Embed Size (px)
Citation preview

THE JOURNALS OF BIOLOGICAL CHE~~ISTRY Vol. 244, No. 15, Issue of August 10, pp. 4089-4094, 1969
Printed in U.S.A.
Cyanide Metabollism by Bacillus megaterium*
(Received for publication, January 24, 1969)
PETER A. CASTRIC$ AND GARY A. STROBEL!
From the Department of Botany and Microbiology, Montana State University, Boxeman, Montana 59715
SUMMARY
Cyanide disappears from a medium supporting actively growing cells of Bacillus megaterium, a bacterium isolated from a cyanide enrichment medium containing Fargo clay soil. Whole cells of this organism convert K14CN to aspara- gine-*4C, aspartic acid-14C and 14C02. Besides cyanide, serine l4C and /3-cyanoalanine-14C are effectively converted by whole cells of this organism to asparagine-14C. An analy- sis of asparagine isolated in an experiment in which whole cells were administered K13C15N and serine-14C shows that the nitrogen and carbon of the amide group is derived from cya- nide. /3-Cyanoalanine-14C was isolated and identified as a product of this experiment. These results as well as results of analyses of the carbon skeletons of P-cyanoalanine-14C and asparagine-14C from this and other experiments collec- tively support the hypothesis that a metabolic pathway in this organism begins with the condensation of serine and cyanide to form P-cyanoalanine which in turn is hydrolyzed to aspara- gine and then to aspartic acid. Furthermore cell-free ex- tracts catalyze the conversion of K14CN and serine to aspara- gine-1% and aspartic acid-14C as well as the conversion of asparagine- 14C to aspartic acid-14C.
Cyanide assimilation occurs in higher plants (1,2), fungi (3,4); algae (2), and bacteria (5, 6). In all cases the first product of metabolism is a nitrile. In higher plants and algae the first in- termediate is P-cyanoalanine (2) which in some plants combines with a glutamyl donor to form the dipeptide y-glutamyl-fl-cy- anoalanine (2, 7). In other plants P-cyanoalanine is hydrolyzed to asparagine (2, 7). Isotope studies indicate that the cyanide carbon becomes the C-4 of the fl-cyanoalanine of the dipeptide (7), and the cyanide carbon and nitrogen become the amide car- bon and nitrogen of asparagine (8). Studies in vitro indicate
* This study was supported in part, by Grant GB-6395 from the National Science Foundation, National Instititutes of Health Training Grant AI-00131-07 from the National Institute of Allergy and Infectious Diseases. Montana Agricultural Experiment Sta- tion Journal Series Paper 953.
$ A portion of a Ph.D. thesis submitted to the Graduate School of Montana State University. Present address, the Department of Biochemistry and Biophysics, University of California, Davis, California 95616.
$ Public Health Service Research Career Development Award 1-K 4-GM-42,475.01 from the National Institute of General Medi- cal Sciences.
that, cyanide condenses with serine (9, 10) or cysteine (8, 9) to form P-cyanoalanine. Hendrickson (11) has purified the en- zyme P-cyanoalanine synthase several hundred-fold from blue lupine which condenses cysteine, and not serine, with cyanide forming /3-cyanoaline. Further aspects of cyanide metabolism in plants are discussed in a review by Conn and Butler (12).
There is a paucity of information on the assimilation of cyanide in bacteria. Studies in vivo by Brysk, Corpe, and Hankes (5) show that Chromobacterium violaceum forms P-cyanoalanine and aspartic acid from cyanide. The 3-carbon precursor was postu- lated to be serine. Dunnill and Fowden (6) have obtained an enzyme preparation from Escherichia coli which forms fi-cyano- alanine from cyanide and serine. The specificity of the a-carbon precursor to fi-cyanoalanine is uncertain since serine appeared to serve as a substrate, however, a competitive effect is noted with cysteine.
This report presents evidence for the formation in vivo and in
vitro of asparagine and aspartic acid from cyanide and serine by Bacillus megaterium as shown in Fig. 1. To the authors’ knowl- edge this is the first report to provide conclusive evidence for this pathway in a bacterium.
EXPERIMENTAL PROCEDURE
Culturing-The culture of B. megaterium used in this study was obtained from a 10-l of KCN enrichment broth of Skow- ronski snd Strobe1 (13) which had been incubated with Fargo clay soil. The morphological and physiological criteria of Breed, Murray, and Smith (14) were used for the taxonomic description of this bacterium.
Cells were grown under aerobic conditions at 35” in Trypticase soy broth with 1% glucose containing 1 mM KCN. Growth appeared in this medium after incubation for 10 to 12 hours while the appearance of growth in this medium minus cyanide was at 3 to 4 hours. Cells mere harvested after 24-hour incuba- tion by centrifugation at 20,000 x g at 4” for 10 min, then rinsed twice and resuspended in distilled water, Trypticase soy broth, or buffer.
Materials
The 1% compounds used in this study were obtained from the following sources: KCN (45.2 mCi per mmole), L-serineJ4C (14.5 mCi per mmole), sodium formate (0.96 mCi per mmole), Nu- clear-Chicago; DL-cysteine-3-l% (25.0 mCi per mmole). The Radiochemical Centre, Amersham, England; acetyl-CoA-1-W (46.2 mCi per mmole), sodium pyruvate-2-W (3.16 mCi per mmole), New England Nuclear; sodium carbonate (10 mCi per mmole), Tracerlab; sodium acetate-lJ4C (48.9 mCi per mmole),
4089
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

4090 Cyanide Metahlisrn by Bacillus ,mcgaterium Vol. 244, So. 13
COOH COOH COOH COOH
Swine p Cyanoalonine Asparagine Asparlic Acid
FIG. 1. A pathway of aspnrtic acid biosynthesis in B. nrega- lerium.
Volk Radiochemical Company; glycolic aldehyde-14C (0.74 pCi/mmole) was prepared by the decarbosylation and deam- ination of L-scrine-14C by a modification of the method of Dakin (15) ; formamide (0.977 PCi per mmole) was prepared by heating ammonium formate-14C at 120” overnight in a scaled bube with subsequent dist,illation and recovery of the formamide; asparagine&% (0.3 /.&i per mmole) was obtained from cultures of B. megaterium that were grown in the presence of KWX. Ethanolamine-W (13.4 mCi per mmole) and L-P-cyanoalanine- 4-W (10 mCi per mmole) were a generous gift of E. E. Conn, University of California, Davis. In addition to these I46 com- pounds, KWN and KC% were purchased from Bio-Rad Laboratories.
Hyamine hydroxide 10X was purchased from Packard Instru- ment Company and NCS protein solubilizer was purchased from Nuclear-Chicago. hspartic decarbosylase was purchased from Sigma. All other chemicals were reagent grade.
-tnalytical Technipues-The technique of Aldridge (16) was used for the quantitative determination of cyanide. Protein concentration was determined by the method of Lowry et al. (17).
The reaction of P-cyanoalanine with ninhydrin was performed by the method of Ressler (18) and its absorption measured in a Beckman DK-2 scanning spectrophotometer. The infrared spectrum of P-cyanoalanine was measured with a Beckman Microspec infrared spcctrophotometcr utilizing KBr micropel- lets.
The Y-bromosuccinimide decarbosylation technique n-as performed according to the method of Blumenthal-Goldschmidt, Butler, and Conn (1). Decarbosylation with aspartic decnr- boxylase was carried out in a gas evolution vessel with 5 s ?;aOH as trapping agent.
Ammonia was determined by the micro-Kjeldahl method. Dilute sulfuric acid was added to the boric acid trap after titra- tion with 0.1 N HCI and the solution was taken to dryness in a 60” oven.
ChronLatography-Sheets of JYhatman ?;o. 541 paper were used for paper chromatography and the following solvent, sys- tems were employed: (a) n-butyl alcohol-acetic acid-water (4: 1:5, v/v/v) ; (b) methanol-pyridine-water (40:2: 10, v/v/v) ; (r) 80% phenol in water. Plain silica gel plates and Eastman Chromatograms were used for one- and two-dimensional thin layer chromatography, respectively. The following solvent sgst,ems were employed for thin layer chromatography; (d) n-butyl alcohol-acetic acid-water (3 : 1: 1) ; (e) isopropyl alcohol- ammonium hydroxide (67 :33). Xmino acids were detected b) spraying the chromatograms with 0.03y0 ethanolic ninhydrin. Radioactivity was detected on two-dimensional thin layer chromatograms with the x-ray film orerlay method used by Strobe1 (3). Radioactivity was also detected on one-dimensional thin layer and paper chromat,ograms lvith a Packard Radio- chromatogram Scanner model 385.
Paper Eleclrophoresis-Sheets of 1Thatman Xo. 311&I mere
used for paper electrophoretiis. The following buffers were used: (n) Pyridine-acetate, pH 5.2 (25; pyridine in 1% aqueous acetic acid), 16 volts per cm, and (b) sodium borate, pH 8.8, 0.05 M, 17 volts per cm. Radioactivity and amino acids were de- tected as indicated previously.
Isofope Determinations---Radio:l~tivit)- was measured quanti- tatively with a l\‘uclear-Chicago scintillation counter model 6804. The scintillation fluid and the method of counting Jvere as indicated by Strobe1 (3). 13C and l5IY enrichments were determined on the Picker-SE1 hlS-10 mass spectrometer. 15S and 13C determinations were performed as indicated by Strobe1
(3). ;Idministrution of Labeled Compounds-Equivalent amounts
of labeled compounds were administered as aqueous solutions to equivalent amounts of bot,h live and boiled cells. Vessels with a side arm or a center well containing 5 x r\‘aOH were used when CO2 was measured. Cnless otherwine indicated, the period of incubation w~as for 24 hours at 23”.
Extmction of Cell T’roducts-Cells were disrupted using a French pressure cell or Braun l\ISK cell homogenizer. Either method resulted in virtually total cell disruption. TWO volumes of 95@/, ethanol were added to the homogenate and the suspension was centrifuged at 20,000 x g at 4” for 10 min. Where indicated the ethanol-insoluble precipitate was dried and resuspended in NC’S protein solubilizer, added to the scintillation fluid in place of Incthanol, and the radioactivity ~-\-as counted in the usual fashion. The supernatant solution WIS passed through cohnnns (I x 2 cm) of Dowex SOW-X8 (H+), 200 to 400 mesh, then 1~0~~s 1-X8 (format?), 200 to 400 mesh. The cation fraction was &ted with 10 ml of 6 s H(I1 and the anion fraction was eluted with 10 ml of 6 N formic arid. hll fractioiis including the neutral fraction were taken to dryness on n hot plate in a stwam of air and stored overnight in a vacuum desiccator containing I’,O, and XaOH.
Carbon dioside formed on osidation of the labeled substrate was trapped as carbonate and precipitated with 0.3 111 Bu(OH)z. The 1)recipitatc was washed twice with distilled water by cen- trifugation at 10,000 X g at 4” for 10 min. Radioactivity was detcrniined by the conversion of a I)ortion of the BaCOs to CO? (by the addition of 0.5 ml of 1 N II$o,) in a gas-trapping vessel containing 1.5 ml of hydroxide of Hyamine in a side arm. The hydroxide of Hyamine was then added to the scintillation fluid in place of methanol.
F’urificniion and &ua,nfi$cafion of -1 sparagi?le-W-The catior fraction was further separated by paper chromatography ir Solvent b. The area mat,ching the area containing authentic asparagine was cut out, and eluted with distilled wat,er. Thi: material \vas dried in a stream of air and chromatographed il Solvent a. This procedure was repeated once with the aspara gine isolated from the triple labeling experiment. &C+anoala, nine n-as isolated by these methods except that it was also chro. rnatographed in Solvent c.
Asparagine was quantitatively determined with the use of thr reagents described in the Technicon Research Bulletin ?r’o. 2( (19) by incubation of a portion of the isolated asparagine wit1 the reaction mixture at 100” for t5 min. Optical density of the mixture was nleasured at 5TO nip. Radioactivity of a portiol of the asparagine was counted yielding specific activity value for the isolated asparagine.
Preparation oj Cell-jree Extracts-Kaahed cells resuspended il appropriate buffers were disrupted in a Braun MSK cell homage
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Issue of August 10, 1969 P. A. Gastric and G. A. Strobe1 4091
nizer and centrifuged twice at 20,000 x g for 10 min at 4”. Boiled (100” for 10 min) and nonboiled portions of the superna- tant solution were incubated with equivalent amounts of 1% compounds.
RESULTS
Uptake of Cyanide by Whole Cells-The first indication that cyanide was assimilated by B. megaterium was the disappearance of cyanide from a medium containing a growing culture of the organism. Fig. 2 shows that bacterial growth coincides with the disappearance of cyanide.
To establish that cyanide was being metabolized by this organism, 4.13 $Zi of K14CN (45.2 PCi per pmole) were admin- istered to washed cells (previously grown in 1 m&l KCN and the equivalent of 125 mg, dry weight) suspended in 20 ml of sterile distilled water. Xfter 24 hours of incubation the cells were harvested, the CO* evolved, and the ethanol-soluble compounds of the cation, anion, and neutral fractions were examined. The radioactivity was incorporated only into t’he cation fraction (1.4 PCi of above boiled control) and COz (7.2 m&i of above boiled control). The examination of the cation fraction by paper and thin layer chromatography and by paper electro- phoresis indicated that two compounds (designated as Unknowns 1 and 2) contained nearly all of the radioactivity. The Rp
values of Unknowns 1 and 2 matched the RF values of asparagine and aspartic acid, respectively, in one-dimensional paper and thin layer chromatography using Solvents a through e. Utiliz- ing paper electrophoresis and Buffers a and b, Unknown 1 and asparagine had the same rates of migration. hs a further sub- stantiation of its identity, Unknown 1 was refluxed for 14 hours in 6 N HCl then dried in a stream of air on a hot plate and stored overnight in a vacuum desiccator containing PZOS and NaOH. The product was then identified by paper and thin layer chro- matography in Solvents a through e. The RF of the radioactive product matched that of authentic aspartic acid in all cases. Further evidence for the identity of Unknowns 1 and 2 and the acid hydrolysis product of Unknown 1 was obtained using two- dimensional thin layer cochromatography (Solvents d and e) using authentic compounds with subsequent s-ray film overlay.
3’o!l.\ ------I:~ . ..I y\-. I
x A.300
Mg KCN I A
Time in hours
FIG. 2. Cyanide disappearance with growth of B. megaterium as a function of time. A-A, sterile 1 mM cyanide medium; X-X, 1 mM cyanide medium inoculated with B. megalerium; O- - -0, growth of B. megaterium in 1 mM cyanide medium. Cy- anide concentration is expressed as the total weight (mil- ligrams of KCN) in the 50 ml of growth medium that was used. Growth of bacteria is expressed as absorbance at 660 mp.
TABLE I
“C labeling patterns of asparagine and j3-cyanoalanine isolated from whole cell suspensions of B. megaterium which had been admin-
istered various ‘V compounds
Exper imenl
Radioactive compound degraded
~-- WC % % %
Asparagine-‘4C 4.3-8.4 K WN 1.7 4.2 94.1 Asparagine-“C 0.3-1.4 K14CN 2.9 0.2 96.7 Asparagine-‘4C 0.6-2.3 SerineJW 32.0 65.4 2.6 AsparagineJ4C 0.1-0.3 B-Cyanoala- 2.0 4.0 94.0
nine-4-14C p-Cyanoala- 1.3-4.5 Serine-“Cd 35.0 62.7 2.3
nineJ4C
4m*yd?J-of activity
sed in deg radations
Radioactive prec”rSOr
Total radioactivity per carbon atom
C-l” C-4=
(i The radioactivity present in the C-l of asparagine and p-cy- anoalanine was the 14COs evolved with the use of the N-bromo- succinimide technique.
*The radioactivity present in the C-2 and C-3 positions of asparagine and p-cyanoalanine was considered to be the radio- activity not accounted for by the C-l and C-4 determinations.
c The radioactivity present in the C-l and C-4 position of asparagine and p-cyanoalanine was considered to be the ‘4CO2 evolved from aspartic acid (the acid hydrolysis product of these compounds) with the N-bromosuccinimide technique. The per- centage of radioactivity in excess of the C-l determination (Fool- note a) was the C-4 radioactivity value. Also, the C-4 radio- activity of asparagine and p-cyanoalanine was measured as the 14C02 evolved from their acid hydrolysis product aspartic acid, by using the aspartic acid decarboxylase technique. The percent- ages given are averages of the values obtained by these two tech- niques.
d Uniformly labeled.
The exposed spots on the films coincided exactly to the reference compounds. Experiment 1 in Table I shows that nearly all of the radioactivity resided in t.he amide carbon of asparagine. The aspartic acid formed by acid hydrolysis of the radioactive asparagine was at least 96% in the L configuration since it reacted to t’his extent with the L-specific aspartic acid decarboxylase.
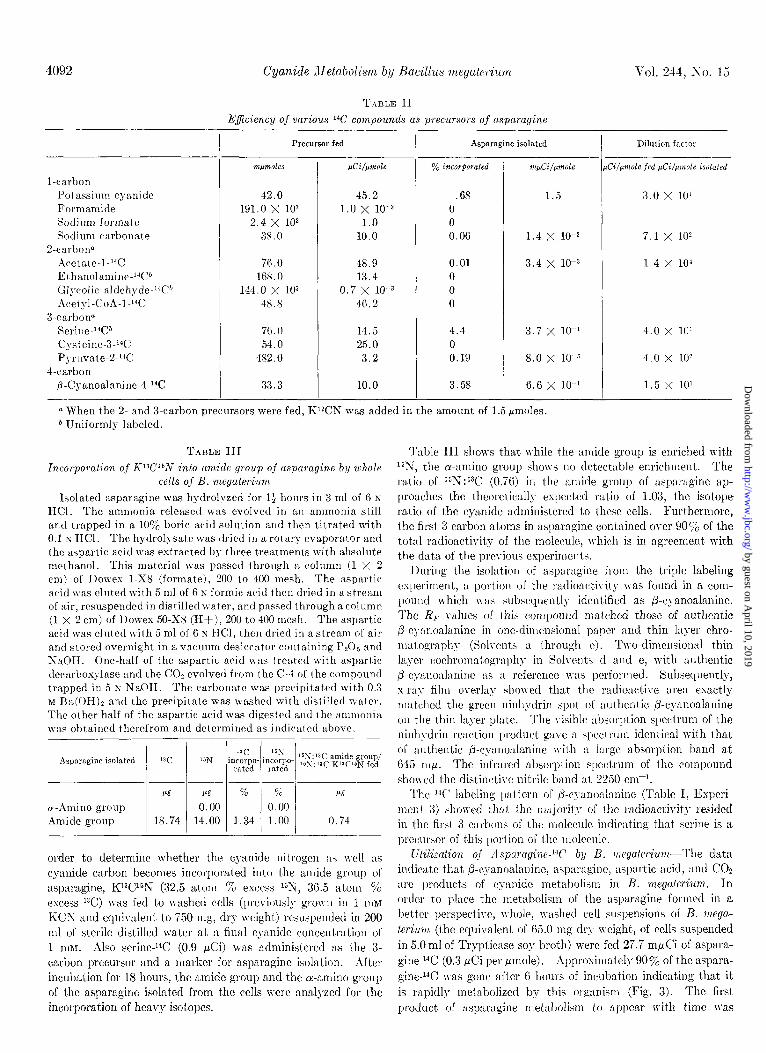
Screening of 14C Precursors of AsparagineSince the cyanide carbon appeared to be going into the amide group of asparagine, it was necessary to determine: (a) what precursors might provide the remainder of the carbon skeleton of asparagine, and (6) what immediate precursor besides cyanide could provide the amide carbon of asparagine. Washed cells (the equivalent. of 125 mg dry weight) suspended in 20 ml of sterile distilled water were fed quantities of 14C compounds thought to be likely precursors. The percentage of incorporation of t’he precursors into aspara- gine and the specific activity of the asparagine isolated as well as the dilution factors of these experiments are cited in Table II. These results show that cyanide-%, serine-‘4C, and P-cyanoala- nine-4-14C were the best precursors to asparagine synthesis as their percentage of incorporation was highest and their dilution factors were lowest. The patterns of lGlabeling in the aspara- gine derived from KCN, serine, or ,&cyanoalanine are given in Experiment 2 in Table I. The results indicate that serine pro- vided the best a-carbon precursor and cyanide, the best l-carbon precursor for asparagine biosynthesis.
Incorporation of KY?5N into Amide Group of Asparagine--In
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
4092 Cyanide ;Iletabolism by l3acillus megatwiwn
T.~BLE II Ejiciency of various 14C compounds as precursors of asparagine
Vol. 244, so. 15
l-carbon Potassi~nn cyanide Formamide Sodiwn formate Sodium carbonate
2-carbolln Acetate-l-14C ~tharlolarnillr,-‘“C” <;lycolic nldehvde-‘“C* Acely1-CoA-1-;4C
3-rarbon” Serine-14C1 Qateine-3-“C Pvrllvate-2V4C
J-carbon fi-Cvanoalanine-4-14C
Precursor fed
?npmoles pCi/pmole I- 42.0 45.2 .68
191.0 x 103 1.0 x 10-S 0 2.4 x 103 1.0 0
38.0 10.0 0.06
7G.0 48.9 16S.O 13.4
144.0 x 103 0.7 x IO-3 48.8 46.2
70.0 14.5 54.0 25.0
482.0 3.2
33.3 10.0
0.01 0 0 0
4.4 0 0.19
3.5s
Asparagine isolated Dilution factor
-
VLpCi/p?l&
1.5
1.4 x 10-Z
3.4 x 10-s
3.7 x 10-l
8.0 x 10-Z
6.6 x 10-l
a When the Z- and S-carbon precursors were fed, Kl*CN was added in the amount of 1.5 rmoles. * Uniformly labeled.
TABLE III Incorporation of K13C15A* into awlide group of usparagine by whole
cells of B. w~egaterium
Isolated asparngine was hydrolyzed for 16 hollrs in 3 ml of 6 in HCl. The ammonia released \v:ts evolved in an ammonia still and trapped in a 10% boric acid solution and then titrated with 0.1 s HCl. The hydrolysate was dried in a rotark- evaporator and the aspartic arid \vas extracted by three treatments with absolllte methanol. This material was passed through a rolumn (1 X 2 cm) of Dowes l-X8 (formate), 200 to 400 mesh. The aspartic acid \vas elllted with 5 ml of 6 x formic acid i hen dried in a stream of air, resuspended in distilled water, and passed through acolnmn (1 X 2 cm) of 1)owex 50-X8 (Hi), 200 to 400 mesh. The aspartic acid was ellltetl \vith 5 ml of 6 h’ IICl, then dried in astream of air and stored overnight in a vacnllm desiccator containing PeOs and Tu’aOH. One-half of the aspartic arid \vas treated with aspartic decnrboxylasc and the CO, evolved from the C-4 of the compound trapped in 5 N NaOH. The rarbonatc was precipitated with 0.3 M Ba(OII)? and the precipitate was washed with distilled water. The other half of the aspartic acid was digesied and the ammonia was obtained therefrom and determined as indicatrtl above
a-Amino group Amide group
The I”(: labeling pattern of p-c~unonl:rnine (Table I, Experi- mcnt 3) showed that the majorii~ of the rndioactivit!- resided in the first 3 carbons of the molecule indicating that scrine is a precursor of this portion of t,lle nlolecwle.
order to determine whether the cyanide nitrogen :M well as Utilisnfion of A spnmyine-1% by B. rtiegateriuw-The data
cyanide carbon becomes incorporated into the amide group of indicate that fi-cyanonlanine, asparaginc, aspartic acid, and COP
aspnragine, K1Q5r\’ (32.5 at,om % excess ‘“lur, 36.5 atone % arc lwoducts of cyanide metabolism in B. megaterium. In
CSC~SS W) was fed to washed cells (previously grown in 1 INM order to lplacc the metabolism of the asparagine formed in a
KCS and equiralent to ‘750 mg, dry n-eight) resuspended in 200 better perspectirc, whole, 17.ashed cell suspensions of B. wega-
ml of sterile distilled water at :L final c)-snide concent,r:ttion of teriuw (the equiralent of 65.0 mg dr>- n-eight, of cells suspended
1 m&r. Also serine-14C (0.9 PCi) was administered as the 3- in 5.0 ml of Trypticase soy broth) were fed 27.7 mpCi of aspara-
carbon precursor and a marker for asparagine isolation. After gine-1% (0.3 MCi per pmole). Approximately 90% of the aspara- incubation for 18 hours, the amide group and the cr-amino group gine-1% xas gone after 6 horns of incubation indicating that it of the asparagine isolated from the cells were analyzed for the is rapidly metabolized by this organism (Fig. 3). The first incorporation of heavy isotopes. product of aspnwgine metabolism to appear trith time was
3.0 x 10’
7.1 x 102
1.4 x 10’
4.0 x lt:’
4.0 x 10’
1.5 x 10’
Table III shows that while the amide group is enriched with lslri, the a-amino group show no detectable enrichment. The ratio of 15N:13C (0.76) iu the amide group of nsparngine ap- proaches the theoretiwllg expected ratio of 1.03, the isotope ratio of the cyanide administered to these cells. Furthermore, the first 3 carbon atoms in asparngine contained over 90% of the total radioactivity of the molecule, which is in agreement with the data of the previous experiments.
During the isolation of asparaginc from the triple labeling csl)eriment, a portion of the radioactivity was found in a com- pound Tvhich was subsequently identified as p-cJ-anoalanine. ‘l’hc RF values of this c~ompound matched those of authentic p-c)-nnoalanine in one-dimensional paper and thin la!-er chro- matography (Solvents a through c). Two-dimensional thin layer cochrolnatogl.nl,h?- in Solvcntv d and e, with authentic &c~auonlanine as :L reference wls performed. Subsequently, x-ra\- film overlay showed that the radioactive are:t exactly matched the green ninh~drin +ot of authentic /?-cynnoalanine on the thin layer plate. The visible ab>orption spectrum of the niuh\-drin reaction product gaye :t spectrum identical with that of authentic p-cyanoalanine Jyith :L large absorption band at 6-15 ml.~. The infrwed absorption spectrum of the compound showed the distinctive nitrile band ai. 2230 cn-I.
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
aspartic acid which was identified by paper and thin layer chromatography in Solvents b, c, and d. The aspartic acid then appears to be metabolized to COZ and ethanol-insoluble material (the composition of which is unknown).
CeZZ;free Eqeriments-Table IV shows the cell-free synthesis of asparagine from KCN and serine. Under the conditions of assay all values reported are above the values of the boiled con- trol. Asparagine was identified by chromatography of portions of the reaction mixture in Solvents a and c. The radioactive
2.5-
2.0 - dm 164
1.5 -
Time in hours
FIG 3. Metabolism of asparagine-4-W in B. megaterium as a function of time. A, asparagine-14C; X, aspartic acid-W; 0, WOZ; 0, W-ethanol-insoluble material.
TABLE IV Conversion of serine-14C and K’TN to asparagine-W’ by cell-free
extracts of B. megaterium
The reaction mixture contained 0.3 mmole of sodium carbonate buffer (pH 9.5), 0.3 Imole of mercaptoethanol, 5.0 pmoles of KCN (when fed as radioactive isotope, 4.95 PC of KWN added for an over-all specific activity of 0.97 pCi per pmole), 10.0 pmoles of nL-serine (when fed as a radioactive isotope, 0.76 pCi of L-serine- 1% added for an over-all specific activity of 0.15 pCi per Hmole of L-serine), 0.4 mg of protein, and distilled water in a total volume of 3.0 ml. Incubation was carried out at 23” and the reaction was stopped by the addition of 2 ml of 95% ethanol to samples of the reaction mixture which were then dried on a hot plate in a stream of air. The samples were assayed using paper chromatography (Solvents a and c). Areas matching the reference were cut out, and added to scintillation vials, and the radioactivity was counted.
PreCUWX
Cyanide and not one of its hydrolysis or oxidation products was the primary l-carbon precursor of asparagine (Table II). Factors that would contribute to the percentage of incorporation and dilution factors of KCN, serine, and P-cyanoalanine as being precursors to asparagine and aspartic acid are (a) proximity of the compound to the final product, and (b) participation of the compound in other enzymatic reactions.
Serine-“C + K’2CN
Serine-1% + K’4CN
1 Time of incubation 1 W-Asparagine isolated
hrs
0 0.00 6 4.6
16 3.6 0 0.00 6 5.0
16 7.0 of the asparagine to aspartic acid in which the amide nitrogen is
The ratio of 16N:laC of the amide group of asparagine isolated from the IQaC?6N and serine-W feeding of whole cells to 16N :13C
of the cyanide fed indicates that the TJ of cyanide appears to be going into the amide group of asparagine to a slightly greater extent than the ISN of cyanide. This discrepancy could be a result of (a) experimental error or (b) the hydrolysis of a portion
Issue of August 10, 1969 P. A. Cast& and G. A. Strobe1 4093
TABLE V
Conversion of asparagine-“C to aspartic acid-W by cell-free extract
The reaction mixture contained 2.5 pmoles of potassium phos- phate buffer (pH 7.2), 11.0 rr$Zi of asparagine-W (36.5 mpmoles), and 16.4 mg of protein in a total volume of 2.0 ml of distilled water. Incubation was carried out at 23”; the reaction was stopped by passing portions of the mixture through columns (0.5 >(: 2 cm) of Dowex l-X8 (formate, 200 to 400 mesh). After elution of the column with 3 ml of 6 N formic acid, both fractions were dried on a hot plate under a stream of air. Portions of these samples were assayed with the use of thin layer chromatography Solvent d. The area matching the reference was scraped into scintillation vials and the radioactivity counted.
Time
hrs
0 3 6
Asparagine
dpm
6100 3600 2500
Aspartic acid
dfim
0 1250 2000
product chromatographed at the same RF as authentic aspara- gine in each case. No radioactive asparagine could be demon- strated when these extracts were fed cysteine-14C and KCN as precursors. Furthermore, the presence of &cyanoalanine could not be shown in any of the reaction mixtures. Radioactive aspartic acid was observed as a product when either KWN or serine-‘4C were used as radioactive precursors. The aspartic acid of the reaction mixture was isolated and assayed according to the procedure in Table V.
The demonstration of the cell free conversion of asparagine to aspartic acid is presented in Table V. Under the conditions of assay noted in Table V, the disappearance of substrate and ap- pearance of product were linear with relation to time and protein concentration. The product of the reaction was identified as aspartic acid by means of two-dimensional thin layer chroma- tography with authentic aspartic acid (Solvents d and e) fol- lowed by x-ray film overlay of the thin layer plate. The exposed spot on the film matched exactly the ninhydrin positive spot on the plate.
DlSCUSSION
The disappearance of cyanide from a growing culture of B.
megaterium (Fig. 2) was the first indication that this organism might metabolize this toxic compound. The administration of Kl4CN to a washed whole cell suspension of this organism showed that radioactivity appeared in asparagine, aspartic acid, and carbonate after incubation of the cell suspension.
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Cyanide Xetabolism by Bacillus megateriwn Vol. 244, No. 15
converted to ammonia and the reverse step of asparagine syn- thesis (reported in bact,eria, see References 20 and 21) from aspartic acid and ammonia slightly, if at all, enriched with 1%.
The question of why p-cyanoalanine was isolated during the Kr3C15S and serine-14C feeding esperiment~ and iu no other experiments deserves comment. 1Vhile Kr4CS nas ordinarily fed in the concentration of 0.1 to 1.0 m~mole lwr rllg of cells, dry weight, the IM?S was administered in the concentration of -MO m~moles per mg of cells, dry weight. -1ssuming that seriue would be generated by t,he cell, the cscess of KC?; in the nredium contributed to a build up of p-cyanoalanine.
The results of the cell-free experiments (Table IV) iho\v that in aliquots of the same extract, Kr4Cn’ and serine were converted to asparaginc to the same extent as KCX and serinem14C’ indicat- ing a single pathway. P-Cya 71 no&nine could not be shown in the reaction mixtures. Either one or both of the following possibilities could give the result,s seen. (n) A\t the times of assay the enzyme of condensatiou had become denatured and the @-cyanoalanine formed converted to asparagine and aspartic acid, or (b) nitrilase activity, necessary to convert p-cyanoala- nine to asparagine, is of the magnitude which does not allow the :mcumulation of p-cyanoalanine under the usual experimental conditions.
The ,significance of the assimilation of cyanide by B. Twge- teriwn can be questioned. This organism, isolated from a soil whicll had supported the growth of flax (a cyanophoric plant) for 52 years, appeared to adapt its respiration to the presence of cyanide’ and converted this toxic molecule into common metabo- lites, indicating a detoxificat,ion process. Cyanide has been postulated to be aa intermediate in the prebiotic synthesis of such nrolecules such as purinr, and amino acids such as aspara- gine and aspartic acid (22). The metabolic activity noted in the llresent report, might have been acquired early in evolution and retained up to the present.
Reports hare indicated that asparagine in Lactobacillw rrrabinosus (21) and Streptococcus bovis (20) is formed from as- partic acid and ammonia by an enzyme termed asparagine syn- thetase. The pathway shown in this report (Fig. 1) could well be a natural mechanism of asparagine synthesis in B. megaterium. Although an asparagine pool is present in this organism when no
1 Unpublished results.
cyanide is administered to it, the possibility exists that cyanide could be evolved by this organism which could condense with serine, ultimntel~- forming asparagine. Precedence for this is the formation of c!-snide from glycine in C. violuceum (23). This point warrants further study.
1.
2. 3.
4. 5.
6. 7.
8.
9.
10.
11. 12.
13.
14.
15. 16. 17.
18. 19.
20.
21.
22.
REFERENCES
HI,~~ESTH.\I,-(;~LI~SCIIJIII)T, S., BUTLER, G. W., .IND COXN, E. E., A\Tature, 197, T18 (1963).
FOKI)EN, L., .\SD BELL, E. .4., -Vatwe, 206, 110 (1965). STKOI~EL, G. .I., J. Bid. Chem., 241, 2618 (1966); 242, 3265
(1967). STROBEI,, C;. A., C'crn. J. Riochwn., 42, 1637 (1964). I~RYsK,RI.LI., C:ORPE, W.A4., ASD II.~sKEs, L. V.,J. Bacteriol.,
97, 322 (lsusj. I)UXNII,L, P. AI., .IXD FOJVDEN, I,., h-atwe, 208, 1206 (1965). RESSLER, C., GIZA, Y. H., .\XD NIG~~M, S. N., J. Amer. Chem.
Sot., 66, 2874 (1963). BLUMENTHAL,S.G., HENDRICKSON, H. 11., ABROL, Y.P., .~XD
CONN, E. E., J. Biol. Chem., 243, 5302 (1968). TSCHIERSCH, B., Flora, rlbt. A, Physiol. Biochem. (Jena), 166,
366 (1965). Floss, 1-T. G., K~DWIGER, L., ASD CONN, E. E., *VToture, 208,
1207 (1965). TIESI)RICKSOS, II. I:., Fed. Proc., 27, 593 (1968). Goss, 14:. E., ~SD BCTLER, G. W. in J. B. HARBORNE AND T.
SKIIS (Editors), Perspectives in phytochemislry, Academic Press, New 1-ork, in press.
SKOWRONRKI, B. s., .YSD STROBEL, G. A., Can. J. Microbial., 16, 93 (1969).
BREED, II. P., ;\~CRR.IY, E. G. II., .IND SMITH, N. R., Bergey’s )wn~aal of tlelerwinalizw bacteriology, Ed. 7, The Williams and Wilkirls Cornparr>-, Baltimore, 1957, p. 616.
D\KIN, II. D., Biochem. J., 11, 79 (1917). AI.DRII~(:E, W. S., Bnalysl, 69, 262 (1944). LOWRY, 0. IJ.,I:USEBROUGH,N. J., F.YRR, A. L., .IND RAND-ILL,
1~. J., J. Biol. Chem., 193, 265 (1951). RESSLER, C., J. Biol. Chem., 237, 733 (1962). Il’echnicon resenxh bulletin So. 20, Technicon Corporation,
Ardsley, New York, 1968. R.~~EI,, J. AI., YORTOX, S. J., HUMPHREYS, J. S., AND SHIVE,
W., J. Biol. Chem., 237. 2815 (19G2). BURCKZLL, J. J., REICHELT,~~. C., .ZNII WOLIN,M.J.,J. Biol.
Chem., 239, 1974 (1964). SINCHEZ, R. A., FERRIS, J. P., .IND ORGEL, L. E., Science,l64,
784 (19G6). 23. MICHAELS, R., EIa.~~css, L. V., AND CORPE, W. A., Arch. Bio-
them. Biophys., 111, 121 (1965).
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Peter A. Castric and Gary A. StrobelBacillus megateriumCyanide Metabolism by
1969, 244:4089-4094.J. Biol. Chem.
http://www.jbc.org/content/244/15/4089Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/244/15/4089.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from














![CLINICAL IMMUNOBIOLOGY color-accurate photomicrographic ... · I:R S S Polyadenylicacid, ammoniumsalt (adenylate-2,8-3H].100CnCimmole Current LOt 1 Ci mmole (rn-bized solid in screw-oap](https://img.pdfslide.us/doc/110x75/5ec52ef065b14f6204607824/clinical-immunobiology-color-accurate-photomicrographic-ir-s-s-polyadenylicacid.jpg)














