protein turnover involving
CPs and RVBs.
Key words: Chloroplast proteins, cysteine proteinase,
photosynthesis, protein turnover, senescence, vesicle
trafficking.
Introduction
Climate change and ongoing ecosystem degradationnecessitate the development of food and bio-energycrops that can support future increasing environmentalfluctuations. Endogenous plant cysteine proteinaseinhibitors or phytocystatins can be used to minimize
* To whom correspondence should be addressed. E-mail: [email protected]: 2-D, 2-dimensional; BSA, bovine serum albumin; CP, cysteine proteinases; GS, glutamine synthetase; LSU, large subunit; OC-1,oryzacystatin-1; OCE, oryzacystatin-1 expressing plants; PBS, phosphate buffered saline; Rubisco, ribulose-1,5-bisphosphate carboxylase/oxygenase;RVBs, Rubisco vesicular bodies; SDS-PAGE, sodium dodecyl sulphate polyacrylamide gel electrophoresis; SSU, small subunit; TCA, trichloroactetic acid.
ª The Author [2008]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved.For Permissions, please e-mail: [email protected]
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018

insect attack (Christou et al., 2006) and improve theyields of useful bio-engineered proteins such asvaccines (Rivard et al., 2006). However, while secondgeneration multiple proteinase inhibitor-containing in-sect-resistant plants are already in production (Christouet al., 2006) little is known about the effects of suchmanipulations on plant productivity. Cysteine protei-nases (CP) are involved with a variety of proteolyticfunctions in higher plants (Granell et al., 1998),particularly those associated with the processing anddegradation of seed storage proteins (Shimada et al.,1994; Toyooka et al., 2000), and fruit ripening (Alonsoand Granell, 1995). They are also induced in responseto stresses such as wounding, cold, and drought(Schaffer and Fischer, 1988; Koizumi et al., 1993;Linthorst et al., 1993; Harrak et al., 2001) and inprogrammed cell death (Solomon et al., 1999; Xu andChye, 1999). Like their CP targets, phytocystatins areregulated by developmental (Lohman et al., 1994) andenvironmental cues (Botella et al., 1996; Pernas et al.,2000; Belenghi et al., 2003; Diop et al., 2004).Two novel tobacco CP-coding sequences have pre-
viously been identified in tobacco including a KDEL-type CP NtCP2 (Beyene et al., 2006). The C-terminalKDEL motif, present in some cysteine proteinases, is anendoplasmic reticulum retention signal for soluble pro-teins that allows CP propeptides to be stored either ina special organelle, called the ricinosome (Schmid et al.,1999), or in KDEL vesicles (KV) before transport tovacuoles through a Golgi complex-independent route(Okamoto et al., 2003). The relatively acidic pH optimaof many of the endogenous plant CPs indicate that theyare localized in the vacuole (Callis, 1995). A papain-likesequence, termed NtCP1, was isolated from senescenttobacco leaves (Beyene et al., 2006). Papain-likecysteine proteinases are often found in senescing organsparticularly leaves (Lohman et al., 1994; Ueda et al.,2000; Gepstein et al., 2003), flowers (Eason et al.,2002), legume nodules (Kardailsky and Brewin, 1996) aswell as in germinating seeds (Ling et al., 2003).Senescence-associated genes (SAGs) are up-regulatedduring leaf senescence (Lohman et al., 1994; Quirinoet al., 1999; Swidzinski et al., 2002; Gepstein et al.,2003; Bhalerao et al., 2003; Lin and Wu, 2004). Of thesethe SAG12 cysteine proteinase is one of the very fewSAGs that are highly senescence-specific (Lohman et al.,1994).The expression of many photosynthesis genes such as
those encoding the chlorophyll a/b binding protein andthe subunits of ribulose-1,5-bisphosphate carboxylase-oxygenase (Rubisco) decreases during senescence andare hence they are classed as senescence down-regulatedgenes (Humbeck et al., 1996). Rubisco degradation canoccur both inside and outside the chloroplast (Irving andRobinson, 2006). Inside the chloroplast, oxidation of
critical cysteine residues on the Rubisco protein modifiesthe proteolytic susceptibility of these or associated aminoacids, causing the protein to adhere to the chloroplastenvelope and ‘marking’ the protein for degradation(Garcia-Ferris and Moreno, 1994). Recent evidencesuggests that the 26S proteasome is activated by carbon-ylation and hence this protein degradation pathway isenhanced when the cellular environment becomes evenmildly oxidizing (Basset et al., 2002). Vacuolar endopep-tidases and globules or vesicles released from thechloroplasts into the cytosol have been implicated inRubisco catabolism, but key questions have remainedregarding the extent to which Rubisco is degraded outsidethe chloroplast and how Rubisco degradation is controlled(Feller et al., 2007). In the chloroplast Rubisco isprotected against degradation by 2-carboxyarabinitol 1-phosphate (CA-I-P) but how this modulates degradationoutside the chloroplast is unknown (Khan et al., 1999).Little information is available on the effects of ectopic
phytocystatin expression on plant growth and develop-ment (Masoud et al., 1993; Guttierrez-Campos et al.,2001; Van der Vyver et al., 2003) as most studies haveconcentrated on effects on insect resistance or proteinproduction (Christou et al., 2006; Rivard et al., 2006).The phenotype resulting from expression of the ricecystatin, OC-1, in transformed tobacco plants has beendescribed previously (Masoud et al., 1993; Guttierrez-Campos et al., 2001; Van der Vyver et al., 2003).However, increased biomass production resulting fromcystatin expression under field conditions is often attrib-uted to enhanced insect resistance rather than to directeffects of the cystatin on endogenous protein turnover inthe plant tissues. Transgenic OC-1 expressing tobaccolines (OCE) grow more slowly with an extended vegeta-tive phase compared to the wild type or empty vectorcontrols (Van der Vyver et al., 2003). They are also moreresistant to chilling-induced inhibition of photosynthesis(Van der Vyver et al., 2003). The following study wasundertaken in order to determine how the constitutiveexpression of the rice cystatin, OC-1, in the cytosol oftobacco leaves alters leaf protein content and compositionand exerts effects on photosynthesis in leaves at differentstages of development.
Materials and methods
Plant material and growth conditions
OCE line T4/5 and wild-type control tobacco (Nicotiana tabacumL.) plants were grown in compost in pots in controlled environmentchambers (Controlled Environments Ltd., Winnipeg, MB, Canada,R3H 0R9) and rooms under a 15/9 h light/dark regime (with a lightintensity of 800–1000 lmol m�2 s�1) and a 26/20 �C day/nighttemperature cycle (Van der Vyver et al., 2003). The leaf ranking at14 weeks is denoted from the base to the tip of the stem. Hence,leaf one is the oldest leaf on the stem.
1936 Prins et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018

Chilling stress treatments
The attached shoots of 6-week-old OCE and control tobacco plantswere chilled in darkness at 5 �C for seven consecutive nights. At theend of each dark period the chilled plants were returned to optimaltemperatures for the subsequent light period. Leaf discs wereharvested from fully expanded leaves of OC-1 transformed (lineT4/5) and wild-type plants at the start of the experiment (day 0) andagain following 7 d of growth at 26/20 �C or 26/5 �C. Metabolismwas arrested in each leaf disc by freeze-clamping at liquid nitrogentemperatures. At the end of the experiment, leaf discs were alsocollected from young expanding leaves that developed during the7 d treatment period. Sampling occurred 4 h after the start of thelight period under full illumination.
Gas exchange measurements
CO2 assimilation was measured in fully expanded leaves of OCEand control tobacco plants. CO2 assimilation rates were measuredwith a portable photosynthesis system (CIRAS-2, PP-systems,Hertz, UK) at a light intensity of 1200 lmol m�2 s�1 and a leaftemperature of 26 �C. Carbon dioxide response curves weregenerated and used for the calculation of ACE (apparent carboxy-lation efficiency) and Jmax (maximal rates of photosynthesis at highCO2 concentrations). Leaf discs were collected from fully expandedleaves for the measurement of initial and maximum Rubisco activityaccording to the radiometric method previously described (Keysand Parry, 1990).
Ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco)
activities
Initial and total Rubisco activities were measured in soluble proteinextracts from the leaf samples according to Keys and Parry (1990).Initial activity is defined here as the activity of the enzyme underthe growth conditions at the time of sampling. Total activity isdefined here as the activity measured following activation of theextracted enzyme with bicarbonate. The total soluble proteincontent of extracts was determined according to the method ofBradford (1976).
Measurements of Rubisco degradation using in vitro assays
The effect of OC-1 expression on protein degradation in vitro wasdetermined according to the method of Yoshida and Minamikawa(1996). Soluble protein extracts (30 lg) from leaves of either wildtype or OC-1 transformed plants were incubated at 37 �C for 0–4 hin the presence or absence of 50 lM E64 (an inhibitor of cysteineproteinases) in 50 mM sodium acetate (pH 5.4) containing 10 mMb-mercaptoethanol. After incubation, samples were immediatelyloaded onto a native polyacrylamide gel (6%). Proteins wereseparated and the gel stained according to the method of Rintamakiet al. (1988).
CP activity measurements
CP activity was measured in leaf discs extracted in citrate phosphatebuffer (0.1 M, pH 6.5) as previously described (Barrett, 1980).
In-gel protease activity assays
Proteolytic activity was detected in plant extracts after mildlydenaturing gelatine-PAGE as previously described (Michaud et al.,1993).
Western blot analysis
Leaf discs were extracted in buffer containing 50 mM TRIS–HCl(pH 7.8), 1 mM EDTA, 3 mM DTT, 6 mM PMSF, and 30 mg
insoluble PVPP. Proteins were separated by standard SDS-PAGEprocedures. After transfer to nitrocellulose membranes (HybondC-extra, Amersham Pharmacia Biotech, UK) protein detection wasconducted using antibodies directed against Rubisco, Rubiscoactivase and glutamine synthetase (Foyer et al., 1993).
Electron microscopy and immunogold labelling
Leaf samples were fixed at 4 �C in 3% paraformaldehyde and0.25% glutaraldehyde in 0.1 M phosphate buffer (pH 7.2) for 2.5 h.The samples were dehydrated with a graded ethanol series andembedded in London Resin White (LR White) acrylic resin.Ultrathin sections (60–70 nm) were made on a Leica EM UC6Ultramicrotome (Leica Microsystems GMBH, Wetzlar). Ultrathinsections on coated nickel grids were incubated for 30 min in PBSplus 5% (w/v) BSA to block non-specific protein binding on thesections. They were then incubated for 3 h with either anti-RbcL(Rubisco Form I and Form II) antibody raised in rabbit (Agrisera,Vannas, Sweden) diluted 1:250 for the RbcL antibody and 1:100 forthe OC-1 antibody with phosphate buffered saline (PBS) plus 5%(w/v) bovine serum albumin (BSA) or with OC-1 antibody raised inrabbit (Van der Vyver et al., 2003). After washing with PBS plus1% (w/v) BSA, the sections were incubated for 1.5 h with thesecondary antibody goat anti-rabbit IgG gold labelled (10 nm,British BioCell International) diluted 1:50 with PBS plus 1% (w/v)BSA and 1% (w/v) Goat Serum (Sigma). The sections were washedsequentially with PBS (two washes) and distilled water (fivewashes). Ultrathin sections were then stained with uranyl acetatefollowed by lead citrate and observed in Philips Tecnai 12transmission electron microscope.
Two-dimensional (2-D) gel electrophoresis
In the following analysis the proteome of leaf 16 only from C andOCE tobacco plants was investigated. Leaf 16 extracts werecompared by 2D electrophoresis according to instructions in thehandbook, 2-D electrophoresis: principles and methods (GEHealthcare). Three technical replicates were prepared from eachextraction. Proteins were precipitated after grinding leaf material inliquid nitrogen. Ground leaf material (200–250 mg) was incubatedover-night at –20 �C in precipitation buffer (1 ml) containing TCA(10%, w/v) and b-mercaptoethanol (0.07% v/v) in acetone (100%,v/v). Precipitated protein was pelleted by centrifuging for 25 min at4 �C at 20 000 g and washed six times with ice-cold washing buffercontaining acetone (90%, v/v) and b-mercaptoethanol (0.07% v/v)in Milli-Q water. Proteins were solubilized in sample buffer (1 ml)containing 8 M urea, 2% (w/v) CHAPS, 61 mM DTT, and 0.5% (v/v)IPG buffer (pH 3–10) (GE Healthcare), by sonication in anultrasonic water bath for 1 h, with vortexing at 15 min intervals.Samples were then incubated in a heating block for 1.5 h at 30 �Cwith vortexing at 15 min intervals before overnight incubation atroom temperature for optimal protein solubilization. Cell debris wasremoved by centrifugation for 25 min at 20 000 g. Solubilizedproteins were quantified using the Bradford assay and ovalbumin(Sigma) as standard (Ramagli, 1999).Samples were diluted in sample buffer containing a few grains of
bromophenol blue to a concentration of 0.6 lg ll�1. Isoelectricfocusing was performed after active rehydration on 150 lg proteinusing Immobiline DryStrip immobilized pH gradient (IPG) strips(13 cm) (GE Healthcare) and the Ettan IPGphor apparatus (GEHealthcare), with voltage being increased stepwise as follows: 30 V(12 h; for rehydration of strip), 100 V (1 h), 500 V (1 h), 1000 V(1 h), 5000 V (1 h), and 8000 V (19 000 Vh) to obtain a total of26 000 Vh. IPG strips were then equilibrated for 15 min each inequilibration buffer (6 M urea, 50 mM TRIS–HCl pH 8.8, 30% v/vglycerol, 2% w/v SDS, a few grains of bromophenol blue)
Cysteine protease activity and Rubisco turnover 1937
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018

containing 65 mM DTT followed by equilibration in equilibrationbuffer containing iodoacetamide (25 mg ml�1).Second dimension focusing of proteins was performed by SDS-
PAGE on a 1 mm, 12% resolving gel with migration at 25 mAgel�1 for 20 min followed by 30 mA gel�1 for approximately 4 h oruntil the blue dye front had reached the bottom of the gel. Proteinswere fixed in the gel overnight by incubation in fixing solution(50% methanol, v/v, 10% acetic acid, v/v) on a rocking platform atlow speed.After fixing of protein, gels were rinsed three times in Milli-Q
water before being stained for 24 h in GelCode Blue (Pierce) ona rocking platform at low speed. Gels were rinsed three times inMilli-Q water before being scanned on a flatbed scanner for imageanalysis. Images were captured using the ImageMaster Labscansoftware, and analysed using Phoretix 2D Expression v2005software.
SELDI-TOF MS and LC-MS/MS
Spots of interest were excised from polyacrylamide gels after 2-Delectrophoresis for peptide fingerprint analysis by surface-enhancedlaser desorption ionization–time of flight mass spectrometry(SELDI-TOF MS) or serial mass spectrometry (LC-MS/MS). LC-MS/MS was performed on excised spots at the McGill ProteomicsPlatform (McGill University, Montreal, Quebec) using an ESI-Quad-TOF mass spectrometer. For SELDI-TOF MS the procedureaccording to Jensen et al. (1999) was followed. Peptide extracts(1–2 ll) from each tryptic digest were spotted onto an H4 Protein-Chip array (Ciphergen) and mixed with a-cyano-4-hydroxycinnamicacid (CHCA) [20%; in acetonitrile (5%)/TFA (0.1%)]. Sampleswere analysed by SELDI-TOF MS in the Ciphergen SELDI-TOFmass spectrometer (GE Healthcare). Spectra were calibrated againstCHCA peaks (643.360 D, 1059.5 D, and 1475.48 D). Peptide peakswith a signal-to-noise ratio >5 were identified using the CiphergenProteinChip Software v3.2.0, and used to identify proteins with theMascot search engine (www.matrixscience.com; Perkins et al.,1999). The type of search performed was a peptide mass fingerprintsearch at the NCBInr database as on 15 June 2007 (Viridiplantaeonly), with trypsin as enzyme, carbamidomethyl (C) as fixedmodification, oxidation (M) as variable modification, using averagemass values, a peptide mass tolerance of 61 Da, and a maximum ofone missed cleavage. LC-MS/MS results were obtained from theMcGill proteomics portal online (http://portal.proteomics.mcgill.ca/portal). An LC-MS/MS ion search was performed using the Mascotsearch engine and a database containing all available nucleotidesequences as on 31 January 2007 in order to find proteinhomologues, with search specifications of trypsin as enzyme,carbamidomethyl (C) as fixed modification, oxidation (M) asvariable modification, using monoisotopic mass values, a peptidemass tolerance and fragment mass tolerance of 60.5 Da, andmaximum of one missed cleavage.
Statistical analysis
The data was statistically analysed using parametric tests ata stringency of P < 0.05. The significance of variation in meanvalues for growth parameters and pigment and protein determi-nations was determined using a t test. The significance of the datafor immunogold labelling measurements was analysed usingANOVA and Tukey HSD tests.
Results
The expression of OC-1 in transgenic tobacco plantsdecreased plant growth, development rate, and protected
photosynthesis from chilling-induced inhibition (Van derVyver et al., 2003). These effects were previouslydocumented in three independent transgenic lines com-pared to the wild type and empty vector controlsconfirming that the slower development, growth, anddelayed senescence traits were linked to the expression ofthe transgene as was the protection of photosynthesis fromchilling-induced inhibition (Van der Vyver et al., 2003).Since Van der Vyver et al. (2003) demonstrated un-equivocally that the altered traits under investigation arerelated to the expression of the transgene, the presentstudy focused on the mechanisms by which altered leafCP activity influences leaf protein composition, photosyn-thesis, Rubisco protein content and activity, and leaf andplant senescence in one transgenic line (line T4/5)compared to wild-type controls.
Leaf protein composition and turnover
To determine whether leaf protein composition wasmodified in the OCE plants, leaf proteins were extractedfrom the youngest mature leaves (number 16) of 8-week-old control and OCE plants and separated using 2-D gelelectrophoresis (Fig. 1). Leaf proteins were extracted andprecipitated by standard proteomic procedures, in whichthe Rubisco large subunit (LSU) has only limitedsolubility (Ramagli, 1999). Since Rubisco generallyaccounts for 30–60% of total soluble proteins in theleaves of C3 species, it is important to use this selectiveprocedure to limit the amount of the Rubisco LSU on thegels, so that other proteins of lower abundance are notobscured.The Phoretix 2-D gel analysis software identified 765
protein spots in the extracts from control leaves and 860protein spots in extracts from OCE leaves. Key parameters(spot volume, pI, and MW) were calculated for all spots.Fifty-one spots were chosen for more intensive character-ization based on visible differences in spot volume. Of the51 spots, 13 were not statistically different in volumebetween C and OCE plants, seven spots had significantlygreater volume in C plants, 26 spots had significantlygreater volume in OCE plants, two spots were below thelevel of detection in OCE plants, and three spots wereonly detected in OCE plants (see Supplementary Table 1at JXB online). Two spots showing a difference in volume(Fig. 1, upper panels, spots 4 and 5) were identified usingSELDI-TOF MS. These proteins were highly homologousto Rubisco activase 2 (accession number Q40565) (spot 4)and Rubisco activase (accession number 1909374A) (spot5) (Table 1). Spot 4 also showed significant homology toRubisco activase (accession number number 1909374A)and Rubisco activase 1 (accession number Q40460), whilespot 5 showed significant homology to Rubisco activase 1(accession number Q40460) and Rubisco activase 2(accession number Q40565). The normalized volumes forspots 4 and 5 in OCE extracts were, respectively, 2.42 and
1938 Prins et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018

2.99 times greater than those found in C extracts. In OCEprotein extracts, spot 4 had a larger volume than spot 5(1.3 times).To characterize the Rubisco activase isoforms present in
these studies further, alignments were performed withthree GenBank tobacco Rubisco activase sequences andtwo Arabidopsis Rubisco activase isoforms (Table 1; seeSupplementary Fig. 1 at JXB online). While the highestscores for spots 4 and 5 were Rubisco activase 2 andRubisco activase from tobacco, the comparison withArabidopsis revealed the absence of the C-terminal amino
acids characteristic of the long isoform of this gene inArabidopsis. Instead of the final 36 C-terminal aminoacids present in the long Arabidopsis isoform, theArabidopsis short isoform has only eight amino acids(TEEKEPSK: Werneke et al., 1989), a difference that isconsidered to result from alternative splicing. The Arabi-dopsis large isoform has a MW of 46 kDa while the smallisoform is approximately 43 kDa. Spot 4 has the highesthomology to NTRA2 (RA2; see Supplementary Fig. 1 atJXB online) which lacks the C-terminal amino acids FAS.Spot 5 had the highest homology to another identified
Fig. 1. The effect of inhibition of CP activity leaf protein on tobacco and composition. Proteins were extracted from leaf 16 of control and OCEplants at 8 weeks and were separated on bi-dimensional gels. Proteins with major differences in abundance are indicated (1–51) in upper panels. Theposition of the proteins with the greatest differences: two Rubisco activase forms (NTRA and NTRA2), histone 4 (H4) and putative pepyidylprolylisomerise (PPI) are indicated in the lower panels.
Cysteine protease activity and Rubisco turnover 1939
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018

tobacco Rubisco activase (1909374A, RuAct; see Supple-mentary Fig. 1 at JXB online). This form lacks the first 59amino acids in the N-terminus and also contains threeadditional amino acids at the C-terminal (FAS) whencompared to RA2. Spot 5 also shows high homology withNTRA1 (RA1; see Supplementary Fig. 1 at JXB online)which has the full-length N-terminal sequence but also hasthe extra amino acids at the C-terminal.Spot number 23 on Fig. 1 upper panels has very low
abundance in the OCE proteome (see Supplementary Table1 at JXB online) and was identified by LC-MS/MS analysisto be highly homologous to volvox histone H4 (P08436),histone H2A.3 from wheat (HSWT93), and rice H2Aprotein (AAF07182) (Table 1). Spot number 24 in theOCE proteome, which is below detection in C extracts, wasidentified by LC-MS/MS and was significantly homolo-gous to rice Os05g0103200 (NP_001054392), which isdescribed as a chloroplast precursor (EC 5.2.1.8) ofpeptidyl-prolyl cis-trans isomerase TLP20. This proteincontains a cyclophilin ABH-like region. Spot 24 is alsosignificantly homologous to an Arabidopsis peptidylprolylisomerase-like protein (CAC05440), which also has a strongsimilarity to the chloroplast stromal cyclophilin, ROC4.The relative abundance of the Rubisco LSU and
Rubisco activase proteins was determined in leaves atdifferent positions on the stem of 14-week-old plants(Fig. 2) using western blot analysis. In the control plants,the amount of Rubisco LSU protein was highest in themature source leaves and least abundant in the youngest(18) and oldest (1) leaves (Fig. 2). However, the relativeabundance of the Rubisco LSU protein was much higherin the leaves of the OCE plants at all ranks on the stem,even in the oldest leaves (Fig. 2). Inhibition of leaf CPactivity resulted in a development-dependent difference in
the Rubisco activase protein bands (Fig. 2). Two bands ofRubisco activase protein were observed on western blotsusing specific antibodies in all but the oldest senescentleaves of the control plants where only the lower bandwas detected (Fig. 2). In marked contrast, only the highermolecular weight band of Rubisco activase protein wasdetected in the young leaves of OCE plants, with twobands becoming evident only in the oldest leaves fromleaf rank 10 and below (Fig. 2). It is important to note thatwe cannot directly relate the two bands of Rubiscoactivase protein observed after SDS-PAGE in Fig. 2 tothose observed on 2-D gels in Fig. 1. For example, thespots characterized after 2-D gel electrophoresis in Fig. 1are of similar molecular weights. A more exhaustiveanalysis of the different Rubisco activase proteins sepa-rated by 2-D gel electrophoresis is required as otherRubisco activase proteins are probably present.
Table 1. Identification of protein spots showing different abundance in control and OCE lines after 2D electrophoresis
Peptide fingerprint analysis (NTRA2 and NTRA) and/or ion analysis (H4 and PPI) using the Mascot search engine was used to establish proteinidentities.
Spot Identificationmethod
Accession Protein name Score e-value Queriesmatched
Peptide sequence(MS/MS)
NTRA2 (4) SELDI-TOF MS Q40565 Rubisco activase 2 (RA 2) 98 6.80E-05 91909374A Rubisco activase 75 1.30E-02 8Q40460 Rubisco activase 1 (RA 1) 71 3.30E-02 8
NTRA (5) SELDI-TOF MS 1909374A Rubisco activase 117 7.70E-07 15Q40460 Rubisco activase 1 (RA 1) 108 6.10E-06 15Q40565 Rubisco activase 2 (RA 2) 94 1.40E-04 13
H4 (23) LC-MS/MS P08436 Histone H4 195 3 ISGLIYEETRDNIQGITKPAIRTVTAMDVVYALK
HSWT93 histone H2A.3 48 1 AGLQFPVGRAAF07182 H2A protein 48 1 AGIQFPVGR
PPI (24) LC-MS/MS NP_001054392 Os05g0103200 357 16 TFKDENFKDFMIQGGDFDKVYFDISIGNPVGKHVVFGQVIEGMDIVKDFMIQGGDFDKGNGTGGK
CAC05440 peptidylprolyl 105 7 TFKDENFKisomerase-like protein
Fig. 2. Western blot analysis of the abundance of the Rubisco largesubunit, and Rubisco activase in leaves at different positions on thestem of 14-week-old plants. Soluble proteins were extracted from leavesat the positions on the stems as indicted, with leaf 1 being at the bottomof each plant and leaf 18 or 28 being the youngest mature leaf on thecontrol and OCE plants, respectively. 10 lg and 30 lg aliquots of leafprotein were loaded per well for the detection of Rubisco and Rubiscoactivase proteins, respectively.
1940 Prins et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018

Rubisco degradation and leaf CP activity
To determine whether tobacco Rubisco is susceptible todegradation by endogenous tobacco CPs in vitro assays wereconducted comparing Rubisco degradation in OCE extractswith that in C extracts in the absence or presence of the CPinhibitor, E64 (Fig. 3A). Rubisco was protected fromdegradation by endogenous CPs in OCE extracts comparedto control extracts; an effect that could be mimicked byinclusion of the CP inhibitor, E64, in the assays of thecontrol extracts (Fig. 3A). The CP activities of leaf extracts,scrutinized by activity staining after SDS-PAGE, revealedthat OCE leaves had much higher CP activities than controls(Fig. 3B), indicating the presence of feedback modulation ofCP expression that enhances CP production when activity isimpaired by constitutive cystatin expression.
Natural senescence and chilling-dependent inhibitionof photosynthesis, decreased Rubisco content andactivity
Dark chilling inhibited photosynthesis in a comparablemanner in three independent transgenic lines relative to
the wild type and empty vector controls and that effects ofconstitutive OC-1 expression on parameters such as theCO2 saturated rates of photosynthesis (Jmax) and apparentcarboxylation efficiency (ACE) were linked to expressionof the transgene (Van der Vyver et al., 2003). Leaf ACEand Jmax values attained maximal values 2 weeks afteremergence in both OCE and control plants (Table 2). Toinvestigate the effect of decreased CP activity on leafsenescence as determined by the age-dependent decreasein photosynthesis, leaf ACE and Jmax values weremeasured on the same leaves from 2–6 weeks (Table 2).The OCE leaves had greater ACE and Jmax values thancontrols at equivalent stages of development. Moreover,the senescence-related decline in photosynthesis wasdelayed in the OCE leaves (Table 2).Photosynthesis (Fig. 4A), extractable Rubisco activities,
and activation states (Fig. 4B) were compared in the leavesof 6-week-old plants maintained at either optimal growthtemperatures or exposed to seven consecutive nights ofchilling. At the beginning of the experiment (day 0), fullyexpanded leaves of control (Fig. 4A, upper panel) and OCEplants (Fig. 4A, lower panel) grown at optimal temper-atures had very similar rates of photosynthesis. At thisstage the OCE plants had higher total Rubisco activities butlower activation states than controls (Fig. 4B). Seven dayslater, the control leaves maintained at optimal temperatureshad about 20% lower photosynthetic rates than the OCEplants (Fig. 4A). Over the same period, total Rubiscoactivity in the control leaves maintained at optimal temper-atures had also decreased (Fig. 4B) with significant effecton Rubisco activation state (Fig. 4B). While Rubiscoactivities were similar in OCE plants at both time pointsthe Rubisco activation state was slightly increased at day 7(Fig. 4B). The chilling-dependent increase in Rubiscoactivation state in the OCE lines is surprising given thatthe overall rate of photosynthesis declined and that theinitial slope of the photosynthesis: intercellular CO2 re-sponse is also slightly decreased. However, these effectsare very small compared to the large effect of chilling onphotosynthesis rates in the control line, where Rubiscoactivation state was unchanged.The leaves of different independent OCE lines contain
about 20% more total soluble protein than those ofcontrols at 6 weeks old (Van der Vyver et al., 2003).Consistent with this observation, the amounts of RubiscoLSU and SSU proteins (Fig. 4C) were similar in theleaves of 6-week-old control and OCE plants at this stage.However, dark-chilling stress led to a pronounced de-crease in the abundance of Rubisco LSU and SSUproteins (Fig. 4C) in the leaves of control plants. Incontrast, dark-chilling had no effect on the amount ofdetectable Rubisco LSU and SSU proteins in the OCEplants. Similar trends were observed in the contents ofRubisco activase protein, but not in the glutaminesynthetase protein (Fig. 4C).
Fig. 3. Protection of Rubisco from degradation by OC-1 in OCE plantsand by E64 in control (C) plants in vitro assays. (A) The abundance ofthe Rubisco holoenzyme protein was detected in soluble protein extractsfrom 4-week-old control (C) and OCE plants on non-denaturing PAGEgels stained with Coomassie Brilliant Blue. (B) In-gel activity assayshowing degradation of the gelatine substrate by endogenous proteinasesfrom extracts of leaves of 14-week-old OCE and control (C) plants.Extracts were prepared from leaves at the bottom (3), middle (8, 14), andtop (18, 27) leaf ranks. Equal amounts of soluble protein (40 lg per well)extracted from C and OCE plants were compared in all instances.
Cysteine protease activity and Rubisco turnover 1941
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018

Intracellular localization of Rubisco protein inchloroplasts and vesicular bodies in the palisade cellsof young leaves
Electron microscopy and immunogold labelling withspecific polyclonal antibodies to the Rubisco LSU wereused to determine the intracellular distribution of theRubisco protein in the youngest mature leaves of controland OCE tobacco at 6 weeks old (Fig. 5). Label wasdetected in the chloroplasts of the palisade cells of control(Fig. 5B) and OCE leaves (Fig. 5C). In addition, Rubiscoprotein was also observed in vesicular bodies outside thechloroplast (Fig. 5B, C). The relative amounts of labelwere quantified in the chloroplasts and in the Rubiscovesicular bodies (RVB) of both control and OCE leave(Table 3). No differences were observed in the relativelocalization of Rubisco protein in the chloroplasts relativeto the RVBs of both control and OCE leaves (Table 3).
Intracellular localization of OC-1 protein in the cytosol,chloroplasts, and vacuoles in the palisade cells ofyoung leaves
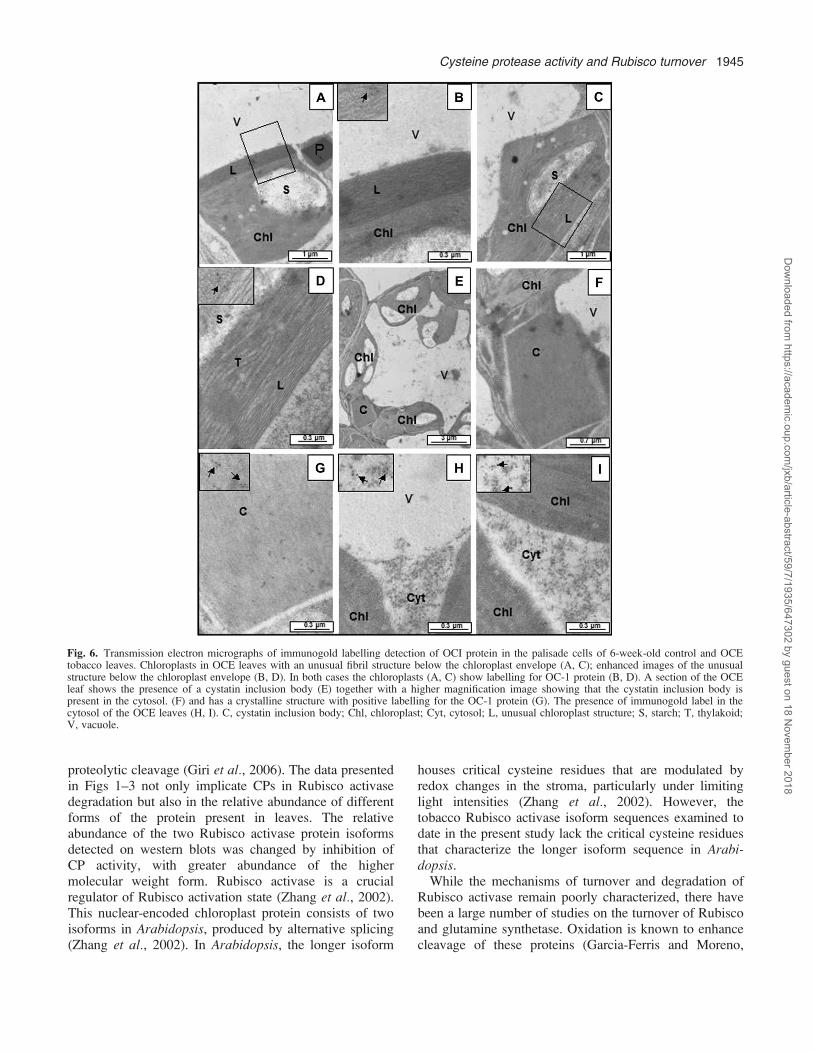
Electron microscopy and immunogold labelling withspecific polyclonal antibodies to the OC-1 protein wereused to determine the intracellular distribution of the OC-1protein in the youngest mature leaves of control and OCEtobacco at 6 weeks old (Fig. 6). The OC-1 protein wasmainly located in the cytosol which had the highestrelative gold particle concentrations (7168 lm2; n¼9).However, label was also detected in the vacuole at a goldparticle concentration of 5.562 lm2 (n¼9) and in thechloroplasts which had a gold particle concentration of20.664.2 lm2 (n¼9). Interestingly, the chloroplasts thatshowed imunogold labelling for the presence of the OC-1protein also had an alteration to the structure at theperiphery of the chloroplast either beneath or adjacent tothe chloroplast envelope (Fig. 6A, C). A higher magnifi-cation of the chloroplast periphery shows that thisstructure possibly has a fibrillar or membranous nature(Fig. 6B, D). These samples had not been fixed withosmium and therefore lipids were not stained in theseimages. This structure is highly stained, suggesting
perhaps that it might have a low lipid content. Thechloroplasts containing these structures clearly show label(Fig. 6B, D, inserts), but further studies are required toexplore the nature of this new structure and how it isformed by inhibition of CP activity.Some of the OCE cells also show the presence of
cytosolic inclusion bodies (Fig. 6E, F), which hasa crystalline structure (Fig. 6G). This structure containslabel (Fig. 6G, insert). It is tempting to suggest that it isformed by the strong interaction of the OC-1 protein withthe endogenous cytosolic CPs, because similar inclusionbodies in the cytosol have been observed previously in trans-genic plants expressing a wound- and methyl jasmonate-inducible 87 kDa tomato cystatin (Madureira et al., 2006).The high level of label in the cytosol (Fig. 6H, I, inserts)is consistent with the mode of expression of the OC-1protein in these studies, where the protein lackedsequences for specific organellar targeting.
Inhibition of CP activity effects on lifespan and leafprotein and chlorophyll contents after flowering
The OCE plants have a slow growth phenotype comparedto wild type or empty vector controls (Van der Vyveret al., 2003). In the present experiments, the control plantsflowered at 58.3361.20 d, at which point vegetativegrowth ceased. The OCE plants also sustained vegetativegrowth until flowering, but in this case, vegetativedevelopment ceased at 80.6761.45 d (Fig. 7). Hence, atthe point where the OCE lines reached sexual maturity (14weeks in the OCE lines) the OCE lines were much taller(Fig. 7A), with greater numbers of larger and heavierleaves than the controls (Fig. 7B–H). The effects ofinhibition of CP activity on leaf protein accumulationwere much more pronounced in 14-week-old (Fig. 8A)than they were in 6–8-week-old tobacco plants (Van derVyver et al., 2003). The increase in leaf protein in OCEplants that had flowered depended on the position on thestem (Fig. 8A). Chlorophyll was also increased but onlyin the youngest tobacco leaves (Fig. 8B). Maximalextractable leaf CP activities were greatly decreased inOCE leaves compared to controls at all positions on the
Table 2. Senescence-related decreases in apparent carboxylation efficiency (ACE) and CO2 saturated rates of photosynthesis (Jmax)in wild-type controls (C) and OCE tobacco leaves
Measured ACE and Jmax values, which were highest in both lines 2 weeks after leaf emergence, were measured in the same leaves for up to 6 weeks.The values represent the means 6SE of four replicates per experiment.
Time after leaf emergence C plants OCE plants C plants OCE plants(weeks) ACE ACE Jmax Jmax
(mol m�2 s�1) (mol m�2 s�1) (lmol m�2 s�1) (lmol m�2 s�1)
2 0.07860.004 0.10760.007 20.361.0 23.260.83 0.06960.004 0.11060.013 15.161.0 21.761.14 0.04060.004 0.06460.002 13.661.3 18.561.75 0.03460.004 0.06360.005 9.660.6 16.660.66 0.01460.001 0.04560.003 3.160.6 11.1960.1
1942 Prins et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018

Fig. 4. Effects of dark chilling on photosynthesis, Rubisco activity and activation state and relative abundance of Rubisco, Rubisco activase, andglutamine synthetase in the leaves of 6-week-old OCE and control tobacco plants. CO2 response curves for photosynthesis (A) in control andOCE leaves were measured at day 1 (closed circle), after 7 d of growth under optimal conditions (Opt: closed square) and after 7 nights of darkchilling (Chilled; open square). Initial and total Rubisco activities and the Rubisco activation state in control and OCE leaves measured at day 1,after 7 d of growth under optimal conditions and after 7 nights of dark chilling (B). Immunodetection of Rubisco, Rubisco activase, and glutaminesynthetase (C) in soluble protein extracts from control and OCE leaves at the beginning of the experiment (lanes 1 and 3) and after 7 nights ofdark chilling (lanes 2 and 4).
Cysteine protease activity and Rubisco turnover 1943
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018

stem (Fig. 8C), suggesting that the OC-1 remains boundto the CP during the extraction and spectrophotometricassay procedures, whereas it is removed by the in-gelassay methods used in Fig. 3B.
Discussion
The results presented here demonstrate that chloroplastproteins, particularly Rubisco and Rubisco activase, aremajor targets of leaf CPs. Moreover, the degradation ofRubisco involves interactions with the cytosol, as it can beblocked by endogenous cytosolic CP inhibitors such asOC-1. The demonstration of Rubisco-containing vesiclesoutside the chloroplast suggests a link between the proteinturnover machinery in the chloroplasts, cytosol, andvacuoles. A detailed discussion of the results and logiccorroborating the above conclusions is provided below.While there are relatively few reports on tobacco leaf pro-
teomics in the literature (Cooper et al., 2003; Franceschettiet al., 2004; Giri et al., 2006), the technique has beenused successfully to study the proteome of leaf plasmamembranes (Rouquie et al., 1997), trichomes (Ammeet al., 2005), and apoplast (Dani et al., 2005). Proteomeinformation is available for tobacco BY2 cell suspensioncultures (Laukens et al., 2004) and for plastids isolatedfrom these cultures (Baginsky et al., 2004). Rubiscoactivase sequences (Q40565, CAA78703, or 1909374A)have been identified previously in Nicotiana attenuataleaves (Giri et al., 2006). In this study concerning proteinselicited by oral secretions from Manduca sexta, sevenspots with homology to Rubisco activase were identified(Giri et al., 2006). Of these, four spots had kDa/pI valuessimilar to reported regulators of complementary activationproteins, with comparable molecular weights but differentpI values. In the present study, spots 4 and 5 in Fig. 1occupy similar positions to the Rubisco activase spotsreported by Giri et al. (2006). Differences in the positionsof the Rubisco activase spots on the gels might arise from
Fig. 5. Immunogold labelling detection of Rubisco protein in palisadecells of 6-week-old control and OCE tobacco leaves. High magnifica-tion of a cross-section of a control leaf showing the structure of thepalisade cells with immunogold labelling (A), and higher magnificationimages of the intracellular structure showing the compartmention of thelabel in wild-type and OCE leaves (B, C). The presence of Rubiscoprotein in chloroplasts and vesicular bodies outside the chloroplast isindicated in (B) and (C). Areas of immunogold are indicated by blackarrows. CY, cytosol; IS, intercellular space; P, plastoglobulus; RVB,Rubisco vesicular body; S, stroma; SG, starch grain; T, thylakoidmembranes; V, vacuole.
Table 3. Quantitation of gold particles (GP) after immunogoldlabelling of Rubisco large subunit in ultrathin leaf sections of Cand OCE tobacco
Gold particles were counted in chloroplasts, Rubisco vesicular bodies(RVB) and cytosol. Values obtained were compared to samples(Control) of both lines in the absence of antibodies. Mean values 6SE(n¼30). The means were compared by analysis of variance and byusing the Tukey multiple range test at P <0.05. Significant differencesbetween treatments are indicated by an asterisk.
GP in chloroplast(lm�2)
GP in RVB(lm�2)
GP in cytosol(lm�2)
Control 0.460.2 0.260.1 0.460.2OCE 27.963.3* 31.466.5* 1.460.8C 26.762.8* 29.165.3* 1.460.7
1944 Prins et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018
proteolytic cleavage (Giri et al., 2006). The data presentedin Figs 1–3 not only implicate CPs in Rubisco activasedegradation but also in the relative abundance of differentforms of the protein present in leaves. The relativeabundance of the two Rubisco activase protein isoformsdetected on western blots was changed by inhibition ofCP activity, with greater abundance of the highermolecular weight form. Rubisco activase is a crucialregulator of Rubisco activation state (Zhang et al., 2002).This nuclear-encoded chloroplast protein consists of twoisoforms in Arabidopsis, produced by alternative splicing(Zhang et al., 2002). In Arabidopsis, the longer isoform
houses critical cysteine residues that are modulated byredox changes in the stroma, particularly under limitinglight intensities (Zhang et al., 2002). However, thetobacco Rubisco activase isoform sequences examined todate in the present study lack the critical cysteine residuesthat characterize the longer isoform sequence in Arabi-dopsis.While the mechanisms of turnover and degradation of
Rubisco activase remain poorly characterized, there havebeen a large number of studies on the turnover of Rubiscoand glutamine synthetase. Oxidation is known to enhancecleavage of these proteins (Garcia-Ferris and Moreno,
Fig. 6. Transmission electron micrographs of immunogold labelling detection of OCI protein in the palisade cells of 6-week-old control and OCEtobacco leaves. Chloroplasts in OCE leaves with an unusual fibril structure below the chloroplast envelope (A, C); enhanced images of the unusualstructure below the chloroplast envelope (B, D). In both cases the chloroplasts (A, C) show labelling for OC-1 protein (B, D). A section of the OCEleaf shows the presence of a cystatin inclusion body (E) together with a higher magnification image showing that the cystatin inclusion body ispresent in the cytosol. (F) and has a crystalline structure with positive labelling for the OC-1 protein (G). The presence of immunogold label in thecytosol of the OCE leaves (H, I). C, cystatin inclusion body; Chl, chloroplast; Cyt, cytosol; L, unusual chloroplast structure; S, starch; T, thylakoid;V, vacuole.
Cysteine protease activity and Rubisco turnover 1945
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018

1994). For example, chloroplastic glutamine synthetasehas been shown to be rapidly degraded under conditionsthat cause photo-oxidative stress in leaves and this hasbeen linked to oxidative carbonylation of histidineresidues on the glutamine synthetase protein (Palatniket al., 1999; Ishida et al., 2002). Oxidatively-modifiedproteins are generally not repaired and are removed byproteolytic degradation. Within the chloroplast, oxidationof critical cysteine residues enhances the binding of theRubisco protein to the chloroplast envelope membranes,marking the protein for degradation (Marın-Navarro andMoreno, 2006). While chaperone-mediated autophagypathways for oxidatively-modified proteins remain to bedemonstrated in plants, stress-induced non-specific auto-phagasitic pathways of protein degradation have beendescribed (Xiong et al., 2007). Once outside the chloro-plast, Rubisco occurs in the vesicular transport system andinvolves cytoplasmic vacuole-type compartments (Chibaet al., 2003). Thereafter, Rubisco degradation productsappear in the vacuoles (Huffaker, 1990). While thisprocess has only previously been considered to be
important in senescing leaves where chloroplast lysisoccurs (Mae et al., 1984; Ono et al., 1995; Hortensteinerand Feller, 2002), the data presented here in Table 3 andFig. 5 show that they are present even in young leaves.These results demonstrate the existence of cytoplasmicvacuole-type compartments that we have called Rubiscovesicular bodies (RVBs) even in young leaves. While theproperties of the RVBs and their interactions with othercompartments are not yet well known, circumstantialevidence as outlined below suggests their involvement inRubisco protein turnover.The data presented here allow the formulation of a new
model for Rubisco degradation of the type illustrated inFig. 9, where vesicles containing CPs continuouslyinteract with those from functional chloroplasts to removeproteins marked for degradation. Rubisco protein ispresent in vesicles as well as in the chloroplasts in bothyoung wild-type and OCE leaves (Fig. 5; Table 3).Interestingly, RVBs have recently been shown to arisefrom the stromules that are continuously produced bychloroplasts even in young leaves (Ishida et al., 2007).
Fig. 7. Comparisons of growth and development in OCE and control plants. Phenotypes of control and OCE plants are shown at (A) 8 weeks and(B) 14 weeks. The white vertical bar indicates a height of 0.5 m. After both genotypes had flowered at 14 weeks the following parameters weremeasured: plant height (C); leaf number (D); total leaf area (E); leaf weight (F); average area per leaf (G); average leaf weight per leaf area (H). Meanvalues 6SE (n¼3). Significant differences between control and OCE plants are indicated by asterisk (P <0.05); ns, not significant.
1946 Prins et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018

While the function of stromules remains a matter ofdebate (Foyer and Noctor, 2007), available evidencesuggests that stromules forge associations between chloro-plasts and other organelles including the vacuole. Thesimilar pattern of Rubisco labelling in the RVBs andchloroplasts observed in the present study is entirelyconsistent with the chloroplast–stromule origin of RVBs.The tobacco chloroplasts observed in the present studyshow protrusions that are like stromules (data not shown).The demonstration of chloroplast–stromule–RVB associa-tions together with the observations of OC-1 protein in the
chloroplasts and vacuoles provide new insights into howchloroplast Rubisco becomes available for degradation byCPs that are absent from the chloroplast. The binding ofoxidized Rubisco protein to the chloroplast envelope(Desimone et al., 1996) might facilitate the accumulationof the protein at the sites where stromules and hencevesicles form. The maximal extractable CP activity ofleaves is greatly decreased by the presence of OC-1 (Fig.8C) unless the cystatin is removed by procedures such asgel electrophoresis. These observations are consistent witha role for extra-chloroplastic CPs in Rubisco proteinturnover. While, the major targets for leaf CPs areconsidered to be vacuolar or cytosolic storage proteins,plant cells contain several different types of vacuole-likecompartments with distinct functions (Chrispeels andHerman, 2000). Vesicle trafficking has been traditionallyviewed as a housekeeping process, but recent findings inplant, yeast and animal cells show that it can also play animportant role in stress responses (Chrispeels and Her-man, 2000; Herman and Schmidt, 2004). The physiolog-ical and biochemical identity of vacuoles is largelydetermined by correct targeting of vesicles and their cargo(Vitale and Pedrazzini, 2005). Since the data presentedhere indicate that the vesicles operate in the routineturnover of chloroplast proteins such as Rubisco, thename RVB seems appropriate. RVBs might be formed aspart of the autophagasitic process and engulf cytosolic aswell as chloroplastic proteins, explaining how cystatins inthe cytosol can effectively inhibit vacuole CPs. The OC-1protein is found largely in the cytosol, but also in thevacuole and in the chloroplasts, consistent with this view.While the data presented here concerning RVBs are
limited to Rubisco, it may be that other chloroplastproteins such as Rubisco activase are also present in theRVBs. This aspect is currently under investigation,together with the role of CPs and cystatins in the
Fig. 8. Comparisons of leaf soluble protein and chlorophyll contentsand cysteine proteinase activities in 14-week-old OCE and controltobacco plants. Soluble protein content (A); chlorophyll content (B);cysteine proteinase (CP) activity (C). Open and closed symbolsrepresent control and OCE plants, respectively. Mean values 6SE(n¼3 for A and B; n¼1 for C).
Fig. 9. A hypothetical model for Rubisco degradation via autophagyand the plant vesicle trafficking system. AA, amino acids; CP, cysteineproteinase; ER, endoplasmic reticulum; OC-1, oryzacystatin-1; RVB,Rubisco vesicular body.
Cysteine protease activity and Rubisco turnover 1947
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018

formation of peripheral structure adjacent to the chloro-plast envelope that has been observed when CP activity isinhibited. To date, only the number of RVBs have beencounted in the leaves of young plants prior to flowering,and values appear to be similar regardless of leaf CPactivity. However, the number may increase as the leafdevelops and senesces. Despite the absence of thisinformation at present, our data suggest that the Rubiscoturnover cycle involving RVBs must have a feedbackinteraction with the chloroplast that requires CP activity,as inhibition of CPs allows the chloroplast to maintaina high level of Rubisco activity and photosynthesis uponexposure to stress. These interactions may be controlledby redox regulation. In the control plants CP activity isregulated by cellular reductants such as glutathione. Themuch decreased CP activity in the OCE lines will allowtime for the re-reduction of reversibly oxidized aminoacids on the Rubisco protein such as cysteine andmethionine residues by thiol-disulphide exchange modu-lators, liberating them from the chloroplast envelope andallowing return of the repaired protein to the stroma forphotosynthetic purposes. While many gaps remain be-tween the hard facts and the hypothetical model describedin Fig. 9, this scheme provides a simple dynamicmechanism for the turnover of chloroplast proteins outsidethe plastid that can be tested readily with appropriatemolecular and cell biology tools.The processes that regulate plant development and
senescence have been widely studied, the concept ofregulation of plant lifespan is very poorly developed incomparison to animals, where a reduction in nutrientintake or dietary restriction extends lifespan in species asdiverse as worm (C. elegans), fly (D. melanogaster), yeast(S. cerevisiae), and mouse (Mus musculus). In theseorganisms, an alteration of nutrient sensing via modula-tion by insulin-signalling genes has been shown to extendlifespan. Evolutionary theory suggests that nutrient sens-ing regulates key genetic pathways that shift the metabolicinvestment that any organism makes from reproductionand growth toward somatic maintenance, allowing theorganism to survive under harsh environments until thereare suitable reproductive conditions. While these theoriesremain to be tested in plants, the data presented here showthat the lifespan of tobacco plants can be extended byinhibition of CP activity and that this manipulation alsoenables the plants to increase biomass production prior tosexual maturity and to avoid stress-induced prematuresenescence.
Supplementary data
Supplementary data can be found at JXB online.Supplementary Table 1. 2-D PAGE analysis of the
leaf proteome of control and OCE tobacco leaves (leaf
16). Normalized spot volume, isoelectric point (pI),molecular weight (MW), and significant differences inspot volume was calculated by Phoretix 2-D Expressionv2005 software from reference gels generated for bothcontrol and OCE tobacco plants (n¼3 for both lines). Allnumbers represent: mean value (SD). Spots are numberedaccording to Fig. 3 in the text.
Supplementary Fig. 1. Pair-wise alignment of RuBiSCOactivase isoforms from Arabidopsis and threeRuBiSCO activase proteins identified in tobacco. RA1,RuBiSCO activase 1 (tobacco); RA2, RuBiSCO activase2 (tobacco); RuAct, RuBiSCO activase (tobacco); AthL,RuBiSCO activase long isoform (Arabidopsis); AthS,RuBiSCO activase short isoform (Arabidopsis).
Acknowledgements
This work was funded by a Royal Society (UK)–National ResearchFoundation (South Africa) joint project (GUN 2068793). AP thanksthe Association of Commonwealth Universities for a Split-site PhDfellowship. We are grateful to Dominique Michaud for criticalreading of the manuscript.
References
Alonso JM, Granell A. 1995. A putative vacuolar processingprotease is regulated by ethylene and also during fruit ripening incitrus fruit. Plant Physiology 109, 541–547.
Amme S, Rutten T, Melzer M, Sonsmann G, Vissers JPC,Schlesier B, Mock H-P. 2005. A proteome approach definesprotective functions of tobacco leaf trichomes. Proteomics 5,2508–2518.
Baginsky S, Siddique A, Gruissem W. 2004. Proteome analysis oftobacco bright yellow-2 (BY-2) cell culture plastids as a modelfor undifferentiated heterotrophic plastids. Journal of ProteomeResearch 3, 1128–1137.
Barrett AJ. 1980. Fluorimetric assays for cathepsin B andcathepsin H with methylcoumarylamide substrates. BiochemicalJournal 187, 909–912.
Basset G, Raymond P, Malek L, Brouquisse R. 2002. Changes inthe expression and the enzymatic properties of the 20S proteo-some in sugar-starved maize roots. Evidence for an in vivooxidation of the proteosome. Plant Physiology 128, 1149–1162.
Belenghi B, Acconcia F, Trovato M, Perazzolli M, Bocedi A,Polticelli F, Ascenzi P, Delledonne M. 2003. AtCYS1, a cystatinfrom Arabidopsis thaliana, suppresses hypersensitive cell death.European Journal of Biochemistry 270, 2593–2604.
Beyene G, Foyer CH, Kunert KJ. 2006. Two new cysteineproteinases with specific expression patterns in mature andsenescent tobacco (Nicotiana tabacum L.) leaves. Journal ofExperimental Botany 57, 1431–1443.
Bhalerao R, Keskitalo J, Sterky F, et al. 2003. Gene expressionin autumn leaves. Plant Physiology 131, 430–442.
Botella MA, Xu Y, Prabha TN, Zhao Y, Narasimhan ML,Wilson KA, Nielsen SS, Bressan RA, Hasegawa PM. 1996.Differential expression of soybean cysteine proteinase inhibitorgenes during development and in response to wounding andmethyl jasmonate. Plant Physiology 112, 1201–1210.
Bradford MM. 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing the
1948 Prins et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018

principle of protein–dye binding. Analytical Biochemistry 72,248–254.
Callis J. 1995. Regulation of protein degradation. The Plant Cell 7,845–857.
Chiba A, Ishida H, Nishizawa NK, Makino A, Mae T. 2003.Exclusion of ribulose-1,5-bisphosphate carboxylase/oxygenasefrom chloroplasts by specific bodies in naturally senescing leavesof wheat. Plant and Cell Physiology 44, 914–921.
Chrispeels MJ, Herman EM. 2000. Endoplasmic reticulum-derived compartments function in storage and as mediators ofvacuolar remodelling via a new type of organelle, precursorprotease vesicles. Plant Physiology 123, 1227–1233.
Christou P, Capell T, Kohli A, Gatehouse JA,Gatehouse AMR. 2006. Recent developments and futureprospects in insect pest control in transgenic crops. Trends inPlant Science 11, 302–308.
Cooper B, Eckert D, Andon NL, Yates JR, Haynes PA. 2003.Investigative proteomics: identification of an unknown plant virusfrom infected plants using mass spectrometry. Journal of theAmerican Society for Mass Spectrometry 14, 736–741.
Dani V, Simon WJ, Duranti M, Croy RRD. 2005. Changes in thetobacco leaf apoplast proteome in response to salt stress.Proteomics 5, 737–745.
Desimone M, Henke A, Wagner E. 1996. Oxidative stress inducespartial degradation of the large subunit of ribulose-1,5-bisphosphatecarboxylase/oxygenase in isolated chloroplasts of barley. PlantPhysiology 111, 789–796.
Diop NN, Kidric M, Repellin A, Gareil M, d’Arcy-Lameta A,Pham Thi AT, Zuily-Fodil Y. 2004. A multicystatin is inducedby drought-stress in cowpea (Vigna unguiculata (L.) Walp.)leaves. FEBS Letters 577, 545–550.
Eason JR, Ryan DJ, Pinkney TT, O’Donoghue EM. 2002.Programmed cell death during flower senescence: isolation andcharacterization of cysteine proteinases from Sandersonia aur-antiaca. Functional Plant Biology 29, 1055–1064.
Feller U, Anders I, Mae T. 2008. Rubiscolytics: fate of Rubiscoafter its enzymatic function in a cell is terminated. Journal ofExperimental Botany 59, (in press).
Foyer CH, Noctor G. 2007. Shape-shifters building bridges?Stromules, matrixules and metabolite channelling in photorespira-tion. Trends in Plant Science 12, 381–383.
Foyer CH, Nurmi A, Dulieu H, Parry MAJ. 1993. Analysis oftwo Rubisco-deficient tobacco mutants, H7 and Sp25; evidencefor the production of Rubisco large subunits in the Sp25 mutantthat forms clusters and are inactive. Journal of ExperimentalBotany 44, 1445–1452.
Franceschetti M, Perry B, Thompson B, Hanfrey C,Michael AJ. 2004. Expression proteomics identifies biochemicaladaptations and defence responses in transgenic plants withperturbed polyamine metabolism. FEBS Letters 576, 477–480.
Garcia-Ferris C, Moreno J. 1994. Oxidative modification andbreakdown of ribulose-1,5-bisphosphate carboxylase oxygenaseinduced in Euglena gracilis by nitrogen starvation. Planta 193,208–215.
Gepstein S, Sabehi G, Carp M-J, Hajouj T, Nesher MFO,Yariv I, Dor C, Bassani M. 2003. Large-scale identificationof leaf senescence-associated genes. The Plant Journal 36,629–642.
Giri AP, Wunsche H, Mitra S, Zavala JA, Muck A, Svatos A,Baldwin IT. 2006. Molecular interactions between the specialistherbivore Manduca sexta (Lepidoptera, Sphingidae) and itsnatural host, Nicotiana attenuata. VII. Changes in the plant’sproteome. Plant Physiology 142, 1621–1641.
Granell A, Cercos M, Carbonell J. 1998. Plant cysteineproteinases in germination and senescence. In: Barrett AJ,
Rawlings ND, Woessner JF, eds. The handbook of proteolyticenzymes. San Diego: Academic Press, 578–583.
Gutierrez-Campos R, Torres-Acosta JA, Perez-Martınez JDJ,Gomez-Lim MA. 2001. Pleiotropic effects in transgenic to-bacco plants expressing oryzacystatin I gene. HortScience 36,118–119.
Harrak H, Azelmat S, Baker EN, Tabaeizadeh Z. 2001. Isolationand characterization of gene encoding a drought-induced cysteineprotease in tomato (Lycopersicon esculentum). Genome 44, 368–374.
Herman E, Schmidt M. 2004. Endoplasmic reticulum to vacuoletrafficking of endoplasmic reticulum bodies provides an alterna-tive pathway for protein transfer to the vacuole. Plant Physiology136, 3440–3446.
Hortensteiner S, Feller U. 2002. Nitrogen metabolism andremobilization during senescence. Journal of ExperimentalBotany 53, 927–937.
Huffaker RC. 1990. Proteolytic activity during senescence ofplants. New Phytologist 116, 199–231.
Humbeck K, Quast S, Krupinska K. 1996. Functional andmolecular changes in the photosynthetic apparatus duringsenescence of flag leaves from field-grown barley plants. Plant.Cell and Environment 19, 337–344.
Irving LJ, Robinson D. 2006. A dynamic model of Rubiscoturnover in cereal leaves. New Phytologist 169, 493–504.
Ishida H, Anzawa D, Kokubun N, Makino A, Mae T. 2002.Direct evidence for non-enzymatic fragmentation of chloroplasticglutamine synthetase by a reactive oxygen species. Plant. Celland Environment 25, 625–631.
Ishida H, Yoshimoto K, Reisen D, Makino A, Ohsumi Y,Hanson M, Mae T. 2007. Visualization of Rubisco-containingbodies derived from chloroplasts in living cells of Arabidopsis.Photosynthesis Research 91, 275–276.
Jensen ON, Wilm M, Shevchenko A, Mann M. 1999. Samplepreparation methods for mass spectrometric peptide mappingdirectly from 2-DE gels. Methods in Molecular Biology 112,513–530.
Kardailsky IV, Brewin NJ. 1996. Expression of cysteine proteasegenes in pea nodule development and senescence. MolecularPlant–Microbe Interactions 8, 689–695.
Keys AJ, Parry MAJ. 1990. Ribulose bisphosphate carboxylase/oxygenase and carbonic anhydrase. Methods in Plant Biochemistry3, 1–15.
Khan S, Andralojc PJ, Lea PJ, Parry MAJ. 1999. 2-carboxy-D-arabinitol 1-phosphate protects ribulose 1,5-bisphosphate carboxy-lase/oxygenase against proteolytic breakdown. European Journalof Biochemistry 266, 840–847.
Koizumi M, Yamaguchi-Shinozaki K, Tsuji H, Shinozaki K.1993. Structure and expression of two genes that encode distinctdrought-inducible cysteine proteinases in Arabidopsis thaliana.Gene 29, 175–182.
Laukens K, Deckers P, Esmans E, Van Onckelen H, Witters E.2004. Construction of a two-dimensional gel electrophoresisprotein database for the Nicotiana tabacum cv. Bright Yellow-2cell suspension culture. Proteomics 4, 720–727.
Lin JS, Wu SH. 2004. Molecular events in senescing Arabidopsisleaves. The Plant Journal 39, 612–628.
Ling J, Kojima T, Shiraiwa M, Takahara H. 2003. Cloning oftwo cysteine proteinases genes, CysP1 and CysP2, from soybeancotyledons by cDNA representational difference analysis. Bio-chimica et Biophysica Acta 1627, 129–139.
Linthorst HJM, Vanderdoes C, Brederode FT, Bol JF. 1993.Circadian expression and induction by wounding of tobaccogenes for cysteine proteinase. Plant Molecular Biology 21, 685–694.
Cysteine protease activity and Rubisco turnover 1949
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018

Lohman KN, Gan S, John MC, Amasino RM. 1994. Molecularanalysis of natural leaf senescence in Arabidopsis thaliana.Physiologia Plantarum 92, 322–328.
Madureira HC, Da Cunh M, Jacinto T. 2006. Immunolocaliza-tion of a defense-related 87 kDa cystatinin leaf blade of tomatoplants. Environmental and Experimental Botany 55, 201–208.
Mae T, Kai N, Makino A, Ohira K. 1984. Relationship betweenribulose bisphosphate carboxylase content and chloroplast num-ber in naturally senescing primary leaves of wheat. Plant and CellPhysiology 25, 333–336.
Marın-Navarro J, Moreno J. 2006. Cysteines 449 and 459modulate the reduction–oxidation conformational changes ofribulose 1�5-bisphosphate carboxylase/oxygenase and the trans-location of the enzyme to membranes during stress. Plant. Celland Environment 29, 898–908.
Masoud SA, Johnson LB, White FF, Reeck GR. 1993.Expression of a cysteine proteinase inhibitor (oryzacystatin-I) intransgenic tobacco plants. Plant Molecular Biology 21, 655–663.
Michaud D, Faye L, Yelle S. 1993. Electrophoretic analysis ofplant cysteine and serine proteinases using gelatin-containingpolyacrylamide gels and class-specific proteinase inhibitors.Electrophoresis 14, 94–98.
Okamoto T, Shimada T, Hara-Nishimura I, Nishimura M,Minamikawa T. 2003. C-terminal KDEL sequence of a KDEL-tailed cysteine proteinase (sulfhydryl-endopeptidase) is involvedin formation of KDEL vesicle and in efficient vacuolar transportof sulfhydryl-endopeptidase. Plant Physiology 132, 1892–1900.
Ono K, Hashimoto H, Katoh S. 1995. Changes in the number andsize of chloroplasts during senescence of primary leaves of wheatgrown under different conditions. Plant and Cell Physiology 36,9–17.
Palatnik JF, Carillo N, Valle EM. 1999. The role of photosyn-thetic electron transport in the oxidative degradation of chloro-plastic glutamine synthetase. Plant Physiology 121, 471–478.
Perkins DN, Pappin DJ, Creasy DM, Cottrell JS. 1999.Probability-based protein identification by searching sequencedatabases using mass spectrometry data. Electrophoresis 20,3551–3567.
Pernas M, Sanchez-Mong R, Salcedo G. 2000. Biotic and abioticstress can induce cystatin expression in chestnut. FEBS Letters467, 206–210.
Quirino BF, Normanly J, Amasino RM. 1999. Diverse range ofgene activity during Arabidopsis thaliana leaf senescenceincludes pathogen-independent induction of defense-relatedgenes. Plant Molecular Bioliogy 40, 267–278.
Ramagli LS. 1999. Quantifying protein in 2-D PAGE solubiliza-tion buffers. In: Link AJ, ed. Methods in molecular biology, Vol.112. Totowa, NJ: Humana Press Inc, 99–103.
Rintamaki E, Keys AJ, Parry MAJ. 1988. Comparison of the spe-cific activity of ribulose-1,5-bisphosphate carboxylase-oxygenasefrom some C3 and C4 plants. Physiologia Plantarum 74, 326–331.
Rivard D, Anguenot R, Brunelle F, Le VQ, Vezina L-P,Trepanier S, Michaud D. 2006. An in-built proteinase inhibitorsystem for the protection of recombinant proteins recovered fromtransgenic plants. Plant Biotechnology Journal 4, 359–368.
Rouquie D, Peltier JB, Marquis-Mansion M, Tournaire C,Doumas P, Rossignol M. 1997. Construction of a directory of
tobacco plasma membrane proteins by combined two-dimensionalgel electrophoresis and protein sequencing. Electrophoresis 18,654–660.
Schaffer MA, Fischer RL. 1988. Analysis of mRNAs thataccumulates in response to low temperature identifies a thiolprotease gene in tomato. Plant Physiology 87, 431–436.
Schmid M, Simpson D, Gietl C. 1999. Programmed cell deathin castor bean endosperm is associated with the accumulationand release of a cysteine endopeptidase from ricinosomes.Proceedings of the National Academy of Sciences, USA 96,14159–14164.
Shimada T, Hiraiwa N, Nishimura M, Hara-Nishimura I. 1994.Vacuolar processing enzyme of soybean that converts proproteinsto the corresponding mature forms. Plant and Cell Physiology 35,713–718.
Solomon M, Belenghi B, Delledonne M, Menachem E,Levine A. 1999. The involvement of cysteine proteases andprotease inhibitor genes in the regulation of programmed celldeath in plants. The Plant Cell 11, 431–444.
Swidzinski JA, Sweetlove LJ, Leaver CJ. 2002. A custommicroarray analysis of gene expression during programmed celldeath in Arabidopsis thaliana. The Plant Journal 30, 431–446.
Toyooka K, Okamoto T, Minamikawa T. 2000. Mass transport ofa proform of a KDEL-tailed cysteine proteinase (SH-EP) toprotein storage vacuoles by endoplasmic reticulum-derivedvesicle is involved in protein mobilization in germinating seeds.Journal of Cell Biology 148, 453–563.
Ueda T, Seo S, Ohashi Y, Hashimoto J. 2000. Circadian andsenescence-enhanced expression of a tobacco cysteine proteasegene. Plant Molecular Biology 44, 649–657.
Van der Vyver C, Schneidereit J, Driscoll S, Turner J,Kunert K, Foyer CH. 2003. Oryzacystatin-1 expression intransformed tobacco produces a conditional growth phenotypeand enhances chilling tolerance. Plant Biotechnology Journal 1,101–112.
Vitale A, Pedrazzini E. 2005. Recombinant pharmaceuticals fromplants: the plant endomembrane system as bioreactor. MolecularInterventions 5, 216–225.
Werneke JM, Chatfield JM, Ogren WL. 1989. AlternativemRNA splicing generates the two ribulose bisphosphate carboxy-lase/oxygenase activase polypeptides in spinach and Arabidopsis.The Plant Cell 1, 815–825.
Xiong Y, Contento AL, Nguyen PQ, Bassham DC. 2007.Degradation of oxidized proteins by autophagy during oxidativestress in Arabidopsis. Plant Physiology 143, 291–299.
Xu X-F, Chye L-M. 1999. Expression of cysteine proteinase duringdevelopmental events associated with programmed cell death inbrinjal. The Plant Journal 17, 321–328.
Yoshida T, Minamikawa T. 1996. Successive amino-terminalproteolysis of the large subunit of ribulose 1,5-bisphosphatecarboxylase/oxygenase by vacuolar enzymes from french beanleaves. European Journal of Biochemistry 238, 317–324.
Zhang N, Kallis RP, Ewy RG, Portis Jr AR. 2002. Lightmodulation of Rubisco in Arabidopsis requires a capacity forredox regulation of the larger Rubisco activase isoform.Proceedings of the National Academy of Sciences, USA 99,3330–3334.
1950 Prins et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article-abstract/59/7/1935/647302 by guest on 18 N
ovember 2018











![Mass Spectrometric Analysis of l-Cysteine Metabolism: … · tion of [U-13C3, 15N]L-cysteine to the culture, the levels of [13C3,15N]L-cysteine increased, and [13C3, 15N]L-cysteine](https://img.pdfslide.us/doc/110x75/5fe663421198753c202620ce/mass-spectrometric-analysis-of-l-cysteine-metabolism-tion-of-u-13c3-15nl-cysteine.jpg)

















