Embed Size (px)
Citation preview

ARTICLE OPEN ACCESS
CYP2C19 variant mitigates Alzheimer diseasepathophysiology in vivo and postmortemAndrea L Benedet MSc Lei Yu PhD Aurelie Labbe PhD Sulantha Mathotaarachchi MSc
Tharick A Pascoal MD Monica Shin MSc Min-Su Kang BSc Serge Gauthier MD Guy A Rouleau MD PhD
Judes Poirier PhD David A Bennett MD and Pedro Rosa-Neto MD PhD for the Alzheimerrsquos Disease
Neuroimaging Initiative
Neurol Genet 20184e216 doi101212NXG0000000000000216
Correspondence
Dr Rosa-Neto
pedrorosamcgillca
AbstractObjectiveTo verify whether CYP polymorphisms are associated with amyloid-β (Aβ) pathology acrossthe spectrum of clinical Alzheimer disease using in vivo and postmortem data from 2 in-dependent cohorts
MethodsA candidate-gene approach tested the association between 5 genes (28 single nucleotidepolymorphisms) and Aβ load measured in vivo by the global [18F]florbetapir PET standardizeduptake value ratio (SUVR) in 338 Alzheimerrsquos Disease Neuroimaging Initiative participantsSignificant results were then tested using plasma Aβ and CSF Aβ and Aβphosphorylated tau(Aβp-tau) ratio in the same cohort The significant association was also generalized topostmortem Aβ load measurement in the Rush Religious Orders StudyMemory and AgingProject cohorts In addition global cognition was used as a phenotype in the analysis in bothcohorts
ResultsAnalysis of Aβ PET identified a variant in the CYP2C19 gene (rs4388808 p = 00006) in whichcarriers of the minor allele (MA) had a lower global SUVR A voxel-wise analysis revealed thatthe variant is associated with a lower Aβ load in the frontal inferior temporal and posteriorcingulate cortices MA carriers also had higher CSF Aβ (p = 0003) and Aβp-tau ratio (p =002) but had no association with Aβ plasma levels In postmortem brains MA carriers hada lower Aβ load (p = 003) Global cognition was higher in MA carriers which was found to bemediated by Aβ
ConclusionsTogether these findings point to an association between CYP2C19 polymorphism and Aβpathology suggesting a protective effect of the MA of rs4388808 Despite the several possi-bilities in which CYP2C19 affects brain Aβ the biological mechanism by which this geneticvariation may act as a protective factor merits further investigation
From the Translational Neuroimaging Laboratory (ALB SM TAP MS M-SK PR-N) and AlzheimerrsquosDisease ResearchUnit (SG PR-N)McGill University ResearchCentre forStudies in Aging Montreal Canada CAPES Foundation (ALB) Ministry of Education of Brazil Brasılia Brazil Rush Alzheimerrsquos Disease Center (LY DAB) Rush University MedicalCenter Chicago IL Department of Decision Sciences (AL) HECMontreal Montreal Canada Department of Epidemiology (AL) Biostatistics ampOccupational HealthMcGill UniversityMontreal Canada Department of Neurology and Neurosurgery (GAR JP PR-N) Douglas Hospital Research Centre (JP PR-N) and Department of Psychiatry (JP) McGillUniversity Montreal Canada and Montreal Neurological Institute (GAR PR-N) Canada
Coinvestigators are listed at httplinkslwwcomNXGA21
Funding information and disclosures are provided at the end of the article Full disclosure form information provided by the authors is available with the full text of this article atNeurologyorgNG
The Article Processing Charge was funded by the authors
This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-NoDerivatives License 40 (CC BY-NC-ND) which permits downloadingand sharing the work provided it is properly cited The work cannot be changed in any way or used commercially without permission from the journal
Copyright copy 2018 The Author(s) Published by Wolters Kluwer Health Inc on behalf of the American Academy of Neurology 1
In sporadic Alzheimer disease (AD) accumulation of amy-loid-β (Aβ) seems to be associated with reduced Aβ clearancebut little is known about the biochemical pathways underlyingcerebral Aβ metabolism
Cytochrome P450 (CYP) is a family of enzymes known tometabolize several endogenous and exogenous substratesBesides metabolizing drugs brain CYPs are also involved inthe modulation of blood flow metabolism of fatty acidscholesterol and neurotransmitters and mobilization of in-tracellular calcium1ndash3 all pathways that have been somehowlinked to AD andor amyloid metabolism In addition studieshave shown that CYP proteins and their genetic variants areassociated with the brain immune response2 and neurode-generative diseases245
Although previous evidence shows CYP expression in areasaffected by AD such as the amygdala frontal and temporalcortices and the hippocampus67 there are only a few reportsdocumenting a link between CYPs and AD pathophysiologyIn one study the overexpression of human amyloid pre-cursor protein (APP) on the Tg2576 mice model was as-sociated with elevated CYP hepatic function while theirrenal counterparts were depressed8 In addition anotherstudy presented evidence that Aβ stimulates NADPH-cytochrome P450 reductase (POR) a CYP inducer in APPtransgenic mice and human AD brains which may affect theredox status9
In line with the evidence that Aβ can stimulate metabolic cas-cades in a similar fashion as foreign compounds910 andknowing that brain CYP could potentially affect amyloid me-tabolism also through other mechanisms we tested the hy-pothesis that variants in CYP genes could be associated with Aβand consequently have an impact in cognitive performanceWe examined the association of CYP variants with biomarkersof Aβ load in vivo and postmortem as further specified
MethodsStudy participantsDiscovery analysis data were obtained from the AD Neu-roimaging Initiative (ADNI) database (adniloniuscedu)For this study cognitively normal (CN) individuals hada Mini-Mental State examination (MMSE) score of 24 orhigher and a clinical dementia rating (CDR) of 0 The
operational criteria adopted for mild cognitive impairment(MCI) were participants who had an MMSE score equal toor greater than 24 a CDR of 05 subjective memory con-cern objective memory loss measured by education-adjusted scores on delayed recall of 1 paragraph fromWechsler Memory Scale Logical Memory II (ge16 years le88ndash15 years le4 and 0ndash7 years le2) and essentially normalactivities of daily living Participants with AD were individ-uals with an MMSE score lower or equal to 26 CDR higherthan 05 who met the National Institute of Neurological andCommunicative Disorders and StrokendashAlzheimerrsquos Diseaseand Related Disorders Association criteria for probable ADAll individuals had absence of any other neuropsychiatricdisorders The ADNI inclusionexclusion criteria are de-scribed in detail at adni-infoorg
Our report also used data from the Religious Orders Study(ROS) and Rush Memory and Aging Project (MAP) 2 longi-tudinal clinical pathologic cohort studies of aging anddementia1112 All participants enrolled without known dementiaand agreed to annual clinical and neuropsychological assess-ments and brain donations after death The clinical diagnosis ofdementia and AD followed the National Institute of Neuro-logical and Communicative Disorders and StrokendashAlzheimerrsquosDisease and Related Disorders Association recommendationsThe MCI diagnosis is given to participants who were rated asimpaired based on MMSE and on the Consortium toEstablish a Registry for Alzheimerrsquos Disease (CERAD) neuro-psychological measures but who were not found to have de-mentia by the examining neurologist Our study includes onlyMCI participants with 1 impaired domain and no other cause ofcognitive impairment Notably the studies are conducted by thesame team of investigators and share a large common core oftesting batteries which allow for combined analyses
The discovery sample consisted of a subset of ADNI partic-ipants who had [18F]florbetapir PET genetic data and CSFdata available The findings obtained with the discoverysample were then generalized to the Rush-ROSMAP par-ticipants who died at age 90 years or younger
Standard protocol approvals registrationsand patient consentsThis secondary analysis study followed all institutional reviewboard regulations which are detailed in the supplementarymethods
GlossaryAA = arachidonic acid Aβ = amyloid-β AD = Alzheimer disease ADNI = AD Neuroimaging Initiative APP = amyloidprecursor protein CDR = clinical dementia rating CN = cognitively normal CYP = cytochrome P450 EET =epoxyeicosatrienoic acid LD = linkage disequilibriumMA = minor alleleMAF = minor allele frequencyMAP =Memory andAging ProjectMCI = mild cognitive impairmentMMSE = Mini-Mental State Examination PHF = paired helical filament p-tau = phosphorylated tau ROS = Religious Orders Study SNP = single nucleotide polymorphism SUVR = standardizeduptake value ratio
2 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
PhenotypesFrom the ADNI we obtained demographics neurocognitivescores CSF and plasmatic data available after passing rigor-ous quality control The preprocessed imaging and geneticdata underwent additional processing and quality controlbefore the statistical analysis in our study as described belowFrom Rush all the data used in this project to perform sta-tistical analyses were already processed and quality controlledwith very rigorous criteria
In the ADNI cohort the brain Aβ load was estimated usingthe [18F]florbetapir PET standardized uptake value ratio(SUVR details are provided in figure 1 and e-methods)Separately Aβ and hyperphosphorylated tau (p-tau) lev-els were measured in the CSF and plasma These data wereobtained by the Biomarkers Consortium CSF and thedescription of the methodology regarding the sampleacquisition processing and analysis is available atthe ADNI website (adniloniuscedudata-samplesbio-specimen-data)
In the Rush-ROSMAP cohort postmortem data of the Aβload and paired helical filament (PHF) tau tangle density wereobtained from 8 brain regions using immunohistochemistry13
Cognitive function was evaluated in both the ADNI and Rush-ROSMAP For the ADNI the global cognition scores weredeveloped by making a z-score of the sum of the memory andthe executive function composite scores In the Rush-ROSMAP the global cognitive scores were based on 17 cognitiveperformance tests and we included only participants whounderwent autopsies thus the last valid cognitive scoreproximate to death was used for this analysis (for furtherdetails see e-Methods httplinkslwwcomNXGA20)
We used gene expression data to examine cis expression quan-titative trait loci (eQTL) For the ADNI total RNAwas obtainedfrom the peripheral bloodmdashat the same visit when the imagingdata were obtainedmdashand after quality control was hybridized tothe Affymetrix Human Genome U219 array plate as described14
The normalized and quality-controlled expression data14 wereused For the Rush-ROSMAP RNA was extracted from thegray matter of dorsolateral prefrontal cortex blocks usingthe miRNeasy Mini Kit (Qiagen Venlo Netherlands) and theRNase-free DNase Set (Qiagen) These samples were quantifiedand qualified as described15 and then sequenced on the IlluminaHiSeq with 101 bp paired-end reads The normalized andquality-controlled expression data were used In both analysesthe RNA integrity measurement was used as a covariate
Figure 1 [18F]florbetapir standardized uptake value ratio analytical method
Flowchart showing acquisition methods (purple) imageprocessing (blue) and outcomes (green) ADNI = Alz-heimerrsquos Disease Neuroimaging Initiative GM = graymatter ROI = region of interest SUVR = standardizeduptake value ratio WM = white matter
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 3
Genotyping imputation and gene selectionWe interrogated single nucleotide polymorphisms (SNPs)from the CYP genes of the 5mdashprobablymdashmost importantmetabolizers CYP3A4 CYP2D6 CYP2C9 CYP1A2 andCYP2C19316ndash18 Methods involved in DNA processing aredetailed in the supplementary methods section (httplinkslwwcomNXGA20) Using PLINK19 the SNPs within thesegenes were recoded based on the dominant model with re-spect to the MAs and the ones in high linkage disequilibrium(LD) (r2 gt 08) were removed from the analysis to avoidunnecessary testing A total of 28 SNPs were used in the initialstep of the association analysis
For Rush-ROSMAP cohorts DNA was extracted genotypedimputed and quality controlled according to the proceduresdescribed in e-Methods (httplinkslwwcomNXGA20)
Statistical analysisThe statistical tests were performed in R statistical software20
where linear regressionmodels were used to test the associationbetween the genotypes and the phenotypes in both cohortsseparately All statistical models included the covariates of agesex and APOE-e4 carriage status except for the models inwhich global cognition was the outcome measure where theAPOE-e4 status was replaced by years of education The initialanalysismdashusing the global [18F]florbetapir SUVR asa phenotypemdashwas performed to interrogate a total of 28 SNPs(table e-1 httplinkslwwcomNXGA19) To correct formultiple testing the statistical significance of the SNP discoverywas set at p lt 00017 using a Bonferroni correction for 28 testsand a type I error α = 005 The significant SNPs were furthertested for associations with other phenotypes in ADNI andRush-ROSMAP cohorts To test for group differences in thediagnostic status we added diagnosis as a covariate in the re-gression models and also tested the interaction between thediagnostic status and desired independent variable Effect sizecalculations (Cohen d) were also used
A voxel-based analysis was performed using the RMINCimaging tool21 where parametric images were obtained con-trasting the [18F]florbetapir SUVR between the genotypegroups of the SNP found in the discovery analysis Afterrandom field theory correction for multiple comparisons the tvalue threshold of significance was leminus31 (p le 0001)
Mediation analysis was used to examine whether the SNPassociations with cognition were mediated through the effect ofAβ To test this hypothesis first the association of CYP SNPswith global cognitive performance was evaluated using linearregression models Then if the association was significant themeasure of Aβ was added to this model to check whether theSNP associations with cognitionwere attenuated because of Aβ
ResultsDemographics and general information of the ADNI andRush-ROSMAP cohorts are presented in table 1 After
imaging and genetic quality control a total of 338 ADNIparticipants were studied including 186 CN 105 single- ormultiple-domain amnestic MCI and 47 AD A total of 738Rush-ROSMAP participants were analyzed including 301CN 179 amnestic MCI and 258 AD As expected in bothcohorts CN individuals had higher MMSE scores and fewerwere APOE-e4 carriers as compared to both MCI and ADThere was no difference in sex or years of formal educationbetween diagnostic groups in both cohorts (details are pro-vided in table e-2 httplinkslwwcomNXGA19)
CYP2C19 polymorphism is associated with theglobal [18F]florbetapir SUVR in the ADNI cohortThe discovery analysis tested the association of 28 SNPs fromCYP genes with the global [18F]florbetapir SUVR The anal-ysis unveiled an association of an SNP (rs4388808) in theCYP2C19 gene (t = minus343 p = 00006) (figure 2A) Theminor allele frequency (MAF) of rs4388808G in the ADNIcohort was 019 and the minor allele (MA) carriers displayedless [18F]florbetapir binding as compared to the noncarriersIt is important that the association remained significant whenadjusting the model for diagnosis In addition no interactionwas found between the SNP and diagnostic status The esti-mated effect size of rs4388808 in the whole sample wasCohen d = 036 Of interest the effect size was greater for thesubset of APOE-e4 carriers (Cohen d = 051) as compared toAPOE-e4 noncarriers (Cohen d = 033)
MA carriers of the CYP2C19 polymorphismhaveless Aβ in AD-related regions in theADNI cohortTo identify the brain areas responsible for the global differ-ence observed in [18F]florbetapir uptake revealed in the dis-covery analysis we performed a voxel-wise analysis Theanalysis showed that carriers of rs4388808G had less [18F]florbetapir binding in the frontal posterior cingulate andinferior temporal cortices (figure 3) with the voxels in theseregions presenting p values equal to or less than 0001
Findings with brain imaging are corroboratedby CSF data in the ADNI cohortBecause of the well-known relationship between brain and CSFlevels of Aβ we tested the association between rs4388808 andCSF Aβ Because not all participants in the ADNI sample hadCSF data we performed the analysis in a subset of 260 indi-viduals As expected MA carriers presented more CSF Aβ thannoncarriers (t = 294 p = 0003) (figure 2B) The CYP2C19polymorphismwas associatedwith the Aβp-tau ratio (t = 229p = 002) and higher ratios were found in carriers ofrs4388808G (figure 2C) No association was found using p-tauas a single outcome measure (t = minus124 p = 021)
Association between the CYP2C19polymorphism and Aβ is exclusive to the Aβbrain levels in the ADNI cohortTo examine whether the CYP2C19 polymorphism is also as-sociated with Aβ levels outside the brain we tested the
4 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
association of rs4388808 with plasmatic levels of Aβ ina subsample of 113 participants We did not find an associa-tion between the SNP and free (t = minus060 p = 054) or total(t = 049 p = 062) plasmatic Aβ1-42 levels (figure 2D) in-dicating that the findings reported above are specific tothe CNS
Results from theADNI cohortwere generalizedto the Rush-ROSMAP cohortWe next generalized the findings to postmortem indices of theAβ load and to neurofibrillary tangles in Rush-ROSMAPcohorts From the 8 brain regions where the Aβ load wasmeasured we selected 6 regions (angular gyrus anterior cin-gulate entorhinal cortex and mesial and inferior temporalcortices) and calculated an average per subject These 6 regionswere chosen to match with the brain areas where we detecteddifferences between genotype groups in the voxel-wise analysisWhen testing the association between the average of the Aβload and the CYP2C19 polymorphism we found thatrs4388808G (MAF = 015) carriers had a lower Aβ load thannoncarriers (t = minus215 p = 003) (figure 2E) consistent withfindings from the ADNI cohort The difference betweenrs4388808G carriers and noncarriers in PHF tau tangle densitydid not reach statistical significance (t = minus165 p = 009)
MA carriers of the CYP2C19 polymorphism hadbetter cognitive performance thannoncarriersin both cohortsTo examine whether the CYP2C19 polymorphism affects thecognitive performance we used composite measures of globalcognition as the outcome in linear regression models In theADNI cohort there was a clear tendency to an associationwith the CYP2C19 polymorphism (t = 192 p = 005) (figure2F) while in Rush-ROSMAP cohorts the association wassuch that rs4388808G carriers had a higher cognition thannoncarriers (t = 208 p = 003) (figure 2G)
The effect of the CYP2C19 polymorphism oncognition is mediated by AβA mediation analysis was used to determine whether theCYP2C19 polymorphism was directly or indirectly associatedwith cognitive performance We retested the association be-tween the CYP2C19 polymorphism and cognition but nowadding the measures of [18F]florbetapir or Aβ load in themodel Neither of the previous genotype-phenotype associa-tions remained significant indicating that the observed effectof the genotype was mediated by Aβ
Association between rs4388808 and geneexpression of CYP2C19 suggests that the SNPis functionalTo check whether the polymorphism has an effect in the geneexpression we tested the association between rs4388808 andCYP2C19 RNA levels In the ADNI cohort with 304 partic-ipants for this analysis we found a cis eQTL such that MAcarriers displayed higher RNA levels in the blood (t = 238 p =001) (figure 2H) In Rush-ROSMAP cohorts very low levelsof CYP2C19 expression were detected in postmortem brainsand the data did not provide consistent results (data notshown) as found in the blood expression data from the ADNI
DiscussionIn the present study we found an association between theCYP2C19 polymorphism and Aβ burden across the spectrumof AD in which carriers of rs4388808G presented a less Aβload and downstream cognitive impairment This associationwas initially detected using the [18F]florbetapir SUVR andposteriorly confirmed with CSF Aβ in the ADNI cohort Noassociation was found between the polymorphism and di-agnostic groups suggesting that the SNP is associated with anamyloid load rather than with the disease status Effect size
Table 1 Demographic and key characteristics of the samples
ADNI Rush-ROSMAP
CN MCI AD CN MCI AD
No of participants 186 105 47 301 179 258
Males n () 91 (489) 64 (61) 26 (553) 182 (605) 101 (564) 154 (597)
Age y mean (SD) 7570 (663) 7429 (812) 7567 (737) 837 (519)a3 8526 (435) 8595 (381)
APOE-laquo4 () 52 (28)a1 57 (543)a1 30 (638)a1 49 (164)a4 50 (279)a4 112 (434)a4
MMSE mean (SD) 2909 (114)a2 2724 (195)a2 2034 (366)a2 2869 (142)a5 2791 (183)a5 2549 (517)a5
Education y 1641 (275) 1667 (268) 1613 (247) 1653 (374) 1634 (347) 1666 (394)
MA n ()b 67 (19) 36 (10) 17 (5) 87 (11) 52 (7) 63 (85)
Abbreviations AD = Alzheimer disease ADNI = AD Neuroimaging Initiative CN = cognitively normal MA = minor allele MAP = Memory and Aging ProjectMCI = mild cognitive impairment MMSE = Mini-Mental State Examination ROS = Religious Orders Studya Statistically different from the other groups of the same sample 1p = 23 times 10minus7 2p = 2 times 10minus16 3p = 92 times 10minus8 4p = 17 times 10minus11 and 5p = 2 times 10minus16b MA (rs4388808G) percentage relative to its major sample (ADNI or Rush-ROSMAP)
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 5
analysis suggests that the benefic effects of the polymorphismare larger in APOE-e4 carriers as compared to noncarriers orthe whole sample in which the effect is similar to what hasbeen described for a BCHE gene polymorphism22 Sub-sequently this association was generalized to postmortemdata from the Rush-ROSMAP cohort Finally both cohortsshowed a protective association between the MA of CYP2C19and cognition It is important that subsequent mediationanalysis suggested that the effects of CYP2C19 on cognitionwere mediated by Aβ
Overall these results support the notion that the MA ofCYP2C19 (rs4388808) is a protective variant against Aβ anddownstream cognitive impairment Of interest voxel-wiseanalysis in in vivo participants revealed that the MA was as-sociated with reduced Aβ burden in AD-related regions in thefrontal inferior temporal and posterior cingulate cortices23
In addition the CYP2C19 SNP was associated with the CSFAβp-tau ratio but not with plasmatic Aβ Beyond the fact thatthe Aβp-tau ratio is postulated to better represent neuriticplaques than the single CSF Aβ information24 these results
Figure 2 Comparison between noncarriers (minus) and carriers (+) of the minor allele of rs4388808 (CYP2C19)
In the ADNI cohort a difference was observedin the brain Aβ load (A) CSF Aβ levels (B) andCSF Aβp-tau ratio (C) but no difference wasdetected in plasmatic levels of Aβ (D) Resultswere generalized to postmortem data of Rush-ROSMAP cohorts where a concordant patternwas observed in the Aβ load (percentage areaoccupied by amyloid) (E) A tendency towardsignificance in global cognitive performance (F)was found in the ADNI cohort while in theRush-ROSMAP it was significant (G) CYP2C19expression levels were also different betweennoncarriers (minus) and carriers (+) of the minorallele of rs4388808 (H) in the ADNI cohort Alllinear models were adjusted for age sex andAPOE-e4 carriage status with the exception ofthe models using global cognition and geneexpression which were adjusted for age sexand years of education or RNA integrity re-spectively Aβ = amyloid-β ADNI = AlzheimerrsquosDisease Neuroimaging Initiative MAP = Mem-ory and Aging Project p-tau = phosphorylatedtau ROS = Religious Orders Study SUVR =standardized uptake value ratio
6 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
provide further evidence that the polymorphism affects thebrain accumulation of neuritic plaques in a tissue- and disease-specific manner24 In both cohorts the referred poly-morphism was not associated with PHF tau or single CSFp-tau agreeing with the framework that Aβ and tau haveindependent upstream triggers25 In line with overall signifi-cant results the CYP2C19 polymorphism also presenteda protective effect in global cognition being supported byprevious studies that linked Aβ pathology to cognitivechanges13 Indeed the mediation analysis supported that theassociation between the genetic factor and cognitive abnor-malities is likely due to the upstream accumulation of Aβ
Levels of the CYP2C19 protein are associated with thers4388808 polymorphism26 which is an intronic variant of theCYP2C19 gene Indeed we found a similar association showingthat MA carriers expressed higher levels of CYP2C19 whencompared with noncarriers suggesting that the SNP affects theexpression of its gene To corroborate this finding we retrievedthe SNPs that were previously excluded because of high LD andverified that none was associated with CYP2C19 expression inthe blood (data not shown) reinforcing the idea that rs4388808is functional This association could not be detected in post-mortem data probably because of the low CYP expression orbecause of its limited detection by the method used7
Despite the SNP not being widely studied to provide in-formation about the association with other phenotypes other
polymorphisms in the gene have been correlated with meta-bolic variability atherosclerosis and behavioral traits27ndash29
CYP2C19 expression has been detected in several brainregions67 however its function in the CNS has not been fullyelucidated Some reports have suggested that besides its roleat metabolizing exogenous substrates CYP2C19 also partic-ipates in other important biological cascades such as themetabolism of serotonin and sexual hormones1 and the me-tabolism of the arachidonic acid (AA) where it functions as anepoxygenase30
The biochemical mechanism underlying the association ofCYP2C19 with Aβ remains speculative A study has shownthat Aβ is able to act as a foreign body and trigger PORactivating CYPs to initiate catabolic reactions9 In turnCYP2C19 is involved in the catalysis of estradiol31 Becauseseveral studies have reported the beneficial effects of estrogenson ADmdashdemonstrating that these steroids are able to act asanti-inflammatory compounds32 and even inhibit Aβproduction33mdashone could think that MA carriers of rs4388808have a reduced expression of this CYP in the brain or that theyhave an increased structural incompatibility betweenCYP2C19 and estradiol Consequently MA carriers couldpresent a reduction in the catabolism of estrogen and a re-duction in the Aβ load Alternatively the Aβ load would beaffected by the levels of epoxyeicosatrienoic acids (EETs)produced via the metabolism of AA by CYP epoxygenases Inthe brain studies have shown that EETs have beneficial effectsat regulating blood flow cortical angiogenesis and at pro-moting anti-inflammatory reactions (for review see reference2) By contrast when AA is metabolized via other cascadesthe protective effect of EETs is lost and there is a possibilityof induction of Aβ production andor accumulation34 Inaddition it was demonstrated that Aβ is able to reduceepoxygenase activity and consequently decrease EET pro-duction35 probably leading to more susceptibility to neuronaldamage (for review see reference 36) In this scenario itwould be possible that the MA of rs4388808 increases theexpression of CYP2C19 leading to an ldquoAβ-resistantrdquo epox-ygenase metabolism of AA with a maintained production ofEETs and a more protective phenotype against amyloidosis
It is also plausible to think however that the associationdescribed here between theCYP2C19 polymorphism and Aβis indirect being mediated by the effect of a drug CYP2C19is known to metabolize several drugs37 including medi-cations to treat depression which has been mentioned asa risk factor for AD (for review see reference 38) Cit-alopram for example is an antidepressant metabolized byCYP2C19 that has been shown to decrease Aβ production39
Thus one may think that the polymorphism rs4388808 isthen associated with an improved action of citalopramleading to the decreased brain Aβ burden rather than as-sociated with Aβ metabolism itself Because it is difficult totrack all the medications the study participants have takenbefore been enrolled it is unfeasible at the moment to checkwhether this association holds true
Figure 3 T-statistical parametric maps showing differencesbetween noncarriers (minus) and carriers (+) of theminor allele of rs4388808 (CYP2C19)
T-statistical parametric maps superimposed on average structural MRI showbrain regions with lower standardized uptake value ratio (SUVR) values inminor allele carriers (CYP2C19 (+)) of the polymorphism of CYP2C19 Statisticaldifferences overlap with brain regions vulnerable to Alzheimer disease path-ophysiology such as the posterior cingulate frontal and temporal cortices
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 7
The interpretation of the results should take into consider-ation a few limitations The use of the 2 cohorts does not allowus to detect and measure Aβ pathology with the samemethodology inserting in the analysis some variability specificto the method used However despite being measured withdifferent techniques both the imaging radiotracer and theantibody used in the brain tissue are expected to bind to thefibrillar form of Aβ13 Similarly the cognitive composite scoresfrom the 2 cohorts have some differences that should beconsidered There are also some intrinsic characteristics of thesamples that have to be taken into account (1) the inclusioncriteria of each study (2) the average age difference betweenthe studies and (3) and the disease stage in which Aβmeasures were performedmdashat the end or during the diseaseprocess Sample size restrictions impose some degree ofcaution when interpreting findings including for examplethe lack of association between the genotype and plasmaticlevels of Aβ or cognition in the ADNI cohort In additionboth cohorts are mostly composed of non-Latino Caucasianslimiting the extrapolation of the present findings to otherpopulation groups Longitudinal analysis together with func-tional genomics and biochemistry experiments not per-formed here would also be necessary to determine the effectof the SNP in the protein function as well as to support anytheoretical framework proposed in the Discussion section
Results obtained from the 2 independent cohorts providecompelling evidences linking the CYP2C19 polymorphismand Aβ pathology suggesting that the MA of rs4388808confers protective effects against Aβ accumulation in the brainand its downstream cognitive consequences These resultscould have implication for antiamyloid clinical trial designs aspreclinical rs4388808G would present a protective factoragainst the amyloid load Therefore the biological mechanismby which the genetic variation would alter Aβ build up andclearance merits further investigation
Author contributionsAndrea L Benedet study concept design analysis and in-terpretation of data composition of figures and drafting themanuscriptLei Yu analysis and interpretation of data and drafting themanuscript Aurelie Labbe supervision of statistical analysisand critical review of the manuscript for intellectual contentSulantha Mathotaarachchi Tharick A Pascoal Monica Shinand Min-Su Kang processing of image data and drafting themanuscript study concept design and drafting the manu-script Serge Gauthier Guy A Rouleau and Judes Poiriercritical review of the manuscript for intellectual content Da-vid A Bennett drafting the manuscript and critical review ofthe manuscript for intellectual content Pedro Rosa-Netostudy concept design study supervision and critical review ofthe manuscript for intellectual content
AcknowledgmentData collection and sharing for this project was funded by theAlzheimerrsquos Disease Neuroimaging Initiative (ADNI) (NIH
grant U01 AG024904) and the DOD ADNI (Department ofDefense award number W81XWH-12-2-0012) The ADNI isfunded by the National Institute on Aging the NationalInstitute of Biomedical Imaging and Bioengineering andthrough generous contributions from the following AbbViethe Alzheimerrsquos Association the Alzheimerrsquos Drug DiscoveryFoundation Araclon Biotech BioClinica Inc BiogenBristol-Myers Squibb CereSpir Inc Cogstate Eisai IncElan Pharmaceuticals Inc Eli Lilly and Company Euro-immun F Hoffmann-La Roche Ltd and its affiliated companyGenentech Inc Fujirebio GE Healthcare IXICO LtdJanssen Alzheimer Immunotherapy Research amp Develop-ment LLC Johnson amp Johnson Pharmaceutical Research ampDevelopment LLC Lumosity Lundbeck Merck amp Co IncMeso Scale Diagnostics LLC NeuroRx Research NeurotrackTechnologies Novartis Pharmaceuticals Corporation PfizerInc Piramal Imaging Servier Takeda Pharmaceutical Com-pany and Transition Therapeutics The Canadian Institutes ofHealth Research is providing funds to support ADNI clinicalsites in Canada Private sector contributions are facilitated bythe Foundation for the NIH (fnihorg) The granteeorganization is the Northern California Institute for Researchand Education and the study is coordinated by the AlzheimerrsquosTherapeutic Research Institute at the University of SouthernCalifornia ADNI data are disseminated by the Laboratory forNeuro Imaging at the University of Southern California
Study fundingThis work was also supported by the Canadian Institutes ofHealth Research (CIHR) (MOP-11-51-31) the AlzheimerrsquosAssociation (NIRG-08-92090) the Canadian Consortium onNeurodegeneration in Aging (CIHR-CCNA) and theWeston Brain Institute Andrea L Benedet is supported bythe CAPES FoundationmdashBrazil (032713-1) Pedro Rosa-Neto is supported by the Fonds de la recherche en sante duQuebec (chercheur boursier) Serge Gauthier and PedroRosa-Neto are members of the CIHR Canadian Consortiumon Neurodegeneration in Aging The Religious Orders Studyand the Rush Memory and Aging Project are funded bythe National Institute on Aging (P30AG10161 andR01AG17917) and the Illinois Department of Public Health
DisclosureAL Benedet has received research support from the CAPESFoundation L Yu has received research support from theNIH A Labbe and S Mathotaarachchi report no disclosuresTA Pascoal has received research support from thePREVENT-AD PhD scholarship M Shin and M-S Kangreport no disclosures S Gauthier has received honoraria forserving on the scientific advisory boards of Alzheon AxovantBoehringer Ingelheim Lilly Lundbeck Novartis SchwabeTauRx on the DSMB of a study sponsored by Eisai and forstudies run by the ADCS and by the ATRI has receivedspeaker honoraria from Schwabe serves on the editorial boardof Alzheimerrsquos amp Dementia has served as a consultant forKalgene and has received research support from the CIHRthe NIH Lundbeck Roche and Lilly GA Rouleau has
8 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
received research support from the CIHR the ALS Society ofCanada and the ALS Association J Poirier serves on theeditorial board of the Journal of Alzheimerrsquos Disease receivespublishing royalties from Trecarre and has received researchsupport from the JL Levesque Foundation DA Bennettserves on the scientific advisory boards of Vigorous MindsTakeda Pharmaceuticals AbbVie EABs (Indian Institute ofScience) DMCs (HRS and MIDUS) and the National Ad-visory Council on Aging is a consultant for Takeda Phar-maceuticals and AbbVie has received research support fromthe NIH and serves on the editorial boards of NeurologyCurrent Alzheimer Research and Neuroepidemiology P Rosa-Neto serves on the scientific advisory boards of EnigmaRadiopharmaceuticals and Kalgene serves on the editorialboard of Frontiers in Aging Neurosciences and has receivedresearch support fromEnigma Radiopharmaceuticals Eli Lillythe CIHR the FRQS Merck (Sharp amp Dohme CorporationMcGill Faculty of Medicine Grant for Translational Re-search) the Weston Brain Institute the Alzheimerrsquos Associa-tion Brain Canada and CQDM Funding information anddisclosures are provided at the end of the article Full disclo-sure form information provided by the authors is availablewith the full text of this article at NeurologyorgNG
Received June 8 2017 Accepted in final form December 4 2017
References1 Miksys S Tyndale RF Cytochrome P450-mediated drug metabolism in the brain
J Psychiatry Neurosci 201338152ndash1632 Iliff JJ Jia J Nelson J Goyagi T Klaus J Alkayed NJ Epoxyeicosanoid signaling in
CNS function and disease Prostaglandins Other Lipid Mediat 20109168ndash843 Stingl JC Brockmoller J Viviani R Genetic variability of drug-metabolizing enzymes
the dual impact on psychiatric therapy and regulation of brain function Mol Psy-chiatry 201318273ndash287
4 McCann SJ Pond SM James KM Le Couteur DG The association between poly-morphisms in the cytochrome P-450 2D6 gene and Parkinsonrsquos disease a case-controlstudy and meta-analysis J Neurol Sci 199715350ndash53
5 Chace C Pang D Weng C et al Variants in CYP17 and CYP19 cytochrome P450genes are associated with onset of Alzheimerrsquos disease in women with down syn-drome JAlzheimer Dis 201228601ndash612
6 McFayden MC Melvin WT Murray GI Regional distribution of individual forms of cyto-chrome P450 mRNA in normal adult human brain Biochem Pharmacol 199855825ndash830
7 Booth Depaz IM Toselli F Wilce PA Gillam EM Differential expression of cyto-chrome P450 enzymes from the CYP2C subfamily in the human brain Drug MetabDispos 201543353ndash357
8 Van Ess PJ Pedersen WA Culmsee C Mattson MP Blouin RA Elevated hepatic anddepressed renal cytochrome P450 activity in the Tg2576 transgenic mouse model ofAlzheimerrsquos disease J Neurochem 200280571ndash578
9 Pappolla MA Omar RA Chyan YJ et al Induction of NADPH cytochrome P450reductase by the Alzheimer beta-protein amyloid as a ldquoforeign bodyrdquo J Neurochem200178121ndash128
10 Liu M Sun A Shin EJ et al Expression of microsomal epoxide hydrolase is elevated inAlzheimerrsquos hippocampus and induced by exogenous beta-amyloid and trimethyl-tinEur J Neurosci 2006232027ndash2034
11 Bennett DA Schneider JA Arvanitakis Z Wilson RS Overview and findings from thereligious orders study Curr Alzheimer Res 20129628ndash645
12 Bennett DA Schneider JA Buchman AS Barnes LL Boyle PA Wilson RS Overviewand findings from the rush Memory and Aging Project Curr Alzheimer Res 20129646ndash663
13 Bennett DA Schneider JA Wilson RS Bienias JL Arnold SE Neurofibrillary tanglesmediate the association of amyloid load with clinical Alzheimer disease and level ofcognitive function Arch Neurol 200461378ndash384
14 Saykin AJ Shen L Yao X et al Genetic studies of quantitative MCI and AD phe-notypes in ADNI Progress opportunities and plans Alzheimers Dement 201511792ndash814
15 Lim AS Srivastava GP Yu L et al 24-hour rhythms of DNA methylation and theirrelation with rhythms of RNA expression in the human dorsolateral prefrontal cortexPLoS Genet 201410e1004792
16 Rogers JF Nafziger AN Bertino JS Jr Pharmacogenetics affects dosing efficacy andtoxicity of cytochrome P450-metabolized drugs Am J Med 2002113746ndash750
17 Anzenbacher P Anzenbacherova E Cytochromes P450 and metabolism of xeno-biotics Cell Mol Life Sci 200158737ndash747
18 Meyer RP Gehlhaus M Knoth R Volk B Expression and function of cytochromep450 in brain drug metabolism Curr Drug Metab 20078297ndash306
19 Purcell S Neale B Todd-Brown K et al PLINK a tool set for whole-genomeassociation and population-based linkage analyses Am J Hum Genet 200781559ndash575
20 R A Language and Environment for Statistical Computing [computer program]Version 310 Vienna R Foundation for Statistical Computing 2015
21 Package RMINC Voxel-Wise Morphometry Using RMINC [computer program]Montreal McGill University 2010
22 Ramanan VK Risacher SL Nho K et al APOE and BCHE as modulators of cerebralamyloid deposition a florbetapir PET genome-wide association study Mol Psychiatry201419351ndash357
23 Braak H Braak E Neuropathological stageing of Alzheimer-related changes ActaNeuropathol 19918220
24 Tapiola T Alafuzoff I Herukka S et al Cerebrospinal fluid β-amyloid 42 and tauproteins as biomarkers of Alzheimer-type pathologic changes in the brain ArchNeurol 200966382ndash389
25 Sperling R Mormino E Johnson K The evolution of preclinical Alzheimerrsquos diseaseimplications for prevention trials Neuron 201484608ndash622
26 Wang D Jiang Z Shen Z et al Functional evaluation of genetic and environmentalregulators of p450 mRNA levels PLoS One 20116e24900
27 Sim SC Risinger C Dahl ML et al A common novel CYP2C19 gene variant causesultrarapid drug metabolism relevant for the drug response to proton pump inhibitorsand antidepressants Clin Pharmacol Ther 200679103ndash113
28 Ishii G Suzuki A Oshino S Shiraishi H Otani K CYP2C19 polymorphism affectspersonality traits of Japanese females Neurosci Lett 200741177ndash80
29 Ercan B Ayaz L Cicek D Tamer L Role of CYP2C9 and CYP2C19 polymorphismsin patients with atherosclerosis Cell Biochem Funct 200826309ndash313
30 Westphal C Konkel A SchunckWH CYP-eicosanoidsmdasha new link between omega-3 fatty acids and cardiac disease Prostaglandins Other Lipid Mediat 20119699ndash108
31 Cheng ZN Shu Y Liu ZQ Wang LS Ou-Yang DS Zhou HH Role of cytochromeP450 in estradiol metabolism in vitro Acta Pharmacol Sin 200122148ndash154
32 Giraud SN Caron CM Pham-Dinh D Kitabgi P Nicot AB Estradiol inhibits ongoingautoimmune neuroinflammation and NFkappaB-dependent CCL2 expression in re-active astrocytes Proc Natl Acad Sci USA 20101078416ndash8421
33 Xu H Wang R Zhang YW Zhang X Estrogen beta-amyloid metabolismtraffickingand Alzheimerrsquos disease Ann NY Acad Sci 20061089324ndash342
34 Amtul Z Uhrig M Wang L Rozmahel RF Beyreuther K Detrimental effects ofarachidonic acid and its metabolites in cellular and mouse models of Alzheimerrsquosdisease structural insight Neurobiol Aging 201233831e21ndashe31
35 Sarkar P Narayanan J Harder DR Differential effect of amyloid beta on the cyto-chrome P450 epoxygenase activity in rat brain Neuroscience 2011194241ndash249
36 Kisler K Nelson AR Montagne A Zlokovic BV Cerebral blood flow regulation andneurovascular dysfunction in Alzheimer disease Nat Rev Neurosci 201718419ndash434
37 Bibi Z Role of cytochrome P450 in drug interactions Nutr Metab (Lond) 200852738 Ownby RL Crocco E Acevedo A John V Loewenstein D Depression and risk for
Alzheimer disease systematic review meta-analysis and metaregression analysisArch Gen Psychiatry 200663530ndash538
39 Sheline YI West T Yarasheski K et al An antidepressant decreases CSF Abetaproduction in healthy individuals and in transgenic AD mice Sci Transl Med 20146236re234
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 9
DOI 101212NXG000000000000021620184 Neurol Genet
Andreacutea L Benedet Lei Yu Aureacutelie Labbe et al variant mitigates Alzheimer disease pathophysiology in vivo and postmortemCYP2C19
This information is current as of January 30 2018
ServicesUpdated Information amp
httpngneurologyorgcontent41e216fullhtmlincluding high resolution figures can be found at
References httpngneurologyorgcontent41e216fullhtmlref-list-1
This article cites 37 articles 2 of which you can access for free at
Subspecialty Collections
httpngneurologyorgcgicollectionpetPET
httpngneurologyorgcgicollectionassociation_studies_in_geneticsAssociation studies in genetics
httpngneurologyorgcgicollectionalzheimers_diseaseAlzheimers diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpngneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpngneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
reserved Online ISSN 2376-7839Published by Wolters Kluwer Health Inc on behalf of the American Academy of Neurology All rightsan open-access online-only continuous publication journal Copyright Copyright copy 2018 The Author(s)
is an official journal of the American Academy of Neurology Published since April 2015 it isNeurol Genet

In sporadic Alzheimer disease (AD) accumulation of amy-loid-β (Aβ) seems to be associated with reduced Aβ clearancebut little is known about the biochemical pathways underlyingcerebral Aβ metabolism
Cytochrome P450 (CYP) is a family of enzymes known tometabolize several endogenous and exogenous substratesBesides metabolizing drugs brain CYPs are also involved inthe modulation of blood flow metabolism of fatty acidscholesterol and neurotransmitters and mobilization of in-tracellular calcium1ndash3 all pathways that have been somehowlinked to AD andor amyloid metabolism In addition studieshave shown that CYP proteins and their genetic variants areassociated with the brain immune response2 and neurode-generative diseases245
Although previous evidence shows CYP expression in areasaffected by AD such as the amygdala frontal and temporalcortices and the hippocampus67 there are only a few reportsdocumenting a link between CYPs and AD pathophysiologyIn one study the overexpression of human amyloid pre-cursor protein (APP) on the Tg2576 mice model was as-sociated with elevated CYP hepatic function while theirrenal counterparts were depressed8 In addition anotherstudy presented evidence that Aβ stimulates NADPH-cytochrome P450 reductase (POR) a CYP inducer in APPtransgenic mice and human AD brains which may affect theredox status9
In line with the evidence that Aβ can stimulate metabolic cas-cades in a similar fashion as foreign compounds910 andknowing that brain CYP could potentially affect amyloid me-tabolism also through other mechanisms we tested the hy-pothesis that variants in CYP genes could be associated with Aβand consequently have an impact in cognitive performanceWe examined the association of CYP variants with biomarkersof Aβ load in vivo and postmortem as further specified
MethodsStudy participantsDiscovery analysis data were obtained from the AD Neu-roimaging Initiative (ADNI) database (adniloniuscedu)For this study cognitively normal (CN) individuals hada Mini-Mental State examination (MMSE) score of 24 orhigher and a clinical dementia rating (CDR) of 0 The
operational criteria adopted for mild cognitive impairment(MCI) were participants who had an MMSE score equal toor greater than 24 a CDR of 05 subjective memory con-cern objective memory loss measured by education-adjusted scores on delayed recall of 1 paragraph fromWechsler Memory Scale Logical Memory II (ge16 years le88ndash15 years le4 and 0ndash7 years le2) and essentially normalactivities of daily living Participants with AD were individ-uals with an MMSE score lower or equal to 26 CDR higherthan 05 who met the National Institute of Neurological andCommunicative Disorders and StrokendashAlzheimerrsquos Diseaseand Related Disorders Association criteria for probable ADAll individuals had absence of any other neuropsychiatricdisorders The ADNI inclusionexclusion criteria are de-scribed in detail at adni-infoorg
Our report also used data from the Religious Orders Study(ROS) and Rush Memory and Aging Project (MAP) 2 longi-tudinal clinical pathologic cohort studies of aging anddementia1112 All participants enrolled without known dementiaand agreed to annual clinical and neuropsychological assess-ments and brain donations after death The clinical diagnosis ofdementia and AD followed the National Institute of Neuro-logical and Communicative Disorders and StrokendashAlzheimerrsquosDisease and Related Disorders Association recommendationsThe MCI diagnosis is given to participants who were rated asimpaired based on MMSE and on the Consortium toEstablish a Registry for Alzheimerrsquos Disease (CERAD) neuro-psychological measures but who were not found to have de-mentia by the examining neurologist Our study includes onlyMCI participants with 1 impaired domain and no other cause ofcognitive impairment Notably the studies are conducted by thesame team of investigators and share a large common core oftesting batteries which allow for combined analyses
The discovery sample consisted of a subset of ADNI partic-ipants who had [18F]florbetapir PET genetic data and CSFdata available The findings obtained with the discoverysample were then generalized to the Rush-ROSMAP par-ticipants who died at age 90 years or younger
Standard protocol approvals registrationsand patient consentsThis secondary analysis study followed all institutional reviewboard regulations which are detailed in the supplementarymethods
GlossaryAA = arachidonic acid Aβ = amyloid-β AD = Alzheimer disease ADNI = AD Neuroimaging Initiative APP = amyloidprecursor protein CDR = clinical dementia rating CN = cognitively normal CYP = cytochrome P450 EET =epoxyeicosatrienoic acid LD = linkage disequilibriumMA = minor alleleMAF = minor allele frequencyMAP =Memory andAging ProjectMCI = mild cognitive impairmentMMSE = Mini-Mental State Examination PHF = paired helical filament p-tau = phosphorylated tau ROS = Religious Orders Study SNP = single nucleotide polymorphism SUVR = standardizeduptake value ratio
2 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
PhenotypesFrom the ADNI we obtained demographics neurocognitivescores CSF and plasmatic data available after passing rigor-ous quality control The preprocessed imaging and geneticdata underwent additional processing and quality controlbefore the statistical analysis in our study as described belowFrom Rush all the data used in this project to perform sta-tistical analyses were already processed and quality controlledwith very rigorous criteria
In the ADNI cohort the brain Aβ load was estimated usingthe [18F]florbetapir PET standardized uptake value ratio(SUVR details are provided in figure 1 and e-methods)Separately Aβ and hyperphosphorylated tau (p-tau) lev-els were measured in the CSF and plasma These data wereobtained by the Biomarkers Consortium CSF and thedescription of the methodology regarding the sampleacquisition processing and analysis is available atthe ADNI website (adniloniuscedudata-samplesbio-specimen-data)
In the Rush-ROSMAP cohort postmortem data of the Aβload and paired helical filament (PHF) tau tangle density wereobtained from 8 brain regions using immunohistochemistry13
Cognitive function was evaluated in both the ADNI and Rush-ROSMAP For the ADNI the global cognition scores weredeveloped by making a z-score of the sum of the memory andthe executive function composite scores In the Rush-ROSMAP the global cognitive scores were based on 17 cognitiveperformance tests and we included only participants whounderwent autopsies thus the last valid cognitive scoreproximate to death was used for this analysis (for furtherdetails see e-Methods httplinkslwwcomNXGA20)
We used gene expression data to examine cis expression quan-titative trait loci (eQTL) For the ADNI total RNAwas obtainedfrom the peripheral bloodmdashat the same visit when the imagingdata were obtainedmdashand after quality control was hybridized tothe Affymetrix Human Genome U219 array plate as described14
The normalized and quality-controlled expression data14 wereused For the Rush-ROSMAP RNA was extracted from thegray matter of dorsolateral prefrontal cortex blocks usingthe miRNeasy Mini Kit (Qiagen Venlo Netherlands) and theRNase-free DNase Set (Qiagen) These samples were quantifiedand qualified as described15 and then sequenced on the IlluminaHiSeq with 101 bp paired-end reads The normalized andquality-controlled expression data were used In both analysesthe RNA integrity measurement was used as a covariate
Figure 1 [18F]florbetapir standardized uptake value ratio analytical method
Flowchart showing acquisition methods (purple) imageprocessing (blue) and outcomes (green) ADNI = Alz-heimerrsquos Disease Neuroimaging Initiative GM = graymatter ROI = region of interest SUVR = standardizeduptake value ratio WM = white matter
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 3
Genotyping imputation and gene selectionWe interrogated single nucleotide polymorphisms (SNPs)from the CYP genes of the 5mdashprobablymdashmost importantmetabolizers CYP3A4 CYP2D6 CYP2C9 CYP1A2 andCYP2C19316ndash18 Methods involved in DNA processing aredetailed in the supplementary methods section (httplinkslwwcomNXGA20) Using PLINK19 the SNPs within thesegenes were recoded based on the dominant model with re-spect to the MAs and the ones in high linkage disequilibrium(LD) (r2 gt 08) were removed from the analysis to avoidunnecessary testing A total of 28 SNPs were used in the initialstep of the association analysis
For Rush-ROSMAP cohorts DNA was extracted genotypedimputed and quality controlled according to the proceduresdescribed in e-Methods (httplinkslwwcomNXGA20)
Statistical analysisThe statistical tests were performed in R statistical software20
where linear regressionmodels were used to test the associationbetween the genotypes and the phenotypes in both cohortsseparately All statistical models included the covariates of agesex and APOE-e4 carriage status except for the models inwhich global cognition was the outcome measure where theAPOE-e4 status was replaced by years of education The initialanalysismdashusing the global [18F]florbetapir SUVR asa phenotypemdashwas performed to interrogate a total of 28 SNPs(table e-1 httplinkslwwcomNXGA19) To correct formultiple testing the statistical significance of the SNP discoverywas set at p lt 00017 using a Bonferroni correction for 28 testsand a type I error α = 005 The significant SNPs were furthertested for associations with other phenotypes in ADNI andRush-ROSMAP cohorts To test for group differences in thediagnostic status we added diagnosis as a covariate in the re-gression models and also tested the interaction between thediagnostic status and desired independent variable Effect sizecalculations (Cohen d) were also used
A voxel-based analysis was performed using the RMINCimaging tool21 where parametric images were obtained con-trasting the [18F]florbetapir SUVR between the genotypegroups of the SNP found in the discovery analysis Afterrandom field theory correction for multiple comparisons the tvalue threshold of significance was leminus31 (p le 0001)
Mediation analysis was used to examine whether the SNPassociations with cognition were mediated through the effect ofAβ To test this hypothesis first the association of CYP SNPswith global cognitive performance was evaluated using linearregression models Then if the association was significant themeasure of Aβ was added to this model to check whether theSNP associations with cognitionwere attenuated because of Aβ
ResultsDemographics and general information of the ADNI andRush-ROSMAP cohorts are presented in table 1 After
imaging and genetic quality control a total of 338 ADNIparticipants were studied including 186 CN 105 single- ormultiple-domain amnestic MCI and 47 AD A total of 738Rush-ROSMAP participants were analyzed including 301CN 179 amnestic MCI and 258 AD As expected in bothcohorts CN individuals had higher MMSE scores and fewerwere APOE-e4 carriers as compared to both MCI and ADThere was no difference in sex or years of formal educationbetween diagnostic groups in both cohorts (details are pro-vided in table e-2 httplinkslwwcomNXGA19)
CYP2C19 polymorphism is associated with theglobal [18F]florbetapir SUVR in the ADNI cohortThe discovery analysis tested the association of 28 SNPs fromCYP genes with the global [18F]florbetapir SUVR The anal-ysis unveiled an association of an SNP (rs4388808) in theCYP2C19 gene (t = minus343 p = 00006) (figure 2A) Theminor allele frequency (MAF) of rs4388808G in the ADNIcohort was 019 and the minor allele (MA) carriers displayedless [18F]florbetapir binding as compared to the noncarriersIt is important that the association remained significant whenadjusting the model for diagnosis In addition no interactionwas found between the SNP and diagnostic status The esti-mated effect size of rs4388808 in the whole sample wasCohen d = 036 Of interest the effect size was greater for thesubset of APOE-e4 carriers (Cohen d = 051) as compared toAPOE-e4 noncarriers (Cohen d = 033)
MA carriers of the CYP2C19 polymorphismhaveless Aβ in AD-related regions in theADNI cohortTo identify the brain areas responsible for the global differ-ence observed in [18F]florbetapir uptake revealed in the dis-covery analysis we performed a voxel-wise analysis Theanalysis showed that carriers of rs4388808G had less [18F]florbetapir binding in the frontal posterior cingulate andinferior temporal cortices (figure 3) with the voxels in theseregions presenting p values equal to or less than 0001
Findings with brain imaging are corroboratedby CSF data in the ADNI cohortBecause of the well-known relationship between brain and CSFlevels of Aβ we tested the association between rs4388808 andCSF Aβ Because not all participants in the ADNI sample hadCSF data we performed the analysis in a subset of 260 indi-viduals As expected MA carriers presented more CSF Aβ thannoncarriers (t = 294 p = 0003) (figure 2B) The CYP2C19polymorphismwas associatedwith the Aβp-tau ratio (t = 229p = 002) and higher ratios were found in carriers ofrs4388808G (figure 2C) No association was found using p-tauas a single outcome measure (t = minus124 p = 021)
Association between the CYP2C19polymorphism and Aβ is exclusive to the Aβbrain levels in the ADNI cohortTo examine whether the CYP2C19 polymorphism is also as-sociated with Aβ levels outside the brain we tested the
4 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
association of rs4388808 with plasmatic levels of Aβ ina subsample of 113 participants We did not find an associa-tion between the SNP and free (t = minus060 p = 054) or total(t = 049 p = 062) plasmatic Aβ1-42 levels (figure 2D) in-dicating that the findings reported above are specific tothe CNS
Results from theADNI cohortwere generalizedto the Rush-ROSMAP cohortWe next generalized the findings to postmortem indices of theAβ load and to neurofibrillary tangles in Rush-ROSMAPcohorts From the 8 brain regions where the Aβ load wasmeasured we selected 6 regions (angular gyrus anterior cin-gulate entorhinal cortex and mesial and inferior temporalcortices) and calculated an average per subject These 6 regionswere chosen to match with the brain areas where we detecteddifferences between genotype groups in the voxel-wise analysisWhen testing the association between the average of the Aβload and the CYP2C19 polymorphism we found thatrs4388808G (MAF = 015) carriers had a lower Aβ load thannoncarriers (t = minus215 p = 003) (figure 2E) consistent withfindings from the ADNI cohort The difference betweenrs4388808G carriers and noncarriers in PHF tau tangle densitydid not reach statistical significance (t = minus165 p = 009)
MA carriers of the CYP2C19 polymorphism hadbetter cognitive performance thannoncarriersin both cohortsTo examine whether the CYP2C19 polymorphism affects thecognitive performance we used composite measures of globalcognition as the outcome in linear regression models In theADNI cohort there was a clear tendency to an associationwith the CYP2C19 polymorphism (t = 192 p = 005) (figure2F) while in Rush-ROSMAP cohorts the association wassuch that rs4388808G carriers had a higher cognition thannoncarriers (t = 208 p = 003) (figure 2G)
The effect of the CYP2C19 polymorphism oncognition is mediated by AβA mediation analysis was used to determine whether theCYP2C19 polymorphism was directly or indirectly associatedwith cognitive performance We retested the association be-tween the CYP2C19 polymorphism and cognition but nowadding the measures of [18F]florbetapir or Aβ load in themodel Neither of the previous genotype-phenotype associa-tions remained significant indicating that the observed effectof the genotype was mediated by Aβ
Association between rs4388808 and geneexpression of CYP2C19 suggests that the SNPis functionalTo check whether the polymorphism has an effect in the geneexpression we tested the association between rs4388808 andCYP2C19 RNA levels In the ADNI cohort with 304 partic-ipants for this analysis we found a cis eQTL such that MAcarriers displayed higher RNA levels in the blood (t = 238 p =001) (figure 2H) In Rush-ROSMAP cohorts very low levelsof CYP2C19 expression were detected in postmortem brainsand the data did not provide consistent results (data notshown) as found in the blood expression data from the ADNI
DiscussionIn the present study we found an association between theCYP2C19 polymorphism and Aβ burden across the spectrumof AD in which carriers of rs4388808G presented a less Aβload and downstream cognitive impairment This associationwas initially detected using the [18F]florbetapir SUVR andposteriorly confirmed with CSF Aβ in the ADNI cohort Noassociation was found between the polymorphism and di-agnostic groups suggesting that the SNP is associated with anamyloid load rather than with the disease status Effect size
Table 1 Demographic and key characteristics of the samples
ADNI Rush-ROSMAP
CN MCI AD CN MCI AD
No of participants 186 105 47 301 179 258
Males n () 91 (489) 64 (61) 26 (553) 182 (605) 101 (564) 154 (597)
Age y mean (SD) 7570 (663) 7429 (812) 7567 (737) 837 (519)a3 8526 (435) 8595 (381)
APOE-laquo4 () 52 (28)a1 57 (543)a1 30 (638)a1 49 (164)a4 50 (279)a4 112 (434)a4
MMSE mean (SD) 2909 (114)a2 2724 (195)a2 2034 (366)a2 2869 (142)a5 2791 (183)a5 2549 (517)a5
Education y 1641 (275) 1667 (268) 1613 (247) 1653 (374) 1634 (347) 1666 (394)
MA n ()b 67 (19) 36 (10) 17 (5) 87 (11) 52 (7) 63 (85)
Abbreviations AD = Alzheimer disease ADNI = AD Neuroimaging Initiative CN = cognitively normal MA = minor allele MAP = Memory and Aging ProjectMCI = mild cognitive impairment MMSE = Mini-Mental State Examination ROS = Religious Orders Studya Statistically different from the other groups of the same sample 1p = 23 times 10minus7 2p = 2 times 10minus16 3p = 92 times 10minus8 4p = 17 times 10minus11 and 5p = 2 times 10minus16b MA (rs4388808G) percentage relative to its major sample (ADNI or Rush-ROSMAP)
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 5
analysis suggests that the benefic effects of the polymorphismare larger in APOE-e4 carriers as compared to noncarriers orthe whole sample in which the effect is similar to what hasbeen described for a BCHE gene polymorphism22 Sub-sequently this association was generalized to postmortemdata from the Rush-ROSMAP cohort Finally both cohortsshowed a protective association between the MA of CYP2C19and cognition It is important that subsequent mediationanalysis suggested that the effects of CYP2C19 on cognitionwere mediated by Aβ
Overall these results support the notion that the MA ofCYP2C19 (rs4388808) is a protective variant against Aβ anddownstream cognitive impairment Of interest voxel-wiseanalysis in in vivo participants revealed that the MA was as-sociated with reduced Aβ burden in AD-related regions in thefrontal inferior temporal and posterior cingulate cortices23
In addition the CYP2C19 SNP was associated with the CSFAβp-tau ratio but not with plasmatic Aβ Beyond the fact thatthe Aβp-tau ratio is postulated to better represent neuriticplaques than the single CSF Aβ information24 these results
Figure 2 Comparison between noncarriers (minus) and carriers (+) of the minor allele of rs4388808 (CYP2C19)
In the ADNI cohort a difference was observedin the brain Aβ load (A) CSF Aβ levels (B) andCSF Aβp-tau ratio (C) but no difference wasdetected in plasmatic levels of Aβ (D) Resultswere generalized to postmortem data of Rush-ROSMAP cohorts where a concordant patternwas observed in the Aβ load (percentage areaoccupied by amyloid) (E) A tendency towardsignificance in global cognitive performance (F)was found in the ADNI cohort while in theRush-ROSMAP it was significant (G) CYP2C19expression levels were also different betweennoncarriers (minus) and carriers (+) of the minorallele of rs4388808 (H) in the ADNI cohort Alllinear models were adjusted for age sex andAPOE-e4 carriage status with the exception ofthe models using global cognition and geneexpression which were adjusted for age sexand years of education or RNA integrity re-spectively Aβ = amyloid-β ADNI = AlzheimerrsquosDisease Neuroimaging Initiative MAP = Mem-ory and Aging Project p-tau = phosphorylatedtau ROS = Religious Orders Study SUVR =standardized uptake value ratio
6 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
provide further evidence that the polymorphism affects thebrain accumulation of neuritic plaques in a tissue- and disease-specific manner24 In both cohorts the referred poly-morphism was not associated with PHF tau or single CSFp-tau agreeing with the framework that Aβ and tau haveindependent upstream triggers25 In line with overall signifi-cant results the CYP2C19 polymorphism also presenteda protective effect in global cognition being supported byprevious studies that linked Aβ pathology to cognitivechanges13 Indeed the mediation analysis supported that theassociation between the genetic factor and cognitive abnor-malities is likely due to the upstream accumulation of Aβ
Levels of the CYP2C19 protein are associated with thers4388808 polymorphism26 which is an intronic variant of theCYP2C19 gene Indeed we found a similar association showingthat MA carriers expressed higher levels of CYP2C19 whencompared with noncarriers suggesting that the SNP affects theexpression of its gene To corroborate this finding we retrievedthe SNPs that were previously excluded because of high LD andverified that none was associated with CYP2C19 expression inthe blood (data not shown) reinforcing the idea that rs4388808is functional This association could not be detected in post-mortem data probably because of the low CYP expression orbecause of its limited detection by the method used7
Despite the SNP not being widely studied to provide in-formation about the association with other phenotypes other
polymorphisms in the gene have been correlated with meta-bolic variability atherosclerosis and behavioral traits27ndash29
CYP2C19 expression has been detected in several brainregions67 however its function in the CNS has not been fullyelucidated Some reports have suggested that besides its roleat metabolizing exogenous substrates CYP2C19 also partic-ipates in other important biological cascades such as themetabolism of serotonin and sexual hormones1 and the me-tabolism of the arachidonic acid (AA) where it functions as anepoxygenase30
The biochemical mechanism underlying the association ofCYP2C19 with Aβ remains speculative A study has shownthat Aβ is able to act as a foreign body and trigger PORactivating CYPs to initiate catabolic reactions9 In turnCYP2C19 is involved in the catalysis of estradiol31 Becauseseveral studies have reported the beneficial effects of estrogenson ADmdashdemonstrating that these steroids are able to act asanti-inflammatory compounds32 and even inhibit Aβproduction33mdashone could think that MA carriers of rs4388808have a reduced expression of this CYP in the brain or that theyhave an increased structural incompatibility betweenCYP2C19 and estradiol Consequently MA carriers couldpresent a reduction in the catabolism of estrogen and a re-duction in the Aβ load Alternatively the Aβ load would beaffected by the levels of epoxyeicosatrienoic acids (EETs)produced via the metabolism of AA by CYP epoxygenases Inthe brain studies have shown that EETs have beneficial effectsat regulating blood flow cortical angiogenesis and at pro-moting anti-inflammatory reactions (for review see reference2) By contrast when AA is metabolized via other cascadesthe protective effect of EETs is lost and there is a possibilityof induction of Aβ production andor accumulation34 Inaddition it was demonstrated that Aβ is able to reduceepoxygenase activity and consequently decrease EET pro-duction35 probably leading to more susceptibility to neuronaldamage (for review see reference 36) In this scenario itwould be possible that the MA of rs4388808 increases theexpression of CYP2C19 leading to an ldquoAβ-resistantrdquo epox-ygenase metabolism of AA with a maintained production ofEETs and a more protective phenotype against amyloidosis
It is also plausible to think however that the associationdescribed here between theCYP2C19 polymorphism and Aβis indirect being mediated by the effect of a drug CYP2C19is known to metabolize several drugs37 including medi-cations to treat depression which has been mentioned asa risk factor for AD (for review see reference 38) Cit-alopram for example is an antidepressant metabolized byCYP2C19 that has been shown to decrease Aβ production39
Thus one may think that the polymorphism rs4388808 isthen associated with an improved action of citalopramleading to the decreased brain Aβ burden rather than as-sociated with Aβ metabolism itself Because it is difficult totrack all the medications the study participants have takenbefore been enrolled it is unfeasible at the moment to checkwhether this association holds true
Figure 3 T-statistical parametric maps showing differencesbetween noncarriers (minus) and carriers (+) of theminor allele of rs4388808 (CYP2C19)
T-statistical parametric maps superimposed on average structural MRI showbrain regions with lower standardized uptake value ratio (SUVR) values inminor allele carriers (CYP2C19 (+)) of the polymorphism of CYP2C19 Statisticaldifferences overlap with brain regions vulnerable to Alzheimer disease path-ophysiology such as the posterior cingulate frontal and temporal cortices
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 7
The interpretation of the results should take into consider-ation a few limitations The use of the 2 cohorts does not allowus to detect and measure Aβ pathology with the samemethodology inserting in the analysis some variability specificto the method used However despite being measured withdifferent techniques both the imaging radiotracer and theantibody used in the brain tissue are expected to bind to thefibrillar form of Aβ13 Similarly the cognitive composite scoresfrom the 2 cohorts have some differences that should beconsidered There are also some intrinsic characteristics of thesamples that have to be taken into account (1) the inclusioncriteria of each study (2) the average age difference betweenthe studies and (3) and the disease stage in which Aβmeasures were performedmdashat the end or during the diseaseprocess Sample size restrictions impose some degree ofcaution when interpreting findings including for examplethe lack of association between the genotype and plasmaticlevels of Aβ or cognition in the ADNI cohort In additionboth cohorts are mostly composed of non-Latino Caucasianslimiting the extrapolation of the present findings to otherpopulation groups Longitudinal analysis together with func-tional genomics and biochemistry experiments not per-formed here would also be necessary to determine the effectof the SNP in the protein function as well as to support anytheoretical framework proposed in the Discussion section
Results obtained from the 2 independent cohorts providecompelling evidences linking the CYP2C19 polymorphismand Aβ pathology suggesting that the MA of rs4388808confers protective effects against Aβ accumulation in the brainand its downstream cognitive consequences These resultscould have implication for antiamyloid clinical trial designs aspreclinical rs4388808G would present a protective factoragainst the amyloid load Therefore the biological mechanismby which the genetic variation would alter Aβ build up andclearance merits further investigation
Author contributionsAndrea L Benedet study concept design analysis and in-terpretation of data composition of figures and drafting themanuscriptLei Yu analysis and interpretation of data and drafting themanuscript Aurelie Labbe supervision of statistical analysisand critical review of the manuscript for intellectual contentSulantha Mathotaarachchi Tharick A Pascoal Monica Shinand Min-Su Kang processing of image data and drafting themanuscript study concept design and drafting the manu-script Serge Gauthier Guy A Rouleau and Judes Poiriercritical review of the manuscript for intellectual content Da-vid A Bennett drafting the manuscript and critical review ofthe manuscript for intellectual content Pedro Rosa-Netostudy concept design study supervision and critical review ofthe manuscript for intellectual content
AcknowledgmentData collection and sharing for this project was funded by theAlzheimerrsquos Disease Neuroimaging Initiative (ADNI) (NIH
grant U01 AG024904) and the DOD ADNI (Department ofDefense award number W81XWH-12-2-0012) The ADNI isfunded by the National Institute on Aging the NationalInstitute of Biomedical Imaging and Bioengineering andthrough generous contributions from the following AbbViethe Alzheimerrsquos Association the Alzheimerrsquos Drug DiscoveryFoundation Araclon Biotech BioClinica Inc BiogenBristol-Myers Squibb CereSpir Inc Cogstate Eisai IncElan Pharmaceuticals Inc Eli Lilly and Company Euro-immun F Hoffmann-La Roche Ltd and its affiliated companyGenentech Inc Fujirebio GE Healthcare IXICO LtdJanssen Alzheimer Immunotherapy Research amp Develop-ment LLC Johnson amp Johnson Pharmaceutical Research ampDevelopment LLC Lumosity Lundbeck Merck amp Co IncMeso Scale Diagnostics LLC NeuroRx Research NeurotrackTechnologies Novartis Pharmaceuticals Corporation PfizerInc Piramal Imaging Servier Takeda Pharmaceutical Com-pany and Transition Therapeutics The Canadian Institutes ofHealth Research is providing funds to support ADNI clinicalsites in Canada Private sector contributions are facilitated bythe Foundation for the NIH (fnihorg) The granteeorganization is the Northern California Institute for Researchand Education and the study is coordinated by the AlzheimerrsquosTherapeutic Research Institute at the University of SouthernCalifornia ADNI data are disseminated by the Laboratory forNeuro Imaging at the University of Southern California
Study fundingThis work was also supported by the Canadian Institutes ofHealth Research (CIHR) (MOP-11-51-31) the AlzheimerrsquosAssociation (NIRG-08-92090) the Canadian Consortium onNeurodegeneration in Aging (CIHR-CCNA) and theWeston Brain Institute Andrea L Benedet is supported bythe CAPES FoundationmdashBrazil (032713-1) Pedro Rosa-Neto is supported by the Fonds de la recherche en sante duQuebec (chercheur boursier) Serge Gauthier and PedroRosa-Neto are members of the CIHR Canadian Consortiumon Neurodegeneration in Aging The Religious Orders Studyand the Rush Memory and Aging Project are funded bythe National Institute on Aging (P30AG10161 andR01AG17917) and the Illinois Department of Public Health
DisclosureAL Benedet has received research support from the CAPESFoundation L Yu has received research support from theNIH A Labbe and S Mathotaarachchi report no disclosuresTA Pascoal has received research support from thePREVENT-AD PhD scholarship M Shin and M-S Kangreport no disclosures S Gauthier has received honoraria forserving on the scientific advisory boards of Alzheon AxovantBoehringer Ingelheim Lilly Lundbeck Novartis SchwabeTauRx on the DSMB of a study sponsored by Eisai and forstudies run by the ADCS and by the ATRI has receivedspeaker honoraria from Schwabe serves on the editorial boardof Alzheimerrsquos amp Dementia has served as a consultant forKalgene and has received research support from the CIHRthe NIH Lundbeck Roche and Lilly GA Rouleau has
8 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
received research support from the CIHR the ALS Society ofCanada and the ALS Association J Poirier serves on theeditorial board of the Journal of Alzheimerrsquos Disease receivespublishing royalties from Trecarre and has received researchsupport from the JL Levesque Foundation DA Bennettserves on the scientific advisory boards of Vigorous MindsTakeda Pharmaceuticals AbbVie EABs (Indian Institute ofScience) DMCs (HRS and MIDUS) and the National Ad-visory Council on Aging is a consultant for Takeda Phar-maceuticals and AbbVie has received research support fromthe NIH and serves on the editorial boards of NeurologyCurrent Alzheimer Research and Neuroepidemiology P Rosa-Neto serves on the scientific advisory boards of EnigmaRadiopharmaceuticals and Kalgene serves on the editorialboard of Frontiers in Aging Neurosciences and has receivedresearch support fromEnigma Radiopharmaceuticals Eli Lillythe CIHR the FRQS Merck (Sharp amp Dohme CorporationMcGill Faculty of Medicine Grant for Translational Re-search) the Weston Brain Institute the Alzheimerrsquos Associa-tion Brain Canada and CQDM Funding information anddisclosures are provided at the end of the article Full disclo-sure form information provided by the authors is availablewith the full text of this article at NeurologyorgNG
Received June 8 2017 Accepted in final form December 4 2017
References1 Miksys S Tyndale RF Cytochrome P450-mediated drug metabolism in the brain
J Psychiatry Neurosci 201338152ndash1632 Iliff JJ Jia J Nelson J Goyagi T Klaus J Alkayed NJ Epoxyeicosanoid signaling in
CNS function and disease Prostaglandins Other Lipid Mediat 20109168ndash843 Stingl JC Brockmoller J Viviani R Genetic variability of drug-metabolizing enzymes
the dual impact on psychiatric therapy and regulation of brain function Mol Psy-chiatry 201318273ndash287
4 McCann SJ Pond SM James KM Le Couteur DG The association between poly-morphisms in the cytochrome P-450 2D6 gene and Parkinsonrsquos disease a case-controlstudy and meta-analysis J Neurol Sci 199715350ndash53
5 Chace C Pang D Weng C et al Variants in CYP17 and CYP19 cytochrome P450genes are associated with onset of Alzheimerrsquos disease in women with down syn-drome JAlzheimer Dis 201228601ndash612
6 McFayden MC Melvin WT Murray GI Regional distribution of individual forms of cyto-chrome P450 mRNA in normal adult human brain Biochem Pharmacol 199855825ndash830
7 Booth Depaz IM Toselli F Wilce PA Gillam EM Differential expression of cyto-chrome P450 enzymes from the CYP2C subfamily in the human brain Drug MetabDispos 201543353ndash357
8 Van Ess PJ Pedersen WA Culmsee C Mattson MP Blouin RA Elevated hepatic anddepressed renal cytochrome P450 activity in the Tg2576 transgenic mouse model ofAlzheimerrsquos disease J Neurochem 200280571ndash578
9 Pappolla MA Omar RA Chyan YJ et al Induction of NADPH cytochrome P450reductase by the Alzheimer beta-protein amyloid as a ldquoforeign bodyrdquo J Neurochem200178121ndash128
10 Liu M Sun A Shin EJ et al Expression of microsomal epoxide hydrolase is elevated inAlzheimerrsquos hippocampus and induced by exogenous beta-amyloid and trimethyl-tinEur J Neurosci 2006232027ndash2034
11 Bennett DA Schneider JA Arvanitakis Z Wilson RS Overview and findings from thereligious orders study Curr Alzheimer Res 20129628ndash645
12 Bennett DA Schneider JA Buchman AS Barnes LL Boyle PA Wilson RS Overviewand findings from the rush Memory and Aging Project Curr Alzheimer Res 20129646ndash663
13 Bennett DA Schneider JA Wilson RS Bienias JL Arnold SE Neurofibrillary tanglesmediate the association of amyloid load with clinical Alzheimer disease and level ofcognitive function Arch Neurol 200461378ndash384
14 Saykin AJ Shen L Yao X et al Genetic studies of quantitative MCI and AD phe-notypes in ADNI Progress opportunities and plans Alzheimers Dement 201511792ndash814
15 Lim AS Srivastava GP Yu L et al 24-hour rhythms of DNA methylation and theirrelation with rhythms of RNA expression in the human dorsolateral prefrontal cortexPLoS Genet 201410e1004792
16 Rogers JF Nafziger AN Bertino JS Jr Pharmacogenetics affects dosing efficacy andtoxicity of cytochrome P450-metabolized drugs Am J Med 2002113746ndash750
17 Anzenbacher P Anzenbacherova E Cytochromes P450 and metabolism of xeno-biotics Cell Mol Life Sci 200158737ndash747
18 Meyer RP Gehlhaus M Knoth R Volk B Expression and function of cytochromep450 in brain drug metabolism Curr Drug Metab 20078297ndash306
19 Purcell S Neale B Todd-Brown K et al PLINK a tool set for whole-genomeassociation and population-based linkage analyses Am J Hum Genet 200781559ndash575
20 R A Language and Environment for Statistical Computing [computer program]Version 310 Vienna R Foundation for Statistical Computing 2015
21 Package RMINC Voxel-Wise Morphometry Using RMINC [computer program]Montreal McGill University 2010
22 Ramanan VK Risacher SL Nho K et al APOE and BCHE as modulators of cerebralamyloid deposition a florbetapir PET genome-wide association study Mol Psychiatry201419351ndash357
23 Braak H Braak E Neuropathological stageing of Alzheimer-related changes ActaNeuropathol 19918220
24 Tapiola T Alafuzoff I Herukka S et al Cerebrospinal fluid β-amyloid 42 and tauproteins as biomarkers of Alzheimer-type pathologic changes in the brain ArchNeurol 200966382ndash389
25 Sperling R Mormino E Johnson K The evolution of preclinical Alzheimerrsquos diseaseimplications for prevention trials Neuron 201484608ndash622
26 Wang D Jiang Z Shen Z et al Functional evaluation of genetic and environmentalregulators of p450 mRNA levels PLoS One 20116e24900
27 Sim SC Risinger C Dahl ML et al A common novel CYP2C19 gene variant causesultrarapid drug metabolism relevant for the drug response to proton pump inhibitorsand antidepressants Clin Pharmacol Ther 200679103ndash113
28 Ishii G Suzuki A Oshino S Shiraishi H Otani K CYP2C19 polymorphism affectspersonality traits of Japanese females Neurosci Lett 200741177ndash80
29 Ercan B Ayaz L Cicek D Tamer L Role of CYP2C9 and CYP2C19 polymorphismsin patients with atherosclerosis Cell Biochem Funct 200826309ndash313
30 Westphal C Konkel A SchunckWH CYP-eicosanoidsmdasha new link between omega-3 fatty acids and cardiac disease Prostaglandins Other Lipid Mediat 20119699ndash108
31 Cheng ZN Shu Y Liu ZQ Wang LS Ou-Yang DS Zhou HH Role of cytochromeP450 in estradiol metabolism in vitro Acta Pharmacol Sin 200122148ndash154
32 Giraud SN Caron CM Pham-Dinh D Kitabgi P Nicot AB Estradiol inhibits ongoingautoimmune neuroinflammation and NFkappaB-dependent CCL2 expression in re-active astrocytes Proc Natl Acad Sci USA 20101078416ndash8421
33 Xu H Wang R Zhang YW Zhang X Estrogen beta-amyloid metabolismtraffickingand Alzheimerrsquos disease Ann NY Acad Sci 20061089324ndash342
34 Amtul Z Uhrig M Wang L Rozmahel RF Beyreuther K Detrimental effects ofarachidonic acid and its metabolites in cellular and mouse models of Alzheimerrsquosdisease structural insight Neurobiol Aging 201233831e21ndashe31
35 Sarkar P Narayanan J Harder DR Differential effect of amyloid beta on the cyto-chrome P450 epoxygenase activity in rat brain Neuroscience 2011194241ndash249
36 Kisler K Nelson AR Montagne A Zlokovic BV Cerebral blood flow regulation andneurovascular dysfunction in Alzheimer disease Nat Rev Neurosci 201718419ndash434
37 Bibi Z Role of cytochrome P450 in drug interactions Nutr Metab (Lond) 200852738 Ownby RL Crocco E Acevedo A John V Loewenstein D Depression and risk for
Alzheimer disease systematic review meta-analysis and metaregression analysisArch Gen Psychiatry 200663530ndash538
39 Sheline YI West T Yarasheski K et al An antidepressant decreases CSF Abetaproduction in healthy individuals and in transgenic AD mice Sci Transl Med 20146236re234
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 9
DOI 101212NXG000000000000021620184 Neurol Genet
Andreacutea L Benedet Lei Yu Aureacutelie Labbe et al variant mitigates Alzheimer disease pathophysiology in vivo and postmortemCYP2C19
This information is current as of January 30 2018
ServicesUpdated Information amp
httpngneurologyorgcontent41e216fullhtmlincluding high resolution figures can be found at
References httpngneurologyorgcontent41e216fullhtmlref-list-1
This article cites 37 articles 2 of which you can access for free at
Subspecialty Collections
httpngneurologyorgcgicollectionpetPET
httpngneurologyorgcgicollectionassociation_studies_in_geneticsAssociation studies in genetics
httpngneurologyorgcgicollectionalzheimers_diseaseAlzheimers diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpngneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpngneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
reserved Online ISSN 2376-7839Published by Wolters Kluwer Health Inc on behalf of the American Academy of Neurology All rightsan open-access online-only continuous publication journal Copyright Copyright copy 2018 The Author(s)
is an official journal of the American Academy of Neurology Published since April 2015 it isNeurol Genet

PhenotypesFrom the ADNI we obtained demographics neurocognitivescores CSF and plasmatic data available after passing rigor-ous quality control The preprocessed imaging and geneticdata underwent additional processing and quality controlbefore the statistical analysis in our study as described belowFrom Rush all the data used in this project to perform sta-tistical analyses were already processed and quality controlledwith very rigorous criteria
In the ADNI cohort the brain Aβ load was estimated usingthe [18F]florbetapir PET standardized uptake value ratio(SUVR details are provided in figure 1 and e-methods)Separately Aβ and hyperphosphorylated tau (p-tau) lev-els were measured in the CSF and plasma These data wereobtained by the Biomarkers Consortium CSF and thedescription of the methodology regarding the sampleacquisition processing and analysis is available atthe ADNI website (adniloniuscedudata-samplesbio-specimen-data)
In the Rush-ROSMAP cohort postmortem data of the Aβload and paired helical filament (PHF) tau tangle density wereobtained from 8 brain regions using immunohistochemistry13
Cognitive function was evaluated in both the ADNI and Rush-ROSMAP For the ADNI the global cognition scores weredeveloped by making a z-score of the sum of the memory andthe executive function composite scores In the Rush-ROSMAP the global cognitive scores were based on 17 cognitiveperformance tests and we included only participants whounderwent autopsies thus the last valid cognitive scoreproximate to death was used for this analysis (for furtherdetails see e-Methods httplinkslwwcomNXGA20)
We used gene expression data to examine cis expression quan-titative trait loci (eQTL) For the ADNI total RNAwas obtainedfrom the peripheral bloodmdashat the same visit when the imagingdata were obtainedmdashand after quality control was hybridized tothe Affymetrix Human Genome U219 array plate as described14
The normalized and quality-controlled expression data14 wereused For the Rush-ROSMAP RNA was extracted from thegray matter of dorsolateral prefrontal cortex blocks usingthe miRNeasy Mini Kit (Qiagen Venlo Netherlands) and theRNase-free DNase Set (Qiagen) These samples were quantifiedand qualified as described15 and then sequenced on the IlluminaHiSeq with 101 bp paired-end reads The normalized andquality-controlled expression data were used In both analysesthe RNA integrity measurement was used as a covariate
Figure 1 [18F]florbetapir standardized uptake value ratio analytical method
Flowchart showing acquisition methods (purple) imageprocessing (blue) and outcomes (green) ADNI = Alz-heimerrsquos Disease Neuroimaging Initiative GM = graymatter ROI = region of interest SUVR = standardizeduptake value ratio WM = white matter
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 3
Genotyping imputation and gene selectionWe interrogated single nucleotide polymorphisms (SNPs)from the CYP genes of the 5mdashprobablymdashmost importantmetabolizers CYP3A4 CYP2D6 CYP2C9 CYP1A2 andCYP2C19316ndash18 Methods involved in DNA processing aredetailed in the supplementary methods section (httplinkslwwcomNXGA20) Using PLINK19 the SNPs within thesegenes were recoded based on the dominant model with re-spect to the MAs and the ones in high linkage disequilibrium(LD) (r2 gt 08) were removed from the analysis to avoidunnecessary testing A total of 28 SNPs were used in the initialstep of the association analysis
For Rush-ROSMAP cohorts DNA was extracted genotypedimputed and quality controlled according to the proceduresdescribed in e-Methods (httplinkslwwcomNXGA20)
Statistical analysisThe statistical tests were performed in R statistical software20
where linear regressionmodels were used to test the associationbetween the genotypes and the phenotypes in both cohortsseparately All statistical models included the covariates of agesex and APOE-e4 carriage status except for the models inwhich global cognition was the outcome measure where theAPOE-e4 status was replaced by years of education The initialanalysismdashusing the global [18F]florbetapir SUVR asa phenotypemdashwas performed to interrogate a total of 28 SNPs(table e-1 httplinkslwwcomNXGA19) To correct formultiple testing the statistical significance of the SNP discoverywas set at p lt 00017 using a Bonferroni correction for 28 testsand a type I error α = 005 The significant SNPs were furthertested for associations with other phenotypes in ADNI andRush-ROSMAP cohorts To test for group differences in thediagnostic status we added diagnosis as a covariate in the re-gression models and also tested the interaction between thediagnostic status and desired independent variable Effect sizecalculations (Cohen d) were also used
A voxel-based analysis was performed using the RMINCimaging tool21 where parametric images were obtained con-trasting the [18F]florbetapir SUVR between the genotypegroups of the SNP found in the discovery analysis Afterrandom field theory correction for multiple comparisons the tvalue threshold of significance was leminus31 (p le 0001)
Mediation analysis was used to examine whether the SNPassociations with cognition were mediated through the effect ofAβ To test this hypothesis first the association of CYP SNPswith global cognitive performance was evaluated using linearregression models Then if the association was significant themeasure of Aβ was added to this model to check whether theSNP associations with cognitionwere attenuated because of Aβ
ResultsDemographics and general information of the ADNI andRush-ROSMAP cohorts are presented in table 1 After
imaging and genetic quality control a total of 338 ADNIparticipants were studied including 186 CN 105 single- ormultiple-domain amnestic MCI and 47 AD A total of 738Rush-ROSMAP participants were analyzed including 301CN 179 amnestic MCI and 258 AD As expected in bothcohorts CN individuals had higher MMSE scores and fewerwere APOE-e4 carriers as compared to both MCI and ADThere was no difference in sex or years of formal educationbetween diagnostic groups in both cohorts (details are pro-vided in table e-2 httplinkslwwcomNXGA19)
CYP2C19 polymorphism is associated with theglobal [18F]florbetapir SUVR in the ADNI cohortThe discovery analysis tested the association of 28 SNPs fromCYP genes with the global [18F]florbetapir SUVR The anal-ysis unveiled an association of an SNP (rs4388808) in theCYP2C19 gene (t = minus343 p = 00006) (figure 2A) Theminor allele frequency (MAF) of rs4388808G in the ADNIcohort was 019 and the minor allele (MA) carriers displayedless [18F]florbetapir binding as compared to the noncarriersIt is important that the association remained significant whenadjusting the model for diagnosis In addition no interactionwas found between the SNP and diagnostic status The esti-mated effect size of rs4388808 in the whole sample wasCohen d = 036 Of interest the effect size was greater for thesubset of APOE-e4 carriers (Cohen d = 051) as compared toAPOE-e4 noncarriers (Cohen d = 033)
MA carriers of the CYP2C19 polymorphismhaveless Aβ in AD-related regions in theADNI cohortTo identify the brain areas responsible for the global differ-ence observed in [18F]florbetapir uptake revealed in the dis-covery analysis we performed a voxel-wise analysis Theanalysis showed that carriers of rs4388808G had less [18F]florbetapir binding in the frontal posterior cingulate andinferior temporal cortices (figure 3) with the voxels in theseregions presenting p values equal to or less than 0001
Findings with brain imaging are corroboratedby CSF data in the ADNI cohortBecause of the well-known relationship between brain and CSFlevels of Aβ we tested the association between rs4388808 andCSF Aβ Because not all participants in the ADNI sample hadCSF data we performed the analysis in a subset of 260 indi-viduals As expected MA carriers presented more CSF Aβ thannoncarriers (t = 294 p = 0003) (figure 2B) The CYP2C19polymorphismwas associatedwith the Aβp-tau ratio (t = 229p = 002) and higher ratios were found in carriers ofrs4388808G (figure 2C) No association was found using p-tauas a single outcome measure (t = minus124 p = 021)
Association between the CYP2C19polymorphism and Aβ is exclusive to the Aβbrain levels in the ADNI cohortTo examine whether the CYP2C19 polymorphism is also as-sociated with Aβ levels outside the brain we tested the
4 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
association of rs4388808 with plasmatic levels of Aβ ina subsample of 113 participants We did not find an associa-tion between the SNP and free (t = minus060 p = 054) or total(t = 049 p = 062) plasmatic Aβ1-42 levels (figure 2D) in-dicating that the findings reported above are specific tothe CNS
Results from theADNI cohortwere generalizedto the Rush-ROSMAP cohortWe next generalized the findings to postmortem indices of theAβ load and to neurofibrillary tangles in Rush-ROSMAPcohorts From the 8 brain regions where the Aβ load wasmeasured we selected 6 regions (angular gyrus anterior cin-gulate entorhinal cortex and mesial and inferior temporalcortices) and calculated an average per subject These 6 regionswere chosen to match with the brain areas where we detecteddifferences between genotype groups in the voxel-wise analysisWhen testing the association between the average of the Aβload and the CYP2C19 polymorphism we found thatrs4388808G (MAF = 015) carriers had a lower Aβ load thannoncarriers (t = minus215 p = 003) (figure 2E) consistent withfindings from the ADNI cohort The difference betweenrs4388808G carriers and noncarriers in PHF tau tangle densitydid not reach statistical significance (t = minus165 p = 009)
MA carriers of the CYP2C19 polymorphism hadbetter cognitive performance thannoncarriersin both cohortsTo examine whether the CYP2C19 polymorphism affects thecognitive performance we used composite measures of globalcognition as the outcome in linear regression models In theADNI cohort there was a clear tendency to an associationwith the CYP2C19 polymorphism (t = 192 p = 005) (figure2F) while in Rush-ROSMAP cohorts the association wassuch that rs4388808G carriers had a higher cognition thannoncarriers (t = 208 p = 003) (figure 2G)
The effect of the CYP2C19 polymorphism oncognition is mediated by AβA mediation analysis was used to determine whether theCYP2C19 polymorphism was directly or indirectly associatedwith cognitive performance We retested the association be-tween the CYP2C19 polymorphism and cognition but nowadding the measures of [18F]florbetapir or Aβ load in themodel Neither of the previous genotype-phenotype associa-tions remained significant indicating that the observed effectof the genotype was mediated by Aβ
Association between rs4388808 and geneexpression of CYP2C19 suggests that the SNPis functionalTo check whether the polymorphism has an effect in the geneexpression we tested the association between rs4388808 andCYP2C19 RNA levels In the ADNI cohort with 304 partic-ipants for this analysis we found a cis eQTL such that MAcarriers displayed higher RNA levels in the blood (t = 238 p =001) (figure 2H) In Rush-ROSMAP cohorts very low levelsof CYP2C19 expression were detected in postmortem brainsand the data did not provide consistent results (data notshown) as found in the blood expression data from the ADNI
DiscussionIn the present study we found an association between theCYP2C19 polymorphism and Aβ burden across the spectrumof AD in which carriers of rs4388808G presented a less Aβload and downstream cognitive impairment This associationwas initially detected using the [18F]florbetapir SUVR andposteriorly confirmed with CSF Aβ in the ADNI cohort Noassociation was found between the polymorphism and di-agnostic groups suggesting that the SNP is associated with anamyloid load rather than with the disease status Effect size
Table 1 Demographic and key characteristics of the samples
ADNI Rush-ROSMAP
CN MCI AD CN MCI AD
No of participants 186 105 47 301 179 258
Males n () 91 (489) 64 (61) 26 (553) 182 (605) 101 (564) 154 (597)
Age y mean (SD) 7570 (663) 7429 (812) 7567 (737) 837 (519)a3 8526 (435) 8595 (381)
APOE-laquo4 () 52 (28)a1 57 (543)a1 30 (638)a1 49 (164)a4 50 (279)a4 112 (434)a4
MMSE mean (SD) 2909 (114)a2 2724 (195)a2 2034 (366)a2 2869 (142)a5 2791 (183)a5 2549 (517)a5
Education y 1641 (275) 1667 (268) 1613 (247) 1653 (374) 1634 (347) 1666 (394)
MA n ()b 67 (19) 36 (10) 17 (5) 87 (11) 52 (7) 63 (85)
Abbreviations AD = Alzheimer disease ADNI = AD Neuroimaging Initiative CN = cognitively normal MA = minor allele MAP = Memory and Aging ProjectMCI = mild cognitive impairment MMSE = Mini-Mental State Examination ROS = Religious Orders Studya Statistically different from the other groups of the same sample 1p = 23 times 10minus7 2p = 2 times 10minus16 3p = 92 times 10minus8 4p = 17 times 10minus11 and 5p = 2 times 10minus16b MA (rs4388808G) percentage relative to its major sample (ADNI or Rush-ROSMAP)
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 5
analysis suggests that the benefic effects of the polymorphismare larger in APOE-e4 carriers as compared to noncarriers orthe whole sample in which the effect is similar to what hasbeen described for a BCHE gene polymorphism22 Sub-sequently this association was generalized to postmortemdata from the Rush-ROSMAP cohort Finally both cohortsshowed a protective association between the MA of CYP2C19and cognition It is important that subsequent mediationanalysis suggested that the effects of CYP2C19 on cognitionwere mediated by Aβ
Overall these results support the notion that the MA ofCYP2C19 (rs4388808) is a protective variant against Aβ anddownstream cognitive impairment Of interest voxel-wiseanalysis in in vivo participants revealed that the MA was as-sociated with reduced Aβ burden in AD-related regions in thefrontal inferior temporal and posterior cingulate cortices23
In addition the CYP2C19 SNP was associated with the CSFAβp-tau ratio but not with plasmatic Aβ Beyond the fact thatthe Aβp-tau ratio is postulated to better represent neuriticplaques than the single CSF Aβ information24 these results
Figure 2 Comparison between noncarriers (minus) and carriers (+) of the minor allele of rs4388808 (CYP2C19)
In the ADNI cohort a difference was observedin the brain Aβ load (A) CSF Aβ levels (B) andCSF Aβp-tau ratio (C) but no difference wasdetected in plasmatic levels of Aβ (D) Resultswere generalized to postmortem data of Rush-ROSMAP cohorts where a concordant patternwas observed in the Aβ load (percentage areaoccupied by amyloid) (E) A tendency towardsignificance in global cognitive performance (F)was found in the ADNI cohort while in theRush-ROSMAP it was significant (G) CYP2C19expression levels were also different betweennoncarriers (minus) and carriers (+) of the minorallele of rs4388808 (H) in the ADNI cohort Alllinear models were adjusted for age sex andAPOE-e4 carriage status with the exception ofthe models using global cognition and geneexpression which were adjusted for age sexand years of education or RNA integrity re-spectively Aβ = amyloid-β ADNI = AlzheimerrsquosDisease Neuroimaging Initiative MAP = Mem-ory and Aging Project p-tau = phosphorylatedtau ROS = Religious Orders Study SUVR =standardized uptake value ratio
6 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
provide further evidence that the polymorphism affects thebrain accumulation of neuritic plaques in a tissue- and disease-specific manner24 In both cohorts the referred poly-morphism was not associated with PHF tau or single CSFp-tau agreeing with the framework that Aβ and tau haveindependent upstream triggers25 In line with overall signifi-cant results the CYP2C19 polymorphism also presenteda protective effect in global cognition being supported byprevious studies that linked Aβ pathology to cognitivechanges13 Indeed the mediation analysis supported that theassociation between the genetic factor and cognitive abnor-malities is likely due to the upstream accumulation of Aβ
Levels of the CYP2C19 protein are associated with thers4388808 polymorphism26 which is an intronic variant of theCYP2C19 gene Indeed we found a similar association showingthat MA carriers expressed higher levels of CYP2C19 whencompared with noncarriers suggesting that the SNP affects theexpression of its gene To corroborate this finding we retrievedthe SNPs that were previously excluded because of high LD andverified that none was associated with CYP2C19 expression inthe blood (data not shown) reinforcing the idea that rs4388808is functional This association could not be detected in post-mortem data probably because of the low CYP expression orbecause of its limited detection by the method used7
Despite the SNP not being widely studied to provide in-formation about the association with other phenotypes other
polymorphisms in the gene have been correlated with meta-bolic variability atherosclerosis and behavioral traits27ndash29
CYP2C19 expression has been detected in several brainregions67 however its function in the CNS has not been fullyelucidated Some reports have suggested that besides its roleat metabolizing exogenous substrates CYP2C19 also partic-ipates in other important biological cascades such as themetabolism of serotonin and sexual hormones1 and the me-tabolism of the arachidonic acid (AA) where it functions as anepoxygenase30
The biochemical mechanism underlying the association ofCYP2C19 with Aβ remains speculative A study has shownthat Aβ is able to act as a foreign body and trigger PORactivating CYPs to initiate catabolic reactions9 In turnCYP2C19 is involved in the catalysis of estradiol31 Becauseseveral studies have reported the beneficial effects of estrogenson ADmdashdemonstrating that these steroids are able to act asanti-inflammatory compounds32 and even inhibit Aβproduction33mdashone could think that MA carriers of rs4388808have a reduced expression of this CYP in the brain or that theyhave an increased structural incompatibility betweenCYP2C19 and estradiol Consequently MA carriers couldpresent a reduction in the catabolism of estrogen and a re-duction in the Aβ load Alternatively the Aβ load would beaffected by the levels of epoxyeicosatrienoic acids (EETs)produced via the metabolism of AA by CYP epoxygenases Inthe brain studies have shown that EETs have beneficial effectsat regulating blood flow cortical angiogenesis and at pro-moting anti-inflammatory reactions (for review see reference2) By contrast when AA is metabolized via other cascadesthe protective effect of EETs is lost and there is a possibilityof induction of Aβ production andor accumulation34 Inaddition it was demonstrated that Aβ is able to reduceepoxygenase activity and consequently decrease EET pro-duction35 probably leading to more susceptibility to neuronaldamage (for review see reference 36) In this scenario itwould be possible that the MA of rs4388808 increases theexpression of CYP2C19 leading to an ldquoAβ-resistantrdquo epox-ygenase metabolism of AA with a maintained production ofEETs and a more protective phenotype against amyloidosis
It is also plausible to think however that the associationdescribed here between theCYP2C19 polymorphism and Aβis indirect being mediated by the effect of a drug CYP2C19is known to metabolize several drugs37 including medi-cations to treat depression which has been mentioned asa risk factor for AD (for review see reference 38) Cit-alopram for example is an antidepressant metabolized byCYP2C19 that has been shown to decrease Aβ production39
Thus one may think that the polymorphism rs4388808 isthen associated with an improved action of citalopramleading to the decreased brain Aβ burden rather than as-sociated with Aβ metabolism itself Because it is difficult totrack all the medications the study participants have takenbefore been enrolled it is unfeasible at the moment to checkwhether this association holds true
Figure 3 T-statistical parametric maps showing differencesbetween noncarriers (minus) and carriers (+) of theminor allele of rs4388808 (CYP2C19)
T-statistical parametric maps superimposed on average structural MRI showbrain regions with lower standardized uptake value ratio (SUVR) values inminor allele carriers (CYP2C19 (+)) of the polymorphism of CYP2C19 Statisticaldifferences overlap with brain regions vulnerable to Alzheimer disease path-ophysiology such as the posterior cingulate frontal and temporal cortices
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 7
The interpretation of the results should take into consider-ation a few limitations The use of the 2 cohorts does not allowus to detect and measure Aβ pathology with the samemethodology inserting in the analysis some variability specificto the method used However despite being measured withdifferent techniques both the imaging radiotracer and theantibody used in the brain tissue are expected to bind to thefibrillar form of Aβ13 Similarly the cognitive composite scoresfrom the 2 cohorts have some differences that should beconsidered There are also some intrinsic characteristics of thesamples that have to be taken into account (1) the inclusioncriteria of each study (2) the average age difference betweenthe studies and (3) and the disease stage in which Aβmeasures were performedmdashat the end or during the diseaseprocess Sample size restrictions impose some degree ofcaution when interpreting findings including for examplethe lack of association between the genotype and plasmaticlevels of Aβ or cognition in the ADNI cohort In additionboth cohorts are mostly composed of non-Latino Caucasianslimiting the extrapolation of the present findings to otherpopulation groups Longitudinal analysis together with func-tional genomics and biochemistry experiments not per-formed here would also be necessary to determine the effectof the SNP in the protein function as well as to support anytheoretical framework proposed in the Discussion section
Results obtained from the 2 independent cohorts providecompelling evidences linking the CYP2C19 polymorphismand Aβ pathology suggesting that the MA of rs4388808confers protective effects against Aβ accumulation in the brainand its downstream cognitive consequences These resultscould have implication for antiamyloid clinical trial designs aspreclinical rs4388808G would present a protective factoragainst the amyloid load Therefore the biological mechanismby which the genetic variation would alter Aβ build up andclearance merits further investigation
Author contributionsAndrea L Benedet study concept design analysis and in-terpretation of data composition of figures and drafting themanuscriptLei Yu analysis and interpretation of data and drafting themanuscript Aurelie Labbe supervision of statistical analysisand critical review of the manuscript for intellectual contentSulantha Mathotaarachchi Tharick A Pascoal Monica Shinand Min-Su Kang processing of image data and drafting themanuscript study concept design and drafting the manu-script Serge Gauthier Guy A Rouleau and Judes Poiriercritical review of the manuscript for intellectual content Da-vid A Bennett drafting the manuscript and critical review ofthe manuscript for intellectual content Pedro Rosa-Netostudy concept design study supervision and critical review ofthe manuscript for intellectual content
AcknowledgmentData collection and sharing for this project was funded by theAlzheimerrsquos Disease Neuroimaging Initiative (ADNI) (NIH
grant U01 AG024904) and the DOD ADNI (Department ofDefense award number W81XWH-12-2-0012) The ADNI isfunded by the National Institute on Aging the NationalInstitute of Biomedical Imaging and Bioengineering andthrough generous contributions from the following AbbViethe Alzheimerrsquos Association the Alzheimerrsquos Drug DiscoveryFoundation Araclon Biotech BioClinica Inc BiogenBristol-Myers Squibb CereSpir Inc Cogstate Eisai IncElan Pharmaceuticals Inc Eli Lilly and Company Euro-immun F Hoffmann-La Roche Ltd and its affiliated companyGenentech Inc Fujirebio GE Healthcare IXICO LtdJanssen Alzheimer Immunotherapy Research amp Develop-ment LLC Johnson amp Johnson Pharmaceutical Research ampDevelopment LLC Lumosity Lundbeck Merck amp Co IncMeso Scale Diagnostics LLC NeuroRx Research NeurotrackTechnologies Novartis Pharmaceuticals Corporation PfizerInc Piramal Imaging Servier Takeda Pharmaceutical Com-pany and Transition Therapeutics The Canadian Institutes ofHealth Research is providing funds to support ADNI clinicalsites in Canada Private sector contributions are facilitated bythe Foundation for the NIH (fnihorg) The granteeorganization is the Northern California Institute for Researchand Education and the study is coordinated by the AlzheimerrsquosTherapeutic Research Institute at the University of SouthernCalifornia ADNI data are disseminated by the Laboratory forNeuro Imaging at the University of Southern California
Study fundingThis work was also supported by the Canadian Institutes ofHealth Research (CIHR) (MOP-11-51-31) the AlzheimerrsquosAssociation (NIRG-08-92090) the Canadian Consortium onNeurodegeneration in Aging (CIHR-CCNA) and theWeston Brain Institute Andrea L Benedet is supported bythe CAPES FoundationmdashBrazil (032713-1) Pedro Rosa-Neto is supported by the Fonds de la recherche en sante duQuebec (chercheur boursier) Serge Gauthier and PedroRosa-Neto are members of the CIHR Canadian Consortiumon Neurodegeneration in Aging The Religious Orders Studyand the Rush Memory and Aging Project are funded bythe National Institute on Aging (P30AG10161 andR01AG17917) and the Illinois Department of Public Health
DisclosureAL Benedet has received research support from the CAPESFoundation L Yu has received research support from theNIH A Labbe and S Mathotaarachchi report no disclosuresTA Pascoal has received research support from thePREVENT-AD PhD scholarship M Shin and M-S Kangreport no disclosures S Gauthier has received honoraria forserving on the scientific advisory boards of Alzheon AxovantBoehringer Ingelheim Lilly Lundbeck Novartis SchwabeTauRx on the DSMB of a study sponsored by Eisai and forstudies run by the ADCS and by the ATRI has receivedspeaker honoraria from Schwabe serves on the editorial boardof Alzheimerrsquos amp Dementia has served as a consultant forKalgene and has received research support from the CIHRthe NIH Lundbeck Roche and Lilly GA Rouleau has
8 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
received research support from the CIHR the ALS Society ofCanada and the ALS Association J Poirier serves on theeditorial board of the Journal of Alzheimerrsquos Disease receivespublishing royalties from Trecarre and has received researchsupport from the JL Levesque Foundation DA Bennettserves on the scientific advisory boards of Vigorous MindsTakeda Pharmaceuticals AbbVie EABs (Indian Institute ofScience) DMCs (HRS and MIDUS) and the National Ad-visory Council on Aging is a consultant for Takeda Phar-maceuticals and AbbVie has received research support fromthe NIH and serves on the editorial boards of NeurologyCurrent Alzheimer Research and Neuroepidemiology P Rosa-Neto serves on the scientific advisory boards of EnigmaRadiopharmaceuticals and Kalgene serves on the editorialboard of Frontiers in Aging Neurosciences and has receivedresearch support fromEnigma Radiopharmaceuticals Eli Lillythe CIHR the FRQS Merck (Sharp amp Dohme CorporationMcGill Faculty of Medicine Grant for Translational Re-search) the Weston Brain Institute the Alzheimerrsquos Associa-tion Brain Canada and CQDM Funding information anddisclosures are provided at the end of the article Full disclo-sure form information provided by the authors is availablewith the full text of this article at NeurologyorgNG
Received June 8 2017 Accepted in final form December 4 2017
References1 Miksys S Tyndale RF Cytochrome P450-mediated drug metabolism in the brain
J Psychiatry Neurosci 201338152ndash1632 Iliff JJ Jia J Nelson J Goyagi T Klaus J Alkayed NJ Epoxyeicosanoid signaling in
CNS function and disease Prostaglandins Other Lipid Mediat 20109168ndash843 Stingl JC Brockmoller J Viviani R Genetic variability of drug-metabolizing enzymes
the dual impact on psychiatric therapy and regulation of brain function Mol Psy-chiatry 201318273ndash287
4 McCann SJ Pond SM James KM Le Couteur DG The association between poly-morphisms in the cytochrome P-450 2D6 gene and Parkinsonrsquos disease a case-controlstudy and meta-analysis J Neurol Sci 199715350ndash53
5 Chace C Pang D Weng C et al Variants in CYP17 and CYP19 cytochrome P450genes are associated with onset of Alzheimerrsquos disease in women with down syn-drome JAlzheimer Dis 201228601ndash612
6 McFayden MC Melvin WT Murray GI Regional distribution of individual forms of cyto-chrome P450 mRNA in normal adult human brain Biochem Pharmacol 199855825ndash830
7 Booth Depaz IM Toselli F Wilce PA Gillam EM Differential expression of cyto-chrome P450 enzymes from the CYP2C subfamily in the human brain Drug MetabDispos 201543353ndash357
8 Van Ess PJ Pedersen WA Culmsee C Mattson MP Blouin RA Elevated hepatic anddepressed renal cytochrome P450 activity in the Tg2576 transgenic mouse model ofAlzheimerrsquos disease J Neurochem 200280571ndash578
9 Pappolla MA Omar RA Chyan YJ et al Induction of NADPH cytochrome P450reductase by the Alzheimer beta-protein amyloid as a ldquoforeign bodyrdquo J Neurochem200178121ndash128
10 Liu M Sun A Shin EJ et al Expression of microsomal epoxide hydrolase is elevated inAlzheimerrsquos hippocampus and induced by exogenous beta-amyloid and trimethyl-tinEur J Neurosci 2006232027ndash2034
11 Bennett DA Schneider JA Arvanitakis Z Wilson RS Overview and findings from thereligious orders study Curr Alzheimer Res 20129628ndash645
12 Bennett DA Schneider JA Buchman AS Barnes LL Boyle PA Wilson RS Overviewand findings from the rush Memory and Aging Project Curr Alzheimer Res 20129646ndash663
13 Bennett DA Schneider JA Wilson RS Bienias JL Arnold SE Neurofibrillary tanglesmediate the association of amyloid load with clinical Alzheimer disease and level ofcognitive function Arch Neurol 200461378ndash384
14 Saykin AJ Shen L Yao X et al Genetic studies of quantitative MCI and AD phe-notypes in ADNI Progress opportunities and plans Alzheimers Dement 201511792ndash814
15 Lim AS Srivastava GP Yu L et al 24-hour rhythms of DNA methylation and theirrelation with rhythms of RNA expression in the human dorsolateral prefrontal cortexPLoS Genet 201410e1004792
16 Rogers JF Nafziger AN Bertino JS Jr Pharmacogenetics affects dosing efficacy andtoxicity of cytochrome P450-metabolized drugs Am J Med 2002113746ndash750
17 Anzenbacher P Anzenbacherova E Cytochromes P450 and metabolism of xeno-biotics Cell Mol Life Sci 200158737ndash747
18 Meyer RP Gehlhaus M Knoth R Volk B Expression and function of cytochromep450 in brain drug metabolism Curr Drug Metab 20078297ndash306
19 Purcell S Neale B Todd-Brown K et al PLINK a tool set for whole-genomeassociation and population-based linkage analyses Am J Hum Genet 200781559ndash575
20 R A Language and Environment for Statistical Computing [computer program]Version 310 Vienna R Foundation for Statistical Computing 2015
21 Package RMINC Voxel-Wise Morphometry Using RMINC [computer program]Montreal McGill University 2010
22 Ramanan VK Risacher SL Nho K et al APOE and BCHE as modulators of cerebralamyloid deposition a florbetapir PET genome-wide association study Mol Psychiatry201419351ndash357
23 Braak H Braak E Neuropathological stageing of Alzheimer-related changes ActaNeuropathol 19918220
24 Tapiola T Alafuzoff I Herukka S et al Cerebrospinal fluid β-amyloid 42 and tauproteins as biomarkers of Alzheimer-type pathologic changes in the brain ArchNeurol 200966382ndash389
25 Sperling R Mormino E Johnson K The evolution of preclinical Alzheimerrsquos diseaseimplications for prevention trials Neuron 201484608ndash622
26 Wang D Jiang Z Shen Z et al Functional evaluation of genetic and environmentalregulators of p450 mRNA levels PLoS One 20116e24900
27 Sim SC Risinger C Dahl ML et al A common novel CYP2C19 gene variant causesultrarapid drug metabolism relevant for the drug response to proton pump inhibitorsand antidepressants Clin Pharmacol Ther 200679103ndash113
28 Ishii G Suzuki A Oshino S Shiraishi H Otani K CYP2C19 polymorphism affectspersonality traits of Japanese females Neurosci Lett 200741177ndash80
29 Ercan B Ayaz L Cicek D Tamer L Role of CYP2C9 and CYP2C19 polymorphismsin patients with atherosclerosis Cell Biochem Funct 200826309ndash313
30 Westphal C Konkel A SchunckWH CYP-eicosanoidsmdasha new link between omega-3 fatty acids and cardiac disease Prostaglandins Other Lipid Mediat 20119699ndash108
31 Cheng ZN Shu Y Liu ZQ Wang LS Ou-Yang DS Zhou HH Role of cytochromeP450 in estradiol metabolism in vitro Acta Pharmacol Sin 200122148ndash154
32 Giraud SN Caron CM Pham-Dinh D Kitabgi P Nicot AB Estradiol inhibits ongoingautoimmune neuroinflammation and NFkappaB-dependent CCL2 expression in re-active astrocytes Proc Natl Acad Sci USA 20101078416ndash8421
33 Xu H Wang R Zhang YW Zhang X Estrogen beta-amyloid metabolismtraffickingand Alzheimerrsquos disease Ann NY Acad Sci 20061089324ndash342
34 Amtul Z Uhrig M Wang L Rozmahel RF Beyreuther K Detrimental effects ofarachidonic acid and its metabolites in cellular and mouse models of Alzheimerrsquosdisease structural insight Neurobiol Aging 201233831e21ndashe31
35 Sarkar P Narayanan J Harder DR Differential effect of amyloid beta on the cyto-chrome P450 epoxygenase activity in rat brain Neuroscience 2011194241ndash249
36 Kisler K Nelson AR Montagne A Zlokovic BV Cerebral blood flow regulation andneurovascular dysfunction in Alzheimer disease Nat Rev Neurosci 201718419ndash434
37 Bibi Z Role of cytochrome P450 in drug interactions Nutr Metab (Lond) 200852738 Ownby RL Crocco E Acevedo A John V Loewenstein D Depression and risk for
Alzheimer disease systematic review meta-analysis and metaregression analysisArch Gen Psychiatry 200663530ndash538
39 Sheline YI West T Yarasheski K et al An antidepressant decreases CSF Abetaproduction in healthy individuals and in transgenic AD mice Sci Transl Med 20146236re234
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 9
DOI 101212NXG000000000000021620184 Neurol Genet
Andreacutea L Benedet Lei Yu Aureacutelie Labbe et al variant mitigates Alzheimer disease pathophysiology in vivo and postmortemCYP2C19
This information is current as of January 30 2018
ServicesUpdated Information amp
httpngneurologyorgcontent41e216fullhtmlincluding high resolution figures can be found at
References httpngneurologyorgcontent41e216fullhtmlref-list-1
This article cites 37 articles 2 of which you can access for free at
Subspecialty Collections
httpngneurologyorgcgicollectionpetPET
httpngneurologyorgcgicollectionassociation_studies_in_geneticsAssociation studies in genetics
httpngneurologyorgcgicollectionalzheimers_diseaseAlzheimers diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpngneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpngneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
reserved Online ISSN 2376-7839Published by Wolters Kluwer Health Inc on behalf of the American Academy of Neurology All rightsan open-access online-only continuous publication journal Copyright Copyright copy 2018 The Author(s)
is an official journal of the American Academy of Neurology Published since April 2015 it isNeurol Genet

Genotyping imputation and gene selectionWe interrogated single nucleotide polymorphisms (SNPs)from the CYP genes of the 5mdashprobablymdashmost importantmetabolizers CYP3A4 CYP2D6 CYP2C9 CYP1A2 andCYP2C19316ndash18 Methods involved in DNA processing aredetailed in the supplementary methods section (httplinkslwwcomNXGA20) Using PLINK19 the SNPs within thesegenes were recoded based on the dominant model with re-spect to the MAs and the ones in high linkage disequilibrium(LD) (r2 gt 08) were removed from the analysis to avoidunnecessary testing A total of 28 SNPs were used in the initialstep of the association analysis
For Rush-ROSMAP cohorts DNA was extracted genotypedimputed and quality controlled according to the proceduresdescribed in e-Methods (httplinkslwwcomNXGA20)
Statistical analysisThe statistical tests were performed in R statistical software20
where linear regressionmodels were used to test the associationbetween the genotypes and the phenotypes in both cohortsseparately All statistical models included the covariates of agesex and APOE-e4 carriage status except for the models inwhich global cognition was the outcome measure where theAPOE-e4 status was replaced by years of education The initialanalysismdashusing the global [18F]florbetapir SUVR asa phenotypemdashwas performed to interrogate a total of 28 SNPs(table e-1 httplinkslwwcomNXGA19) To correct formultiple testing the statistical significance of the SNP discoverywas set at p lt 00017 using a Bonferroni correction for 28 testsand a type I error α = 005 The significant SNPs were furthertested for associations with other phenotypes in ADNI andRush-ROSMAP cohorts To test for group differences in thediagnostic status we added diagnosis as a covariate in the re-gression models and also tested the interaction between thediagnostic status and desired independent variable Effect sizecalculations (Cohen d) were also used
A voxel-based analysis was performed using the RMINCimaging tool21 where parametric images were obtained con-trasting the [18F]florbetapir SUVR between the genotypegroups of the SNP found in the discovery analysis Afterrandom field theory correction for multiple comparisons the tvalue threshold of significance was leminus31 (p le 0001)
Mediation analysis was used to examine whether the SNPassociations with cognition were mediated through the effect ofAβ To test this hypothesis first the association of CYP SNPswith global cognitive performance was evaluated using linearregression models Then if the association was significant themeasure of Aβ was added to this model to check whether theSNP associations with cognitionwere attenuated because of Aβ
ResultsDemographics and general information of the ADNI andRush-ROSMAP cohorts are presented in table 1 After
imaging and genetic quality control a total of 338 ADNIparticipants were studied including 186 CN 105 single- ormultiple-domain amnestic MCI and 47 AD A total of 738Rush-ROSMAP participants were analyzed including 301CN 179 amnestic MCI and 258 AD As expected in bothcohorts CN individuals had higher MMSE scores and fewerwere APOE-e4 carriers as compared to both MCI and ADThere was no difference in sex or years of formal educationbetween diagnostic groups in both cohorts (details are pro-vided in table e-2 httplinkslwwcomNXGA19)
CYP2C19 polymorphism is associated with theglobal [18F]florbetapir SUVR in the ADNI cohortThe discovery analysis tested the association of 28 SNPs fromCYP genes with the global [18F]florbetapir SUVR The anal-ysis unveiled an association of an SNP (rs4388808) in theCYP2C19 gene (t = minus343 p = 00006) (figure 2A) Theminor allele frequency (MAF) of rs4388808G in the ADNIcohort was 019 and the minor allele (MA) carriers displayedless [18F]florbetapir binding as compared to the noncarriersIt is important that the association remained significant whenadjusting the model for diagnosis In addition no interactionwas found between the SNP and diagnostic status The esti-mated effect size of rs4388808 in the whole sample wasCohen d = 036 Of interest the effect size was greater for thesubset of APOE-e4 carriers (Cohen d = 051) as compared toAPOE-e4 noncarriers (Cohen d = 033)
MA carriers of the CYP2C19 polymorphismhaveless Aβ in AD-related regions in theADNI cohortTo identify the brain areas responsible for the global differ-ence observed in [18F]florbetapir uptake revealed in the dis-covery analysis we performed a voxel-wise analysis Theanalysis showed that carriers of rs4388808G had less [18F]florbetapir binding in the frontal posterior cingulate andinferior temporal cortices (figure 3) with the voxels in theseregions presenting p values equal to or less than 0001
Findings with brain imaging are corroboratedby CSF data in the ADNI cohortBecause of the well-known relationship between brain and CSFlevels of Aβ we tested the association between rs4388808 andCSF Aβ Because not all participants in the ADNI sample hadCSF data we performed the analysis in a subset of 260 indi-viduals As expected MA carriers presented more CSF Aβ thannoncarriers (t = 294 p = 0003) (figure 2B) The CYP2C19polymorphismwas associatedwith the Aβp-tau ratio (t = 229p = 002) and higher ratios were found in carriers ofrs4388808G (figure 2C) No association was found using p-tauas a single outcome measure (t = minus124 p = 021)
Association between the CYP2C19polymorphism and Aβ is exclusive to the Aβbrain levels in the ADNI cohortTo examine whether the CYP2C19 polymorphism is also as-sociated with Aβ levels outside the brain we tested the
4 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
association of rs4388808 with plasmatic levels of Aβ ina subsample of 113 participants We did not find an associa-tion between the SNP and free (t = minus060 p = 054) or total(t = 049 p = 062) plasmatic Aβ1-42 levels (figure 2D) in-dicating that the findings reported above are specific tothe CNS
Results from theADNI cohortwere generalizedto the Rush-ROSMAP cohortWe next generalized the findings to postmortem indices of theAβ load and to neurofibrillary tangles in Rush-ROSMAPcohorts From the 8 brain regions where the Aβ load wasmeasured we selected 6 regions (angular gyrus anterior cin-gulate entorhinal cortex and mesial and inferior temporalcortices) and calculated an average per subject These 6 regionswere chosen to match with the brain areas where we detecteddifferences between genotype groups in the voxel-wise analysisWhen testing the association between the average of the Aβload and the CYP2C19 polymorphism we found thatrs4388808G (MAF = 015) carriers had a lower Aβ load thannoncarriers (t = minus215 p = 003) (figure 2E) consistent withfindings from the ADNI cohort The difference betweenrs4388808G carriers and noncarriers in PHF tau tangle densitydid not reach statistical significance (t = minus165 p = 009)
MA carriers of the CYP2C19 polymorphism hadbetter cognitive performance thannoncarriersin both cohortsTo examine whether the CYP2C19 polymorphism affects thecognitive performance we used composite measures of globalcognition as the outcome in linear regression models In theADNI cohort there was a clear tendency to an associationwith the CYP2C19 polymorphism (t = 192 p = 005) (figure2F) while in Rush-ROSMAP cohorts the association wassuch that rs4388808G carriers had a higher cognition thannoncarriers (t = 208 p = 003) (figure 2G)
The effect of the CYP2C19 polymorphism oncognition is mediated by AβA mediation analysis was used to determine whether theCYP2C19 polymorphism was directly or indirectly associatedwith cognitive performance We retested the association be-tween the CYP2C19 polymorphism and cognition but nowadding the measures of [18F]florbetapir or Aβ load in themodel Neither of the previous genotype-phenotype associa-tions remained significant indicating that the observed effectof the genotype was mediated by Aβ
Association between rs4388808 and geneexpression of CYP2C19 suggests that the SNPis functionalTo check whether the polymorphism has an effect in the geneexpression we tested the association between rs4388808 andCYP2C19 RNA levels In the ADNI cohort with 304 partic-ipants for this analysis we found a cis eQTL such that MAcarriers displayed higher RNA levels in the blood (t = 238 p =001) (figure 2H) In Rush-ROSMAP cohorts very low levelsof CYP2C19 expression were detected in postmortem brainsand the data did not provide consistent results (data notshown) as found in the blood expression data from the ADNI
DiscussionIn the present study we found an association between theCYP2C19 polymorphism and Aβ burden across the spectrumof AD in which carriers of rs4388808G presented a less Aβload and downstream cognitive impairment This associationwas initially detected using the [18F]florbetapir SUVR andposteriorly confirmed with CSF Aβ in the ADNI cohort Noassociation was found between the polymorphism and di-agnostic groups suggesting that the SNP is associated with anamyloid load rather than with the disease status Effect size
Table 1 Demographic and key characteristics of the samples
ADNI Rush-ROSMAP
CN MCI AD CN MCI AD
No of participants 186 105 47 301 179 258
Males n () 91 (489) 64 (61) 26 (553) 182 (605) 101 (564) 154 (597)
Age y mean (SD) 7570 (663) 7429 (812) 7567 (737) 837 (519)a3 8526 (435) 8595 (381)
APOE-laquo4 () 52 (28)a1 57 (543)a1 30 (638)a1 49 (164)a4 50 (279)a4 112 (434)a4
MMSE mean (SD) 2909 (114)a2 2724 (195)a2 2034 (366)a2 2869 (142)a5 2791 (183)a5 2549 (517)a5
Education y 1641 (275) 1667 (268) 1613 (247) 1653 (374) 1634 (347) 1666 (394)
MA n ()b 67 (19) 36 (10) 17 (5) 87 (11) 52 (7) 63 (85)
Abbreviations AD = Alzheimer disease ADNI = AD Neuroimaging Initiative CN = cognitively normal MA = minor allele MAP = Memory and Aging ProjectMCI = mild cognitive impairment MMSE = Mini-Mental State Examination ROS = Religious Orders Studya Statistically different from the other groups of the same sample 1p = 23 times 10minus7 2p = 2 times 10minus16 3p = 92 times 10minus8 4p = 17 times 10minus11 and 5p = 2 times 10minus16b MA (rs4388808G) percentage relative to its major sample (ADNI or Rush-ROSMAP)
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 5
analysis suggests that the benefic effects of the polymorphismare larger in APOE-e4 carriers as compared to noncarriers orthe whole sample in which the effect is similar to what hasbeen described for a BCHE gene polymorphism22 Sub-sequently this association was generalized to postmortemdata from the Rush-ROSMAP cohort Finally both cohortsshowed a protective association between the MA of CYP2C19and cognition It is important that subsequent mediationanalysis suggested that the effects of CYP2C19 on cognitionwere mediated by Aβ
Overall these results support the notion that the MA ofCYP2C19 (rs4388808) is a protective variant against Aβ anddownstream cognitive impairment Of interest voxel-wiseanalysis in in vivo participants revealed that the MA was as-sociated with reduced Aβ burden in AD-related regions in thefrontal inferior temporal and posterior cingulate cortices23
In addition the CYP2C19 SNP was associated with the CSFAβp-tau ratio but not with plasmatic Aβ Beyond the fact thatthe Aβp-tau ratio is postulated to better represent neuriticplaques than the single CSF Aβ information24 these results
Figure 2 Comparison between noncarriers (minus) and carriers (+) of the minor allele of rs4388808 (CYP2C19)
In the ADNI cohort a difference was observedin the brain Aβ load (A) CSF Aβ levels (B) andCSF Aβp-tau ratio (C) but no difference wasdetected in plasmatic levels of Aβ (D) Resultswere generalized to postmortem data of Rush-ROSMAP cohorts where a concordant patternwas observed in the Aβ load (percentage areaoccupied by amyloid) (E) A tendency towardsignificance in global cognitive performance (F)was found in the ADNI cohort while in theRush-ROSMAP it was significant (G) CYP2C19expression levels were also different betweennoncarriers (minus) and carriers (+) of the minorallele of rs4388808 (H) in the ADNI cohort Alllinear models were adjusted for age sex andAPOE-e4 carriage status with the exception ofthe models using global cognition and geneexpression which were adjusted for age sexand years of education or RNA integrity re-spectively Aβ = amyloid-β ADNI = AlzheimerrsquosDisease Neuroimaging Initiative MAP = Mem-ory and Aging Project p-tau = phosphorylatedtau ROS = Religious Orders Study SUVR =standardized uptake value ratio
6 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
provide further evidence that the polymorphism affects thebrain accumulation of neuritic plaques in a tissue- and disease-specific manner24 In both cohorts the referred poly-morphism was not associated with PHF tau or single CSFp-tau agreeing with the framework that Aβ and tau haveindependent upstream triggers25 In line with overall signifi-cant results the CYP2C19 polymorphism also presenteda protective effect in global cognition being supported byprevious studies that linked Aβ pathology to cognitivechanges13 Indeed the mediation analysis supported that theassociation between the genetic factor and cognitive abnor-malities is likely due to the upstream accumulation of Aβ
Levels of the CYP2C19 protein are associated with thers4388808 polymorphism26 which is an intronic variant of theCYP2C19 gene Indeed we found a similar association showingthat MA carriers expressed higher levels of CYP2C19 whencompared with noncarriers suggesting that the SNP affects theexpression of its gene To corroborate this finding we retrievedthe SNPs that were previously excluded because of high LD andverified that none was associated with CYP2C19 expression inthe blood (data not shown) reinforcing the idea that rs4388808is functional This association could not be detected in post-mortem data probably because of the low CYP expression orbecause of its limited detection by the method used7
Despite the SNP not being widely studied to provide in-formation about the association with other phenotypes other
polymorphisms in the gene have been correlated with meta-bolic variability atherosclerosis and behavioral traits27ndash29
CYP2C19 expression has been detected in several brainregions67 however its function in the CNS has not been fullyelucidated Some reports have suggested that besides its roleat metabolizing exogenous substrates CYP2C19 also partic-ipates in other important biological cascades such as themetabolism of serotonin and sexual hormones1 and the me-tabolism of the arachidonic acid (AA) where it functions as anepoxygenase30
The biochemical mechanism underlying the association ofCYP2C19 with Aβ remains speculative A study has shownthat Aβ is able to act as a foreign body and trigger PORactivating CYPs to initiate catabolic reactions9 In turnCYP2C19 is involved in the catalysis of estradiol31 Becauseseveral studies have reported the beneficial effects of estrogenson ADmdashdemonstrating that these steroids are able to act asanti-inflammatory compounds32 and even inhibit Aβproduction33mdashone could think that MA carriers of rs4388808have a reduced expression of this CYP in the brain or that theyhave an increased structural incompatibility betweenCYP2C19 and estradiol Consequently MA carriers couldpresent a reduction in the catabolism of estrogen and a re-duction in the Aβ load Alternatively the Aβ load would beaffected by the levels of epoxyeicosatrienoic acids (EETs)produced via the metabolism of AA by CYP epoxygenases Inthe brain studies have shown that EETs have beneficial effectsat regulating blood flow cortical angiogenesis and at pro-moting anti-inflammatory reactions (for review see reference2) By contrast when AA is metabolized via other cascadesthe protective effect of EETs is lost and there is a possibilityof induction of Aβ production andor accumulation34 Inaddition it was demonstrated that Aβ is able to reduceepoxygenase activity and consequently decrease EET pro-duction35 probably leading to more susceptibility to neuronaldamage (for review see reference 36) In this scenario itwould be possible that the MA of rs4388808 increases theexpression of CYP2C19 leading to an ldquoAβ-resistantrdquo epox-ygenase metabolism of AA with a maintained production ofEETs and a more protective phenotype against amyloidosis
It is also plausible to think however that the associationdescribed here between theCYP2C19 polymorphism and Aβis indirect being mediated by the effect of a drug CYP2C19is known to metabolize several drugs37 including medi-cations to treat depression which has been mentioned asa risk factor for AD (for review see reference 38) Cit-alopram for example is an antidepressant metabolized byCYP2C19 that has been shown to decrease Aβ production39
Thus one may think that the polymorphism rs4388808 isthen associated with an improved action of citalopramleading to the decreased brain Aβ burden rather than as-sociated with Aβ metabolism itself Because it is difficult totrack all the medications the study participants have takenbefore been enrolled it is unfeasible at the moment to checkwhether this association holds true
Figure 3 T-statistical parametric maps showing differencesbetween noncarriers (minus) and carriers (+) of theminor allele of rs4388808 (CYP2C19)
T-statistical parametric maps superimposed on average structural MRI showbrain regions with lower standardized uptake value ratio (SUVR) values inminor allele carriers (CYP2C19 (+)) of the polymorphism of CYP2C19 Statisticaldifferences overlap with brain regions vulnerable to Alzheimer disease path-ophysiology such as the posterior cingulate frontal and temporal cortices
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 7
The interpretation of the results should take into consider-ation a few limitations The use of the 2 cohorts does not allowus to detect and measure Aβ pathology with the samemethodology inserting in the analysis some variability specificto the method used However despite being measured withdifferent techniques both the imaging radiotracer and theantibody used in the brain tissue are expected to bind to thefibrillar form of Aβ13 Similarly the cognitive composite scoresfrom the 2 cohorts have some differences that should beconsidered There are also some intrinsic characteristics of thesamples that have to be taken into account (1) the inclusioncriteria of each study (2) the average age difference betweenthe studies and (3) and the disease stage in which Aβmeasures were performedmdashat the end or during the diseaseprocess Sample size restrictions impose some degree ofcaution when interpreting findings including for examplethe lack of association between the genotype and plasmaticlevels of Aβ or cognition in the ADNI cohort In additionboth cohorts are mostly composed of non-Latino Caucasianslimiting the extrapolation of the present findings to otherpopulation groups Longitudinal analysis together with func-tional genomics and biochemistry experiments not per-formed here would also be necessary to determine the effectof the SNP in the protein function as well as to support anytheoretical framework proposed in the Discussion section
Results obtained from the 2 independent cohorts providecompelling evidences linking the CYP2C19 polymorphismand Aβ pathology suggesting that the MA of rs4388808confers protective effects against Aβ accumulation in the brainand its downstream cognitive consequences These resultscould have implication for antiamyloid clinical trial designs aspreclinical rs4388808G would present a protective factoragainst the amyloid load Therefore the biological mechanismby which the genetic variation would alter Aβ build up andclearance merits further investigation
Author contributionsAndrea L Benedet study concept design analysis and in-terpretation of data composition of figures and drafting themanuscriptLei Yu analysis and interpretation of data and drafting themanuscript Aurelie Labbe supervision of statistical analysisand critical review of the manuscript for intellectual contentSulantha Mathotaarachchi Tharick A Pascoal Monica Shinand Min-Su Kang processing of image data and drafting themanuscript study concept design and drafting the manu-script Serge Gauthier Guy A Rouleau and Judes Poiriercritical review of the manuscript for intellectual content Da-vid A Bennett drafting the manuscript and critical review ofthe manuscript for intellectual content Pedro Rosa-Netostudy concept design study supervision and critical review ofthe manuscript for intellectual content
AcknowledgmentData collection and sharing for this project was funded by theAlzheimerrsquos Disease Neuroimaging Initiative (ADNI) (NIH
grant U01 AG024904) and the DOD ADNI (Department ofDefense award number W81XWH-12-2-0012) The ADNI isfunded by the National Institute on Aging the NationalInstitute of Biomedical Imaging and Bioengineering andthrough generous contributions from the following AbbViethe Alzheimerrsquos Association the Alzheimerrsquos Drug DiscoveryFoundation Araclon Biotech BioClinica Inc BiogenBristol-Myers Squibb CereSpir Inc Cogstate Eisai IncElan Pharmaceuticals Inc Eli Lilly and Company Euro-immun F Hoffmann-La Roche Ltd and its affiliated companyGenentech Inc Fujirebio GE Healthcare IXICO LtdJanssen Alzheimer Immunotherapy Research amp Develop-ment LLC Johnson amp Johnson Pharmaceutical Research ampDevelopment LLC Lumosity Lundbeck Merck amp Co IncMeso Scale Diagnostics LLC NeuroRx Research NeurotrackTechnologies Novartis Pharmaceuticals Corporation PfizerInc Piramal Imaging Servier Takeda Pharmaceutical Com-pany and Transition Therapeutics The Canadian Institutes ofHealth Research is providing funds to support ADNI clinicalsites in Canada Private sector contributions are facilitated bythe Foundation for the NIH (fnihorg) The granteeorganization is the Northern California Institute for Researchand Education and the study is coordinated by the AlzheimerrsquosTherapeutic Research Institute at the University of SouthernCalifornia ADNI data are disseminated by the Laboratory forNeuro Imaging at the University of Southern California
Study fundingThis work was also supported by the Canadian Institutes ofHealth Research (CIHR) (MOP-11-51-31) the AlzheimerrsquosAssociation (NIRG-08-92090) the Canadian Consortium onNeurodegeneration in Aging (CIHR-CCNA) and theWeston Brain Institute Andrea L Benedet is supported bythe CAPES FoundationmdashBrazil (032713-1) Pedro Rosa-Neto is supported by the Fonds de la recherche en sante duQuebec (chercheur boursier) Serge Gauthier and PedroRosa-Neto are members of the CIHR Canadian Consortiumon Neurodegeneration in Aging The Religious Orders Studyand the Rush Memory and Aging Project are funded bythe National Institute on Aging (P30AG10161 andR01AG17917) and the Illinois Department of Public Health
DisclosureAL Benedet has received research support from the CAPESFoundation L Yu has received research support from theNIH A Labbe and S Mathotaarachchi report no disclosuresTA Pascoal has received research support from thePREVENT-AD PhD scholarship M Shin and M-S Kangreport no disclosures S Gauthier has received honoraria forserving on the scientific advisory boards of Alzheon AxovantBoehringer Ingelheim Lilly Lundbeck Novartis SchwabeTauRx on the DSMB of a study sponsored by Eisai and forstudies run by the ADCS and by the ATRI has receivedspeaker honoraria from Schwabe serves on the editorial boardof Alzheimerrsquos amp Dementia has served as a consultant forKalgene and has received research support from the CIHRthe NIH Lundbeck Roche and Lilly GA Rouleau has
8 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
received research support from the CIHR the ALS Society ofCanada and the ALS Association J Poirier serves on theeditorial board of the Journal of Alzheimerrsquos Disease receivespublishing royalties from Trecarre and has received researchsupport from the JL Levesque Foundation DA Bennettserves on the scientific advisory boards of Vigorous MindsTakeda Pharmaceuticals AbbVie EABs (Indian Institute ofScience) DMCs (HRS and MIDUS) and the National Ad-visory Council on Aging is a consultant for Takeda Phar-maceuticals and AbbVie has received research support fromthe NIH and serves on the editorial boards of NeurologyCurrent Alzheimer Research and Neuroepidemiology P Rosa-Neto serves on the scientific advisory boards of EnigmaRadiopharmaceuticals and Kalgene serves on the editorialboard of Frontiers in Aging Neurosciences and has receivedresearch support fromEnigma Radiopharmaceuticals Eli Lillythe CIHR the FRQS Merck (Sharp amp Dohme CorporationMcGill Faculty of Medicine Grant for Translational Re-search) the Weston Brain Institute the Alzheimerrsquos Associa-tion Brain Canada and CQDM Funding information anddisclosures are provided at the end of the article Full disclo-sure form information provided by the authors is availablewith the full text of this article at NeurologyorgNG
Received June 8 2017 Accepted in final form December 4 2017
References1 Miksys S Tyndale RF Cytochrome P450-mediated drug metabolism in the brain
J Psychiatry Neurosci 201338152ndash1632 Iliff JJ Jia J Nelson J Goyagi T Klaus J Alkayed NJ Epoxyeicosanoid signaling in
CNS function and disease Prostaglandins Other Lipid Mediat 20109168ndash843 Stingl JC Brockmoller J Viviani R Genetic variability of drug-metabolizing enzymes
the dual impact on psychiatric therapy and regulation of brain function Mol Psy-chiatry 201318273ndash287
4 McCann SJ Pond SM James KM Le Couteur DG The association between poly-morphisms in the cytochrome P-450 2D6 gene and Parkinsonrsquos disease a case-controlstudy and meta-analysis J Neurol Sci 199715350ndash53
5 Chace C Pang D Weng C et al Variants in CYP17 and CYP19 cytochrome P450genes are associated with onset of Alzheimerrsquos disease in women with down syn-drome JAlzheimer Dis 201228601ndash612
6 McFayden MC Melvin WT Murray GI Regional distribution of individual forms of cyto-chrome P450 mRNA in normal adult human brain Biochem Pharmacol 199855825ndash830
7 Booth Depaz IM Toselli F Wilce PA Gillam EM Differential expression of cyto-chrome P450 enzymes from the CYP2C subfamily in the human brain Drug MetabDispos 201543353ndash357
8 Van Ess PJ Pedersen WA Culmsee C Mattson MP Blouin RA Elevated hepatic anddepressed renal cytochrome P450 activity in the Tg2576 transgenic mouse model ofAlzheimerrsquos disease J Neurochem 200280571ndash578
9 Pappolla MA Omar RA Chyan YJ et al Induction of NADPH cytochrome P450reductase by the Alzheimer beta-protein amyloid as a ldquoforeign bodyrdquo J Neurochem200178121ndash128
10 Liu M Sun A Shin EJ et al Expression of microsomal epoxide hydrolase is elevated inAlzheimerrsquos hippocampus and induced by exogenous beta-amyloid and trimethyl-tinEur J Neurosci 2006232027ndash2034
11 Bennett DA Schneider JA Arvanitakis Z Wilson RS Overview and findings from thereligious orders study Curr Alzheimer Res 20129628ndash645
12 Bennett DA Schneider JA Buchman AS Barnes LL Boyle PA Wilson RS Overviewand findings from the rush Memory and Aging Project Curr Alzheimer Res 20129646ndash663
13 Bennett DA Schneider JA Wilson RS Bienias JL Arnold SE Neurofibrillary tanglesmediate the association of amyloid load with clinical Alzheimer disease and level ofcognitive function Arch Neurol 200461378ndash384
14 Saykin AJ Shen L Yao X et al Genetic studies of quantitative MCI and AD phe-notypes in ADNI Progress opportunities and plans Alzheimers Dement 201511792ndash814
15 Lim AS Srivastava GP Yu L et al 24-hour rhythms of DNA methylation and theirrelation with rhythms of RNA expression in the human dorsolateral prefrontal cortexPLoS Genet 201410e1004792
16 Rogers JF Nafziger AN Bertino JS Jr Pharmacogenetics affects dosing efficacy andtoxicity of cytochrome P450-metabolized drugs Am J Med 2002113746ndash750
17 Anzenbacher P Anzenbacherova E Cytochromes P450 and metabolism of xeno-biotics Cell Mol Life Sci 200158737ndash747
18 Meyer RP Gehlhaus M Knoth R Volk B Expression and function of cytochromep450 in brain drug metabolism Curr Drug Metab 20078297ndash306
19 Purcell S Neale B Todd-Brown K et al PLINK a tool set for whole-genomeassociation and population-based linkage analyses Am J Hum Genet 200781559ndash575
20 R A Language and Environment for Statistical Computing [computer program]Version 310 Vienna R Foundation for Statistical Computing 2015
21 Package RMINC Voxel-Wise Morphometry Using RMINC [computer program]Montreal McGill University 2010
22 Ramanan VK Risacher SL Nho K et al APOE and BCHE as modulators of cerebralamyloid deposition a florbetapir PET genome-wide association study Mol Psychiatry201419351ndash357
23 Braak H Braak E Neuropathological stageing of Alzheimer-related changes ActaNeuropathol 19918220
24 Tapiola T Alafuzoff I Herukka S et al Cerebrospinal fluid β-amyloid 42 and tauproteins as biomarkers of Alzheimer-type pathologic changes in the brain ArchNeurol 200966382ndash389
25 Sperling R Mormino E Johnson K The evolution of preclinical Alzheimerrsquos diseaseimplications for prevention trials Neuron 201484608ndash622
26 Wang D Jiang Z Shen Z et al Functional evaluation of genetic and environmentalregulators of p450 mRNA levels PLoS One 20116e24900
27 Sim SC Risinger C Dahl ML et al A common novel CYP2C19 gene variant causesultrarapid drug metabolism relevant for the drug response to proton pump inhibitorsand antidepressants Clin Pharmacol Ther 200679103ndash113
28 Ishii G Suzuki A Oshino S Shiraishi H Otani K CYP2C19 polymorphism affectspersonality traits of Japanese females Neurosci Lett 200741177ndash80
29 Ercan B Ayaz L Cicek D Tamer L Role of CYP2C9 and CYP2C19 polymorphismsin patients with atherosclerosis Cell Biochem Funct 200826309ndash313
30 Westphal C Konkel A SchunckWH CYP-eicosanoidsmdasha new link between omega-3 fatty acids and cardiac disease Prostaglandins Other Lipid Mediat 20119699ndash108
31 Cheng ZN Shu Y Liu ZQ Wang LS Ou-Yang DS Zhou HH Role of cytochromeP450 in estradiol metabolism in vitro Acta Pharmacol Sin 200122148ndash154
32 Giraud SN Caron CM Pham-Dinh D Kitabgi P Nicot AB Estradiol inhibits ongoingautoimmune neuroinflammation and NFkappaB-dependent CCL2 expression in re-active astrocytes Proc Natl Acad Sci USA 20101078416ndash8421
33 Xu H Wang R Zhang YW Zhang X Estrogen beta-amyloid metabolismtraffickingand Alzheimerrsquos disease Ann NY Acad Sci 20061089324ndash342
34 Amtul Z Uhrig M Wang L Rozmahel RF Beyreuther K Detrimental effects ofarachidonic acid and its metabolites in cellular and mouse models of Alzheimerrsquosdisease structural insight Neurobiol Aging 201233831e21ndashe31
35 Sarkar P Narayanan J Harder DR Differential effect of amyloid beta on the cyto-chrome P450 epoxygenase activity in rat brain Neuroscience 2011194241ndash249
36 Kisler K Nelson AR Montagne A Zlokovic BV Cerebral blood flow regulation andneurovascular dysfunction in Alzheimer disease Nat Rev Neurosci 201718419ndash434
37 Bibi Z Role of cytochrome P450 in drug interactions Nutr Metab (Lond) 200852738 Ownby RL Crocco E Acevedo A John V Loewenstein D Depression and risk for
Alzheimer disease systematic review meta-analysis and metaregression analysisArch Gen Psychiatry 200663530ndash538
39 Sheline YI West T Yarasheski K et al An antidepressant decreases CSF Abetaproduction in healthy individuals and in transgenic AD mice Sci Transl Med 20146236re234
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 9
DOI 101212NXG000000000000021620184 Neurol Genet
Andreacutea L Benedet Lei Yu Aureacutelie Labbe et al variant mitigates Alzheimer disease pathophysiology in vivo and postmortemCYP2C19
This information is current as of January 30 2018
ServicesUpdated Information amp
httpngneurologyorgcontent41e216fullhtmlincluding high resolution figures can be found at
References httpngneurologyorgcontent41e216fullhtmlref-list-1
This article cites 37 articles 2 of which you can access for free at
Subspecialty Collections
httpngneurologyorgcgicollectionpetPET
httpngneurologyorgcgicollectionassociation_studies_in_geneticsAssociation studies in genetics
httpngneurologyorgcgicollectionalzheimers_diseaseAlzheimers diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpngneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpngneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
reserved Online ISSN 2376-7839Published by Wolters Kluwer Health Inc on behalf of the American Academy of Neurology All rightsan open-access online-only continuous publication journal Copyright Copyright copy 2018 The Author(s)
is an official journal of the American Academy of Neurology Published since April 2015 it isNeurol Genet

association of rs4388808 with plasmatic levels of Aβ ina subsample of 113 participants We did not find an associa-tion between the SNP and free (t = minus060 p = 054) or total(t = 049 p = 062) plasmatic Aβ1-42 levels (figure 2D) in-dicating that the findings reported above are specific tothe CNS
Results from theADNI cohortwere generalizedto the Rush-ROSMAP cohortWe next generalized the findings to postmortem indices of theAβ load and to neurofibrillary tangles in Rush-ROSMAPcohorts From the 8 brain regions where the Aβ load wasmeasured we selected 6 regions (angular gyrus anterior cin-gulate entorhinal cortex and mesial and inferior temporalcortices) and calculated an average per subject These 6 regionswere chosen to match with the brain areas where we detecteddifferences between genotype groups in the voxel-wise analysisWhen testing the association between the average of the Aβload and the CYP2C19 polymorphism we found thatrs4388808G (MAF = 015) carriers had a lower Aβ load thannoncarriers (t = minus215 p = 003) (figure 2E) consistent withfindings from the ADNI cohort The difference betweenrs4388808G carriers and noncarriers in PHF tau tangle densitydid not reach statistical significance (t = minus165 p = 009)
MA carriers of the CYP2C19 polymorphism hadbetter cognitive performance thannoncarriersin both cohortsTo examine whether the CYP2C19 polymorphism affects thecognitive performance we used composite measures of globalcognition as the outcome in linear regression models In theADNI cohort there was a clear tendency to an associationwith the CYP2C19 polymorphism (t = 192 p = 005) (figure2F) while in Rush-ROSMAP cohorts the association wassuch that rs4388808G carriers had a higher cognition thannoncarriers (t = 208 p = 003) (figure 2G)
The effect of the CYP2C19 polymorphism oncognition is mediated by AβA mediation analysis was used to determine whether theCYP2C19 polymorphism was directly or indirectly associatedwith cognitive performance We retested the association be-tween the CYP2C19 polymorphism and cognition but nowadding the measures of [18F]florbetapir or Aβ load in themodel Neither of the previous genotype-phenotype associa-tions remained significant indicating that the observed effectof the genotype was mediated by Aβ
Association between rs4388808 and geneexpression of CYP2C19 suggests that the SNPis functionalTo check whether the polymorphism has an effect in the geneexpression we tested the association between rs4388808 andCYP2C19 RNA levels In the ADNI cohort with 304 partic-ipants for this analysis we found a cis eQTL such that MAcarriers displayed higher RNA levels in the blood (t = 238 p =001) (figure 2H) In Rush-ROSMAP cohorts very low levelsof CYP2C19 expression were detected in postmortem brainsand the data did not provide consistent results (data notshown) as found in the blood expression data from the ADNI
DiscussionIn the present study we found an association between theCYP2C19 polymorphism and Aβ burden across the spectrumof AD in which carriers of rs4388808G presented a less Aβload and downstream cognitive impairment This associationwas initially detected using the [18F]florbetapir SUVR andposteriorly confirmed with CSF Aβ in the ADNI cohort Noassociation was found between the polymorphism and di-agnostic groups suggesting that the SNP is associated with anamyloid load rather than with the disease status Effect size
Table 1 Demographic and key characteristics of the samples
ADNI Rush-ROSMAP
CN MCI AD CN MCI AD
No of participants 186 105 47 301 179 258
Males n () 91 (489) 64 (61) 26 (553) 182 (605) 101 (564) 154 (597)
Age y mean (SD) 7570 (663) 7429 (812) 7567 (737) 837 (519)a3 8526 (435) 8595 (381)
APOE-laquo4 () 52 (28)a1 57 (543)a1 30 (638)a1 49 (164)a4 50 (279)a4 112 (434)a4
MMSE mean (SD) 2909 (114)a2 2724 (195)a2 2034 (366)a2 2869 (142)a5 2791 (183)a5 2549 (517)a5
Education y 1641 (275) 1667 (268) 1613 (247) 1653 (374) 1634 (347) 1666 (394)
MA n ()b 67 (19) 36 (10) 17 (5) 87 (11) 52 (7) 63 (85)
Abbreviations AD = Alzheimer disease ADNI = AD Neuroimaging Initiative CN = cognitively normal MA = minor allele MAP = Memory and Aging ProjectMCI = mild cognitive impairment MMSE = Mini-Mental State Examination ROS = Religious Orders Studya Statistically different from the other groups of the same sample 1p = 23 times 10minus7 2p = 2 times 10minus16 3p = 92 times 10minus8 4p = 17 times 10minus11 and 5p = 2 times 10minus16b MA (rs4388808G) percentage relative to its major sample (ADNI or Rush-ROSMAP)
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 5
analysis suggests that the benefic effects of the polymorphismare larger in APOE-e4 carriers as compared to noncarriers orthe whole sample in which the effect is similar to what hasbeen described for a BCHE gene polymorphism22 Sub-sequently this association was generalized to postmortemdata from the Rush-ROSMAP cohort Finally both cohortsshowed a protective association between the MA of CYP2C19and cognition It is important that subsequent mediationanalysis suggested that the effects of CYP2C19 on cognitionwere mediated by Aβ
Overall these results support the notion that the MA ofCYP2C19 (rs4388808) is a protective variant against Aβ anddownstream cognitive impairment Of interest voxel-wiseanalysis in in vivo participants revealed that the MA was as-sociated with reduced Aβ burden in AD-related regions in thefrontal inferior temporal and posterior cingulate cortices23
In addition the CYP2C19 SNP was associated with the CSFAβp-tau ratio but not with plasmatic Aβ Beyond the fact thatthe Aβp-tau ratio is postulated to better represent neuriticplaques than the single CSF Aβ information24 these results
Figure 2 Comparison between noncarriers (minus) and carriers (+) of the minor allele of rs4388808 (CYP2C19)
In the ADNI cohort a difference was observedin the brain Aβ load (A) CSF Aβ levels (B) andCSF Aβp-tau ratio (C) but no difference wasdetected in plasmatic levels of Aβ (D) Resultswere generalized to postmortem data of Rush-ROSMAP cohorts where a concordant patternwas observed in the Aβ load (percentage areaoccupied by amyloid) (E) A tendency towardsignificance in global cognitive performance (F)was found in the ADNI cohort while in theRush-ROSMAP it was significant (G) CYP2C19expression levels were also different betweennoncarriers (minus) and carriers (+) of the minorallele of rs4388808 (H) in the ADNI cohort Alllinear models were adjusted for age sex andAPOE-e4 carriage status with the exception ofthe models using global cognition and geneexpression which were adjusted for age sexand years of education or RNA integrity re-spectively Aβ = amyloid-β ADNI = AlzheimerrsquosDisease Neuroimaging Initiative MAP = Mem-ory and Aging Project p-tau = phosphorylatedtau ROS = Religious Orders Study SUVR =standardized uptake value ratio
6 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
provide further evidence that the polymorphism affects thebrain accumulation of neuritic plaques in a tissue- and disease-specific manner24 In both cohorts the referred poly-morphism was not associated with PHF tau or single CSFp-tau agreeing with the framework that Aβ and tau haveindependent upstream triggers25 In line with overall signifi-cant results the CYP2C19 polymorphism also presenteda protective effect in global cognition being supported byprevious studies that linked Aβ pathology to cognitivechanges13 Indeed the mediation analysis supported that theassociation between the genetic factor and cognitive abnor-malities is likely due to the upstream accumulation of Aβ
Levels of the CYP2C19 protein are associated with thers4388808 polymorphism26 which is an intronic variant of theCYP2C19 gene Indeed we found a similar association showingthat MA carriers expressed higher levels of CYP2C19 whencompared with noncarriers suggesting that the SNP affects theexpression of its gene To corroborate this finding we retrievedthe SNPs that were previously excluded because of high LD andverified that none was associated with CYP2C19 expression inthe blood (data not shown) reinforcing the idea that rs4388808is functional This association could not be detected in post-mortem data probably because of the low CYP expression orbecause of its limited detection by the method used7
Despite the SNP not being widely studied to provide in-formation about the association with other phenotypes other
polymorphisms in the gene have been correlated with meta-bolic variability atherosclerosis and behavioral traits27ndash29
CYP2C19 expression has been detected in several brainregions67 however its function in the CNS has not been fullyelucidated Some reports have suggested that besides its roleat metabolizing exogenous substrates CYP2C19 also partic-ipates in other important biological cascades such as themetabolism of serotonin and sexual hormones1 and the me-tabolism of the arachidonic acid (AA) where it functions as anepoxygenase30
The biochemical mechanism underlying the association ofCYP2C19 with Aβ remains speculative A study has shownthat Aβ is able to act as a foreign body and trigger PORactivating CYPs to initiate catabolic reactions9 In turnCYP2C19 is involved in the catalysis of estradiol31 Becauseseveral studies have reported the beneficial effects of estrogenson ADmdashdemonstrating that these steroids are able to act asanti-inflammatory compounds32 and even inhibit Aβproduction33mdashone could think that MA carriers of rs4388808have a reduced expression of this CYP in the brain or that theyhave an increased structural incompatibility betweenCYP2C19 and estradiol Consequently MA carriers couldpresent a reduction in the catabolism of estrogen and a re-duction in the Aβ load Alternatively the Aβ load would beaffected by the levels of epoxyeicosatrienoic acids (EETs)produced via the metabolism of AA by CYP epoxygenases Inthe brain studies have shown that EETs have beneficial effectsat regulating blood flow cortical angiogenesis and at pro-moting anti-inflammatory reactions (for review see reference2) By contrast when AA is metabolized via other cascadesthe protective effect of EETs is lost and there is a possibilityof induction of Aβ production andor accumulation34 Inaddition it was demonstrated that Aβ is able to reduceepoxygenase activity and consequently decrease EET pro-duction35 probably leading to more susceptibility to neuronaldamage (for review see reference 36) In this scenario itwould be possible that the MA of rs4388808 increases theexpression of CYP2C19 leading to an ldquoAβ-resistantrdquo epox-ygenase metabolism of AA with a maintained production ofEETs and a more protective phenotype against amyloidosis
It is also plausible to think however that the associationdescribed here between theCYP2C19 polymorphism and Aβis indirect being mediated by the effect of a drug CYP2C19is known to metabolize several drugs37 including medi-cations to treat depression which has been mentioned asa risk factor for AD (for review see reference 38) Cit-alopram for example is an antidepressant metabolized byCYP2C19 that has been shown to decrease Aβ production39
Thus one may think that the polymorphism rs4388808 isthen associated with an improved action of citalopramleading to the decreased brain Aβ burden rather than as-sociated with Aβ metabolism itself Because it is difficult totrack all the medications the study participants have takenbefore been enrolled it is unfeasible at the moment to checkwhether this association holds true
Figure 3 T-statistical parametric maps showing differencesbetween noncarriers (minus) and carriers (+) of theminor allele of rs4388808 (CYP2C19)
T-statistical parametric maps superimposed on average structural MRI showbrain regions with lower standardized uptake value ratio (SUVR) values inminor allele carriers (CYP2C19 (+)) of the polymorphism of CYP2C19 Statisticaldifferences overlap with brain regions vulnerable to Alzheimer disease path-ophysiology such as the posterior cingulate frontal and temporal cortices
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 7
The interpretation of the results should take into consider-ation a few limitations The use of the 2 cohorts does not allowus to detect and measure Aβ pathology with the samemethodology inserting in the analysis some variability specificto the method used However despite being measured withdifferent techniques both the imaging radiotracer and theantibody used in the brain tissue are expected to bind to thefibrillar form of Aβ13 Similarly the cognitive composite scoresfrom the 2 cohorts have some differences that should beconsidered There are also some intrinsic characteristics of thesamples that have to be taken into account (1) the inclusioncriteria of each study (2) the average age difference betweenthe studies and (3) and the disease stage in which Aβmeasures were performedmdashat the end or during the diseaseprocess Sample size restrictions impose some degree ofcaution when interpreting findings including for examplethe lack of association between the genotype and plasmaticlevels of Aβ or cognition in the ADNI cohort In additionboth cohorts are mostly composed of non-Latino Caucasianslimiting the extrapolation of the present findings to otherpopulation groups Longitudinal analysis together with func-tional genomics and biochemistry experiments not per-formed here would also be necessary to determine the effectof the SNP in the protein function as well as to support anytheoretical framework proposed in the Discussion section
Results obtained from the 2 independent cohorts providecompelling evidences linking the CYP2C19 polymorphismand Aβ pathology suggesting that the MA of rs4388808confers protective effects against Aβ accumulation in the brainand its downstream cognitive consequences These resultscould have implication for antiamyloid clinical trial designs aspreclinical rs4388808G would present a protective factoragainst the amyloid load Therefore the biological mechanismby which the genetic variation would alter Aβ build up andclearance merits further investigation
Author contributionsAndrea L Benedet study concept design analysis and in-terpretation of data composition of figures and drafting themanuscriptLei Yu analysis and interpretation of data and drafting themanuscript Aurelie Labbe supervision of statistical analysisand critical review of the manuscript for intellectual contentSulantha Mathotaarachchi Tharick A Pascoal Monica Shinand Min-Su Kang processing of image data and drafting themanuscript study concept design and drafting the manu-script Serge Gauthier Guy A Rouleau and Judes Poiriercritical review of the manuscript for intellectual content Da-vid A Bennett drafting the manuscript and critical review ofthe manuscript for intellectual content Pedro Rosa-Netostudy concept design study supervision and critical review ofthe manuscript for intellectual content
AcknowledgmentData collection and sharing for this project was funded by theAlzheimerrsquos Disease Neuroimaging Initiative (ADNI) (NIH
grant U01 AG024904) and the DOD ADNI (Department ofDefense award number W81XWH-12-2-0012) The ADNI isfunded by the National Institute on Aging the NationalInstitute of Biomedical Imaging and Bioengineering andthrough generous contributions from the following AbbViethe Alzheimerrsquos Association the Alzheimerrsquos Drug DiscoveryFoundation Araclon Biotech BioClinica Inc BiogenBristol-Myers Squibb CereSpir Inc Cogstate Eisai IncElan Pharmaceuticals Inc Eli Lilly and Company Euro-immun F Hoffmann-La Roche Ltd and its affiliated companyGenentech Inc Fujirebio GE Healthcare IXICO LtdJanssen Alzheimer Immunotherapy Research amp Develop-ment LLC Johnson amp Johnson Pharmaceutical Research ampDevelopment LLC Lumosity Lundbeck Merck amp Co IncMeso Scale Diagnostics LLC NeuroRx Research NeurotrackTechnologies Novartis Pharmaceuticals Corporation PfizerInc Piramal Imaging Servier Takeda Pharmaceutical Com-pany and Transition Therapeutics The Canadian Institutes ofHealth Research is providing funds to support ADNI clinicalsites in Canada Private sector contributions are facilitated bythe Foundation for the NIH (fnihorg) The granteeorganization is the Northern California Institute for Researchand Education and the study is coordinated by the AlzheimerrsquosTherapeutic Research Institute at the University of SouthernCalifornia ADNI data are disseminated by the Laboratory forNeuro Imaging at the University of Southern California
Study fundingThis work was also supported by the Canadian Institutes ofHealth Research (CIHR) (MOP-11-51-31) the AlzheimerrsquosAssociation (NIRG-08-92090) the Canadian Consortium onNeurodegeneration in Aging (CIHR-CCNA) and theWeston Brain Institute Andrea L Benedet is supported bythe CAPES FoundationmdashBrazil (032713-1) Pedro Rosa-Neto is supported by the Fonds de la recherche en sante duQuebec (chercheur boursier) Serge Gauthier and PedroRosa-Neto are members of the CIHR Canadian Consortiumon Neurodegeneration in Aging The Religious Orders Studyand the Rush Memory and Aging Project are funded bythe National Institute on Aging (P30AG10161 andR01AG17917) and the Illinois Department of Public Health
DisclosureAL Benedet has received research support from the CAPESFoundation L Yu has received research support from theNIH A Labbe and S Mathotaarachchi report no disclosuresTA Pascoal has received research support from thePREVENT-AD PhD scholarship M Shin and M-S Kangreport no disclosures S Gauthier has received honoraria forserving on the scientific advisory boards of Alzheon AxovantBoehringer Ingelheim Lilly Lundbeck Novartis SchwabeTauRx on the DSMB of a study sponsored by Eisai and forstudies run by the ADCS and by the ATRI has receivedspeaker honoraria from Schwabe serves on the editorial boardof Alzheimerrsquos amp Dementia has served as a consultant forKalgene and has received research support from the CIHRthe NIH Lundbeck Roche and Lilly GA Rouleau has
8 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
received research support from the CIHR the ALS Society ofCanada and the ALS Association J Poirier serves on theeditorial board of the Journal of Alzheimerrsquos Disease receivespublishing royalties from Trecarre and has received researchsupport from the JL Levesque Foundation DA Bennettserves on the scientific advisory boards of Vigorous MindsTakeda Pharmaceuticals AbbVie EABs (Indian Institute ofScience) DMCs (HRS and MIDUS) and the National Ad-visory Council on Aging is a consultant for Takeda Phar-maceuticals and AbbVie has received research support fromthe NIH and serves on the editorial boards of NeurologyCurrent Alzheimer Research and Neuroepidemiology P Rosa-Neto serves on the scientific advisory boards of EnigmaRadiopharmaceuticals and Kalgene serves on the editorialboard of Frontiers in Aging Neurosciences and has receivedresearch support fromEnigma Radiopharmaceuticals Eli Lillythe CIHR the FRQS Merck (Sharp amp Dohme CorporationMcGill Faculty of Medicine Grant for Translational Re-search) the Weston Brain Institute the Alzheimerrsquos Associa-tion Brain Canada and CQDM Funding information anddisclosures are provided at the end of the article Full disclo-sure form information provided by the authors is availablewith the full text of this article at NeurologyorgNG
Received June 8 2017 Accepted in final form December 4 2017
References1 Miksys S Tyndale RF Cytochrome P450-mediated drug metabolism in the brain
J Psychiatry Neurosci 201338152ndash1632 Iliff JJ Jia J Nelson J Goyagi T Klaus J Alkayed NJ Epoxyeicosanoid signaling in
CNS function and disease Prostaglandins Other Lipid Mediat 20109168ndash843 Stingl JC Brockmoller J Viviani R Genetic variability of drug-metabolizing enzymes
the dual impact on psychiatric therapy and regulation of brain function Mol Psy-chiatry 201318273ndash287
4 McCann SJ Pond SM James KM Le Couteur DG The association between poly-morphisms in the cytochrome P-450 2D6 gene and Parkinsonrsquos disease a case-controlstudy and meta-analysis J Neurol Sci 199715350ndash53
5 Chace C Pang D Weng C et al Variants in CYP17 and CYP19 cytochrome P450genes are associated with onset of Alzheimerrsquos disease in women with down syn-drome JAlzheimer Dis 201228601ndash612
6 McFayden MC Melvin WT Murray GI Regional distribution of individual forms of cyto-chrome P450 mRNA in normal adult human brain Biochem Pharmacol 199855825ndash830
7 Booth Depaz IM Toselli F Wilce PA Gillam EM Differential expression of cyto-chrome P450 enzymes from the CYP2C subfamily in the human brain Drug MetabDispos 201543353ndash357
8 Van Ess PJ Pedersen WA Culmsee C Mattson MP Blouin RA Elevated hepatic anddepressed renal cytochrome P450 activity in the Tg2576 transgenic mouse model ofAlzheimerrsquos disease J Neurochem 200280571ndash578
9 Pappolla MA Omar RA Chyan YJ et al Induction of NADPH cytochrome P450reductase by the Alzheimer beta-protein amyloid as a ldquoforeign bodyrdquo J Neurochem200178121ndash128
10 Liu M Sun A Shin EJ et al Expression of microsomal epoxide hydrolase is elevated inAlzheimerrsquos hippocampus and induced by exogenous beta-amyloid and trimethyl-tinEur J Neurosci 2006232027ndash2034
11 Bennett DA Schneider JA Arvanitakis Z Wilson RS Overview and findings from thereligious orders study Curr Alzheimer Res 20129628ndash645
12 Bennett DA Schneider JA Buchman AS Barnes LL Boyle PA Wilson RS Overviewand findings from the rush Memory and Aging Project Curr Alzheimer Res 20129646ndash663
13 Bennett DA Schneider JA Wilson RS Bienias JL Arnold SE Neurofibrillary tanglesmediate the association of amyloid load with clinical Alzheimer disease and level ofcognitive function Arch Neurol 200461378ndash384
14 Saykin AJ Shen L Yao X et al Genetic studies of quantitative MCI and AD phe-notypes in ADNI Progress opportunities and plans Alzheimers Dement 201511792ndash814
15 Lim AS Srivastava GP Yu L et al 24-hour rhythms of DNA methylation and theirrelation with rhythms of RNA expression in the human dorsolateral prefrontal cortexPLoS Genet 201410e1004792
16 Rogers JF Nafziger AN Bertino JS Jr Pharmacogenetics affects dosing efficacy andtoxicity of cytochrome P450-metabolized drugs Am J Med 2002113746ndash750
17 Anzenbacher P Anzenbacherova E Cytochromes P450 and metabolism of xeno-biotics Cell Mol Life Sci 200158737ndash747
18 Meyer RP Gehlhaus M Knoth R Volk B Expression and function of cytochromep450 in brain drug metabolism Curr Drug Metab 20078297ndash306
19 Purcell S Neale B Todd-Brown K et al PLINK a tool set for whole-genomeassociation and population-based linkage analyses Am J Hum Genet 200781559ndash575
20 R A Language and Environment for Statistical Computing [computer program]Version 310 Vienna R Foundation for Statistical Computing 2015
21 Package RMINC Voxel-Wise Morphometry Using RMINC [computer program]Montreal McGill University 2010
22 Ramanan VK Risacher SL Nho K et al APOE and BCHE as modulators of cerebralamyloid deposition a florbetapir PET genome-wide association study Mol Psychiatry201419351ndash357
23 Braak H Braak E Neuropathological stageing of Alzheimer-related changes ActaNeuropathol 19918220
24 Tapiola T Alafuzoff I Herukka S et al Cerebrospinal fluid β-amyloid 42 and tauproteins as biomarkers of Alzheimer-type pathologic changes in the brain ArchNeurol 200966382ndash389
25 Sperling R Mormino E Johnson K The evolution of preclinical Alzheimerrsquos diseaseimplications for prevention trials Neuron 201484608ndash622
26 Wang D Jiang Z Shen Z et al Functional evaluation of genetic and environmentalregulators of p450 mRNA levels PLoS One 20116e24900
27 Sim SC Risinger C Dahl ML et al A common novel CYP2C19 gene variant causesultrarapid drug metabolism relevant for the drug response to proton pump inhibitorsand antidepressants Clin Pharmacol Ther 200679103ndash113
28 Ishii G Suzuki A Oshino S Shiraishi H Otani K CYP2C19 polymorphism affectspersonality traits of Japanese females Neurosci Lett 200741177ndash80
29 Ercan B Ayaz L Cicek D Tamer L Role of CYP2C9 and CYP2C19 polymorphismsin patients with atherosclerosis Cell Biochem Funct 200826309ndash313
30 Westphal C Konkel A SchunckWH CYP-eicosanoidsmdasha new link between omega-3 fatty acids and cardiac disease Prostaglandins Other Lipid Mediat 20119699ndash108
31 Cheng ZN Shu Y Liu ZQ Wang LS Ou-Yang DS Zhou HH Role of cytochromeP450 in estradiol metabolism in vitro Acta Pharmacol Sin 200122148ndash154
32 Giraud SN Caron CM Pham-Dinh D Kitabgi P Nicot AB Estradiol inhibits ongoingautoimmune neuroinflammation and NFkappaB-dependent CCL2 expression in re-active astrocytes Proc Natl Acad Sci USA 20101078416ndash8421
33 Xu H Wang R Zhang YW Zhang X Estrogen beta-amyloid metabolismtraffickingand Alzheimerrsquos disease Ann NY Acad Sci 20061089324ndash342
34 Amtul Z Uhrig M Wang L Rozmahel RF Beyreuther K Detrimental effects ofarachidonic acid and its metabolites in cellular and mouse models of Alzheimerrsquosdisease structural insight Neurobiol Aging 201233831e21ndashe31
35 Sarkar P Narayanan J Harder DR Differential effect of amyloid beta on the cyto-chrome P450 epoxygenase activity in rat brain Neuroscience 2011194241ndash249
36 Kisler K Nelson AR Montagne A Zlokovic BV Cerebral blood flow regulation andneurovascular dysfunction in Alzheimer disease Nat Rev Neurosci 201718419ndash434
37 Bibi Z Role of cytochrome P450 in drug interactions Nutr Metab (Lond) 200852738 Ownby RL Crocco E Acevedo A John V Loewenstein D Depression and risk for
Alzheimer disease systematic review meta-analysis and metaregression analysisArch Gen Psychiatry 200663530ndash538
39 Sheline YI West T Yarasheski K et al An antidepressant decreases CSF Abetaproduction in healthy individuals and in transgenic AD mice Sci Transl Med 20146236re234
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 9
DOI 101212NXG000000000000021620184 Neurol Genet
Andreacutea L Benedet Lei Yu Aureacutelie Labbe et al variant mitigates Alzheimer disease pathophysiology in vivo and postmortemCYP2C19
This information is current as of January 30 2018
ServicesUpdated Information amp
httpngneurologyorgcontent41e216fullhtmlincluding high resolution figures can be found at
References httpngneurologyorgcontent41e216fullhtmlref-list-1
This article cites 37 articles 2 of which you can access for free at
Subspecialty Collections
httpngneurologyorgcgicollectionpetPET
httpngneurologyorgcgicollectionassociation_studies_in_geneticsAssociation studies in genetics
httpngneurologyorgcgicollectionalzheimers_diseaseAlzheimers diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpngneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpngneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
reserved Online ISSN 2376-7839Published by Wolters Kluwer Health Inc on behalf of the American Academy of Neurology All rightsan open-access online-only continuous publication journal Copyright Copyright copy 2018 The Author(s)
is an official journal of the American Academy of Neurology Published since April 2015 it isNeurol Genet
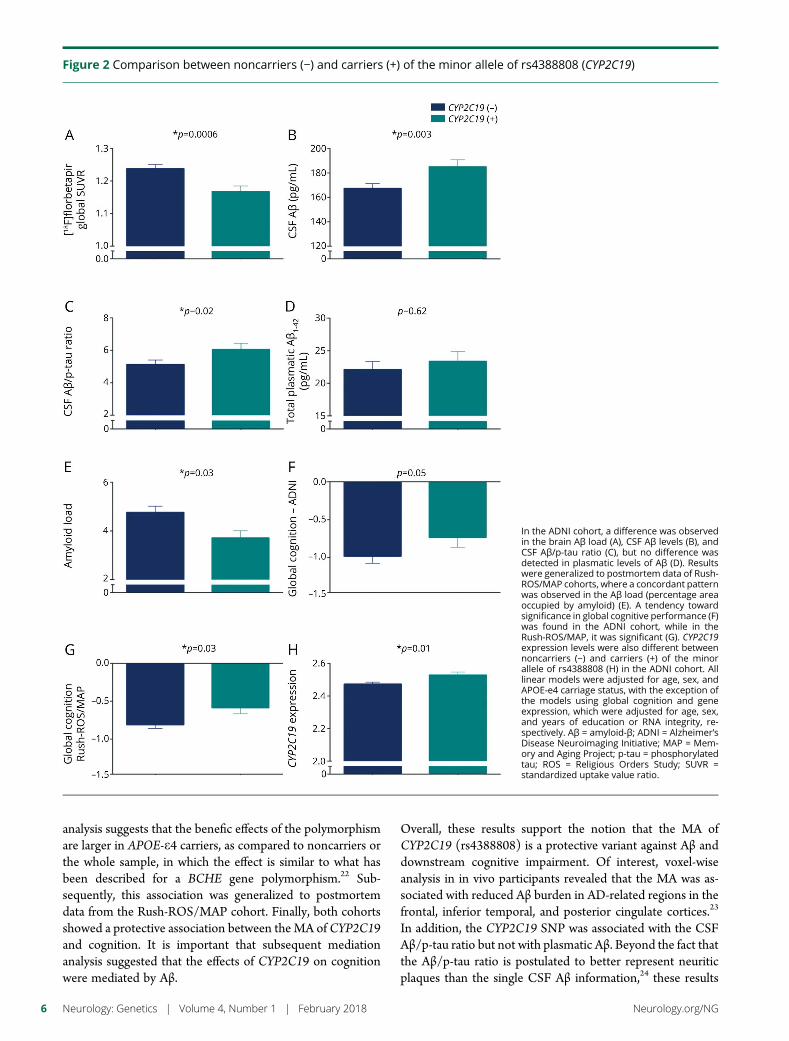
analysis suggests that the benefic effects of the polymorphismare larger in APOE-e4 carriers as compared to noncarriers orthe whole sample in which the effect is similar to what hasbeen described for a BCHE gene polymorphism22 Sub-sequently this association was generalized to postmortemdata from the Rush-ROSMAP cohort Finally both cohortsshowed a protective association between the MA of CYP2C19and cognition It is important that subsequent mediationanalysis suggested that the effects of CYP2C19 on cognitionwere mediated by Aβ
Overall these results support the notion that the MA ofCYP2C19 (rs4388808) is a protective variant against Aβ anddownstream cognitive impairment Of interest voxel-wiseanalysis in in vivo participants revealed that the MA was as-sociated with reduced Aβ burden in AD-related regions in thefrontal inferior temporal and posterior cingulate cortices23
In addition the CYP2C19 SNP was associated with the CSFAβp-tau ratio but not with plasmatic Aβ Beyond the fact thatthe Aβp-tau ratio is postulated to better represent neuriticplaques than the single CSF Aβ information24 these results
Figure 2 Comparison between noncarriers (minus) and carriers (+) of the minor allele of rs4388808 (CYP2C19)
In the ADNI cohort a difference was observedin the brain Aβ load (A) CSF Aβ levels (B) andCSF Aβp-tau ratio (C) but no difference wasdetected in plasmatic levels of Aβ (D) Resultswere generalized to postmortem data of Rush-ROSMAP cohorts where a concordant patternwas observed in the Aβ load (percentage areaoccupied by amyloid) (E) A tendency towardsignificance in global cognitive performance (F)was found in the ADNI cohort while in theRush-ROSMAP it was significant (G) CYP2C19expression levels were also different betweennoncarriers (minus) and carriers (+) of the minorallele of rs4388808 (H) in the ADNI cohort Alllinear models were adjusted for age sex andAPOE-e4 carriage status with the exception ofthe models using global cognition and geneexpression which were adjusted for age sexand years of education or RNA integrity re-spectively Aβ = amyloid-β ADNI = AlzheimerrsquosDisease Neuroimaging Initiative MAP = Mem-ory and Aging Project p-tau = phosphorylatedtau ROS = Religious Orders Study SUVR =standardized uptake value ratio
6 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
provide further evidence that the polymorphism affects thebrain accumulation of neuritic plaques in a tissue- and disease-specific manner24 In both cohorts the referred poly-morphism was not associated with PHF tau or single CSFp-tau agreeing with the framework that Aβ and tau haveindependent upstream triggers25 In line with overall signifi-cant results the CYP2C19 polymorphism also presenteda protective effect in global cognition being supported byprevious studies that linked Aβ pathology to cognitivechanges13 Indeed the mediation analysis supported that theassociation between the genetic factor and cognitive abnor-malities is likely due to the upstream accumulation of Aβ
Levels of the CYP2C19 protein are associated with thers4388808 polymorphism26 which is an intronic variant of theCYP2C19 gene Indeed we found a similar association showingthat MA carriers expressed higher levels of CYP2C19 whencompared with noncarriers suggesting that the SNP affects theexpression of its gene To corroborate this finding we retrievedthe SNPs that were previously excluded because of high LD andverified that none was associated with CYP2C19 expression inthe blood (data not shown) reinforcing the idea that rs4388808is functional This association could not be detected in post-mortem data probably because of the low CYP expression orbecause of its limited detection by the method used7
Despite the SNP not being widely studied to provide in-formation about the association with other phenotypes other
polymorphisms in the gene have been correlated with meta-bolic variability atherosclerosis and behavioral traits27ndash29
CYP2C19 expression has been detected in several brainregions67 however its function in the CNS has not been fullyelucidated Some reports have suggested that besides its roleat metabolizing exogenous substrates CYP2C19 also partic-ipates in other important biological cascades such as themetabolism of serotonin and sexual hormones1 and the me-tabolism of the arachidonic acid (AA) where it functions as anepoxygenase30
The biochemical mechanism underlying the association ofCYP2C19 with Aβ remains speculative A study has shownthat Aβ is able to act as a foreign body and trigger PORactivating CYPs to initiate catabolic reactions9 In turnCYP2C19 is involved in the catalysis of estradiol31 Becauseseveral studies have reported the beneficial effects of estrogenson ADmdashdemonstrating that these steroids are able to act asanti-inflammatory compounds32 and even inhibit Aβproduction33mdashone could think that MA carriers of rs4388808have a reduced expression of this CYP in the brain or that theyhave an increased structural incompatibility betweenCYP2C19 and estradiol Consequently MA carriers couldpresent a reduction in the catabolism of estrogen and a re-duction in the Aβ load Alternatively the Aβ load would beaffected by the levels of epoxyeicosatrienoic acids (EETs)produced via the metabolism of AA by CYP epoxygenases Inthe brain studies have shown that EETs have beneficial effectsat regulating blood flow cortical angiogenesis and at pro-moting anti-inflammatory reactions (for review see reference2) By contrast when AA is metabolized via other cascadesthe protective effect of EETs is lost and there is a possibilityof induction of Aβ production andor accumulation34 Inaddition it was demonstrated that Aβ is able to reduceepoxygenase activity and consequently decrease EET pro-duction35 probably leading to more susceptibility to neuronaldamage (for review see reference 36) In this scenario itwould be possible that the MA of rs4388808 increases theexpression of CYP2C19 leading to an ldquoAβ-resistantrdquo epox-ygenase metabolism of AA with a maintained production ofEETs and a more protective phenotype against amyloidosis
It is also plausible to think however that the associationdescribed here between theCYP2C19 polymorphism and Aβis indirect being mediated by the effect of a drug CYP2C19is known to metabolize several drugs37 including medi-cations to treat depression which has been mentioned asa risk factor for AD (for review see reference 38) Cit-alopram for example is an antidepressant metabolized byCYP2C19 that has been shown to decrease Aβ production39
Thus one may think that the polymorphism rs4388808 isthen associated with an improved action of citalopramleading to the decreased brain Aβ burden rather than as-sociated with Aβ metabolism itself Because it is difficult totrack all the medications the study participants have takenbefore been enrolled it is unfeasible at the moment to checkwhether this association holds true
Figure 3 T-statistical parametric maps showing differencesbetween noncarriers (minus) and carriers (+) of theminor allele of rs4388808 (CYP2C19)
T-statistical parametric maps superimposed on average structural MRI showbrain regions with lower standardized uptake value ratio (SUVR) values inminor allele carriers (CYP2C19 (+)) of the polymorphism of CYP2C19 Statisticaldifferences overlap with brain regions vulnerable to Alzheimer disease path-ophysiology such as the posterior cingulate frontal and temporal cortices
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 7
The interpretation of the results should take into consider-ation a few limitations The use of the 2 cohorts does not allowus to detect and measure Aβ pathology with the samemethodology inserting in the analysis some variability specificto the method used However despite being measured withdifferent techniques both the imaging radiotracer and theantibody used in the brain tissue are expected to bind to thefibrillar form of Aβ13 Similarly the cognitive composite scoresfrom the 2 cohorts have some differences that should beconsidered There are also some intrinsic characteristics of thesamples that have to be taken into account (1) the inclusioncriteria of each study (2) the average age difference betweenthe studies and (3) and the disease stage in which Aβmeasures were performedmdashat the end or during the diseaseprocess Sample size restrictions impose some degree ofcaution when interpreting findings including for examplethe lack of association between the genotype and plasmaticlevels of Aβ or cognition in the ADNI cohort In additionboth cohorts are mostly composed of non-Latino Caucasianslimiting the extrapolation of the present findings to otherpopulation groups Longitudinal analysis together with func-tional genomics and biochemistry experiments not per-formed here would also be necessary to determine the effectof the SNP in the protein function as well as to support anytheoretical framework proposed in the Discussion section
Results obtained from the 2 independent cohorts providecompelling evidences linking the CYP2C19 polymorphismand Aβ pathology suggesting that the MA of rs4388808confers protective effects against Aβ accumulation in the brainand its downstream cognitive consequences These resultscould have implication for antiamyloid clinical trial designs aspreclinical rs4388808G would present a protective factoragainst the amyloid load Therefore the biological mechanismby which the genetic variation would alter Aβ build up andclearance merits further investigation
Author contributionsAndrea L Benedet study concept design analysis and in-terpretation of data composition of figures and drafting themanuscriptLei Yu analysis and interpretation of data and drafting themanuscript Aurelie Labbe supervision of statistical analysisand critical review of the manuscript for intellectual contentSulantha Mathotaarachchi Tharick A Pascoal Monica Shinand Min-Su Kang processing of image data and drafting themanuscript study concept design and drafting the manu-script Serge Gauthier Guy A Rouleau and Judes Poiriercritical review of the manuscript for intellectual content Da-vid A Bennett drafting the manuscript and critical review ofthe manuscript for intellectual content Pedro Rosa-Netostudy concept design study supervision and critical review ofthe manuscript for intellectual content
AcknowledgmentData collection and sharing for this project was funded by theAlzheimerrsquos Disease Neuroimaging Initiative (ADNI) (NIH
grant U01 AG024904) and the DOD ADNI (Department ofDefense award number W81XWH-12-2-0012) The ADNI isfunded by the National Institute on Aging the NationalInstitute of Biomedical Imaging and Bioengineering andthrough generous contributions from the following AbbViethe Alzheimerrsquos Association the Alzheimerrsquos Drug DiscoveryFoundation Araclon Biotech BioClinica Inc BiogenBristol-Myers Squibb CereSpir Inc Cogstate Eisai IncElan Pharmaceuticals Inc Eli Lilly and Company Euro-immun F Hoffmann-La Roche Ltd and its affiliated companyGenentech Inc Fujirebio GE Healthcare IXICO LtdJanssen Alzheimer Immunotherapy Research amp Develop-ment LLC Johnson amp Johnson Pharmaceutical Research ampDevelopment LLC Lumosity Lundbeck Merck amp Co IncMeso Scale Diagnostics LLC NeuroRx Research NeurotrackTechnologies Novartis Pharmaceuticals Corporation PfizerInc Piramal Imaging Servier Takeda Pharmaceutical Com-pany and Transition Therapeutics The Canadian Institutes ofHealth Research is providing funds to support ADNI clinicalsites in Canada Private sector contributions are facilitated bythe Foundation for the NIH (fnihorg) The granteeorganization is the Northern California Institute for Researchand Education and the study is coordinated by the AlzheimerrsquosTherapeutic Research Institute at the University of SouthernCalifornia ADNI data are disseminated by the Laboratory forNeuro Imaging at the University of Southern California
Study fundingThis work was also supported by the Canadian Institutes ofHealth Research (CIHR) (MOP-11-51-31) the AlzheimerrsquosAssociation (NIRG-08-92090) the Canadian Consortium onNeurodegeneration in Aging (CIHR-CCNA) and theWeston Brain Institute Andrea L Benedet is supported bythe CAPES FoundationmdashBrazil (032713-1) Pedro Rosa-Neto is supported by the Fonds de la recherche en sante duQuebec (chercheur boursier) Serge Gauthier and PedroRosa-Neto are members of the CIHR Canadian Consortiumon Neurodegeneration in Aging The Religious Orders Studyand the Rush Memory and Aging Project are funded bythe National Institute on Aging (P30AG10161 andR01AG17917) and the Illinois Department of Public Health
DisclosureAL Benedet has received research support from the CAPESFoundation L Yu has received research support from theNIH A Labbe and S Mathotaarachchi report no disclosuresTA Pascoal has received research support from thePREVENT-AD PhD scholarship M Shin and M-S Kangreport no disclosures S Gauthier has received honoraria forserving on the scientific advisory boards of Alzheon AxovantBoehringer Ingelheim Lilly Lundbeck Novartis SchwabeTauRx on the DSMB of a study sponsored by Eisai and forstudies run by the ADCS and by the ATRI has receivedspeaker honoraria from Schwabe serves on the editorial boardof Alzheimerrsquos amp Dementia has served as a consultant forKalgene and has received research support from the CIHRthe NIH Lundbeck Roche and Lilly GA Rouleau has
8 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
received research support from the CIHR the ALS Society ofCanada and the ALS Association J Poirier serves on theeditorial board of the Journal of Alzheimerrsquos Disease receivespublishing royalties from Trecarre and has received researchsupport from the JL Levesque Foundation DA Bennettserves on the scientific advisory boards of Vigorous MindsTakeda Pharmaceuticals AbbVie EABs (Indian Institute ofScience) DMCs (HRS and MIDUS) and the National Ad-visory Council on Aging is a consultant for Takeda Phar-maceuticals and AbbVie has received research support fromthe NIH and serves on the editorial boards of NeurologyCurrent Alzheimer Research and Neuroepidemiology P Rosa-Neto serves on the scientific advisory boards of EnigmaRadiopharmaceuticals and Kalgene serves on the editorialboard of Frontiers in Aging Neurosciences and has receivedresearch support fromEnigma Radiopharmaceuticals Eli Lillythe CIHR the FRQS Merck (Sharp amp Dohme CorporationMcGill Faculty of Medicine Grant for Translational Re-search) the Weston Brain Institute the Alzheimerrsquos Associa-tion Brain Canada and CQDM Funding information anddisclosures are provided at the end of the article Full disclo-sure form information provided by the authors is availablewith the full text of this article at NeurologyorgNG
Received June 8 2017 Accepted in final form December 4 2017
References1 Miksys S Tyndale RF Cytochrome P450-mediated drug metabolism in the brain
J Psychiatry Neurosci 201338152ndash1632 Iliff JJ Jia J Nelson J Goyagi T Klaus J Alkayed NJ Epoxyeicosanoid signaling in
CNS function and disease Prostaglandins Other Lipid Mediat 20109168ndash843 Stingl JC Brockmoller J Viviani R Genetic variability of drug-metabolizing enzymes
the dual impact on psychiatric therapy and regulation of brain function Mol Psy-chiatry 201318273ndash287
4 McCann SJ Pond SM James KM Le Couteur DG The association between poly-morphisms in the cytochrome P-450 2D6 gene and Parkinsonrsquos disease a case-controlstudy and meta-analysis J Neurol Sci 199715350ndash53
5 Chace C Pang D Weng C et al Variants in CYP17 and CYP19 cytochrome P450genes are associated with onset of Alzheimerrsquos disease in women with down syn-drome JAlzheimer Dis 201228601ndash612
6 McFayden MC Melvin WT Murray GI Regional distribution of individual forms of cyto-chrome P450 mRNA in normal adult human brain Biochem Pharmacol 199855825ndash830
7 Booth Depaz IM Toselli F Wilce PA Gillam EM Differential expression of cyto-chrome P450 enzymes from the CYP2C subfamily in the human brain Drug MetabDispos 201543353ndash357
8 Van Ess PJ Pedersen WA Culmsee C Mattson MP Blouin RA Elevated hepatic anddepressed renal cytochrome P450 activity in the Tg2576 transgenic mouse model ofAlzheimerrsquos disease J Neurochem 200280571ndash578
9 Pappolla MA Omar RA Chyan YJ et al Induction of NADPH cytochrome P450reductase by the Alzheimer beta-protein amyloid as a ldquoforeign bodyrdquo J Neurochem200178121ndash128
10 Liu M Sun A Shin EJ et al Expression of microsomal epoxide hydrolase is elevated inAlzheimerrsquos hippocampus and induced by exogenous beta-amyloid and trimethyl-tinEur J Neurosci 2006232027ndash2034
11 Bennett DA Schneider JA Arvanitakis Z Wilson RS Overview and findings from thereligious orders study Curr Alzheimer Res 20129628ndash645
12 Bennett DA Schneider JA Buchman AS Barnes LL Boyle PA Wilson RS Overviewand findings from the rush Memory and Aging Project Curr Alzheimer Res 20129646ndash663
13 Bennett DA Schneider JA Wilson RS Bienias JL Arnold SE Neurofibrillary tanglesmediate the association of amyloid load with clinical Alzheimer disease and level ofcognitive function Arch Neurol 200461378ndash384
14 Saykin AJ Shen L Yao X et al Genetic studies of quantitative MCI and AD phe-notypes in ADNI Progress opportunities and plans Alzheimers Dement 201511792ndash814
15 Lim AS Srivastava GP Yu L et al 24-hour rhythms of DNA methylation and theirrelation with rhythms of RNA expression in the human dorsolateral prefrontal cortexPLoS Genet 201410e1004792
16 Rogers JF Nafziger AN Bertino JS Jr Pharmacogenetics affects dosing efficacy andtoxicity of cytochrome P450-metabolized drugs Am J Med 2002113746ndash750
17 Anzenbacher P Anzenbacherova E Cytochromes P450 and metabolism of xeno-biotics Cell Mol Life Sci 200158737ndash747
18 Meyer RP Gehlhaus M Knoth R Volk B Expression and function of cytochromep450 in brain drug metabolism Curr Drug Metab 20078297ndash306
19 Purcell S Neale B Todd-Brown K et al PLINK a tool set for whole-genomeassociation and population-based linkage analyses Am J Hum Genet 200781559ndash575
20 R A Language and Environment for Statistical Computing [computer program]Version 310 Vienna R Foundation for Statistical Computing 2015
21 Package RMINC Voxel-Wise Morphometry Using RMINC [computer program]Montreal McGill University 2010
22 Ramanan VK Risacher SL Nho K et al APOE and BCHE as modulators of cerebralamyloid deposition a florbetapir PET genome-wide association study Mol Psychiatry201419351ndash357
23 Braak H Braak E Neuropathological stageing of Alzheimer-related changes ActaNeuropathol 19918220
24 Tapiola T Alafuzoff I Herukka S et al Cerebrospinal fluid β-amyloid 42 and tauproteins as biomarkers of Alzheimer-type pathologic changes in the brain ArchNeurol 200966382ndash389
25 Sperling R Mormino E Johnson K The evolution of preclinical Alzheimerrsquos diseaseimplications for prevention trials Neuron 201484608ndash622
26 Wang D Jiang Z Shen Z et al Functional evaluation of genetic and environmentalregulators of p450 mRNA levels PLoS One 20116e24900
27 Sim SC Risinger C Dahl ML et al A common novel CYP2C19 gene variant causesultrarapid drug metabolism relevant for the drug response to proton pump inhibitorsand antidepressants Clin Pharmacol Ther 200679103ndash113
28 Ishii G Suzuki A Oshino S Shiraishi H Otani K CYP2C19 polymorphism affectspersonality traits of Japanese females Neurosci Lett 200741177ndash80
29 Ercan B Ayaz L Cicek D Tamer L Role of CYP2C9 and CYP2C19 polymorphismsin patients with atherosclerosis Cell Biochem Funct 200826309ndash313
30 Westphal C Konkel A SchunckWH CYP-eicosanoidsmdasha new link between omega-3 fatty acids and cardiac disease Prostaglandins Other Lipid Mediat 20119699ndash108
31 Cheng ZN Shu Y Liu ZQ Wang LS Ou-Yang DS Zhou HH Role of cytochromeP450 in estradiol metabolism in vitro Acta Pharmacol Sin 200122148ndash154
32 Giraud SN Caron CM Pham-Dinh D Kitabgi P Nicot AB Estradiol inhibits ongoingautoimmune neuroinflammation and NFkappaB-dependent CCL2 expression in re-active astrocytes Proc Natl Acad Sci USA 20101078416ndash8421
33 Xu H Wang R Zhang YW Zhang X Estrogen beta-amyloid metabolismtraffickingand Alzheimerrsquos disease Ann NY Acad Sci 20061089324ndash342
34 Amtul Z Uhrig M Wang L Rozmahel RF Beyreuther K Detrimental effects ofarachidonic acid and its metabolites in cellular and mouse models of Alzheimerrsquosdisease structural insight Neurobiol Aging 201233831e21ndashe31
35 Sarkar P Narayanan J Harder DR Differential effect of amyloid beta on the cyto-chrome P450 epoxygenase activity in rat brain Neuroscience 2011194241ndash249
36 Kisler K Nelson AR Montagne A Zlokovic BV Cerebral blood flow regulation andneurovascular dysfunction in Alzheimer disease Nat Rev Neurosci 201718419ndash434
37 Bibi Z Role of cytochrome P450 in drug interactions Nutr Metab (Lond) 200852738 Ownby RL Crocco E Acevedo A John V Loewenstein D Depression and risk for
Alzheimer disease systematic review meta-analysis and metaregression analysisArch Gen Psychiatry 200663530ndash538
39 Sheline YI West T Yarasheski K et al An antidepressant decreases CSF Abetaproduction in healthy individuals and in transgenic AD mice Sci Transl Med 20146236re234
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 9
DOI 101212NXG000000000000021620184 Neurol Genet
Andreacutea L Benedet Lei Yu Aureacutelie Labbe et al variant mitigates Alzheimer disease pathophysiology in vivo and postmortemCYP2C19
This information is current as of January 30 2018
ServicesUpdated Information amp
httpngneurologyorgcontent41e216fullhtmlincluding high resolution figures can be found at
References httpngneurologyorgcontent41e216fullhtmlref-list-1
This article cites 37 articles 2 of which you can access for free at
Subspecialty Collections
httpngneurologyorgcgicollectionpetPET
httpngneurologyorgcgicollectionassociation_studies_in_geneticsAssociation studies in genetics
httpngneurologyorgcgicollectionalzheimers_diseaseAlzheimers diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpngneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpngneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
reserved Online ISSN 2376-7839Published by Wolters Kluwer Health Inc on behalf of the American Academy of Neurology All rightsan open-access online-only continuous publication journal Copyright Copyright copy 2018 The Author(s)
is an official journal of the American Academy of Neurology Published since April 2015 it isNeurol Genet
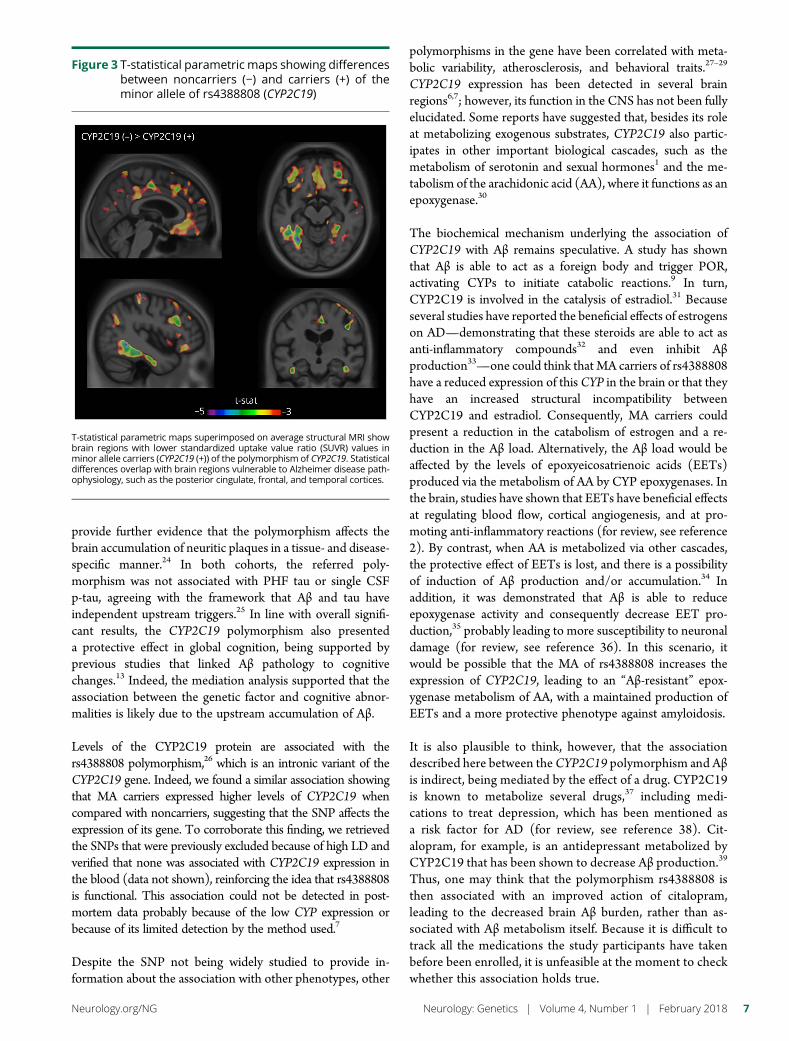
provide further evidence that the polymorphism affects thebrain accumulation of neuritic plaques in a tissue- and disease-specific manner24 In both cohorts the referred poly-morphism was not associated with PHF tau or single CSFp-tau agreeing with the framework that Aβ and tau haveindependent upstream triggers25 In line with overall signifi-cant results the CYP2C19 polymorphism also presenteda protective effect in global cognition being supported byprevious studies that linked Aβ pathology to cognitivechanges13 Indeed the mediation analysis supported that theassociation between the genetic factor and cognitive abnor-malities is likely due to the upstream accumulation of Aβ
Levels of the CYP2C19 protein are associated with thers4388808 polymorphism26 which is an intronic variant of theCYP2C19 gene Indeed we found a similar association showingthat MA carriers expressed higher levels of CYP2C19 whencompared with noncarriers suggesting that the SNP affects theexpression of its gene To corroborate this finding we retrievedthe SNPs that were previously excluded because of high LD andverified that none was associated with CYP2C19 expression inthe blood (data not shown) reinforcing the idea that rs4388808is functional This association could not be detected in post-mortem data probably because of the low CYP expression orbecause of its limited detection by the method used7
Despite the SNP not being widely studied to provide in-formation about the association with other phenotypes other
polymorphisms in the gene have been correlated with meta-bolic variability atherosclerosis and behavioral traits27ndash29
CYP2C19 expression has been detected in several brainregions67 however its function in the CNS has not been fullyelucidated Some reports have suggested that besides its roleat metabolizing exogenous substrates CYP2C19 also partic-ipates in other important biological cascades such as themetabolism of serotonin and sexual hormones1 and the me-tabolism of the arachidonic acid (AA) where it functions as anepoxygenase30
The biochemical mechanism underlying the association ofCYP2C19 with Aβ remains speculative A study has shownthat Aβ is able to act as a foreign body and trigger PORactivating CYPs to initiate catabolic reactions9 In turnCYP2C19 is involved in the catalysis of estradiol31 Becauseseveral studies have reported the beneficial effects of estrogenson ADmdashdemonstrating that these steroids are able to act asanti-inflammatory compounds32 and even inhibit Aβproduction33mdashone could think that MA carriers of rs4388808have a reduced expression of this CYP in the brain or that theyhave an increased structural incompatibility betweenCYP2C19 and estradiol Consequently MA carriers couldpresent a reduction in the catabolism of estrogen and a re-duction in the Aβ load Alternatively the Aβ load would beaffected by the levels of epoxyeicosatrienoic acids (EETs)produced via the metabolism of AA by CYP epoxygenases Inthe brain studies have shown that EETs have beneficial effectsat regulating blood flow cortical angiogenesis and at pro-moting anti-inflammatory reactions (for review see reference2) By contrast when AA is metabolized via other cascadesthe protective effect of EETs is lost and there is a possibilityof induction of Aβ production andor accumulation34 Inaddition it was demonstrated that Aβ is able to reduceepoxygenase activity and consequently decrease EET pro-duction35 probably leading to more susceptibility to neuronaldamage (for review see reference 36) In this scenario itwould be possible that the MA of rs4388808 increases theexpression of CYP2C19 leading to an ldquoAβ-resistantrdquo epox-ygenase metabolism of AA with a maintained production ofEETs and a more protective phenotype against amyloidosis
It is also plausible to think however that the associationdescribed here between theCYP2C19 polymorphism and Aβis indirect being mediated by the effect of a drug CYP2C19is known to metabolize several drugs37 including medi-cations to treat depression which has been mentioned asa risk factor for AD (for review see reference 38) Cit-alopram for example is an antidepressant metabolized byCYP2C19 that has been shown to decrease Aβ production39
Thus one may think that the polymorphism rs4388808 isthen associated with an improved action of citalopramleading to the decreased brain Aβ burden rather than as-sociated with Aβ metabolism itself Because it is difficult totrack all the medications the study participants have takenbefore been enrolled it is unfeasible at the moment to checkwhether this association holds true
Figure 3 T-statistical parametric maps showing differencesbetween noncarriers (minus) and carriers (+) of theminor allele of rs4388808 (CYP2C19)
T-statistical parametric maps superimposed on average structural MRI showbrain regions with lower standardized uptake value ratio (SUVR) values inminor allele carriers (CYP2C19 (+)) of the polymorphism of CYP2C19 Statisticaldifferences overlap with brain regions vulnerable to Alzheimer disease path-ophysiology such as the posterior cingulate frontal and temporal cortices
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 7
The interpretation of the results should take into consider-ation a few limitations The use of the 2 cohorts does not allowus to detect and measure Aβ pathology with the samemethodology inserting in the analysis some variability specificto the method used However despite being measured withdifferent techniques both the imaging radiotracer and theantibody used in the brain tissue are expected to bind to thefibrillar form of Aβ13 Similarly the cognitive composite scoresfrom the 2 cohorts have some differences that should beconsidered There are also some intrinsic characteristics of thesamples that have to be taken into account (1) the inclusioncriteria of each study (2) the average age difference betweenthe studies and (3) and the disease stage in which Aβmeasures were performedmdashat the end or during the diseaseprocess Sample size restrictions impose some degree ofcaution when interpreting findings including for examplethe lack of association between the genotype and plasmaticlevels of Aβ or cognition in the ADNI cohort In additionboth cohorts are mostly composed of non-Latino Caucasianslimiting the extrapolation of the present findings to otherpopulation groups Longitudinal analysis together with func-tional genomics and biochemistry experiments not per-formed here would also be necessary to determine the effectof the SNP in the protein function as well as to support anytheoretical framework proposed in the Discussion section
Results obtained from the 2 independent cohorts providecompelling evidences linking the CYP2C19 polymorphismand Aβ pathology suggesting that the MA of rs4388808confers protective effects against Aβ accumulation in the brainand its downstream cognitive consequences These resultscould have implication for antiamyloid clinical trial designs aspreclinical rs4388808G would present a protective factoragainst the amyloid load Therefore the biological mechanismby which the genetic variation would alter Aβ build up andclearance merits further investigation
Author contributionsAndrea L Benedet study concept design analysis and in-terpretation of data composition of figures and drafting themanuscriptLei Yu analysis and interpretation of data and drafting themanuscript Aurelie Labbe supervision of statistical analysisand critical review of the manuscript for intellectual contentSulantha Mathotaarachchi Tharick A Pascoal Monica Shinand Min-Su Kang processing of image data and drafting themanuscript study concept design and drafting the manu-script Serge Gauthier Guy A Rouleau and Judes Poiriercritical review of the manuscript for intellectual content Da-vid A Bennett drafting the manuscript and critical review ofthe manuscript for intellectual content Pedro Rosa-Netostudy concept design study supervision and critical review ofthe manuscript for intellectual content
AcknowledgmentData collection and sharing for this project was funded by theAlzheimerrsquos Disease Neuroimaging Initiative (ADNI) (NIH
grant U01 AG024904) and the DOD ADNI (Department ofDefense award number W81XWH-12-2-0012) The ADNI isfunded by the National Institute on Aging the NationalInstitute of Biomedical Imaging and Bioengineering andthrough generous contributions from the following AbbViethe Alzheimerrsquos Association the Alzheimerrsquos Drug DiscoveryFoundation Araclon Biotech BioClinica Inc BiogenBristol-Myers Squibb CereSpir Inc Cogstate Eisai IncElan Pharmaceuticals Inc Eli Lilly and Company Euro-immun F Hoffmann-La Roche Ltd and its affiliated companyGenentech Inc Fujirebio GE Healthcare IXICO LtdJanssen Alzheimer Immunotherapy Research amp Develop-ment LLC Johnson amp Johnson Pharmaceutical Research ampDevelopment LLC Lumosity Lundbeck Merck amp Co IncMeso Scale Diagnostics LLC NeuroRx Research NeurotrackTechnologies Novartis Pharmaceuticals Corporation PfizerInc Piramal Imaging Servier Takeda Pharmaceutical Com-pany and Transition Therapeutics The Canadian Institutes ofHealth Research is providing funds to support ADNI clinicalsites in Canada Private sector contributions are facilitated bythe Foundation for the NIH (fnihorg) The granteeorganization is the Northern California Institute for Researchand Education and the study is coordinated by the AlzheimerrsquosTherapeutic Research Institute at the University of SouthernCalifornia ADNI data are disseminated by the Laboratory forNeuro Imaging at the University of Southern California
Study fundingThis work was also supported by the Canadian Institutes ofHealth Research (CIHR) (MOP-11-51-31) the AlzheimerrsquosAssociation (NIRG-08-92090) the Canadian Consortium onNeurodegeneration in Aging (CIHR-CCNA) and theWeston Brain Institute Andrea L Benedet is supported bythe CAPES FoundationmdashBrazil (032713-1) Pedro Rosa-Neto is supported by the Fonds de la recherche en sante duQuebec (chercheur boursier) Serge Gauthier and PedroRosa-Neto are members of the CIHR Canadian Consortiumon Neurodegeneration in Aging The Religious Orders Studyand the Rush Memory and Aging Project are funded bythe National Institute on Aging (P30AG10161 andR01AG17917) and the Illinois Department of Public Health
DisclosureAL Benedet has received research support from the CAPESFoundation L Yu has received research support from theNIH A Labbe and S Mathotaarachchi report no disclosuresTA Pascoal has received research support from thePREVENT-AD PhD scholarship M Shin and M-S Kangreport no disclosures S Gauthier has received honoraria forserving on the scientific advisory boards of Alzheon AxovantBoehringer Ingelheim Lilly Lundbeck Novartis SchwabeTauRx on the DSMB of a study sponsored by Eisai and forstudies run by the ADCS and by the ATRI has receivedspeaker honoraria from Schwabe serves on the editorial boardof Alzheimerrsquos amp Dementia has served as a consultant forKalgene and has received research support from the CIHRthe NIH Lundbeck Roche and Lilly GA Rouleau has
8 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
received research support from the CIHR the ALS Society ofCanada and the ALS Association J Poirier serves on theeditorial board of the Journal of Alzheimerrsquos Disease receivespublishing royalties from Trecarre and has received researchsupport from the JL Levesque Foundation DA Bennettserves on the scientific advisory boards of Vigorous MindsTakeda Pharmaceuticals AbbVie EABs (Indian Institute ofScience) DMCs (HRS and MIDUS) and the National Ad-visory Council on Aging is a consultant for Takeda Phar-maceuticals and AbbVie has received research support fromthe NIH and serves on the editorial boards of NeurologyCurrent Alzheimer Research and Neuroepidemiology P Rosa-Neto serves on the scientific advisory boards of EnigmaRadiopharmaceuticals and Kalgene serves on the editorialboard of Frontiers in Aging Neurosciences and has receivedresearch support fromEnigma Radiopharmaceuticals Eli Lillythe CIHR the FRQS Merck (Sharp amp Dohme CorporationMcGill Faculty of Medicine Grant for Translational Re-search) the Weston Brain Institute the Alzheimerrsquos Associa-tion Brain Canada and CQDM Funding information anddisclosures are provided at the end of the article Full disclo-sure form information provided by the authors is availablewith the full text of this article at NeurologyorgNG
Received June 8 2017 Accepted in final form December 4 2017
References1 Miksys S Tyndale RF Cytochrome P450-mediated drug metabolism in the brain
J Psychiatry Neurosci 201338152ndash1632 Iliff JJ Jia J Nelson J Goyagi T Klaus J Alkayed NJ Epoxyeicosanoid signaling in
CNS function and disease Prostaglandins Other Lipid Mediat 20109168ndash843 Stingl JC Brockmoller J Viviani R Genetic variability of drug-metabolizing enzymes
the dual impact on psychiatric therapy and regulation of brain function Mol Psy-chiatry 201318273ndash287
4 McCann SJ Pond SM James KM Le Couteur DG The association between poly-morphisms in the cytochrome P-450 2D6 gene and Parkinsonrsquos disease a case-controlstudy and meta-analysis J Neurol Sci 199715350ndash53
5 Chace C Pang D Weng C et al Variants in CYP17 and CYP19 cytochrome P450genes are associated with onset of Alzheimerrsquos disease in women with down syn-drome JAlzheimer Dis 201228601ndash612
6 McFayden MC Melvin WT Murray GI Regional distribution of individual forms of cyto-chrome P450 mRNA in normal adult human brain Biochem Pharmacol 199855825ndash830
7 Booth Depaz IM Toselli F Wilce PA Gillam EM Differential expression of cyto-chrome P450 enzymes from the CYP2C subfamily in the human brain Drug MetabDispos 201543353ndash357
8 Van Ess PJ Pedersen WA Culmsee C Mattson MP Blouin RA Elevated hepatic anddepressed renal cytochrome P450 activity in the Tg2576 transgenic mouse model ofAlzheimerrsquos disease J Neurochem 200280571ndash578
9 Pappolla MA Omar RA Chyan YJ et al Induction of NADPH cytochrome P450reductase by the Alzheimer beta-protein amyloid as a ldquoforeign bodyrdquo J Neurochem200178121ndash128
10 Liu M Sun A Shin EJ et al Expression of microsomal epoxide hydrolase is elevated inAlzheimerrsquos hippocampus and induced by exogenous beta-amyloid and trimethyl-tinEur J Neurosci 2006232027ndash2034
11 Bennett DA Schneider JA Arvanitakis Z Wilson RS Overview and findings from thereligious orders study Curr Alzheimer Res 20129628ndash645
12 Bennett DA Schneider JA Buchman AS Barnes LL Boyle PA Wilson RS Overviewand findings from the rush Memory and Aging Project Curr Alzheimer Res 20129646ndash663
13 Bennett DA Schneider JA Wilson RS Bienias JL Arnold SE Neurofibrillary tanglesmediate the association of amyloid load with clinical Alzheimer disease and level ofcognitive function Arch Neurol 200461378ndash384
14 Saykin AJ Shen L Yao X et al Genetic studies of quantitative MCI and AD phe-notypes in ADNI Progress opportunities and plans Alzheimers Dement 201511792ndash814
15 Lim AS Srivastava GP Yu L et al 24-hour rhythms of DNA methylation and theirrelation with rhythms of RNA expression in the human dorsolateral prefrontal cortexPLoS Genet 201410e1004792
16 Rogers JF Nafziger AN Bertino JS Jr Pharmacogenetics affects dosing efficacy andtoxicity of cytochrome P450-metabolized drugs Am J Med 2002113746ndash750
17 Anzenbacher P Anzenbacherova E Cytochromes P450 and metabolism of xeno-biotics Cell Mol Life Sci 200158737ndash747
18 Meyer RP Gehlhaus M Knoth R Volk B Expression and function of cytochromep450 in brain drug metabolism Curr Drug Metab 20078297ndash306
19 Purcell S Neale B Todd-Brown K et al PLINK a tool set for whole-genomeassociation and population-based linkage analyses Am J Hum Genet 200781559ndash575
20 R A Language and Environment for Statistical Computing [computer program]Version 310 Vienna R Foundation for Statistical Computing 2015
21 Package RMINC Voxel-Wise Morphometry Using RMINC [computer program]Montreal McGill University 2010
22 Ramanan VK Risacher SL Nho K et al APOE and BCHE as modulators of cerebralamyloid deposition a florbetapir PET genome-wide association study Mol Psychiatry201419351ndash357
23 Braak H Braak E Neuropathological stageing of Alzheimer-related changes ActaNeuropathol 19918220
24 Tapiola T Alafuzoff I Herukka S et al Cerebrospinal fluid β-amyloid 42 and tauproteins as biomarkers of Alzheimer-type pathologic changes in the brain ArchNeurol 200966382ndash389
25 Sperling R Mormino E Johnson K The evolution of preclinical Alzheimerrsquos diseaseimplications for prevention trials Neuron 201484608ndash622
26 Wang D Jiang Z Shen Z et al Functional evaluation of genetic and environmentalregulators of p450 mRNA levels PLoS One 20116e24900
27 Sim SC Risinger C Dahl ML et al A common novel CYP2C19 gene variant causesultrarapid drug metabolism relevant for the drug response to proton pump inhibitorsand antidepressants Clin Pharmacol Ther 200679103ndash113
28 Ishii G Suzuki A Oshino S Shiraishi H Otani K CYP2C19 polymorphism affectspersonality traits of Japanese females Neurosci Lett 200741177ndash80
29 Ercan B Ayaz L Cicek D Tamer L Role of CYP2C9 and CYP2C19 polymorphismsin patients with atherosclerosis Cell Biochem Funct 200826309ndash313
30 Westphal C Konkel A SchunckWH CYP-eicosanoidsmdasha new link between omega-3 fatty acids and cardiac disease Prostaglandins Other Lipid Mediat 20119699ndash108
31 Cheng ZN Shu Y Liu ZQ Wang LS Ou-Yang DS Zhou HH Role of cytochromeP450 in estradiol metabolism in vitro Acta Pharmacol Sin 200122148ndash154
32 Giraud SN Caron CM Pham-Dinh D Kitabgi P Nicot AB Estradiol inhibits ongoingautoimmune neuroinflammation and NFkappaB-dependent CCL2 expression in re-active astrocytes Proc Natl Acad Sci USA 20101078416ndash8421
33 Xu H Wang R Zhang YW Zhang X Estrogen beta-amyloid metabolismtraffickingand Alzheimerrsquos disease Ann NY Acad Sci 20061089324ndash342
34 Amtul Z Uhrig M Wang L Rozmahel RF Beyreuther K Detrimental effects ofarachidonic acid and its metabolites in cellular and mouse models of Alzheimerrsquosdisease structural insight Neurobiol Aging 201233831e21ndashe31
35 Sarkar P Narayanan J Harder DR Differential effect of amyloid beta on the cyto-chrome P450 epoxygenase activity in rat brain Neuroscience 2011194241ndash249
36 Kisler K Nelson AR Montagne A Zlokovic BV Cerebral blood flow regulation andneurovascular dysfunction in Alzheimer disease Nat Rev Neurosci 201718419ndash434
37 Bibi Z Role of cytochrome P450 in drug interactions Nutr Metab (Lond) 200852738 Ownby RL Crocco E Acevedo A John V Loewenstein D Depression and risk for
Alzheimer disease systematic review meta-analysis and metaregression analysisArch Gen Psychiatry 200663530ndash538
39 Sheline YI West T Yarasheski K et al An antidepressant decreases CSF Abetaproduction in healthy individuals and in transgenic AD mice Sci Transl Med 20146236re234
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 9
DOI 101212NXG000000000000021620184 Neurol Genet
Andreacutea L Benedet Lei Yu Aureacutelie Labbe et al variant mitigates Alzheimer disease pathophysiology in vivo and postmortemCYP2C19
This information is current as of January 30 2018
ServicesUpdated Information amp
httpngneurologyorgcontent41e216fullhtmlincluding high resolution figures can be found at
References httpngneurologyorgcontent41e216fullhtmlref-list-1
This article cites 37 articles 2 of which you can access for free at
Subspecialty Collections
httpngneurologyorgcgicollectionpetPET
httpngneurologyorgcgicollectionassociation_studies_in_geneticsAssociation studies in genetics
httpngneurologyorgcgicollectionalzheimers_diseaseAlzheimers diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpngneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpngneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
reserved Online ISSN 2376-7839Published by Wolters Kluwer Health Inc on behalf of the American Academy of Neurology All rightsan open-access online-only continuous publication journal Copyright Copyright copy 2018 The Author(s)
is an official journal of the American Academy of Neurology Published since April 2015 it isNeurol Genet

The interpretation of the results should take into consider-ation a few limitations The use of the 2 cohorts does not allowus to detect and measure Aβ pathology with the samemethodology inserting in the analysis some variability specificto the method used However despite being measured withdifferent techniques both the imaging radiotracer and theantibody used in the brain tissue are expected to bind to thefibrillar form of Aβ13 Similarly the cognitive composite scoresfrom the 2 cohorts have some differences that should beconsidered There are also some intrinsic characteristics of thesamples that have to be taken into account (1) the inclusioncriteria of each study (2) the average age difference betweenthe studies and (3) and the disease stage in which Aβmeasures were performedmdashat the end or during the diseaseprocess Sample size restrictions impose some degree ofcaution when interpreting findings including for examplethe lack of association between the genotype and plasmaticlevels of Aβ or cognition in the ADNI cohort In additionboth cohorts are mostly composed of non-Latino Caucasianslimiting the extrapolation of the present findings to otherpopulation groups Longitudinal analysis together with func-tional genomics and biochemistry experiments not per-formed here would also be necessary to determine the effectof the SNP in the protein function as well as to support anytheoretical framework proposed in the Discussion section
Results obtained from the 2 independent cohorts providecompelling evidences linking the CYP2C19 polymorphismand Aβ pathology suggesting that the MA of rs4388808confers protective effects against Aβ accumulation in the brainand its downstream cognitive consequences These resultscould have implication for antiamyloid clinical trial designs aspreclinical rs4388808G would present a protective factoragainst the amyloid load Therefore the biological mechanismby which the genetic variation would alter Aβ build up andclearance merits further investigation
Author contributionsAndrea L Benedet study concept design analysis and in-terpretation of data composition of figures and drafting themanuscriptLei Yu analysis and interpretation of data and drafting themanuscript Aurelie Labbe supervision of statistical analysisand critical review of the manuscript for intellectual contentSulantha Mathotaarachchi Tharick A Pascoal Monica Shinand Min-Su Kang processing of image data and drafting themanuscript study concept design and drafting the manu-script Serge Gauthier Guy A Rouleau and Judes Poiriercritical review of the manuscript for intellectual content Da-vid A Bennett drafting the manuscript and critical review ofthe manuscript for intellectual content Pedro Rosa-Netostudy concept design study supervision and critical review ofthe manuscript for intellectual content
AcknowledgmentData collection and sharing for this project was funded by theAlzheimerrsquos Disease Neuroimaging Initiative (ADNI) (NIH
grant U01 AG024904) and the DOD ADNI (Department ofDefense award number W81XWH-12-2-0012) The ADNI isfunded by the National Institute on Aging the NationalInstitute of Biomedical Imaging and Bioengineering andthrough generous contributions from the following AbbViethe Alzheimerrsquos Association the Alzheimerrsquos Drug DiscoveryFoundation Araclon Biotech BioClinica Inc BiogenBristol-Myers Squibb CereSpir Inc Cogstate Eisai IncElan Pharmaceuticals Inc Eli Lilly and Company Euro-immun F Hoffmann-La Roche Ltd and its affiliated companyGenentech Inc Fujirebio GE Healthcare IXICO LtdJanssen Alzheimer Immunotherapy Research amp Develop-ment LLC Johnson amp Johnson Pharmaceutical Research ampDevelopment LLC Lumosity Lundbeck Merck amp Co IncMeso Scale Diagnostics LLC NeuroRx Research NeurotrackTechnologies Novartis Pharmaceuticals Corporation PfizerInc Piramal Imaging Servier Takeda Pharmaceutical Com-pany and Transition Therapeutics The Canadian Institutes ofHealth Research is providing funds to support ADNI clinicalsites in Canada Private sector contributions are facilitated bythe Foundation for the NIH (fnihorg) The granteeorganization is the Northern California Institute for Researchand Education and the study is coordinated by the AlzheimerrsquosTherapeutic Research Institute at the University of SouthernCalifornia ADNI data are disseminated by the Laboratory forNeuro Imaging at the University of Southern California
Study fundingThis work was also supported by the Canadian Institutes ofHealth Research (CIHR) (MOP-11-51-31) the AlzheimerrsquosAssociation (NIRG-08-92090) the Canadian Consortium onNeurodegeneration in Aging (CIHR-CCNA) and theWeston Brain Institute Andrea L Benedet is supported bythe CAPES FoundationmdashBrazil (032713-1) Pedro Rosa-Neto is supported by the Fonds de la recherche en sante duQuebec (chercheur boursier) Serge Gauthier and PedroRosa-Neto are members of the CIHR Canadian Consortiumon Neurodegeneration in Aging The Religious Orders Studyand the Rush Memory and Aging Project are funded bythe National Institute on Aging (P30AG10161 andR01AG17917) and the Illinois Department of Public Health
DisclosureAL Benedet has received research support from the CAPESFoundation L Yu has received research support from theNIH A Labbe and S Mathotaarachchi report no disclosuresTA Pascoal has received research support from thePREVENT-AD PhD scholarship M Shin and M-S Kangreport no disclosures S Gauthier has received honoraria forserving on the scientific advisory boards of Alzheon AxovantBoehringer Ingelheim Lilly Lundbeck Novartis SchwabeTauRx on the DSMB of a study sponsored by Eisai and forstudies run by the ADCS and by the ATRI has receivedspeaker honoraria from Schwabe serves on the editorial boardof Alzheimerrsquos amp Dementia has served as a consultant forKalgene and has received research support from the CIHRthe NIH Lundbeck Roche and Lilly GA Rouleau has
8 Neurology Genetics | Volume 4 Number 1 | February 2018 NeurologyorgNG
received research support from the CIHR the ALS Society ofCanada and the ALS Association J Poirier serves on theeditorial board of the Journal of Alzheimerrsquos Disease receivespublishing royalties from Trecarre and has received researchsupport from the JL Levesque Foundation DA Bennettserves on the scientific advisory boards of Vigorous MindsTakeda Pharmaceuticals AbbVie EABs (Indian Institute ofScience) DMCs (HRS and MIDUS) and the National Ad-visory Council on Aging is a consultant for Takeda Phar-maceuticals and AbbVie has received research support fromthe NIH and serves on the editorial boards of NeurologyCurrent Alzheimer Research and Neuroepidemiology P Rosa-Neto serves on the scientific advisory boards of EnigmaRadiopharmaceuticals and Kalgene serves on the editorialboard of Frontiers in Aging Neurosciences and has receivedresearch support fromEnigma Radiopharmaceuticals Eli Lillythe CIHR the FRQS Merck (Sharp amp Dohme CorporationMcGill Faculty of Medicine Grant for Translational Re-search) the Weston Brain Institute the Alzheimerrsquos Associa-tion Brain Canada and CQDM Funding information anddisclosures are provided at the end of the article Full disclo-sure form information provided by the authors is availablewith the full text of this article at NeurologyorgNG
Received June 8 2017 Accepted in final form December 4 2017
References1 Miksys S Tyndale RF Cytochrome P450-mediated drug metabolism in the brain
J Psychiatry Neurosci 201338152ndash1632 Iliff JJ Jia J Nelson J Goyagi T Klaus J Alkayed NJ Epoxyeicosanoid signaling in
CNS function and disease Prostaglandins Other Lipid Mediat 20109168ndash843 Stingl JC Brockmoller J Viviani R Genetic variability of drug-metabolizing enzymes
the dual impact on psychiatric therapy and regulation of brain function Mol Psy-chiatry 201318273ndash287
4 McCann SJ Pond SM James KM Le Couteur DG The association between poly-morphisms in the cytochrome P-450 2D6 gene and Parkinsonrsquos disease a case-controlstudy and meta-analysis J Neurol Sci 199715350ndash53
5 Chace C Pang D Weng C et al Variants in CYP17 and CYP19 cytochrome P450genes are associated with onset of Alzheimerrsquos disease in women with down syn-drome JAlzheimer Dis 201228601ndash612
6 McFayden MC Melvin WT Murray GI Regional distribution of individual forms of cyto-chrome P450 mRNA in normal adult human brain Biochem Pharmacol 199855825ndash830
7 Booth Depaz IM Toselli F Wilce PA Gillam EM Differential expression of cyto-chrome P450 enzymes from the CYP2C subfamily in the human brain Drug MetabDispos 201543353ndash357
8 Van Ess PJ Pedersen WA Culmsee C Mattson MP Blouin RA Elevated hepatic anddepressed renal cytochrome P450 activity in the Tg2576 transgenic mouse model ofAlzheimerrsquos disease J Neurochem 200280571ndash578
9 Pappolla MA Omar RA Chyan YJ et al Induction of NADPH cytochrome P450reductase by the Alzheimer beta-protein amyloid as a ldquoforeign bodyrdquo J Neurochem200178121ndash128
10 Liu M Sun A Shin EJ et al Expression of microsomal epoxide hydrolase is elevated inAlzheimerrsquos hippocampus and induced by exogenous beta-amyloid and trimethyl-tinEur J Neurosci 2006232027ndash2034
11 Bennett DA Schneider JA Arvanitakis Z Wilson RS Overview and findings from thereligious orders study Curr Alzheimer Res 20129628ndash645
12 Bennett DA Schneider JA Buchman AS Barnes LL Boyle PA Wilson RS Overviewand findings from the rush Memory and Aging Project Curr Alzheimer Res 20129646ndash663
13 Bennett DA Schneider JA Wilson RS Bienias JL Arnold SE Neurofibrillary tanglesmediate the association of amyloid load with clinical Alzheimer disease and level ofcognitive function Arch Neurol 200461378ndash384
14 Saykin AJ Shen L Yao X et al Genetic studies of quantitative MCI and AD phe-notypes in ADNI Progress opportunities and plans Alzheimers Dement 201511792ndash814
15 Lim AS Srivastava GP Yu L et al 24-hour rhythms of DNA methylation and theirrelation with rhythms of RNA expression in the human dorsolateral prefrontal cortexPLoS Genet 201410e1004792
16 Rogers JF Nafziger AN Bertino JS Jr Pharmacogenetics affects dosing efficacy andtoxicity of cytochrome P450-metabolized drugs Am J Med 2002113746ndash750
17 Anzenbacher P Anzenbacherova E Cytochromes P450 and metabolism of xeno-biotics Cell Mol Life Sci 200158737ndash747
18 Meyer RP Gehlhaus M Knoth R Volk B Expression and function of cytochromep450 in brain drug metabolism Curr Drug Metab 20078297ndash306
19 Purcell S Neale B Todd-Brown K et al PLINK a tool set for whole-genomeassociation and population-based linkage analyses Am J Hum Genet 200781559ndash575
20 R A Language and Environment for Statistical Computing [computer program]Version 310 Vienna R Foundation for Statistical Computing 2015
21 Package RMINC Voxel-Wise Morphometry Using RMINC [computer program]Montreal McGill University 2010
22 Ramanan VK Risacher SL Nho K et al APOE and BCHE as modulators of cerebralamyloid deposition a florbetapir PET genome-wide association study Mol Psychiatry201419351ndash357
23 Braak H Braak E Neuropathological stageing of Alzheimer-related changes ActaNeuropathol 19918220
24 Tapiola T Alafuzoff I Herukka S et al Cerebrospinal fluid β-amyloid 42 and tauproteins as biomarkers of Alzheimer-type pathologic changes in the brain ArchNeurol 200966382ndash389
25 Sperling R Mormino E Johnson K The evolution of preclinical Alzheimerrsquos diseaseimplications for prevention trials Neuron 201484608ndash622
26 Wang D Jiang Z Shen Z et al Functional evaluation of genetic and environmentalregulators of p450 mRNA levels PLoS One 20116e24900
27 Sim SC Risinger C Dahl ML et al A common novel CYP2C19 gene variant causesultrarapid drug metabolism relevant for the drug response to proton pump inhibitorsand antidepressants Clin Pharmacol Ther 200679103ndash113
28 Ishii G Suzuki A Oshino S Shiraishi H Otani K CYP2C19 polymorphism affectspersonality traits of Japanese females Neurosci Lett 200741177ndash80
29 Ercan B Ayaz L Cicek D Tamer L Role of CYP2C9 and CYP2C19 polymorphismsin patients with atherosclerosis Cell Biochem Funct 200826309ndash313
30 Westphal C Konkel A SchunckWH CYP-eicosanoidsmdasha new link between omega-3 fatty acids and cardiac disease Prostaglandins Other Lipid Mediat 20119699ndash108
31 Cheng ZN Shu Y Liu ZQ Wang LS Ou-Yang DS Zhou HH Role of cytochromeP450 in estradiol metabolism in vitro Acta Pharmacol Sin 200122148ndash154
32 Giraud SN Caron CM Pham-Dinh D Kitabgi P Nicot AB Estradiol inhibits ongoingautoimmune neuroinflammation and NFkappaB-dependent CCL2 expression in re-active astrocytes Proc Natl Acad Sci USA 20101078416ndash8421
33 Xu H Wang R Zhang YW Zhang X Estrogen beta-amyloid metabolismtraffickingand Alzheimerrsquos disease Ann NY Acad Sci 20061089324ndash342
34 Amtul Z Uhrig M Wang L Rozmahel RF Beyreuther K Detrimental effects ofarachidonic acid and its metabolites in cellular and mouse models of Alzheimerrsquosdisease structural insight Neurobiol Aging 201233831e21ndashe31
35 Sarkar P Narayanan J Harder DR Differential effect of amyloid beta on the cyto-chrome P450 epoxygenase activity in rat brain Neuroscience 2011194241ndash249
36 Kisler K Nelson AR Montagne A Zlokovic BV Cerebral blood flow regulation andneurovascular dysfunction in Alzheimer disease Nat Rev Neurosci 201718419ndash434
37 Bibi Z Role of cytochrome P450 in drug interactions Nutr Metab (Lond) 200852738 Ownby RL Crocco E Acevedo A John V Loewenstein D Depression and risk for
Alzheimer disease systematic review meta-analysis and metaregression analysisArch Gen Psychiatry 200663530ndash538
39 Sheline YI West T Yarasheski K et al An antidepressant decreases CSF Abetaproduction in healthy individuals and in transgenic AD mice Sci Transl Med 20146236re234
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 9
DOI 101212NXG000000000000021620184 Neurol Genet
Andreacutea L Benedet Lei Yu Aureacutelie Labbe et al variant mitigates Alzheimer disease pathophysiology in vivo and postmortemCYP2C19
This information is current as of January 30 2018
ServicesUpdated Information amp
httpngneurologyorgcontent41e216fullhtmlincluding high resolution figures can be found at
References httpngneurologyorgcontent41e216fullhtmlref-list-1
This article cites 37 articles 2 of which you can access for free at
Subspecialty Collections
httpngneurologyorgcgicollectionpetPET
httpngneurologyorgcgicollectionassociation_studies_in_geneticsAssociation studies in genetics
httpngneurologyorgcgicollectionalzheimers_diseaseAlzheimers diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpngneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpngneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
reserved Online ISSN 2376-7839Published by Wolters Kluwer Health Inc on behalf of the American Academy of Neurology All rightsan open-access online-only continuous publication journal Copyright Copyright copy 2018 The Author(s)
is an official journal of the American Academy of Neurology Published since April 2015 it isNeurol Genet

received research support from the CIHR the ALS Society ofCanada and the ALS Association J Poirier serves on theeditorial board of the Journal of Alzheimerrsquos Disease receivespublishing royalties from Trecarre and has received researchsupport from the JL Levesque Foundation DA Bennettserves on the scientific advisory boards of Vigorous MindsTakeda Pharmaceuticals AbbVie EABs (Indian Institute ofScience) DMCs (HRS and MIDUS) and the National Ad-visory Council on Aging is a consultant for Takeda Phar-maceuticals and AbbVie has received research support fromthe NIH and serves on the editorial boards of NeurologyCurrent Alzheimer Research and Neuroepidemiology P Rosa-Neto serves on the scientific advisory boards of EnigmaRadiopharmaceuticals and Kalgene serves on the editorialboard of Frontiers in Aging Neurosciences and has receivedresearch support fromEnigma Radiopharmaceuticals Eli Lillythe CIHR the FRQS Merck (Sharp amp Dohme CorporationMcGill Faculty of Medicine Grant for Translational Re-search) the Weston Brain Institute the Alzheimerrsquos Associa-tion Brain Canada and CQDM Funding information anddisclosures are provided at the end of the article Full disclo-sure form information provided by the authors is availablewith the full text of this article at NeurologyorgNG
Received June 8 2017 Accepted in final form December 4 2017
References1 Miksys S Tyndale RF Cytochrome P450-mediated drug metabolism in the brain
J Psychiatry Neurosci 201338152ndash1632 Iliff JJ Jia J Nelson J Goyagi T Klaus J Alkayed NJ Epoxyeicosanoid signaling in
CNS function and disease Prostaglandins Other Lipid Mediat 20109168ndash843 Stingl JC Brockmoller J Viviani R Genetic variability of drug-metabolizing enzymes
the dual impact on psychiatric therapy and regulation of brain function Mol Psy-chiatry 201318273ndash287
4 McCann SJ Pond SM James KM Le Couteur DG The association between poly-morphisms in the cytochrome P-450 2D6 gene and Parkinsonrsquos disease a case-controlstudy and meta-analysis J Neurol Sci 199715350ndash53
5 Chace C Pang D Weng C et al Variants in CYP17 and CYP19 cytochrome P450genes are associated with onset of Alzheimerrsquos disease in women with down syn-drome JAlzheimer Dis 201228601ndash612
6 McFayden MC Melvin WT Murray GI Regional distribution of individual forms of cyto-chrome P450 mRNA in normal adult human brain Biochem Pharmacol 199855825ndash830
7 Booth Depaz IM Toselli F Wilce PA Gillam EM Differential expression of cyto-chrome P450 enzymes from the CYP2C subfamily in the human brain Drug MetabDispos 201543353ndash357
8 Van Ess PJ Pedersen WA Culmsee C Mattson MP Blouin RA Elevated hepatic anddepressed renal cytochrome P450 activity in the Tg2576 transgenic mouse model ofAlzheimerrsquos disease J Neurochem 200280571ndash578
9 Pappolla MA Omar RA Chyan YJ et al Induction of NADPH cytochrome P450reductase by the Alzheimer beta-protein amyloid as a ldquoforeign bodyrdquo J Neurochem200178121ndash128
10 Liu M Sun A Shin EJ et al Expression of microsomal epoxide hydrolase is elevated inAlzheimerrsquos hippocampus and induced by exogenous beta-amyloid and trimethyl-tinEur J Neurosci 2006232027ndash2034
11 Bennett DA Schneider JA Arvanitakis Z Wilson RS Overview and findings from thereligious orders study Curr Alzheimer Res 20129628ndash645
12 Bennett DA Schneider JA Buchman AS Barnes LL Boyle PA Wilson RS Overviewand findings from the rush Memory and Aging Project Curr Alzheimer Res 20129646ndash663
13 Bennett DA Schneider JA Wilson RS Bienias JL Arnold SE Neurofibrillary tanglesmediate the association of amyloid load with clinical Alzheimer disease and level ofcognitive function Arch Neurol 200461378ndash384
14 Saykin AJ Shen L Yao X et al Genetic studies of quantitative MCI and AD phe-notypes in ADNI Progress opportunities and plans Alzheimers Dement 201511792ndash814
15 Lim AS Srivastava GP Yu L et al 24-hour rhythms of DNA methylation and theirrelation with rhythms of RNA expression in the human dorsolateral prefrontal cortexPLoS Genet 201410e1004792
16 Rogers JF Nafziger AN Bertino JS Jr Pharmacogenetics affects dosing efficacy andtoxicity of cytochrome P450-metabolized drugs Am J Med 2002113746ndash750
17 Anzenbacher P Anzenbacherova E Cytochromes P450 and metabolism of xeno-biotics Cell Mol Life Sci 200158737ndash747
18 Meyer RP Gehlhaus M Knoth R Volk B Expression and function of cytochromep450 in brain drug metabolism Curr Drug Metab 20078297ndash306
19 Purcell S Neale B Todd-Brown K et al PLINK a tool set for whole-genomeassociation and population-based linkage analyses Am J Hum Genet 200781559ndash575
20 R A Language and Environment for Statistical Computing [computer program]Version 310 Vienna R Foundation for Statistical Computing 2015
21 Package RMINC Voxel-Wise Morphometry Using RMINC [computer program]Montreal McGill University 2010
22 Ramanan VK Risacher SL Nho K et al APOE and BCHE as modulators of cerebralamyloid deposition a florbetapir PET genome-wide association study Mol Psychiatry201419351ndash357
23 Braak H Braak E Neuropathological stageing of Alzheimer-related changes ActaNeuropathol 19918220
24 Tapiola T Alafuzoff I Herukka S et al Cerebrospinal fluid β-amyloid 42 and tauproteins as biomarkers of Alzheimer-type pathologic changes in the brain ArchNeurol 200966382ndash389
25 Sperling R Mormino E Johnson K The evolution of preclinical Alzheimerrsquos diseaseimplications for prevention trials Neuron 201484608ndash622
26 Wang D Jiang Z Shen Z et al Functional evaluation of genetic and environmentalregulators of p450 mRNA levels PLoS One 20116e24900
27 Sim SC Risinger C Dahl ML et al A common novel CYP2C19 gene variant causesultrarapid drug metabolism relevant for the drug response to proton pump inhibitorsand antidepressants Clin Pharmacol Ther 200679103ndash113
28 Ishii G Suzuki A Oshino S Shiraishi H Otani K CYP2C19 polymorphism affectspersonality traits of Japanese females Neurosci Lett 200741177ndash80
29 Ercan B Ayaz L Cicek D Tamer L Role of CYP2C9 and CYP2C19 polymorphismsin patients with atherosclerosis Cell Biochem Funct 200826309ndash313
30 Westphal C Konkel A SchunckWH CYP-eicosanoidsmdasha new link between omega-3 fatty acids and cardiac disease Prostaglandins Other Lipid Mediat 20119699ndash108
31 Cheng ZN Shu Y Liu ZQ Wang LS Ou-Yang DS Zhou HH Role of cytochromeP450 in estradiol metabolism in vitro Acta Pharmacol Sin 200122148ndash154
32 Giraud SN Caron CM Pham-Dinh D Kitabgi P Nicot AB Estradiol inhibits ongoingautoimmune neuroinflammation and NFkappaB-dependent CCL2 expression in re-active astrocytes Proc Natl Acad Sci USA 20101078416ndash8421
33 Xu H Wang R Zhang YW Zhang X Estrogen beta-amyloid metabolismtraffickingand Alzheimerrsquos disease Ann NY Acad Sci 20061089324ndash342
34 Amtul Z Uhrig M Wang L Rozmahel RF Beyreuther K Detrimental effects ofarachidonic acid and its metabolites in cellular and mouse models of Alzheimerrsquosdisease structural insight Neurobiol Aging 201233831e21ndashe31
35 Sarkar P Narayanan J Harder DR Differential effect of amyloid beta on the cyto-chrome P450 epoxygenase activity in rat brain Neuroscience 2011194241ndash249
36 Kisler K Nelson AR Montagne A Zlokovic BV Cerebral blood flow regulation andneurovascular dysfunction in Alzheimer disease Nat Rev Neurosci 201718419ndash434
37 Bibi Z Role of cytochrome P450 in drug interactions Nutr Metab (Lond) 200852738 Ownby RL Crocco E Acevedo A John V Loewenstein D Depression and risk for
Alzheimer disease systematic review meta-analysis and metaregression analysisArch Gen Psychiatry 200663530ndash538
39 Sheline YI West T Yarasheski K et al An antidepressant decreases CSF Abetaproduction in healthy individuals and in transgenic AD mice Sci Transl Med 20146236re234
NeurologyorgNG Neurology Genetics | Volume 4 Number 1 | February 2018 9
DOI 101212NXG000000000000021620184 Neurol Genet
Andreacutea L Benedet Lei Yu Aureacutelie Labbe et al variant mitigates Alzheimer disease pathophysiology in vivo and postmortemCYP2C19
This information is current as of January 30 2018
ServicesUpdated Information amp
httpngneurologyorgcontent41e216fullhtmlincluding high resolution figures can be found at
References httpngneurologyorgcontent41e216fullhtmlref-list-1
This article cites 37 articles 2 of which you can access for free at
Subspecialty Collections
httpngneurologyorgcgicollectionpetPET
httpngneurologyorgcgicollectionassociation_studies_in_geneticsAssociation studies in genetics
httpngneurologyorgcgicollectionalzheimers_diseaseAlzheimers diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpngneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpngneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
reserved Online ISSN 2376-7839Published by Wolters Kluwer Health Inc on behalf of the American Academy of Neurology All rightsan open-access online-only continuous publication journal Copyright Copyright copy 2018 The Author(s)
is an official journal of the American Academy of Neurology Published since April 2015 it isNeurol Genet

DOI 101212NXG000000000000021620184 Neurol Genet
Andreacutea L Benedet Lei Yu Aureacutelie Labbe et al variant mitigates Alzheimer disease pathophysiology in vivo and postmortemCYP2C19
This information is current as of January 30 2018
ServicesUpdated Information amp
httpngneurologyorgcontent41e216fullhtmlincluding high resolution figures can be found at
References httpngneurologyorgcontent41e216fullhtmlref-list-1
This article cites 37 articles 2 of which you can access for free at
Subspecialty Collections
httpngneurologyorgcgicollectionpetPET
httpngneurologyorgcgicollectionassociation_studies_in_geneticsAssociation studies in genetics
httpngneurologyorgcgicollectionalzheimers_diseaseAlzheimers diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpngneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpngneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
reserved Online ISSN 2376-7839Published by Wolters Kluwer Health Inc on behalf of the American Academy of Neurology All rightsan open-access online-only continuous publication journal Copyright Copyright copy 2018 The Author(s)
is an official journal of the American Academy of Neurology Published since April 2015 it isNeurol Genet





























