Embed Size (px)
Citation preview

Cyclooxygenase-2 inhibition inhibits PI3K/AKT kinase activityin epithelial ovarian cancer
Shahab Uddin1, Maqbool Ahmed1, Azhar Hussain1, Lina Assad2, Fouad Al-Dayel2, Prashant Bavi1, Khawla S. Al-Kuraya1
and Adnan Munkarah3
1 Human Cancer Genomic Research, Research Center, King Faisal Specialist Hospital and Research Center, Riyadh, Saudi Arabia2 Department of Pathology, King Faisal Specialist Hospital and Research Center, Riyadh, Saudi Arabia3 Department of Gynecology, King Faisal Specialist Hospital and Research Center, Riyadh, Saudi Arabia
Cyclooxygenase-2 (COX-2) expression contributes to tumor growth and invasion in epithelial ovarian cancer (EOC). COX-2
inhibitors exhibit important anticarcinogenic potential against EOC, but the molecular mechanisms underlying this effect and
relation with PI3-kinase/AKT signaling remain the subject of intense investigations. Therefore, the role of COX-2 in EOC and
its cross talk with PI3-kinase/AKT pathway were investigated using a large series of EOC tissues in a tissue micro array (TMA)
format followed by in vitro and in vivo studies using EOC cell lines and NUDE mice. Clinically, COX-2 was overexpressed in
60.3% of EOC and was significantly associated with activated AKT (p < 0.0001). Cox-1 expression was seen in 59.9% but did
not associate with AKT. Our in vitro data using EOC cell line showed that inhibition of COX-2 by aspirin, selective inhibitor
NS398 and gene silencing by COX-2 specific siRNA impaired phosphorylation of AKT resulting decreased downstream
signaling leading to cell growth inhibition and induction of apoptosis. Finally, treatment of MDAH2774 cell line xenografts
with aspirin resulted in growth inhibition of tumors in NUDE mice via down-regulation of COX-2 and AKT activity. These data
identify COX-2 as a potential biomarker and therapeutic target in distinct molecular subtypes of ovarian cancer.
More than 85% of human ovarian cancers are epithelial ovar-ian carcinomas (EOCs), which originate from ovarian surfaceepithelial (OSE) cells.1 The high mortality rate associatedwith epithelial ovarian cancer seems to be multifactorial.First, there is a lack of effective screening and early detectionstrategies; therefore, a high percentage of patients are diag-nosed in advanced stage disease.2 In addition, despite goodresponse to primary surgery and chemotherapy treatments,recurrence rates are high and available salvage therapies arenot curative.3–6
Although the exact etiology of EOCs is not determined,an inflammatory process is one factor suggested to participatein tumorigenesis.7 Some studies have reported an associationbetween chronic inflammatory conditions caused by talc orasbestos exposure, endometriosis or other pelvic inflamma-tory diseases and an increased incidence of EOC.8 In addi-tion, repetitive incessant ovulation has been identified as one
of the major risk factors for developing EOC.9 Since severalclassical mediators of inflammation participate in the ovula-tory process, one can speculate that each ovulation event rep-resents a localized inflammatory reaction.10 The importanceof inflammation/ovulation in ovarian tumorigenesis is furthersupported by the observed reduction in the relative risk ofovarian cancer in women with a decrease in the total numberof ovulations during a lifetime, whether due to childbearingor the use of contraceptive pills.11 In addition, evidence fromin vitro and in vivo studies suggests an important role forprostaglandins and their synthesizing enzyme COX in ovar-ian carcinogenesis. Cyclooxygenases (COX-1 and -2) are keyenzymes in the production of prostaglandins (PGs), whichare short-lived lipid signaling molecules involved in a num-ber of biologic functions.12 COX-1 expression is generallyconstitutive, whereas COX-2 is usually induced by stimuliinvolved in inflammatory responses. Prostaglandin E2(PGE2), a primary metabolite of COX-2, has been shown topromote cell survival, proliferation, and angiogenesis andprohibit apoptosis, all processes influencing cancer develop-ment.13 Recently COX-2 overexpression has been shown aslinking factor of chronic inflammation with neoplasticchanges.14 Several reports have shown that either COX-1 orCOX-2 is upregulated in EOC.15–20 Ovarian tumors withincreased content of COX-2 were associated with chemother-apy resistance and poor prognosis while COX-1 was sug-gested to participate in neovascularization.17,21,22 However,how cyclooxygenase (COX2) enhances ovarian cancer pro-gression remains poorly understood.
Key words: Cox-2, AKT, apoptosis, ovarian cancer
Additional Supporting Information may be found in the online
version of this article
The first two authors contributed equally to this work
DOI: 10.1002/ijc.24757
History: Received 21 Dec 2008; Accepted 2 Jul 2009; Online 20
Jul 2009
Correspondence to: Shahab Uddin, Human Cancer Genomic
Research, Research Center, King Faisal Specialist Hospital and
Research Cancer, MBC 98-16, P.O. Box 3354, Riyadh 11211, Saudi
Arabia, Fax: þ966-1-205-5170, E-mail: [email protected]
Can
cerCellBiology
Int. J. Cancer: 126, 382–394 (2010) VC 2009 UICC
International Journal of Cancer
IJC

Epidemiologic studies to examine whether nonsteroidolalanti-inflammatory drugs (NSAIDs) prevent or delay the de-velopment of ovarian cancers remain inconclusive. Althoughseveral population- and hospital-based case–control studiespresent evidence for time- and dose-dependent decreases inthe risk of developing ovarian cancers with the consumptionof several NSAIDs,23 other studies failed to find any signifi-cant correlation or found that the reduction in risk was asso-ciated only with the use of particular NSAIDs.24 MostNSAIDs currently in use inhibit both cyclooxygenase (COX-1) and COX-2 at their recommended dosages.25–27
Several investigators have shown that inhibition of COX2activity is cytotoxic to cancer cells in vitro and in vivo.28
Some have reported that these cytotoxic effects are inducedby modulating the activity PI3K/AKT signaling pathway.29,30
It is known that upon activation of PI3K pathway a numberof signaling proteins will be phosphorylated including proteinkinase B (AKT). AKT, once activated exert its oncogenic andantiapoptotic activity by activating several downstream tar-gets such as Bad, Foxo transcription factors and GSK3.
There are conflicting reports regarding role of COX-2 inthe pathogenesis of ovarian cancer. A recent study suggestedthat cellular proliferative activity in ovarian tumors is unre-lated to COX-231; however, there are several other studiesthat indicated a role for COX-2 in ovarian carcinogesis.32,33
This prompted us to analyze the expression of COX-2 pro-tein in a large series of EOC and explore its prognostic sig-nificance. In addition, in vitro and in vivo studies were per-formed to delineate the possible mechanisms by which COX-2 contributes to ovarian cancer pathogenesis.
Material and methodsPatient selection
One hundred fifty-six patients with ovarian carcinoma diag-nosed between 1991 and 2007 were selected from the files ofthe King Faisal Specialist Hospital and Research Centre. Allsamples were analyzed in a tissue microarray (TMA) format.The Institutional Review Board of the King Faisal SpecialistHospital and Research Centre approved the study.
The patients included in this study had their diagnosis,treatment and follow-up care in the departments of Obstet-rics and Gynecology and Oncology at King Faisal SpecialistHospital and Research Centre. The histological subtype ofeach ovarian tumor sample was determined according toestablished criteria.34 The distribution of tumors by histologictype was as follows: 125 serous (80.1%), 22 endometrioid(14.1%), 4 clear cell (2.6%) and 5 undifferentiated/mixed Epi-thelial (3.2%). The median age of the patient population was56 years with a range from 19 to 86 yr. The majority ofpatients underwent primary surgical staging or cytoreduction.In some patients who were not fit for primary surgery, pri-mary neoadjuvant chemotherapy was followed by intervaldebulking surgery. The distribution by FIGO stage at diagno-sis was: Stage I–II in 8 patients (5.1%), Stage III–IV in 137(87.8%), and unknown in 11(6.1%).
Tissue microarray(TMA) construction
Tissue microarrays were constructed from formalin-fixed,paraffin-embedded ovarian cancer specimens as describedpreviously.35,36 In addition, we also constructed a tissuemicroarray comprising of non-neoplastic ovarian surface epi-thelium from FFPE from 13 patients, who had been operatedfor hysterectomy with salpingoopherectomy. Tumor regionswere mapped by a pathologist for coring. The tissue microar-ray was constructed with 0.6-mm diameter cores spaced 0.8mm apart using a modified Tissue Microarrayer (BeecherInstruments, Sun Prairie, WI). The tissue microarray blockwas cut into 5 mm sections, adhered to the slide by an adhe-sive tape-transfer method (Instrumedics Inc., Hackensack,NJ) and UV crosslinked.
Antibodies and immunohistochemistry
Immunohistochemical studies on formalin-fixed, paraffin-em-bedded tissue sections were performed as describer in earlierstudies.35,36 For antigen retrieval, Dako Target Retrieval Solu-tion pH 6.0 (Catalogue number S1700) was used, and theslides were microwaved at 750 W for 5 minutes and then at250 W for 30 minutes. The sections were incubated overnightwith COX-2 (rabbit polycloncal COX-2 Abcam) and COX-1(mouse monoclonal, ABR,12E12) the Dako Envision PlusSystem kit was used as the secondary detection system withDAB as chromogen. IHC for p-AKT was Eperformed bystaining 3–4 lm thick tissue micro array sections with the p-AKT (Ser 473) antibody [Survival Marker: Signal Stain Phos-pho-AKT (Ser 473) IHC detection kit Product No 8100 CellSignaling Technology, Beverly, MA]. The IHC protocolincluded with the kit was followed with no modifications.Incubating the tissue in blocking solution blocked nonspecificbinding. Endogenous peroxidase activity was quenched usingperoxidase quench supplied along with the kit. Endogenousbiotin was blocked and all slides were counterstained withhematoxylin, dehydrated, cleared, and cover slipped with pre-mount. Only fresh cut TMA slides were stained simultane-ously to minimize the influence of slide aging and maximizerepeatability and reproducibility of the experiment. Two typesof negative controls were used. One was the negative controlin the kit in which the primary antibody was omitted. A pre-absorption experiment using p-AKT Ser 473 blocking peptide(Cell Signaling Technology, Beverly, MA, Product No 1140)was used as the second negative control.
Immunohistochemistry assessment
There are no established cut points available for quantitativeCOX-2 expression as there are relatively few articles in ovar-ian carcinomas,37–39 reporting a wide variations in the inci-dence of COX-2 overexpression ranging from 32% to 67.7 %.In this study, COX-2 expression was categorized by doing anH score, which combines intensity of staining in each celland percentage of stained cells. In brief, each TMA spot wasassigned a staining intensity score from 0 to 3(I0, I1–3), and a
Can
cerCellBiology
Uddin et al. 383
Int. J. Cancer: 126, 382–394 (2010) VC 2009 UICC

percent of stained tumor cells that was recorded in 5% incre-ments from a range of 0 to 100(P0, P1–3). For each spot ana-lyzed, a score was generated form the product of intensityand percent of tumor cells stained. A final H score (range 0–300) was obtained by adding the sum of individual scoresobtained for each tissue microarray spot (H score ¼ I1 � P1þ I2 � P2 þ I3 � P3). Ovarian tumors were categorized into2 groups based on H score and using the X-tile plat asdescribed below.
X-tile plots are constructed for assessment of biomarkerand optimization of cut off points based on outcome.40,41
The X-Tile plots allow determination of an optimal cut pointwhile correcting for the use of minimum P statistics. Usingthe X-Tile program, an optimal cut point for COX-2 expres-sion was determined at 160, with a Miller-Seigmund p valueof 0.5950 as determined by X-Tile. Tumors with H score<160 were classified as low expressers (n ¼ 58; 39.7%), andthose with H score >160 were classified as high expressers(n ¼ 88; 60.3%; Figs. 1a and 1c).
p-AKT scoring was done as described earlier.42,43 Briefly,phospho-AKT was scored as levels on an intensity scale rang-ing from 0 to 3. Scoring was performed as follows: 0, noappreciable staining in tumor cells; 1, barely detectable stain-ing in tumor cells; 2, appreciable staining of moderate inten-sity, distinctly marking tumor cells and 3, readily appreciablestaining of strong intensity). For purposes of statistical analy-sis, all cases staining at level 0 or 1 were grouped as p-AKTnegative and all cases staining at Level 2 and Level 3 weregrouped as p-AKT positive.
Statistical analysis
The software used for statistical analysis was statview 5.0 (SASInstitute Inc., NC). v2 tests were used to examine relationshipbetween nominal variables. The limit of significance for allanalyses was defined as a p value of 0.05; two-sided tests wereused in all calculations. Survival curves were generated usingthe Kaplan–Meier method and log rank test was used to com-pare survival in patients with COX-2 overexpressing tumorscompared with those with low COX-2 expression.
Cell culture
Epithelial ovarian cancer (EOC) cell lines SKOV3 andMDAH2774 cells were cultured in RPMI 1640 medium sup-plemented with 10% (vol/vol) fetal bovine serum, 100 U/mlPenicillin and 100 U/ml Streptomycin at 37�C in humidifiedatmosphere containing 5% CO2.
Reagents and antibodies
Direct AKT inhibitor and zVAD-fmk inhibitor were pur-chased from Calbiochem (San Diego, CA). MTT and aspirinwere purchased from Sigma (St. Louis, MO). NS398 was pur-chased from Cayman Chemical (Ann Arbor, MI) Antibodiesagainst phospho-AKT, phospho-FKHR, phospho-GSK3,cleaved caspase-3 and BID antibodies were purchased fromCell Signaling Technologies (Beverly, MA). COX-2, cyto-
chrome c, beta-actin, caspase-3, and PARP antibodies werepurchased from Santa Cruz Biotechnology, Inc. (Santa Cruz,CA). Caspase-8 antibody was purchased from R&D (USA).Annexin V kit was purchased from Molecular Probes(Eugene OR). Apoptotic DNA-ladder kit was obtained fromRoche (Penzberg, Germany).
Cell cycle analysis
Cell lines were either treated with and without aspirin for48 hr and resuspended in 500 ll hypotonic staining buffer(sodium citrate 250 mg, Triton X 0.75 ml, propidium iodide25 lg, ribonuclease A 5 lg and 250 ml water) and analyzedby flow cytometry as described previously.44
Annexin V/propidium iodide dual staining
Ovarian cancer cell lines were treated with the indicated con-centrations of aspirin. The cells were harvested and the per-centage of cells undergoing apoptosis was measured by flowcytometry after staining with fluorescein-conjugated Annex-inV/ propidium iodide as previously described.44
Cell lysis and immunoblotting
Cells were treated with aspirin as described in the legends andlysed as previously described.45 Proteins were immunoblottedwith different antibodies and visualized by the enhancedchemiluminescence (Amersham, Piscataway, NJ) method.
Measurement of mitochondrial potential using the JC1 (5,50, 6, 60-teterachloro-1, 10, 3,30- tetraethylbenzimidazolylcar-bocyanine iodide) assay kit.
After treatment of ovarian cancer cell lines with aspirinfor 48 hr, cells were incubated with 10 lM JC1 at 37�C indark for 15 minutes and mitochondrial membrane potential(% of green and red aggregates) was determined by flowcytometry as described previously.46
Assay for cytochrome c release
Release of cytochrome c from mitochondria was assayed asdescribed in a previous publication.47 Briefly, cells weretreated with and without aspirin as described in figure legendand proteins were extracted. Twenty micrograms of proteinsfrom cytosolic and mitochondrial fraction of each sampleswere analyzed by immunoblotting using anti-cytochrome cantibody.
Gene silencing using siRNA
COX-2 siRNA and Scrambled control siRNA were purchasedfrom Qiagen. For transient expression, cell lines were trans-fected by using LipofectAMINE 2000 reagent (Invitrogen)according to the manufacturer’s instructions. After incubatingthe cells for 6 hr, the lipid and siRNA complex was removedand fresh growth medium was added. Cells were lysed 48 hrafter transfection and specific protein levels were determinedby Western Blot analysis with specific antibodies against thetargeted proteins and actin as a loading control.
Can
cerCellBiology
384 COX-2 mediated signaling in ovarian cancer
Int. J. Cancer: 126, 382–394 (2010) VC 2009 UICC

Animals and xenograft study
Six weeks old NUDE mice were obtained from Jackson Labo-ratories (Maine) and maintained in a pathogen free animalfacility at least 1 week before use. All animal studies weredone in accordance with institutional guidelines. For xeno-graft study, mice were inoculated subcutaneously into theright abdominal quadrant with 5 � 106 MDAH2774 cells in200 ll PBS. After 1 week, mice were randomly assigned into3 groups; 2 groups received intraperitoneal aspirin at 1 of 2doses (100 mg/kg or 200 mg/kg); the remaining groupreceived 0.9% saline. The body weight and tumor volume ofeach mouse were monitored weekly. The tumor volume wasmeasured as previously described.48 After 5 weeks treatment,
mice were sacrificed and individual tumors were weighed,then snap-frozen in liquid nitrogen for storage.
ResultsCOX-2 expression and its correlation with p-AKT and
other clinicopathological parameters
Levels of COX-2 were examined by immunohistochemistryin a series of 156 ovarian carcinomas. High levels of COX-2expression were seen in 60.3% (88 of 146) of the ovarian car-cinomas. p-AKT over expression was seen in 52.1 %(75 of144) of the ovarian carcinoma (Fig. 1a).
As shown in Table 1 COX-2 overexpression was signifi-cantly associated with overexpression of p-AKT (p < 0.0001).
Figure 1. Expression of COX-2 and p-AKT in ovarian carcinoma. (a) COX-2 overexpression in ovarian carcinoma, (b) p-AKT overexpression in
ovarian carcinoma, (c) Reduced COX-2 expression in ovarian carcinoma, (d) Reduced p-AKT expression in ovarian carcinoma Original
magnification �200 with the inset showing a higher magnification �400. (immunoperoxidase stain) and (e) Overview of tissue micro array
slides of ovarian carcinoma.
Can
cerCellBiology
Uddin et al. 385
Int. J. Cancer: 126, 382–394 (2010) VC 2009 UICC
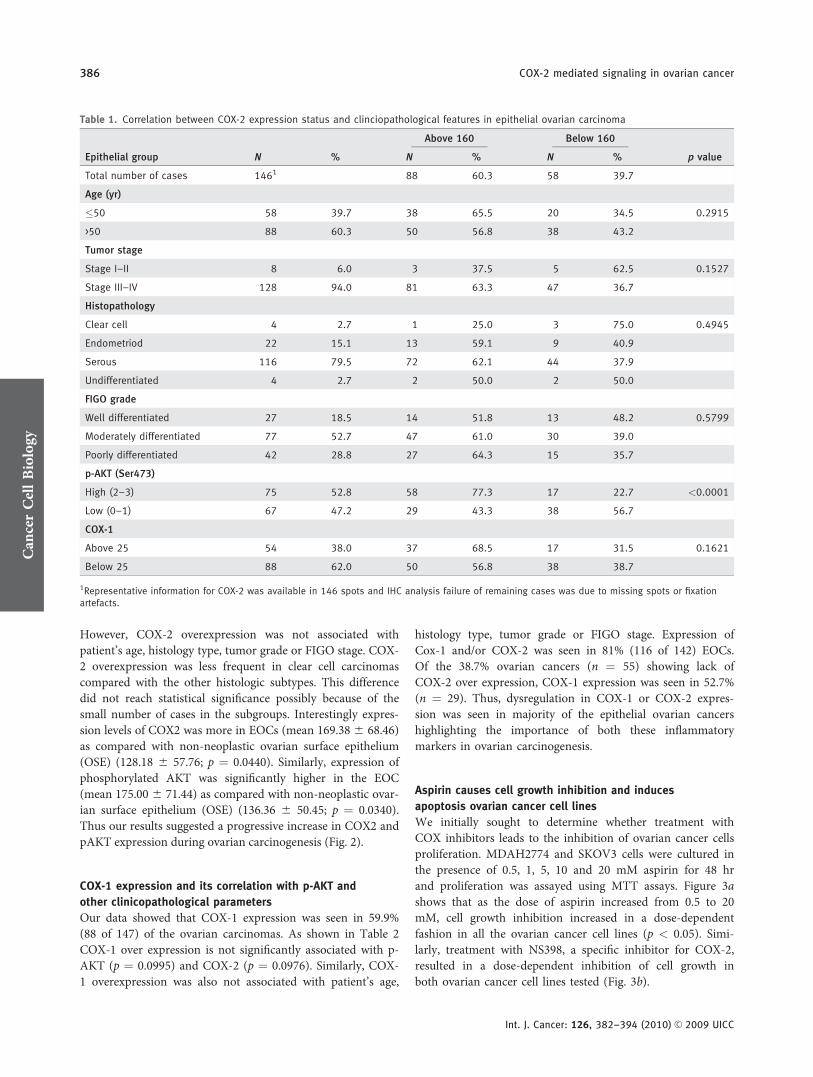
However, COX-2 overexpression was not associated withpatient’s age, histology type, tumor grade or FIGO stage. COX-2 overexpression was less frequent in clear cell carcinomascompared with the other histologic subtypes. This differencedid not reach statistical significance possibly because of thesmall number of cases in the subgroups. Interestingly expres-sion levels of COX2 was more in EOCs (mean 169.38 6 68.46)as compared with non-neoplastic ovarian surface epithelium(OSE) (128.18 6 57.76; p ¼ 0.0440). Similarly, expression ofphosphorylated AKT was significantly higher in the EOC(mean 175.00 6 71.44) as compared with non-neoplastic ovar-ian surface epithelium (OSE) (136.36 6 50.45; p ¼ 0.0340).Thus our results suggested a progressive increase in COX2 andpAKT expression during ovarian carcinogenesis (Fig. 2).
COX-1 expression and its correlation with p-AKT and
other clinicopathological parameters
Our data showed that COX-1 expression was seen in 59.9%(88 of 147) of the ovarian carcinomas. As shown in Table 2COX-1 over expression is not significantly associated with p-AKT (p ¼ 0.0995) and COX-2 (p ¼ 0.0976). Similarly, COX-1 overexpression was also not associated with patient’s age,
histology type, tumor grade or FIGO stage. Expression ofCox-1 and/or COX-2 was seen in 81% (116 of 142) EOCs.Of the 38.7% ovarian cancers (n ¼ 55) showing lack ofCOX-2 over expression, COX-1 expression was seen in 52.7%(n ¼ 29). Thus, dysregulation in COX-1 or COX-2 expres-sion was seen in majority of the epithelial ovarian cancershighlighting the importance of both these inflammatorymarkers in ovarian carcinogenesis.
Aspirin causes cell growth inhibition and induces
apoptosis ovarian cancer cell lines
We initially sought to determine whether treatment withCOX inhibitors leads to the inhibition of ovarian cancer cellsproliferation. MDAH2774 and SKOV3 cells were cultured inthe presence of 0.5, 1, 5, 10 and 20 mM aspirin for 48 hrand proliferation was assayed using MTT assays. Figure 3ashows that as the dose of aspirin increased from 0.5 to 20mM, cell growth inhibition increased in a dose-dependentfashion in all the ovarian cancer cell lines (p < 0.05). Simi-larly, treatment with NS398, a specific inhibitor for COX-2,resulted in a dose-dependent inhibition of cell growth inboth ovarian cancer cell lines tested (Fig. 3b).
Table 1. Correlation between COX-2 expression status and clinciopathological features in epithelial ovarian carcinoma
Epithelial group N %
Above 160 Below 160
p valueN % N %
Total number of cases 1461 88 60.3 58 39.7
Age (yr)
�50 58 39.7 38 65.5 20 34.5 0.2915
>50 88 60.3 50 56.8 38 43.2
Tumor stage
Stage I–II 8 6.0 3 37.5 5 62.5 0.1527
Stage III–IV 128 94.0 81 63.3 47 36.7
Histopathology
Clear cell 4 2.7 1 25.0 3 75.0 0.4945
Endometriod 22 15.1 13 59.1 9 40.9
Serous 116 79.5 72 62.1 44 37.9
Undifferentiated 4 2.7 2 50.0 2 50.0
FIGO grade
Well differentiated 27 18.5 14 51.8 13 48.2 0.5799
Moderately differentiated 77 52.7 47 61.0 30 39.0
Poorly differentiated 42 28.8 27 64.3 15 35.7
p-AKT (Ser473)
High (2–3) 75 52.8 58 77.3 17 22.7 <0.0001
Low (0–1) 67 47.2 29 43.3 38 56.7
COX-1
Above 25 54 38.0 37 68.5 17 31.5 0.1621
Below 25 88 62.0 50 56.8 38 38.7
1Representative information for COX-2 was available in 146 spots and IHC analysis failure of remaining cases was due to missing spots or fixationartefacts.
Can
cerCellBiology
386 COX-2 mediated signaling in ovarian cancer
Int. J. Cancer: 126, 382–394 (2010) VC 2009 UICC

In subsequent experiments, we determined whether theobserved suppressive effects of aspirin and NS398 in MTTassays were due to induction of cell cycle arrest or apoptosis.Ovarian cancer cell lines were treated with 5 and 10 mM ofaspirin for 48 hr or vehicle alone. The cells were stained andcell cycle fractions were determined by flow cytometry. Asshown in Supporting Information Figure 1, the sub-G1 popu-lation of cells increased from 3.77% in control cells to 23.67and 36.85% in MDAH2774 cells treated with 5 or 10 mM as-pirin, respectively. Similarly in SKOV3 cells, the sub-G1 pop-ulation was 4.33% in the control group compared with 18.12and 31.55% in cells treated with 5 or 10 mM aspirin, respec-tively. Annexin V/PI dual staining and DNA laddering detec-tion methods confirmed those apoptotic effects. Ovarian can-cer cells were treated with 5 and 10 mM aspirin for 48 hrand apoptosis was measured by annexinV/PI dual staining.As shown in Figure 3c, treatment of MDAH2774 cells with 5mM and 10 mM aspirin induces apoptosis in 21.10 and39.20% of cells, respectively. Similarly, treatment of SKOV3cells with 5 or 10 mM aspirin induced apoptosis in 21.4 and50%, respectively. Finally, ovarian cancer cells were treated
with 100 lM NS398 or 10 mM aspirin for 48 hr, DNA wasisolated using apoptotic DNA-laddering kit from Roche. Asshown in Figure 3d, inhibition of COX-2 causes fragmenta-tion formation of DNA, a characteristic of apoptotic celldeath. These results confirm that suppression of growth byNS398 and aspirin treatment in ovarian carcinoma cellsoccurs via inducing apoptosis.
Constitutive expression of COX-2 and activation of AKT
signaling pathways in ovarian cancer cell lines
Using 2 ovarian cancer cell lines, we determined the constitu-tive expression of COX-2 and AKT in these cells as well as theeffect of aspirin treatment on that expression. MDAH2774cells were treated for 24 hr with 50 or 100 lM NS398, a spe-cific COX-2 inhibitor. As shown in Figure 4a, both ovariancancer cell lines exhibited constitutive expression of COX-2and p-AKT. Treatment of ovarian cancer cells with NS398(Fig. 4a), and aspirin (Fig. 4b) suppressed COX-2 expressionand induced dephosphorylation of AKT significantly (p <
0.001) (Supporting Information Figs. 2a and 2b). Inhibition ofCOX-2 also suppressed the downstream target of AKT,
Figure 2. Comaprision of COX2 and pAKT expression in non neoplastic ovarian surface epithelium (OSE) and epithelial ovarian carinomas
(EOC). Section of a non-neoplastic ovarian surface epithelium (OSE), (a) heamtoxylin and eosin (H&E) stained (b) COX-2 expression and (c)
p-AKT expression. Original magnification �200 with the inset showing a higher magnification �400 (immunoperoxidase stain). (d and e)
Box plot charts indicate the mean and standard deviation of COX-2 expression in 2 groups—non-neoplastic ovarian surface epithelium(OSE)
and epithelial ovarian carcinomas(EOC).
Can
cerCellBiology
Uddin et al. 387
Int. J. Cancer: 126, 382–394 (2010) VC 2009 UICC

FKHR/FOXO1. These data suggest that the activity of AKT isregulated by COX-2. To verify that the effects of aspirin andNS398 are attributed to their COX-2 inhibitory effects, similarexperiments were conducted with siRNA-targeting COX-2.The COX-2-targeting siRNA, down-regulated the expressionof COX-2 protein and decreased the phosphorylation of AKT(Fig. 4c). In addition, downregulation of COX-2 and inhibi-tion of AKT caused activation of caspase-3 On the otherhand, treatment of MDAH2774 cells with specific AKT inhibi-tor or transfection of these cells with AKT-specific siRNAdecreased AKT phosphorylation but did not affect COX-2expression proving that AKT is downstream to COX-2 in thesignal pathway (Fig. 4d and Supporting Information Fig. 2c).
Inhibition of COX-2 with aspirin in ovarian cancer cells
induces apoptosis via mitochondrial pathway and caspase
activation
Next, we sought to determine whether inhibition of COX-2and p-AKT signaling involves the mitochondria in the ovar-ian cancer cells. Activated caspase-8 is capable of cleavingcaspase-3 either directly or by digesting BID to its activeform (tBID) that subsequently leads to loss of mitochondrial
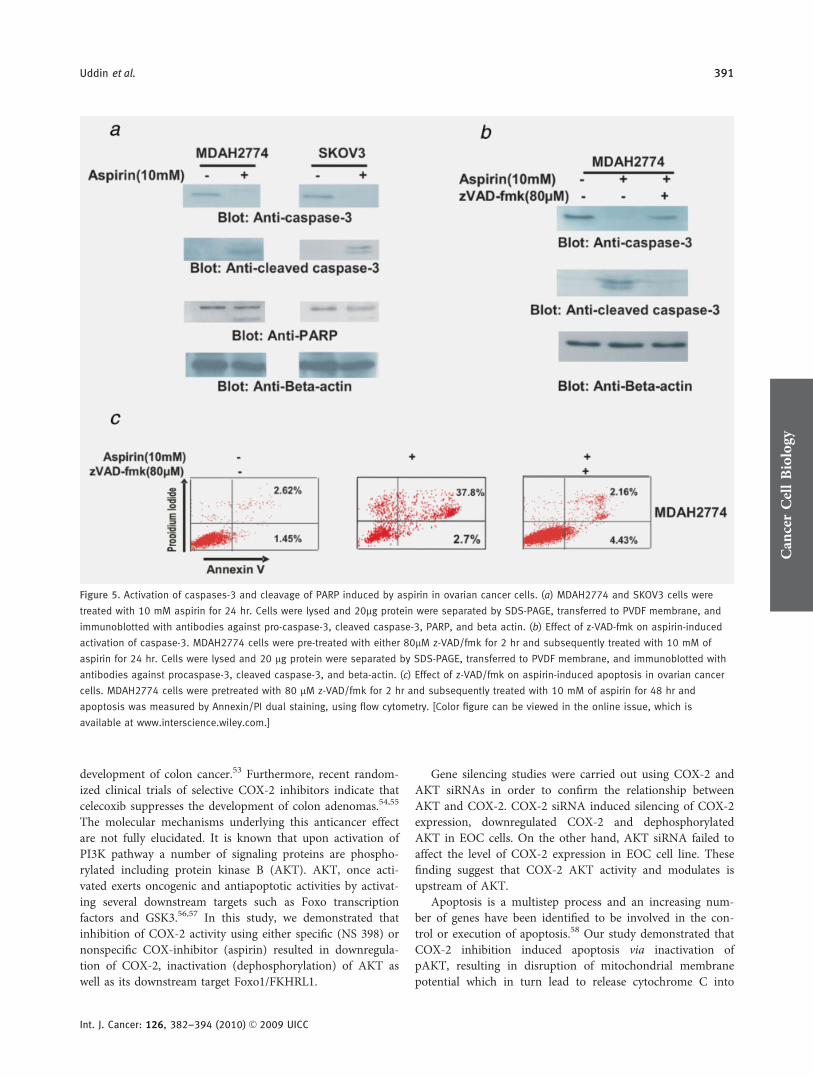
membrane potential and release of cytochrome c from mito-chondria. Aspirin treatment of MDAH2774 and SKOV3 cellsresulted in activation of caspase-8 leading to truncation ofBID in both cell lines tested (Supporting Information Fig.3a). In addition it resulted in loss of mitochondrial mem-brane potential and release of cytochrome c from mitochon-dria into cytosole (Supporting Information Figs. 3b–d). Wethen sought to determine whether aspirin-induced release ofcytochrome c is capable of activation of caspase-3 and PARP.Figure 5a shows that aspirin treatment resulted in the activa-tion of caspase-3 and cleavage of PARP in MDAH2774, andSKOV3 cells. In addition, pretreatment of these cells with 80lM z-VAD-fmk, a universal inhibitor of caspases, followedby aspirin treatment, prevented caspase-3 and PARP activa-tion induced by aspirin and abrogated apoptosis (Figs. 5band 5c), clearly indicating that caspases play a critical role inaspirin induced apoptosis in ovarian cancer cells.
In vivo activity of aspirin against ovarian cancer cells
xenograft
Our observation that ovarian cancer cells exhibit enhancedsensitivity to aspirin-induced apoptosis in vitro suggests a
Table 2. Correlation between COX-1 expression status and clinciopathological features in epithelial ovarian carcinoma
Epithelial group N %
Above 25 Below 25
p valueN % N %
Total number of cases 1471 54 36.7 93 63.3
Age (yr)
�50 57 38.8 23 40.3 34 59.7 0.4701
>50 90 61.2 31 34.4 59 65.6
Tumor stage
Stage I–II 8 5.8 2 25.0 6 75.0 0.5293
Stage III–IV 129 94.2 46 35.7 83 64.3
Histopathology
Clear cell 4 2.7 1 25.0 3 75.0 0.1752
Endometriod 21 14.3 4 19.0 17 81.0
Serous 117 79.6 48 41.0 69 59.0
Undifferentiated 5 3.4 1 20.0 4 80.0
FIGO grade
Well differentiated 27 18.4 10 37.0 17 63.0 0.8553
Moderately differentiated 78 53.1 30 38.5 48 61.5
Poorly differentiated 42 28.6 14 33.3 28 66.7
p-AKT (Ser473)
High (2–3) 73 51.4 30 41.1 43 58.9 0.2541
Low (0–1) 69 48.6 22 31.9 47 68.1
COX-2
Above 160 87 61.3 37 42.5 50 57.5 0.1621
Below 160 55 38.7 17 30.9 38 69.1
1Representative information for COX-1 was available in 147 spots and IHC analysis failure of remaining cases was due to missing spots or fixationartefacts.
Can
cerCellBiology
388 COX-2 mediated signaling in ovarian cancer
Int. J. Cancer: 126, 382–394 (2010) VC 2009 UICC

potential therapeutic role in treatment of ovarian cancer. Weexamined the ability of aspirin to inhibit ovarian cancer tu-mor growth in a mouse xenograft model of ovarian cancer.NUDE mice were inoculated subcutaneously in the right ab-dominal quadrant with 5 million MDAH2774 cells. The ani-mals were then treated intraperitoneally with 1 of 2 doses ofaspirin (100 mg/kg/dose and 200 mg/kg/dose) twice perweek,49 or vehicle saline-treated control groups (n ¼ 6). After5 weeks of treatment, mice were sacrificed and tumors werecollected. Aspirin treatment caused a time-dependent regres-sion of MDAH2774 xenograft tumors in mice as comparedwith vehicle-treated mice (6A). The regression reached signif-icance (p < 0.05) at the end of third week of treatment. Asignificant reduction in tumor weight (Fig. 6b) was observedin mice treated with aspirin (p < 0.05). Additionally, images
of tumor before and after necropsy showed that aspirin treat-ment resulted in shrinkage of tumor size (Fig. 6c). The levelof COX-2 and p-AKT proteins were markedly decreased intumors of mice treated with aspirin as compared to vehicle-treated mice as detected by western blotting (Fig. 6d).
DiscussionIn this study, we first investigated the incidence and clinico-pathological associations of both COX-1 and COX-2 expres-sion in a large series of EOC patient samples. COX-2 overex-pression was significantly associated with activated AKT (p-AKT) expression in the clinical samples studied. These datasuggest an interaction between COX-2 enzymatic activity andPI3Kinase/AKT signaling pathway in EOC. COX-1 expres-sion showed a direct trend with expression of p-AKT and
Figure 3. Aspirin (a) and NS398 (b) inhibit the proliferation of ovarian cancer cells. MDAH2774 and SKOV3 cells were incubated with 0.5,
1, 5, 10 and 20 mM aspirin and with 10, 25, 50 and 100 lM NS398 for 48 hr. Cell proliferation assays were performed using MTT as
described in Material and methods. The graph displays the mean 6 standard deviation (SD) of 3 independent experiments with replicates
of 6 wells for all the doses and vehicle control for each experiment *p < 0.05, statistically significant (Student’s t-test). (c) Aspirin-induced
apoptosis detected by Annexin V/PI dual staining. MDAH2774 and SKOV3 cells were treated with 5 and 10 mM for 48 hr and cells were
subsequently stained with flourescein-conjugated annexin-V and propidium iodide (PI). (d) MDAH2774 cells were treated with 10 mM
aspirin and 100 lM NS-398 for 48 hr cells and DNA was extracted and separated by electrophoresis on 1.5% agarose gel. [Color figure can
be viewed in the online issue, which is available at www.interscience.wiley.com.]
Can
cerCellBiology
Uddin et al. 389
Int. J. Cancer: 126, 382–394 (2010) VC 2009 UICC

COX-2. Almost half of the EOCs showing lack of COX2expression showed overexpression of COX-1. Thus, dysregu-lation in COX-1 or COX-2 expression was seen in 81.7%(116 of 142) highlighting the importance of both theseinflammatory markers in ovarian carcinogenesis.
A number of studies have shown that COX-2 inhibitorscan be used for the prevention and/or treatment of a numberof malignancies.50–52 In fact, epidemiological studies andrandomized clinical trials provide convincing evidence thatregular intake of aspirin and other NSAIDs inhibit the
Figure 4. NS398 and aspirin treatment causes downregulation of COX-2 and dephosphorylation of constitutive phosphorylation of AKT and
Foxo1 in ovarian cancer cell lines. (a) MDAH2774 cells were treated with 50 and 100 lM for 24 hr. After cell lysis, equal amounts of
proteins were separated by SDS-PAGE, transferred to immobilon membrane, and immunoblotted with antibodies against COX-2, p-AKT, AKT,
p-Foxo1, Foxo1 and beta actin as indicated. (b) MDAH2774 and SKOV3 cells were treated with 5 and 10 mM for 24 hr. After cell lysis,
equal amounts of proteins were separated by SDS-PAGE, transferred to immobilon membrane, and immunoblotted with antibodies against
COX-2, p-AKT, AKT, p-Foxo1, Foxo1 and beta actin as indicated. (c) MDAH2774 cells were transfected with scrambled siRNA (100 nM) and
COX-2 siRNA (50 and 100 nM) with Lipofectamine 2000. After 48 hr, cells were lysed and proteins were immunoblotted with antibodies
against COX-2, p-AKT-Ser473, AKT, caspase-3 and beta actin. (d) MDAH2774 cells were transfected with scrambled siRNA (100nM) and
AKT1 siRNA (50 and 100 nM) with Lipofectamine 2000. After 48 hr, cells were lysed and proteins were immunoblotted with antibodies
against COX-2, p-AKT-Ser473, AKT, caspase-3 and beta actin. Every experiment is representative of a minimum of 3 independent
experiments. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
Can
cerCellBiology
390 COX-2 mediated signaling in ovarian cancer
Int. J. Cancer: 126, 382–394 (2010) VC 2009 UICC
development of colon cancer.53 Furthermore, recent random-ized clinical trials of selective COX-2 inhibitors indicate thatcelecoxib suppresses the development of colon adenomas.54,55
The molecular mechanisms underlying this anticancer effectare not fully elucidated. It is known that upon activation ofPI3K pathway a number of signaling proteins are phospho-rylated including protein kinase B (AKT). AKT, once acti-vated exerts oncogenic and antiapoptotic activities by activat-ing several downstream targets such as Foxo transcriptionfactors and GSK3.56,57 In this study, we demonstrated thatinhibition of COX-2 activity using either specific (NS 398) ornonspecific COX-inhibitor (aspirin) resulted in downregula-tion of COX-2, inactivation (dephosphorylation) of AKT aswell as its downstream target Foxo1/FKHRL1.
Gene silencing studies were carried out using COX-2 andAKT siRNAs in order to confirm the relationship betweenAKT and COX-2. COX-2 siRNA induced silencing of COX-2expression, downregulated COX-2 and dephosphorylatedAKT in EOC cells. On the other hand, AKT siRNA failed toaffect the level of COX-2 expression in EOC cell line. Thesefinding suggest that COX-2 AKT activity and modulates isupstream of AKT.
Apoptosis is a multistep process and an increasing num-ber of genes have been identified to be involved in the con-trol or execution of apoptosis.58 Our study demonstrated thatCOX-2 inhibition induced apoptosis via inactivation ofpAKT, resulting in disruption of mitochondrial membranepotential which in turn lead to release cytochrome C into
Figure 5. Activation of caspases-3 and cleavage of PARP induced by aspirin in ovarian cancer cells. (a) MDAH2774 and SKOV3 cells were
treated with 10 mM aspirin for 24 hr. Cells were lysed and 20lg protein were separated by SDS-PAGE, transferred to PVDF membrane, and
immunoblotted with antibodies against pro-caspase-3, cleaved caspase-3, PARP, and beta actin. (b) Effect of z-VAD-fmk on aspirin-induced
activation of caspase-3. MDAH2774 cells were pre-treated with either 80lM z-VAD/fmk for 2 hr and subsequently treated with 10 mM of
aspirin for 24 hr. Cells were lysed and 20 lg protein were separated by SDS-PAGE, transferred to PVDF membrane, and immunoblotted with
antibodies against procaspase-3, cleaved caspase-3, and beta-actin. (c) Effect of z-VAD/fmk on aspirin-induced apoptosis in ovarian cancer
cells. MDAH2774 cells were pretreated with 80 lM z-VAD/fmk for 2 hr and subsequently treated with 10 mM of aspirin for 48 hr and
apoptosis was measured by Annexin/PI dual staining, using flow cytometry. [Color figure can be viewed in the online issue, which is
available at www.interscience.wiley.com.]
Can
cerCellBiology
Uddin et al. 391
Int. J. Cancer: 126, 382–394 (2010) VC 2009 UICC

cytosol. Release of cytochrome C was associated with activa-tion of caspase activity, eventually resulting in apoptosis.
We have used MDAH2774 cell line xenograft tumor inNUDE mice to translate the in vitro results into an in vivo
animal model, Treatment of xenograft tumors with aspirinresulted in a reduction of tumors’ size in a time and dose-de-pendent manner. The body weighs of control and experimen-tal groups of animal did not differ through out the
Figure 6. Aspirin-inhibits growth of MDAH2774 xenograft and down-regulates COX-2 and inactivates AKT in vivo. Nude mice at 6 weeks of
age were injected s.c. with five million MDAH2774 cells. After 1 week, mice were treated with aspirin at 100 mg/kg/dose, 200 mg/kg/dose
or with 5% DMSO in PBS as a vehicle control. (a) Inhibition of MDAH2774 tumor growth by aspirin. The volume of each tumor was
measured every week. The average (n ¼ 6) tumor volume in vehicle-treated control mice and treated with aspirin was plotted, *p < 0.05.
(b) After 5 weeks of treatment, mice were sacrificed and tumor weights were measured *p < 0.05 compared with vehicle-treated mice by
Student’s t-test. (c) Representative tumor images of vehicle and aspirin treated mice. (d) Whole cell lysates from mice treated with vehicle,
aspirin (100 mg/kg) and aspirin (200 mg/kg) were prepared, and 20 lg protein were separated by SDS-PAGE, transferred to PVDF
membrane, and immunoblotted with antibodies against COX-2, p-Akt, Akt and beta-actin.
Can
cerCellBiology
392 COX-2 mediated signaling in ovarian cancer
Int. J. Cancer: 126, 382–394 (2010) VC 2009 UICC

experiments suggesting that aspirin treatment significantlyinhibited MDAH2774 xenograft growth without causing anyside effects to mice. Tumor regression was accompanied by adecrease the level of phosphorylated AKT and COX-2 in thetumor cells.
Altogether these data suggest that COX-2 may play rolein tumor growth through regulation of key oncogenic path-ways (PI3K/AKT) on which cancer cells depend for survival.Furthermore, our data also support the hypothesis that COX-2 inhibition may have therapeutic potential in EOC.
References
1. Auersperg N, Wong AS, Choi KC, KangSK, Leung PC. Ovarian surface epithelium:biology, endocrinology, and pathology.Endocr Rev 2001;22:255–8.
2. Fishman DA, Bozorgi K. The scientificbasis of early detection of epithelial ovariancancer: the national ovarian cancer earlydetection program (NOCEDP). In: StackMS, Fishman DA, eds. Cancer treatmentand research: ovarian cancer. Boston:Kluwer Academic Publishers, 2001. 3–28.
3. Fader AN, Rose PG. Role of surgery inovarian carcinoma. J Clin Oncol 2007;25:2873–83.
4. Salzberg M, Thurlimann B, Bonnefois H,Fink D, Rochlitz C, von Moos R, Senn H.Current concepts of treatment strategies inadvanced or recurrent ovarian cancer.Oncology 2005;68:293–8.
5. McGuire WP, Ozols RF. Chemotherapy ofadvanced ovarian cancer. Semin Oncol1998;25:340–8.
6. Fung-Kee-Fung M, Oliver T, Elit L, Oza A,Hirte HW, Bryson P. Optimalchemotherapy treatment for women withrecurrent ovarian cancer. Curr Oncol 2007;4:195–208.
7. Ness RB, Cottreau C. Possible role ofovarian epithelial inflammationin ovariancancer. J Natl Cancer Inst 1999;91:1459–67.
8. Ness RB, Grisso JA, Cottreau C, Klapper J,Vergona R, Wheeler JE, Morgan M,Schlesselman JJ. Factors related toinflammation of the ovarian epitheliumand risk of ovarian cancer. Epidemiology2000;11:111–17.
9. Fathalla MF. Incessant ovulation–a factorin ovarian neoplasia?Lancet 1971;2:63.
10. Brannstrom M, Enskog A. Leukocytenetworks and ovulation. J Reprod Immunol2002;57:47–60.
11. Riman T, Nilsson S, Persson IR. Review ofepidemiological evidence for reproductiveand hormonal factors in relation to therisk of epithelial ovarian malignancies. ActaObstet Gynecol Scand 2004;83:783–95.
12. Williams CS, DuBois RN. Prostaglandinsendoperoxide synthase: why two isoforms?Am J Physiol 1996;270:G393–G400.
13. Gierse JK, Hauser SD, Creely DP, KoboldtC, Rangwala SH, Isakson PC, Seibert K.Expression and selective inhibition of theconstitutive and inducible forms of humancyclo-oxygenase. Biochem J 1995;15:479–84.
14. Colby JK, Klein DR, McArthur MJ, ContiCJ, Kiguchi K, Kawamoto T, Riggs PK,
Pavone AI, Sawicki J, Fischer SJ.Progressive metaplastic and dysplasticchanges in mouse pancreas induced bycyclooxygenase-2 overexpression. Neoplasia2008;10:782–96.
15. Daikoku T, Tranguch S, Trofimova IN,Dinulescu DM, Jacks T, Nikitin AY,Connolly DC, Dey SK. Cyclooxygenase-1 isoverexpressed in multiple geneticallyengineered mouse models of epithelialovarian cancer. Cancer Res 2006;66:2527–31.
16. Denkert C, Furstenberg A, Daniel PT,Koch I, Kobel M, Weichert W, SiegertAHauptmann S. Induction of G0/G1 cellcycle arrest in ovarian carcinoma cells bythe anti-inflammatory drug NS-398, butnot by COX-2-specific RNA interference.Oncogene 2003;22:8653–61.
17. Gupta RA, Tejada LV, Tong BJ, Das SK,Morrow JD, Dey SK, DuBois RN.Cyclooxygenase-1 is overexpressed andpromotes angiogenic growth factorproduction in ovarian cancer. Cancer Res2003;63:906–11.
18. Symowicz J, Adley BP, Woo MM,Auersperg N, Hudson LG, Stack MS.Cyclooxygenase-2 functions as adownstream mediator of lysophosphatidicacid to promote aggressive behavior inovarian carcinoma cells. Cancer Res 2005;65:2234–42.
19. Athanassiadou P, Grapsa D, AthanassiadesP, Gonidi M, Athanassiadou AM, Tsipis A,Patsouris E. The prognostic significance ofCOX-2 and survivin expression in ovariancancer. Pathol Res Pract 2008;204:241–9.
20. Lee JS, Choi YD, Lee JH, Nam JH, Choi C,Lee MC, Park CS, Juhng SW, Min KW.Expression of cyclooxygenase-2 inepithelial ovarian tumors and its relation tovascular endothelial growth factor and p53expression. Int J Gynecol Cancer 2006;16:247–53.
21. Ferrandina G, Lauriola L, Zannoni GF,Fagotti A, Fanfani F, Legge F, Maggiano N,Gessi M, Mancuso S, Ranelletti FO,Scambia G. Increased clyclooxygenase-2(COX-2) expression is associated withchemotherapy resistance and outcome inovarian cancer patients. Ann Oncol 2002;13:1205–11.
22. Denkert C, Kobel M, Pest S, Koch I,Berger S, Schwabe M, Siegert A, Reles A,Klosterhalfen B, and Hauptmann S.Expression of clyclooxygenase-2 is an
independent prognostic factor in humanovarian carcinoma. Am J Pathol 2002;160:893–903.
23. Harris RE, Beebe-Donk J, Doss H, BurrDoss D. Aspirin, ibuprofen, and other non-steroidal anti-inflammatory drugs in cancerprevention: a critical review of non-selective COX-2 blockade. Oncol Rep 2005;13:559–83.
24. Ulrich CM, Bigler J, Potter JD. Non-steroidal anti-inflammatory drugs forcancer prevention: promise, perils andpharmacogenetics. Nat Rev Cancer 2006;6:130–40.
25. Meade EA, Smith WL, DeWitt DL.Differential inhibition of prostaglandinendoperoxide synthase (cyclooxygenase)isozymes by aspirin and other non-steroidal anti-inflammatory drugs. J BiolChem 1993;268:6610–14.
26. Laneuville O, Breuer DK, Dewitt DL, HlaT, Funk CD, Smith WL. Differentialinhibition of human prostaglandinendoperoxide H synthases-1 and -2 bynonsteroidal anti-inflammatory drugs. JPharmacol Exp Ther 1994;271:927–34.
27. O’Neill GP, Mancini JA, Kargman S,Yergey J, Kwan MY, Falgueyret JP,Abramovitz M, Kennedy BP, Ouellet M,Cromlish W. Overexpression of humanprostaglandin G/H synthase-1 and -2 byrecombinant vaccinia virus: inhibition bynonsteroidal anti-inflammatory drugs andbiosynthesis of 15-hydroxyeicosatetraenoicacid. Mol Pharmacol 1994;45:245–54.
28. Adhami VM, Malik A, Zaman N, SarfarazS, Siddiqui IA, Syed DN, Afaq F, Pasha FS,Saleem M, Mukhtar H. Combinedinhibitory effects of green tea polyphenolsand selective cyclooxygenase-2 inhibitorson the growth of human prostate cancercells both in vitro and in vivo. Clin CancerRes 2007;13:1611–19.
29. Hsu AL, Ching TT, Wang DS, Song X,Rangnekar VM, Chen CS. Thecyclooxygenase-2 inhibitor celecoxibinduces apoptosis by blocking Aktactivation in human prostate cancer cellsindependently of Bcl-2. J Biol Chem 2000;275:11397–403.
30. Fan XM, Jiang XH, Gu Q, Ching YP, HeH, Xia HH, Lin MC, Chan AO, Yuen MF,Kung HF, Wong BC. Inhibition of Akt/PKB by a COX-2 inhibitor inducesapoptosis in gastric cancer cells. Digestion2006;73:75–83.
Can
cerCellBiology
Uddin et al. 393
Int. J. Cancer: 126, 382–394 (2010) VC 2009 UICC

31. Yoshida A, Sarian LO, Andrade LA,Pignataro F, Pinto GA, Derchain SF. Cellproliferation activity unrelated to COX-2expression in ovarian tumors. Int J GynecolCancer 2007;17:607–14.
32. Gu P, Su Y, Guo S, Teng L, Xu Y, Qi J,Gong H, Cai Y. Over-expression of COX-2induces human ovarian cancer cells(CAOV-3) viability, migration andproliferation in association with PI3-k/Aktactivation. Cancer Invest 2008;26:822–9.
33. Bijman MN, Hermelink CA, van BerkelMP, Laan AC, Janmaat ML, Peters GJ,Boven E. Interaction between celecoxib anddocetaxel or cisplatin in human cell linesof ovarian cancer and colon cancer isindependent of COX-2 expression levels.Biochem Pharmacol 2008;75:427–37.
34. Russel P. Surface epithelial-stromal tumorsof the ovary. In: Kurman RJ, ed.Blaustein’s pathology of the female genitaltract. New York: Springer-Verlag, 1995.705–82.
35. Siraj AK, Bavi P, Abubaker J, Jehan Z,Sultana M, Al-Dayel F, Al-Nuaim A,Alzahrani A, Ahmed M, Al-Sanea O,Uddin S, Al-Kuraya KS. Genome-wideexpression analysis of Middle Easternpapillary thyroid cancer reveals c-MET as anovel target for cancer therapy. J Pathol2007;213:190–9.
36. Abubaker J, Jehan Z, Sultana M, Al-HarbiS, Ibrahim M, Al-Nuaim A, Ahmed M,Amin T, Al-Fehaily M, Al-Sanea O, Al-Dayel F, Uddin S, Al-Kuraya KS.Clinicopathological analysis of papillarythyroid cancer with PIK3CA alterations inMiddle Eastern Population. J ClinEndocrionol Metab 2008;93:611–18.
37. Symowicz J, Adley BP, Woo MM,Auersperg N, Hudson LG, Stack MS.Cyclooxygenase-2 functions as adownstream mediator of lysophosphatidicacid to promote aggressive behavior inovarian carcinoma cells. Cancer Res 2005;65:2234–2242.
38. Steffensen KD, Waldstrøm M, Jeppesen U,Jakobsen E, Brandslund I, Jakobsen A. Theprognostic importance of cyclooxygenase 2and HER2 expression in epithelial ovariancancer. Int J Gynecol Cancer 2007;17:798–807.
39. Ali-Fehmi R, Che M, Khalifeh I, MaloneJM, Morris R, Lawrence WD, MunkarahAR. The effect of cyclooxygenase-2expression on tumor vascularity inadvanced stage ovarian serous carcinoma.Cancer 2003;98:1423–9.
40. McCarthy MM, Sznol M, DiVito KA,Camp RL, Rimm DL, Kluger HM.Evaluating the expression and prognosticvalue of TRAIL-R1 and TRAIL-R2 inbreast cancer. Clin Cancer Res 2005;11:5188–94.
41. Camp RL, Dolled-Filhart M, Rimm DL. X-tile: a new bio-informatics tool forbiomarker assessment and outcome-basedcut-point optimization. Clin Cancer Res2004;10:7252–9.
42. Uddin S, Hussain AR, Siraj AK,Manogaran PS, Al-Jomah NA, Moorji A,Atizado V, Al-Dayel F, Belgaumi A, El-Solh H, Ezzat A, Bavi P, et al. Role ofphosphatidylinositol 30-kinase/AKTpathway in diffuse large B-cell lymphomasurvival. Blood 2006;108:4178–86.
43. Abubaker J, Bavi P, Al-Harbi S, IbrahimM, Siraj AK, Al-Sanea N, Abduljabbar A,Ashari LH, Alhomoud S, Al-Dayel F,Uddin S, Al-Kuraya KS.Clinicopathological analysis of colorectalcancers with PIK3CA mutations in MiddleEastern population. Oncogene 2008;27:3539–45.
44. Uddin S, Siraj AK, Al-Rasheed M, AhmedM, Bu R, Myers JN, Al-Nuaim A, Al-SobhiS, Al-Dayel F, Bavi P, Hussain AR, Al-Kuraya KS. Fatty acid synthase and AKTpathway signaling in a subset of papillarythyroid cancers. J Clin Endocrinol Metab2008;93:4088–97.
45. Uddin S, Ah-Kang J, Ulaszek J, MahmudD, Wickrema A. Differentiation stage-specific activation of p38 mitogen-activatedprotein kinase isoforms in primary humanerythroid cells. Proc Natl Acad Sci USA2004;101:147–52.
46. Uddin S, Hussain AR, Al-Hussein KA,Manogaran PS, Wickrema A, Gutierrez MI,Bhatia KG. Inhibition ofphosphatidylinositol 30-kinase/AKTsignaling promotes apoptosis of primaryeffusion lymphoma cells. Clin Cancer Res2005;11:3102–8.
47. Uddin S, Hussain AR, Manogaran PS, Al-Hussein K, Platanias LC, Gutierrez MI,Bhatia KG. Curcumin suppresses growthand induces apoptosis in primary effusionlymphoma. Oncogene 2005;24:7022–30.
48. Uddin S, Ahmed M, Bavi P, El-Sayed R,Al-Sanea N, abduljabbar A, Ashari LH,Alhomoud S, Al-Dayel F, Hussain A, Al-Kuraya KS. Bortezomib (VelcadeVR ) inducesp27Kip1 expression through SKP2degradation in colorectal cancer. CancerRes 2008;68:3379–88.
49. Stark LA, Reid K, Sansom OJ, Din FV,Guichard S, Mayer I, Jodrell DI, ClarkeAR, Dunlop MG. Aspirin activates the NF-KB signalling pathway and inducesapoptosis in intestinal neoplasia in two invivo models of human colorectal cancer.Carcinogenesis 2007;28:968–76.
50. Sun WH, Zhu F, Chen GS, Su H, Luo C,Zhao QS, Zhang Y, Shao Y, Sun J, ZhouSM, Ding GX, Cheng YL. Blockade ofcholecystokinin-2 receptor andcyclooxygenase-2 synergistically inducescell apoptosis, and inhibits the proliferationof human gastric cancer cells invitroCancer Lett 2008;263:302–11.
51. Li QB, Chen ZC, You Y, Zou P. Smallinterfering RNA of cyclooxygenase-2induces growth inhibition and apoptosisindependently of Bcl-2 in human myelomaRPMI8226 cells. Acta Pharmacol Sin 2007;28:1031–6.
52. Alam M, Wang JH, Coffey JC, Qadri SS,O’Donnell A, Aherne T, Redmond P.Characterization of the effects ofcyclooxygenase-2 inhibition in theregulation of apoptosis in human smalland non-small cell lung cancer cell lines.Ann Surg Oncol 2007;14:2678–84.
53. Harris RE. Cyclooxygenase-2 (COX-2) andthe inflammogenesis of cancer. SubcellBiochem 2007;42:93–126.
54. Bertagnolli,MM, Eagle CJ, Zauber AG,Redston M, Solomon,SD, Kim K, Tang J,Rosenstein RB, Wittes J, Core D, Hess TM,Woloj GM, et al.; APC Study Investigators.Celecoxib for the prevention of sporadiccolorectal adenomas. N Engl J Med 2006;355:873–884.
55. Arber N, Eagle CJ, Spicak J, Racz,I, Dite P,Hajer J, Zavoral M, Lechuga MJ, Gerletti P,Tang J, Rosenstein RB, Macdonald K,et al.; PreSAP Tri al Investigators.Celecoxib for the prevention of colorectaladenomatous polyps. N Engl J Med 2006;355:885–95.
56. Brunet A, Park J, Tran H, Hu LS,Hemmings BA, Greenberg ME. Proteinkinase SGK mediates survival signals byphosphorylating the forkhead transcriptionfactor FKHRL1 (FOXO3a). Mol. Cell Biol2001;21:952–65.
57. Alvarez B, Martinez- AC, Burgering BM,Carrera AC. Forkhead transcription factorscontribute to execution of the mitoticprogramme in mammals. Nature 2001;413:744–7.
58. Gastman BR. Apoptosis and its clinicalimpact. Head Neck 2001;23:409–25.
Can
cerCellBiology
394 COX-2 mediated signaling in ovarian cancer
Int. J. Cancer: 126, 382–394 (2010) VC 2009 UICC







![Targeting of PI3K/AKT/mTOR pathway to inhibit T cell activation … · 2017. 8. 25. · AKT/mammalian target of rapamycin (PI3K/AKT/ mTOR) [1]. This pathway controls numerous cellular](https://img.pdfslide.us/doc/110x75/60af5eaa6ab71f4bc15363aa/targeting-of-pi3kaktmtor-pathway-to-inhibit-t-cell-activation-2017-8-25-aktmammalian.jpg)





















