Embed Size (px)
Citation preview

Cyclic Changes in the Squamo-columnar Junction of the Mouse Cervix Uteri
CHARLES E. GRAHAM' Department of Anatomy, Yale University School of Medicine, New Haven, Connecticut
ABSTRACT The histological structure of the squamo-columnar junction of the cervix uteri of mice was studied during the estrous cycle, pregnancy, lactation, and in ovariectomized and estrogen-stimulaied animals. In the absence of endogenous or exogenous estrogen, there was no clear demarcation between the squamous and the columnar epithelium; the epithelium in the zone of transition resembled types pre- viously described in the human cervix that have been regarded as evidence of a metaplasia of columnar to stratified epithelium. However, in the mouse, estrogen stimulation caused the disappearance of the transitional zone so that an abrupt junc- tion resulted. It was concluded that the type of junction is controlled by estrogen, and that the stratified epithelium of the transitional zone may differentiate to pro- duce either keratinized cells or cells of uterine type at its surface.
Although there have been many studies on the normal and diseased human cervix uteri (summarized by Fluhmann, '61 ) , no attempts have been made to relate the histological structure of the actual squamo- columnar junction to the stages of the reproductive cycle in any species. This omission is surprising, because it appears from the work of Fluhmann ('59) that the junction region is an unstable zone of transition between two epithelial types, one of which is an extension of the estro- gen-sensitive vaginal epithelium, the other a mucus-secreting columnar epithelium.
The junction area is the site of origin of most cervical carcinoma (Fluhmann, '60): a full understanding of cyclic changes in this region would seem to be an im- portant prerequisite for histological stud- ies of the etiology of this disease. In the present paper a study of cyclic changes in the mouse cervix is presented.
MATERIALS AND METHODS
Animals. For most of the study, White Wistar mice, three-four months old, were used. A smaller number of hybrid mice (CBA X C,,) were used: both types gave similar results. The stage of estrous cycle was established by the vaginal smear technique (Allen, '22) and confirmed by histological examination of the vagina.
Histological methods. All tissues were fixed in Zenker-acetic. Sections of the
ANAT. REC., 155: 251-260.
cervices were cut serially in the frontal plane at 8.0 v. Every twentieth section was mounted on a slide, and these were stained with Ehrlich's haematoxylin and eosin or Papanicolaou's polychrome stain (Papanicolaou, '42). Usually duplicate sec- tions were incubated with diastase and then stained in periodic-acid Schiff (PAS) followed by Erhlichs haematoxylin, so demonstrating neutral mucopolysacchar- ides, and adjacent sections with alcian blue, specific for acid mucopolysacchar- ides, followed by neutral red (Pearse, '62).
Ovariectomy was carried out under anes- thesia with sodium pentobarbital (Pilgrim and DeOme, '55) through two dorso-lateral incisions in the body wall; complete re- moval of the ovaries was c o n h e d at autopsy. Estradiol benzoate was adminis- tered to three animals one week after ovariectomy. The estradiol benzoate, ob- tained from the National Biochemicals Corporation, was dissolved in sesame oil to give a concentration of 64.0 wg hor- mone/ml. Each animal was injected sub- cutaneously with 0.02 ml of this solution; control mice received oil alone.
RESULTS The genera1 anatomical and histological
relationships of the mouse cervix are illus- trated in figure 1. Just above the point of
1 Present address: Yerkes Regional Primate Research Center, Emory University, Atlanta, Georgia 30322.
25 1

252 CHARLES E. GRAHAM
bifurcation of the endocervical canal, the cervical stratified epithelium, which is continuous with that of the vagina, gives way to a columnar epithelium similar to and continuous with the epithelium lining the lumen of the uterine cornua. In ana- tomical terms there are two squamo- columnar junctions in each cervix, one in each half of the bifurcated cervical canal; thus in each coronal section colum- nar and stratified epithelium is seen to meet in four places (fig. 1).
Many observations of the anatomical junctions were made in each cervix, be- cause every twentieth section was ex- amined (excluding sections that were too tangential to permit unequivocal interpre- tation). Each meeting-point of the two types of epithelia seen in sections will be regarded as an individual junction. This avoids the semantic difficulties which arise because different parts of the same anatomical junction sometimes differ in structure, and it allows the classification of the many junctions observed in sec- tions into several different types, the rela- tive frequency of which can be expressed as a percentage of the total number of observations of junctions made in each group of animals (see table 1).
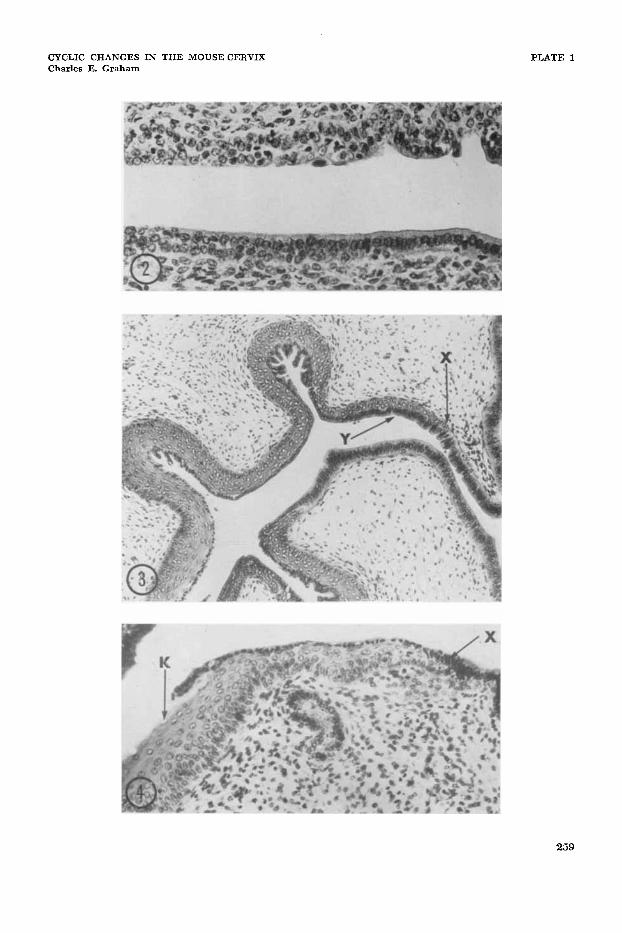
Two basic types of squamo-columnar junction were recognized. One of these, termed “transitional,” showed a gradual increase in number of cell layers between the uterine columnar and vaginal strati- fied epithelium so that the two types were confiuent (figs. 2-4). In the second type
COLUMNAR \\ V h
VAGINAL FORNL
EXTERNAL 0s
Fig. 1 Schematic frontal section through mouse cervix, showing location of squarno- columnar junction and distribution of squamous epithelium (thick line),
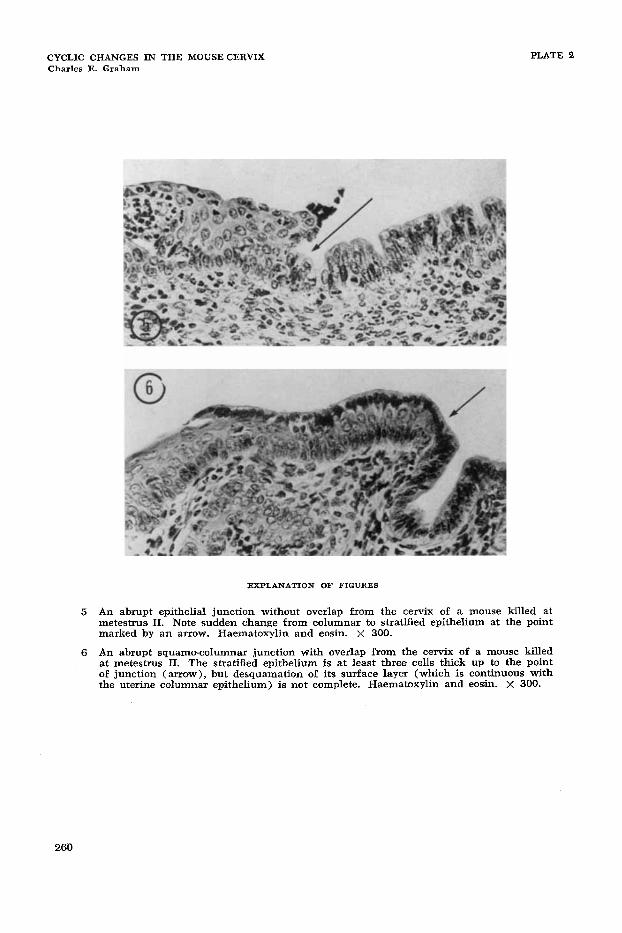
of junction, termed “abrupt,” there was an immediate change from one to several layers of cells (figs. 5 and 6).
In junctions of both types the surface layer of the stratified epithelium close to the junction was sometimes composed of specialized columnar cells, and this layer of cells was always continuous with the uterine columnar epithelium. This con- tinuity of the surface layer of the cervical stratified epithelium with the uterine c01- umnar epithelium will be termed “over- lap.” This term is used descriptively, be- ing in no way intended to imply any par- ticular mode of origin of the appearance. Overlap may be associated with transi-
TABLE 1 Frequency of junction types in the mouse cervix during the estrus cycle, pregnancy,
kctation, exogenous estrogen stimulation, and after ovariectomy
Hormonal status
% of 5% of junctions junctions No. of No. of junctions with
animals examined abrupt overlap
Lkiestrus Proestrus Estrus Metestrus I Metestrus I1 Pregnant, day 7 Pregnant, day 17 7 days postpartum 14 days postpartum Ovariectomized 6 weeks previously Ovariectarnized, estrogen treated Estrogen controls
6 14 10 16
7
102 178 87 155 65 38 32 30 26 28 95 24
3.4 1.7 58 58 97 0
15.6 0 0 0
95 0
13 60
77 23 87
88
3.1 83.5
7.7 100 27 100

253 CYCLIC CHANGES IN THE MOUSE CERVIX
tional junctions (figs. 3 and 4), or with abrupt junctions (fig. 6).
The incidence of the four junction types in relation to the stages of the estrous cycle, pregnancy and lactation, following ovariectomy, and following ovariectomy and injection of estradiol benzoate, is shown in table 1. In the penultimate col- umn of the table the percentage of junc- tions which were abrupt are shown, the remainder being transitional. The per- centage of junctions showing overlap is assessed independently (last column), as this scheme clarifies interpretation of the results.
The stages of the estrous cycle were originally described by Allen ('22). At anestrus the vaginal and cervical epithelium is atrophic, being composed of two or three layers of cells. During the present study the sur- face layer of cells in the vagina was some- times observed to contain small quantities of mucopolysaccharide at anestrus. Dur- ing proestrus proliferation of the stratified epithelium takes place, and in the vagina a layer of very large PAS and alcian blue- positive mucified cells appears at its sur- face. In the endocervical canal this layer becomes composed of flatter cells contain- ing little or no mucin. During estrus, the next stage of the cycle, layers of keratin- ized cells appear beneath the mucified layer, first at the base of the vagina, then progressively towards the cervix. During the following stage of metestrus I, the keratinized layer begins to desquamate. Consequently, the mucified layer is lost at this time; the loss of the mucified layer is a progressive process which begins at the base of the vagina. During metestrus 11, the stratified epithelium is infiltrated by leucocytes, the remainder of keratin is desquamated, and the epithelium begins to regress to the anestrus condition. Dur- ing the estrous cycle and under other con- ditions of hormonal stimulation the cer- vical stratified epithelium reacted simi- larly to the vaginal epithelium, but later and less intensely.
The epithelium that occupies the upper part of the cervix and the uterus is cu- boidal during anestrus. During proestrus it becomes columnar and frequently showed PAS and alcian blue staining in
Junctions during the estrous cycle.
the striated border; at this time prolifera- tion takes place and many uterine glands are formed. During metestrus I1 leuco- cytes infiltrate the epithelium and the epi- thelium begins to regress.
Junctions from animals killed at pro- estrus and early estrus were predominantly transitional, usually with overlap (figs. 2, 3 ) . The increase in number of cell layers between the stratified and colum- nar epithelium was at first very gradual (fig. 3) but as the cycle progressed the cervical stratified epithelium became in- volved in the general vaginal response of proliferation and so became thicker. Con- sequently, the areas of transition became shorter (fig. 4). Indeed a fluent transi- tion between the two types of epithelium became less frequent, and abrupt junctions became more common (fig. 5); the pro- portion of abrupt junctions rose from 1.7% at proestrus to 97% at metestrus I1 (see table 1 ). At the same time junctions with overlap became less frequent.
Overlap was seen in 60% of junctions at proestrus and in 88% at estrus. Near such junctions, the surface layer of the stratified epithelium was composed of a layer of columnar cells that resembled typical uterine epithelial cells; indeed this layer was continuous with the uterine epithelium (fig. 3 ) . Like the uterine cells, the cells forming this surface layer of the stratified epithelium had large oval deeply basophil nuclei, slight cytoplasmic basophilia, and showed PAS and alcian blue staining in the striated border. To- wards the vagina this layer of cells be- came increasingly flattened, lost the mu- copolysaccharide staining reaction and be- came identical with the more flattened cells which formed the surface layer of the stratified epithelium lining the endo- cervical canal; frequently these flattened cells were in turn continuous with the layer of enlarged mucified cells that formed at the surface of the vaginal epithelium dur- ing proestrus.
The surface layer of the vaginal and cervical stratified epithelium was desquam- ated progressively in a cranial direction during metestrus (fig. 4) consequent to the formation of a subjacent layer of keratin, a process which likewise began at the base of the vagina and proceeded

254 CHARLES E. GRAHAM
cranially. In 77% of junctions at metes- trUS 11, keratinization and desquamation reached the junction (which W;LS usually abrupt by this time) so that all the uterine- like cells that had formed the surface layer in this region were lost: thus in these junc- tions overlap was not seen (fig. 5 and table 1, column 5). Occasionally junctions occurred which although abrupt, still showed a small area of overlap (fig. 6 ) .
Junctions in the pregnant and lactating mouse. Cervices were examined from animals killed on the seventh and seven- teenth days of pregnancy, and on the seventh and fourteenth days following de- livery. Almost all of the junctions in these cervices were transitional (see table 1) . Over 80% of the junctions from cervices of the 7-day pregnant and 7-day post- partum mice showed overlap; the appear- ance was similar to that in the cervices at proestrus and early estrus. Overlap was relatively infrequent in cervices at the seventeenth day of pregnancy and 14 days postpartum: typically there was a transi- tional junction without clear relationship of layers.
Junctions in mariectomized animals. In animals killed six weeks after ovariectomy, all the junctions examined were transi- tional with overlap.
Junctions in animals after estrogen treat- ment. To determine whether or not abrupt junctions result from estrogen stim- ulation. three mice ovariectomized one week previously were each given a sub- cutaneous injection of 1.28 vg estradiol benzoate. Three control animals received sesame oil. The animals were killed five days after the injections, and 95% of the junctions in cervices from the estrogen treated mice were abrupt. The cervical epithelium in the animals which received oil was atrophic, and all the junctions were transitional.
DISCUSSION
The ovarian hormones control the ac- tivity of the vaginal and uterine epithelia in rodents and other mammals; estrogens cause proliferation and keratinization in the rodent vagina (Eckstein, '62). The stratified cervical epithelium also is under ovarian control, because it is continuous with the vaginal epithelium and responded
similarly to it during the estrous cycle (though less intensely, as remarked by Allen, '22). The configuration of the junc- tion results in part from changes in the cervical squamous epithelium; the junc- tion itself is therefore under ovarian con- trol. Changes in the columnar epithelium during the cycle did not affect the funda- mental structure of the junction.
Abrupt junctions were predominant dur- ing estrus and metestrus in normal ani- mals. This observation suggested that abrupt junctions are related to stimula- tion by estrogen, because i t is generally agreed that maximal ovarian output of estrogen occurs at estrus (Eckstein, '62), and estrogens cause proliferation of vagi- nal and cervical epithelium. Furthermore, transitional junctions occurred in ovari- ectomized animals, and when ovariecto- mized animals were given estradiol ben- zoate, 95% of the junctions were abrupt five days later.
The cervical epithelium near the junc- tion sometimes bore a layer of uterine-like cells at its surface and this layer was al- ways continuous with the cervical colum- nar epithelium (overlap). The mode of origin of this layer was not immediately obvious. Three possible explanations sug- gest themselves, of which the first two have previously been advanced to account for the origin of similar appearances in the human cervix (review, Fluhmann, '61).
( a ) The columnar epithelium prolifer- ates and grows out over the adjacent stratified epithelium, and/or the stratified epithelium proliferates and burrows be- tween the columnar epithelium and the basement membrane (Novak, '62). If bur- rowing or overgrowth have taken place, then in a horizontal section one would always see a clear demarcation between the two epithelial types at the edge of the overlapping sheet of columnar cells. This arrangement is seen in figure 6, but in this and similar cases it was shown (p. 253) that such appearances resulted from the desquamation of more posterior elements of what was a continuous layer: at proestrus and estrus the uterine-like cells forming the surface layer of the stratified epithelium close to the junction gradually merged caudally with the flat- tened surface layer of the stratified epi-

CYCLIC CHANGES IN THE MOUSE CERVIX 255
thelium which occupied the undivided portion of the cervical canal. Because the continuity of the uterine columnar epi- thelium and the surface layer of the cer- vical stratified epithelium was uninter- rupted, and because this layer could be followed into the cervical canal and some- times even into the vagina, it is concluded that this layer could not have arisen by extension and overgrowth of the uterine columnar epithelium or burrowing of stratified epithelium.
(b) Isolated areas of stratified epithe- lium bearing a surface layer of columnar cells occur in the cervix of women (Car- michael and Jeaffreson, ’41 ; Fluhmann, ’ 5 3 ) . These foci differ in the number of cell layers which they possess, and in the preservation of the columnar surface layer. Fluhmann ( ’53 ) showed that the different types can be arranged in a series which suggests that each is a stage in the trans- formation of columnar epithelium into stratified epithelium. Such transfonna- tions have been termed “squamous meta- plasia” (Willis, ’58). Fluhmann ( ’53) postulated that squamous metaplasia pro- ceeds by the following series of events, and he termed this specific process “proso- plasia.” The columnar cells divide to form a row of small new cells (reserve cells) between themselves and the basement membrane. The reserve cells then pro- liferate and form a stratified layer of from two to eight layers, the original col- umnar cells forming the outer layer. The columnar cells are eventually cast off and the proliferating new cells differentiate into mature squamous epithelium.
The above hypothesis can be extended to account for the origin of the surface layer of columnar cells associated with transitional junctions in the mouse cervix. (Abrupt junctions with overlap need not be considered at this point because it was previously shown on p. 253 how these de- velop from transitional junctions). If one imagines that prosoplasia takes place in the mouse cervix in such a way that the cervical stratified epithelium is progres- sively added to cranially, then one would have to envisage exactly the type of tran- sitional junction under discussion (fig. 3 ) ; this type of junction can be inter- preted as showing a gradation from nor-
mal columnar epithelium through all the stages of prosoplasia involving two, three, four or more layers of cells (the outer layer being the original columnar epithe- lium) and eventually terminating in a normal type of stratified epithelium.
The main weakness of this hypothesis is that there is no evidence that a real extension of cervical stratified epithelium takes place concurrently with the exis- tence of transitional junctions at proes- trus, or in pregnant or lactating animals for example. Further, transitional junc- tions were seen in ovariectomized animals, although metaplasia could hardly be ex- pected in such animals since ovariectomy is known to drastically reduce the rate of cell proliferation in the genital tract (Allen et al., ’24). On the other hand, as a result of the acceleration of cellular proliferation during proestrus and estrus the evidence for metaplasia actually dis- appears, because the area of transition quickly becomes shorter, and abrupt junc- tions without overlap appear.’
(c) The third hypothesis to account for overlap and the transitional junction was evolved during the present study and does not seem to have been previously considered even in relation to the human cervix. It is postulated that the mouse’s cervical epithelium is intrinsically inter- mediate in character. in the sense that its potentialities become progressively more “uterine” cranially; under appropri- ate conditions these potentialities become expressed in the formation of a transi- tional junction in which the two epithe- lial types merge with one another and in which the differentiation of the surface layer of the cervical stratified epithelium becomes progessively more like the typical uterine epithelial cells with which it is continuous. Just as the vaginal stratified epithelium is capable of differentiating to produce either keratinized or mucified cells under appropriate conditions, so un- der comparable conditions the cervical stratified epithelium of the transitional zone is able to produce keratinized cells, or
2 Extension of stratified epithelium does take place under prolonged estrogen stimulation in rodents and primates (Review by Zuckerman, ’40) but under these conditions the junction i s always. abrupt (Graham, in preparation) so that again there is no evidence for the type of metaplasia under discussion, and some other mechanism would seem to be involved.

256 CHARLES E. GRAHAM
columnar cells of more uterine type. Thus the continuity of the surface layer of the cervical stratified epithelium with the uterine columnar epithelium is interpreted as the expression of a continuous gradient of epithelial properties between vagina and uterus. The concept of gradients in the genital tissues has previously been dis- cussed by Zuckennan ('40). Although the cervical stratified epithelium is re- garded as an intermediate type, it is ad- mitted that this characteristic can be ob- scured by the development of an abrupt junction : the differentiation of the transi- tional zone is swung towards the vaginal type by estrogenic stimulation.
The ability of vaginal and cervical strati- fied epithelium to show alternative types of differentiation might be considered to be a type of metaplasia. But the use of this term is avoided in the present context, to emphasize that the postulated dual dif- ferentiative ability of the stratified epithe- lium is quite a different phenomenon from that of prosoplasia, since only the latter process involves a metamorphosis of one epithelial type into another. "Modu- lation" is perhaps a more suitable term.
Now that hormone-controlled cyclic changes have been demonstrated in the mouse cervix, it will be of interest to in- quire if similar changes take place in the human cervix. The mode of origin in the human cervix of areas of epithelium bear- ing a layer of columnar cells also deserves reassessment, since a third hypothesis has now been added to the two classical explanations of the origin of such appear- ances. This latter problem is of consider- able interest; its elucidation would con- tribute to a clarification of the status of metaplasia in the cervix. The possible re- lationship of metaplasia to carcinogenesis has been discussed by Howard, Erickson and Stoddard ('51), Wheeler and Hertig ( '55), Olivi, Barbieri and Paoletti ('62), and Forsberg ( '63) .
ACKNOWLEDGMENTS
This work was carried out in the Depart- ments of Zoology and Comparative Physi- ology, and Obstetrics and Gynaecology, University of Birmingham, England, while supported by a D.S.I.R. Research Student- ship, and in the Department of Anatomy,
Yale University School of Medicine, dur- ing tenure of an Anna Fuller Postdoctoral Research Fellowship. I wish to thank Pro- fessor 0. E. Lowenstein, F.R.Z.S., Profes- sor H. C. McLaren, Dr. J. Cohen and Dr. W. U. Gardner for their advice and en- couragement and for making this work possible.
LITERATURE CITED
Allen, E. 1922 The oestrous cycle in the mouse Am. J. Anat., 30: 297-391.
Allen, E., B. F. Francis, L. L. Robertson, E. C. Colgate, C. G. Johnston, E. A. Doisy, W. B. Kountz and H. V. Gibson 1924 The hormone of the ovarian follicle, its localization and ac- tion in test animals, and additional points bearing upon the internal secretion of the ovary. Am. J. Anat., 34: 133-181.
Carmichael, R., and B. L. Jeaffreson 1941 Squamous metaplasia of the columnar epithe- lium in the human cervical canal. J. Path. Bact.. 49: 63-68.
Eckstein, P. 1962 The Ovary. ed. by S. Zucker- man. Academic Press, New York and London. VoI. I, Chap. VI, 311-359.
Fluhmann, C. F. 1953 The histogenesis of squamous cell metaplasia of the cervix uteri and endometrium. Am. J. Obstet. Gynec., 97:
-- 1959 The squamo-columnar transition zone of the cervix uteri. Obstet. and Gynec., 14: 133-142. - 1960 Carcinoma in situ and the transi-
tional zone of the cervix uteri. Obstet. and Gynec., 16: 424-437. - 1961 The Cervix Uteri and its Dis-
eases. W. B. Saunders Company, Philadelphia and London. Chap. 111, 46-102.
Forsberg, J. G. 1963 Embryological aspects on the origin of atypical epithelia in the cervix uteri. K. frsiogr. Sallsk. Lund Forh., 33: 39- 43.
Howard, L., C. C. Erickson and L. D. Stoddard 1951 A study of the incidence and histo- genesis of endocervical metaplasia and intra- epithelial carcinoma. Cancer, 4: 1210-1223.
Novak, E. R. 1952 Gynaecologic and Obstetric Pathology. Third edition. W. B. Saunders Company, Philadelphia and London. Chap. V, 70-89.
Papanicolaou, G. N. 1942 A new procedure for staining vaginal smears. Science, 95: 384.
Pearse, A. G. 1962 Histochemistry; theoreti- cal and applied. Little, Brown and Company, Boston. Second edition. appendix 9, pp. 831- 843.
Pilgrim, M. I., and K. B. DeOme 1955 Intra- peritoneal pentobarbital anesthesia in mice. Exp. Med. Surg., 23: 401-403.
Olivi, M., G. Barbieri and I. Paoletti 1962 Changes of uterus epithelium induced in mice by 20-rnethylcholanthrene. The Morphological Precursors of Cancer: Proceedings of Interna-
45-58.

CYCLIC CHANGES IN THE MOUSE CERVIX 257
tional Conference, ed. by L. Severi. Perugia: Willis, R. A. 1958 The Borderland of Embry- 467-477. ology and Pathology. Chap. XIV, 506-570.
Wheeler, J. D., and A. T. Hertig 1955 The Zuckerman, S. 1940 The histogenesis of tis- pathologic anatomy of carcinoma of the uterus. sues sensitive to oestrogens. Biol. Rev. 15: 1. Squamous carcinoma of the cervix. Am. J. 231-271. Clin. Path., 25: 345-375.

PLATE 1
EXPLANATION OF FIGURES
2 A transitional junction from the cervix of a mouse killed at anestrus. There is no clear relationship of particular cell layers between the stratified epithelium at left, and the columnar epithelium to the right. Haematoxylin and eosin. x 300.
3 Cervix from mouse killed during early estrus. At X a region of transition between columnar and stratified epithelium begins: note gradual increase in number of cell layers. The surface layer of cells of the stratified epithelium at Y is composed of cells morphologically similar to, and continuous with the cells of the columnar epithelium (overlap). Haematoxylin and eosin. X 180.
A transitional junction with overlap from the cervix of an animal killed at late estrus. The stratified epithelium is much thicker than at early estrus. At the point of junction ( X ) the columnar epi- thelium is continuous with the surface layer of the stratified epi- thelium. Keratin ( K ) has formed beneath the surface layer of the stratified epithelium. and in this region the surface cells are desqua- mating. Haematoxylin and eosin. x 300.
4
258
CYCLIC CHANGES IN THE MOUSE CERVIX Charles E. Graham
PLATE 1
259
CYCLIC CHANGES IN THE MOUSE CERVIX Charles E. Graham
PLATE 2
EXPLANATION OF FIGURES
5 An abrupt epithelial junction without overlap from the cervix of a mouse killed at metestrus, 11. Note sudden change from columnar to stratified epithelium at the point marked by an arrow, Haematoxylin and eosin. X 300.
An abrupt squamo-columnar junction with overlap from the cervix of a mouse killed at metestrus 11. The stratified epithelium is at least three cells thick up to the point of junction (arrow), but desquamation of its surface layer (which is continuous with the uterine columnar epithelium) is not complete. Haematoxylin and wsin. X 300.
6
260

















![ResearchEarly clinical experience of radiotherapy of ... · anal canal, cervix uteri cancer and other indications [3-9]. The potential role of RA in the treatment of prostate can-cer](https://img.pdfslide.us/doc/110x75/5fc0d7aef0fedd574d35b5b4/researchearly-clinical-experience-of-radiotherapy-of-anal-canal-cervix-uteri.jpg)











