Embed Size (px)
Citation preview

JOUFWALOFNEUROPHYSIOLOGY Vol. 43, No. 5, May 1980. Printed in U.S.A.
Current Source-Density Analysis of the b-Wave of Frog Retina
ERIC A. NEWMAN
Research Laboratory of Electronics, Massachusetts Institute of Technology, Cambridge, Massachusetts 02139
SUMMARY AND CONCLUSIONS
1. Current source-density analysis was used to study the spatial and temporal properties of b-wave current sources and sinks in the frog eye cup.
2. The analysis shows that the b-wave arises primarily from a current source near the retinal surface and from two current sinks, one in the distal portion of the inner plexiform layer and the other in the outer plexiform layer.
3. The magnitude of the distal (outer plexiform layer) sink peaks at an earlier time than does the proximal sink, and decays at a faster rate. At early times, however, the proximal sink is greater in magnitude.
4. The results demonstrate that the cur- rents of the proximal and distal sinks both contribute positively to the transretinal voltage, summing to give the b-wave response.
5. The spatial and temporal properties of the two current sinks closely match those of the two light-evoked extracellular K+ ([K+],) increases observed by others. This cor- respondence supports the hypothesis that increases in [K+], lead to the generation of b-wave currents. The distribution of these currents indicate that they flow through the glial cells of Muller.
INTRODUCTION
Faber (7) and Miller and Dowling (18) were the first to suggest that the glial cells of Muller play a role in generating the b- wave of the electroretinogram (ERG), the gross electrical response of the eye. They proposed that changes in the extracellular potassium ion concentration ([K+],) result- ing from neuronal activity lead to local
changes in the Muller cell membrane poten- tial and to the establishment of a trans- retinal current flow. This notion has been supported in recent years by measurements of [K+], made by sever21 laboratories using ion-selective micropipettes. These experi- ments reveal two sites of light-evoked [K+], increase, one in the proximal retina (6, 13- 15,24) and the other in the distal retina(6,15).
A previous investigation of b-wave cur- rents (21) supports this K+ hypothesis of b- wave generation. In that study, I showed that b-wave currents are generated by a current source near the retinal surface and by two current sinks in the proximal and distal retina. The locations of these sinks roughly corresponded to the positions of the two sources of light-evoked [K+], increase, suggesting that the current sinks represent regions of increased [K+],.
The two sources of light-evoked K+ ef- flux can be distinguished by their time courses as well as their retinal locations. Recent observations by Kline et al. (15) and by Dick and Miller (6) have demonstrated that the distal [K+], increase is a more transient response than is the proximal one. If these [K+], increases are responsible for generating b-wave currents, their time courses should match the time courses of the two b-wave current sinks.
In the present study, I have conducted a detailed current source-density analysis of the b-wave in order to determine the spatial and temporal properties of the two b-wave current sinks. The computed current source- density distributions permit, for the first time, an accurate localization of the two current sinks within the retina. In addition, I have determined the time courses of the two sinks and compared them to the two
0022-3077/80/0000-0000$01.25 Copyright 0 1980 The American Physiological Society 1355

1356 E. A. NEWMAN
light-evoked [K+], increases observed by others. The results support the hypothesis that b-wave currents are generated by these [Kflo increases. A preliminary report of some of this work has appeared else- where (20).
METHODS
Preparation The eye cup preparation used in these experi-
ments is described in Newman and Lettvin (22) and Newman (21). In brief, eyes from dark- adapted Rana pipiens, the northern leopard frog, were transected under dim red illumination and placed on a pad of Ringer-soaked gauze. They were maintained in a moist 95% 02-5% CO, atmosphere at HOC. Eyes were dark-adapted for an additional 2 h before experiments were begun.
The DC, intraretinal ERG was recorded differ- entially between two Ringer-filled micropipettes: an active intraretinal electrode (-3-pm tip diam- eter) and a reference, vitreal electrode (-20-pm tip diameter). The tip of the vitreal electrode was positioned within 100 pm of the retinal surface, near the entry point of the intraretinal electrode. With this electrode configuration, b- waves within the retina were recorded as negative-going potentials at all retinal depths. b-Wave amplitudes were measured from the pre- stimulus base line (21). Retinal tissue resistance was measured by passing current pulses of con- stant amplitude (10 PA) across the eye cup (1, 2 1). Eye cups had a surface area of approxi- mately 0.25 cm2.
Stimuli consisted of diffuse white light flashes presented in the dark. The stimuli illuminated the entire eye cup uniformly and lOO-ms flashes were repeated once per minute for the duration of an experiment. Light intensities are given as log intensity; a lOO-ms flash at 0 intensity represents the scotopic equivalent of 7.8 x 10m4 photons/ pm2 of SOO-nm light. This was the dimmest stimulus that produced a detectable ERG re- sponse in the most sensitive preparations (22).
Current source-density analysis I calculated current source-density distribu-
tions by a simple difference method discussed by Freeman and Nicholson (8). This analysis is based on the following principles and assump- tions, which are founded on previous experimen- tal work:
1) The analysis I use is a one-dimensional treatment of the generalized three-dimensional calculation of current source density (12,23). As Nicholson and Freeman (23) showed, a one- dimensional calculation may be used when deal-
ing with a thin sheet (much wider than it is thick) of uniformly responding neural tissue, such as the retina. To ensure uniformity of response, the entire retina was evenly il- luminated during stimulus presentation. The con- sistency of the current source-density plots be- tween penetrations in different areas of the retina established that the retina was responding uni- formly to stimuli.
2) Nicholson and Freeman (23) showed that tissue resistance does not vary significantly dur- ing electrical activation of nervous tissue (in toad cerebellar cortex). I assumed, therefore, that retinal resistance values, which were meas- ured at -20 s following each stimulus presenta- tion, accurately reflect retinal resistance during the b-wave response.
3) My previous study of b-wave currents (21) demonstrated that the entire proximal current source of the b-wave is localized to an extremely narrow region (probably not more than 5- 10 pm in depth) bordering the inner limiting membrane. This was determined from electrode-advance ex- periments (advanced in 4-pm steps), which were not repeated in the present study. I have used this finding in calculating the current source- density distributions presented here, where I as- sume that the proximal current source occurs within one electrode step (8 or 12 pm) of the retinal surface. The actual impact of this as- sumption is small and affects only the shape, and not magnitude, of the proximal source.
4) I identify the proximal and distal boun- daries of the retina by resistance landmarks. The inner limiting membrane (0% retinal depth) is defined by the border of the low-resistance vitreous and the higher resistance retina (indi- cated in Fig. IB by the change in slope at 0% retinal depth). The inner surface of the pigment epithelium (100% retinal depth) coincides with the distal border of the high-resistance R mem- brane (2, 4). This is indicated by another large change in slope in Fig. l.B at a retinal depth of 192 pm.
RESUCTS
Computation of b-wave current source- density distribution
In this study I use current source-density analysis to obtain a quantitative description of the sources and sinks of retinal current that generate the b-wave response. This analysis is based on the calculation of the second spatial derivative of the intraretinal b-wave voltage, once it has been corrected for variations in retinal resistance (12, 23). I calculate these derivatives by a simple
ANALYSIS OF RETINAL b-WAVE 1357
RETINAL DEPTH (pm)
192
180 168
156
132
108
96
84
72
60
48
36 24 I2 0
B RETINAL DEPTH (pm)
I -24 illlllIlllllilllllr1 0 24 40 72 96 120 144 168 192
0.2mV] ’ _-
Ill I I I I J I I I I I I I I I 0
ThE (SE:) 3 0
% RETIN: DEPTH 100
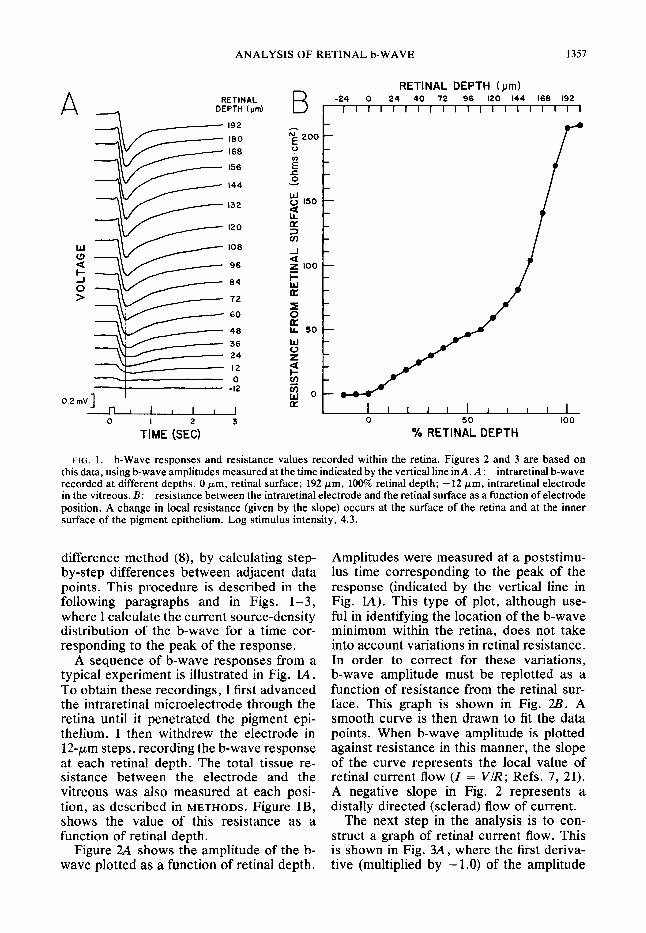
FIG. 1. b-Wave responses and resistance values recorded within the retina. Figures 2 and 3 are based on this data, using b-wave amplitudes measured at the time indicated by the vertical line in A. A : intraretinal b-wave recorded at different depths. 0 pm, retinal surface; 192 pm, 100% retinal depth; - 12 pm, intraretinal electrode in the vitreous. B: resistance between the intraretinal electrode and the retinal surface as a function of electrode position. A change in local resistance (given by the slope) occurs at the surface of the retina and at the inner surface of the pigment epithelium. Log stimulus intensity, 4.3.
difference method (8)) by calculating step- by-step differences between adjacent data points. This procedure is described in the following paragraphs and in Figs. l-3, where I calculate the current source-density distribution of the b-w ave for a time cor- responding to the peak of the response.
A sequence of b-wave responses from a typical experiment is illustrated in Fig. lA. To obtain these recordings, I first advanced the intraretinal microelectrode through the retina until it penetrated the pigment epi- thelium. I then withdrew the electrode in 12-pm step s, ret .ording the b-wave respon se at each retinal depth. The total tissue re- sistance between the electrode and the vitreous was also measured at each posi- tion, as described in METHODS. Figure lB, shows the value of this resistance as a function of retinal depth.
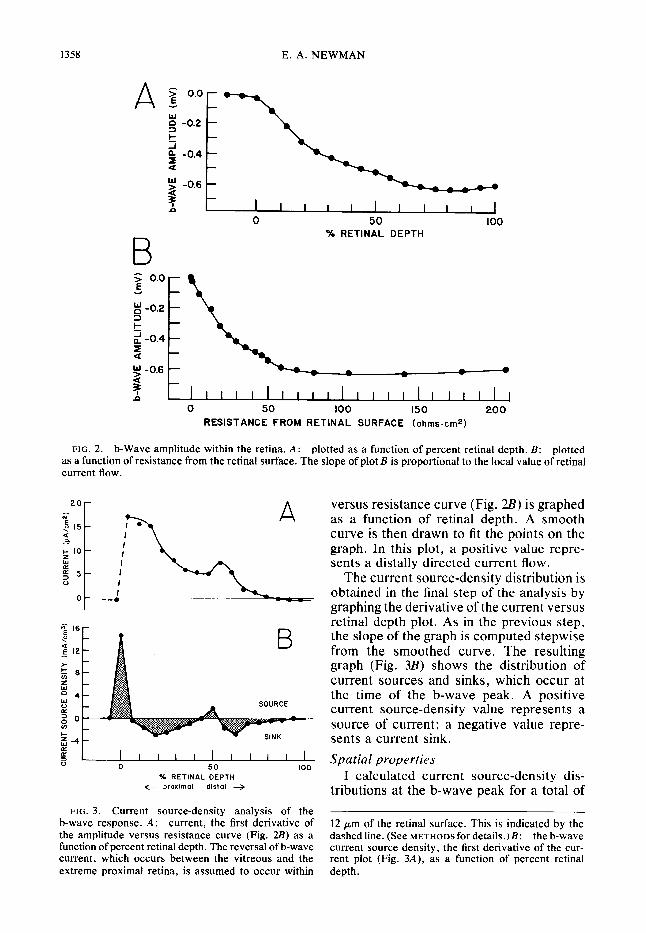
Figure 2A shows the amplitude of the b- wave plotted as a function of retinal depth.
Amplitudes were measured at a poststimu- lus time corresponding to the peak of the response (indicated by the vertical line in Fig. LA). This type of plot, although use- ful in identifying the location of the b-wave minimum within the retina, does not take into account variations in retinal resistance. In order to correct for these variations, b-wave amplitude must be replotted as a function of resistance from the retinal sur- face. This graph is shown in Fig. 2Z?. A smooth curve is then drawn to fit the data points. When b-wave amplitude is plotted against resistance in this manner, the slope of the curve represents the local value of retinal current flow (I = V/R; Refs. 7, 21). A negative slope in Fig. 2 represents a distally directed (sclerad) flow of current.
The next step in the analysis is to con- struct a graph of retinal current flow. This is shown in Fig. 3A, where the first deriva- tive (multiplied by - 1 .O) of the amplitude
1358 E. A. NEWMAN
z -0.2 2 2 g -0.4 a
9 -0.6
4 0 50 100
% RETINAL DEPTH
m -a
a v a
I Ill I II I I I I l l l 11 l 1 11 1 J 0 50 100 150 200
RESISTANCE FROM RETINAL SURFACE (ohmscm2)
FIG. 2. b-Wave amplitude within the retina. A: plotted as a function of percent retinal depth. B: plotted as a function of resistance from the retinal surface. The slope of plot B is proportional to the local value of retinal current flow.
0 I I I I II I II I I
0 50 100
% RETINAL DEPTH + proximal distal +
FIG. 3. Current source-density analysis of the b-wave response. A : current, the first derivative of the amplitude versus resistance curve (Fig. 2B) as a function of percent retinal depth. The reversal of b-wave current, which occurs between the vitreous and the extreme proximal retina, is assumed to occur within
versus resistance curve (Fig. 2B) is graphed as a function of retinal depth. A smooth curve is then drawn to fit the points on the graph. In this plot, a positive value repre- sents a distally directed current flow.
The current source-density distribution is obtained in the final step of the analysis by graphing the derivative of the current versus retinal depth plot. As in the previous step, the slope of the graph is computed stepwise from the smoothed curve. The resulting graph (Fig. 3B) shows the distribution of current sources and sinks, which occur at the time of the b-wave peak. A positive current source-density value represents a source of current; a negative value repre- sents a current sink. Spatial properties
I calculated current source-density dis- tributions at the b-wave peak for a total of
12 pm of the retinal surface. This is indicated by the dashed line. (See METHODS for details.)& the b-wave current source density, the first derivative of the cur- rent plot (Fig. 3A), as a function of percent retinal depth.
ANALYSIS OF RETINAL b-WAVE 1359
I I I I I I I I 0 IO 20 30 40 50 60 70
% RETINAL DEPTH
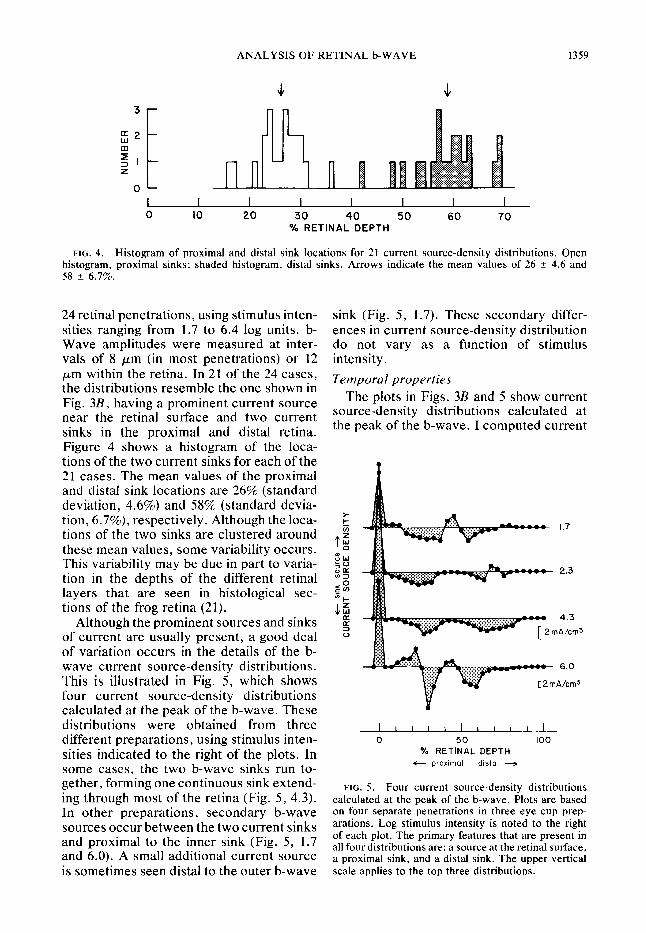
FIG. 4. Histogram of proximal and distal sink locations for 21 current source-density distributions. Open histogram, proximal sinks; shaded histogram, distal sinks. Arrows indicate the mean values of 26 f- 4.6 and 58 + 6.7%.
24 retinal penetrations, using stimulus inten- sink (Fig. 5, 1.7). These secondary differ- sities ranging from 1.7 to 6.4 log units. b- ences in current source-density distribution Wave amplitudes were measured at inter- do not vary as a function of stimulus vals of 8 ,um (in most penetrations) or 12 intensity.
Temporal properties pm within the retina. In 21 of the 24 cases, the distributions resemble the one shown in Fig. 3B, having a prominent current source near the retinal surface and two current sinks in the proximal and distal retina. Figure 4 shows a histogram of the loca- tions of the two current sinks for each of the 21 cases. The mean values of the proximal and distal sink locations are 26% (standard deviation, 4.6%) and 58% (standard devia- tion, 6.7%), respectively. Although the loca- tions of the two sinks are clustered around these mean values, some variability occurs. This variability may be due in part to varia- tion in the depths of the different retinal layers that are seen in histological sec- tions of the frog retina (2 1).
Although the prominent sources and sinks of current are usually present, a good deal of variation occurs in the details of the b- wave current source-density distributions. This is illustrated in Fig. 5, which shows four current source-density distributions calculated at the peak of the b-wave. These
The plots in Figs. 3B and 5 show current source-density distributions calculated at the peak of the b-wave. I computed current
distributions were obtained from three I I I I I I I I I I I
different preparations, using stimulus inten- 0 50 100
sities indicated to the right of the plots. In O/o RETINAL DEPTH
some cases, the two b-wave sinks run to- c- proximal distal -
gether, forming one continuous sink extend- FIG. 5. ing through most of the retina (Fig. 5, 4.3).
Four current source-density distributions calculated at the peak of the b-wave. Plots are based
In other nrenarations. secondarv b-wave on four separate penetrations in three eye cup prep- sources o&&between the two current sinks arations. Log stimulus intensity is noted to the right
and proximal to the inner sink (Fig. 5, 1.7 of each plot. The primary features that are present in
and 6.0). A small additional current source all four distributions are: a source at the retinal surface, a proximal sink, and a distal sink. The upper vertical
is sometimes seen distal to the outer b-wave scale applies to the top three distributions.
1360 E. A. NEWMAN
A a b C
I I I
> ” I 1 I I
0 I TIM: (SEC)
3 4 > ” 1 I 1 1
0 I TIM: 3 4 (SEC)
I I I I I I I I I I I
0 50 100 % RETINAL DEPTH
I II 1 I I I I II I
0 50 100
% RETINAL DEPTH
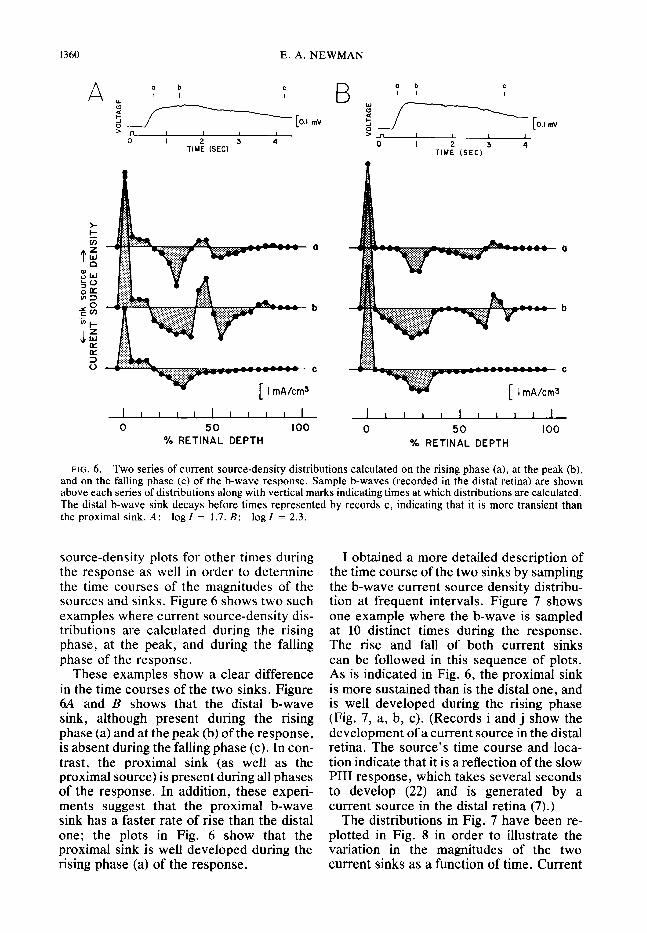
FIG. 6. Two series of current source-density distributions calculated on the rising phase (a), at the peak (b), and on the falling phase (c) of the b-wave response. Sample b-waves (recorded in the distal retina) are shown above each series of distributions along with vertical marks indicating times at which distributions are calculated. The distal b-wave sink decays before times represented by records c, indicating that it is more transient than the proximal sink. A: log1 = 1.7. B: log1 = 2.3.
source-density plots for other times during the response as well in order to determine the time courses of the magnitudes of the sources and sinks. Figure 6 shows two such examples where current source-density dis- tributions are calculated during the rising phase, at the peak, and during the falling phase of the response.
These examples show a clear difference in the time courses of the two sinks. Figure 6A and B shows that the distal b-wave sink, although present during the rising phase (a) and at the peak (b) of the response, is absent during the falling phase (c). In con- trast, the proximal sink (as well as the proximal source) is present during all phases of the response. In addition, these experi- ments suggest that the proximal b-wave sink has a faster rate of rise than the distal one; the plots in Fig. 6 show that the proximal sink is well developed during the rising phase (a) of the response.
I obtained a more detailed description of the time course of the two sinks by sampling the b-wave current source density distribu- tion at frequent intervals. Figure 7 shows one example where the b-wave is sampled at 10 distinct times during the response. The rise and fall of both current sinks can be followed in this sequence of plots. As is indicated in Fig. 6, the proximal sink is more sustained than is the distal one, and is well developed during the rising phase (Fig. 7, a, b, c). (Records i and j show the development of a current source in the distal retina. The source’s time course and loca- tion indicate that it is a reflection of the slow PI11 response, which takes several seconds to develop (22) and is generated by a current source in the distal retina (7).)
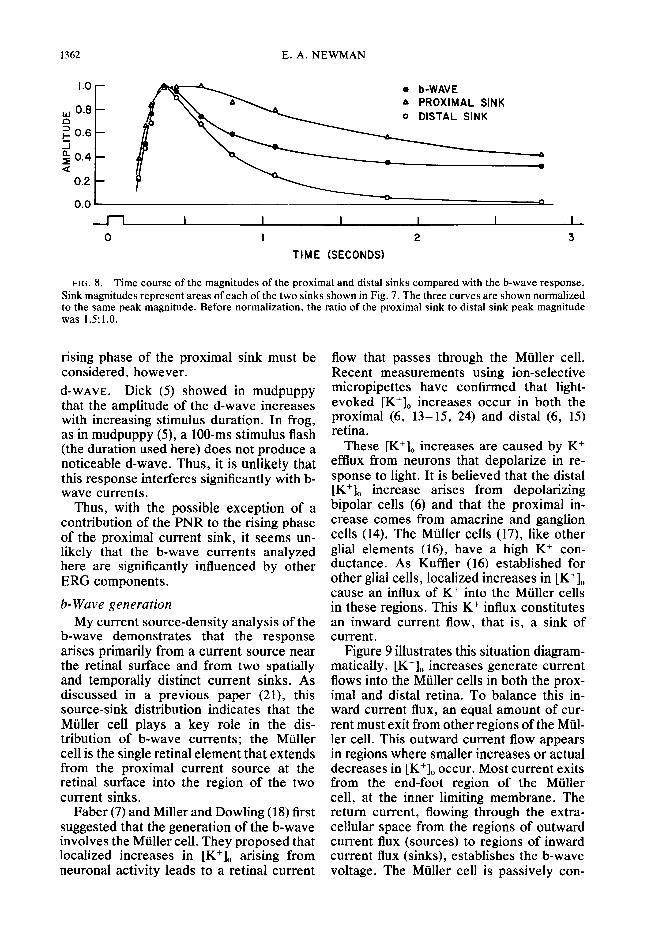
The distributions in Fig. 7 have been re- plotted in Fig. 8 in order to illustrate the variation in the magnitudes of the two current sinks as a function of time. Current
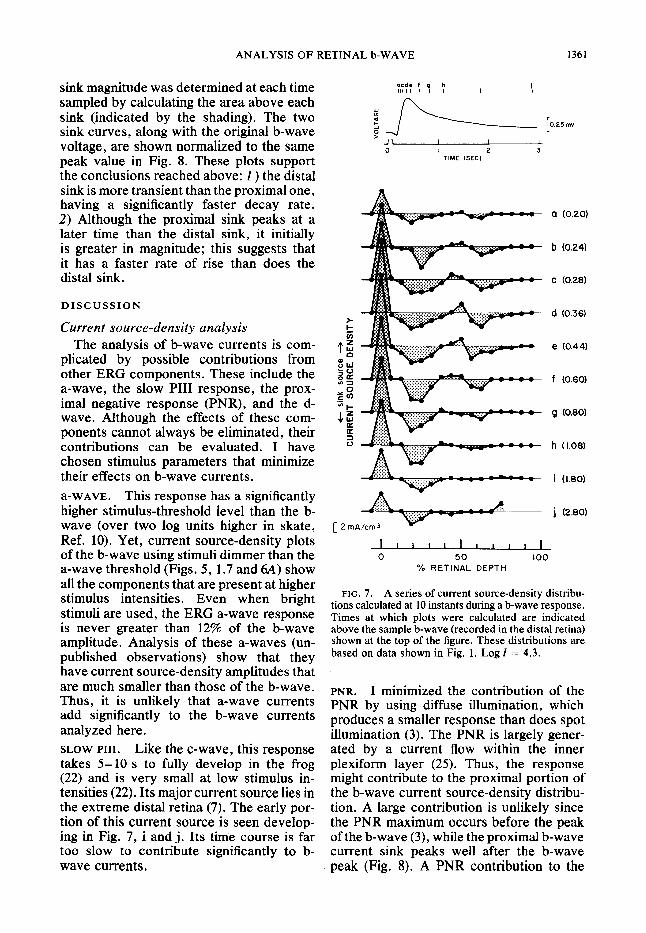
ANALYSIS OF RETINAL b-WAVE 1361
sink magnitude was determined at each time sampled by calculating the area above each sink (indicated by the shading). The two sink curves, along with the original b-wave voltage, are shown normalized to the same peak value in Fig. 8. These plots support the conclusions reached above: 1) the distal sink is more transient than the proximal one, having a significantly faster decay rate. 2) Although the proximal sink peaks at a later time than the distal sink, it initially is greater in magnitude; this suggests that it has a faster rate of rise than does the distal sink.
DISCUSSION
Current source-density analysis The analysis of b-wave currents is com-
plicated by possible contributions from other ERG components. These include the a-wave, the slow PI11 response, the prox- imal negative response (PNR), and the d- wave. Although the effects of these com- ponents cannot always be eliminated, their contributions can be evaluated. I have chosen stimulus parameters that minimize their effects on b-wave currents. a-WAVE. This response has a significantly higher stimulus-threshold level than the b- wave (over two log units higher in skate, Ref. 10). Yet, current source-density plots of the b-wave using stimuli dimmer than the a-wave threshold (Figs. 5, 1.7 and 64) show all the components that are present at higher stimulus intensities. Even when bright stimuli are used, the ERG a-wave response is never greater than 12% of the b-wave amplitude. Analysis of these a-waves (un- published observations) show that they have current source-density amplitudes that are much smaller than those of the b-wave. Thus, it is unlikely that a-wave currents add significantly to the b-wave currents analyzed here. SLOW PIII. Like the c-wave, this response takes 5- 10 s to fully develop in the frog (22) and is very small at low stimulus in- tensities (22). Its major current source lies in the extreme distal retina (7). The early por- tion of this current source is seen develop- ing in Fig. 7, i and j. Its time course is far too slow to contribute significantly to b- wave currents.
acde f III I I I s: t ‘I
TIME (SEC)
a (0.20)
b (0.24)
c (0.28)
d (0.36)
e (0.44)
f (0.60)
g (0.80)
h (I.081
i (I.801
j (2.80)
I I I I I I I I 11 I
0 50 100 % RETINAL DEPTH
FIG. 7. A series of current source-density distribu- tions calculated at 10 instants during a b-wave response. Times at which plots were calculated are indicated above the sample b-wave (recorded in the distal retina) shown at the top of the figure. These distributions are based on data shown in Fig. 1. Log Z = 4.3.
PNR. I minimized the contribution of the PNR by using diffuse illumination, which produces a smaller response than does spot illumination (3). The PNR is largely gener- ated by a current flow within the inner plexiform layer (25). Thus, the response might contribute to the proximal portion of the b-wave current source-density distribu- tion. A large contribution is unlikely since the PNR maximum occurs before the peak of the b-wave (3), while the proximal b-wave current sink peaks well after the b-wave peak (Fig. 8). A PNR contribution to the
1362 E. A. NEWMAN
ki 0.8
z 0.6 i g 0.4 a
. b-WAVE A PROXIMAL SINK 0 DISTAL SINK
I I I I I I I
0 I 2 3
TIME (SECONDS)
FIG. 8. Time course of the magnitudes of the proximal and distal sinks compared with the b-wave response. Sink magnitudes represent areas of each of the two sinks shown in Fig. 7. The three curves are shown normalized to the same peak magnitude. Before normalization, the ratio of the proximal sink to distal sink peak magnitude was 1.5:l.O.
rising phase of the proximal sink must be considered, however. d-WAVE. Dick (5) showed in mudpuppy that the amplitude of the d-wave increases with increasing stimulus duration. In frog, as in mudpuppy (S), a lOO-ms stimulus flash (the duration used here) does not produce a noticeable d-wave. Thus, it is unlikely that this response interferes significantly with b- wave currents.
Thus, with the possible exception of a contribution of the PNR to the rising phase of the proximal current sink, it seems un- likely that the b-wave currents analyzed here are significantly influenced by other ERG components.
b-Wave generation My current source-density analysis of the
b-wave demonstrates that the response arises primarily from a current source near the retinal surface and from two spatially and temporally distinct current sinks. As discussed in a previous paper (21), this source-sink distribution indicates that the Mtiller cell plays a key role in the dis- tribution of b-wave currents; the Mtiller cell is the single retinal element that extends from the proximal current source at the retinal surface into the region of the two current sinks.
Faber (7) and Miller and Dowling (18) first suggested that the generation of the b-wave involves the Muller cell. They proposed that localized increases in [K+], arising from neuronal activity leads to a retinal current
flow that passes through the Mtiller cell. Recent measurements using ion-selective micropipettes have confirmed that light- evoked [K+], increases occur in both the proximal (6, 13-15, 24) and distal (6, 15) retina.
These [K+], increases are caused by K+ efflux from neurons that depolarize in re- sponse to light. It is believed that the distal lK+1 increase arises from depolarizing bipolar cells (6) and that the proximal in- crease comes from amacrine and ganglion cells (14). The Mtiller cells (17), like other glial elements (16), have a high K+ con- ductance. As Kuffler (16) established for other glial cells, localized increases in [K+], cause an influx of K+ into the Muller cells in these regions. This K+ influx constitutes an inward current flow, that is, a sink of current.
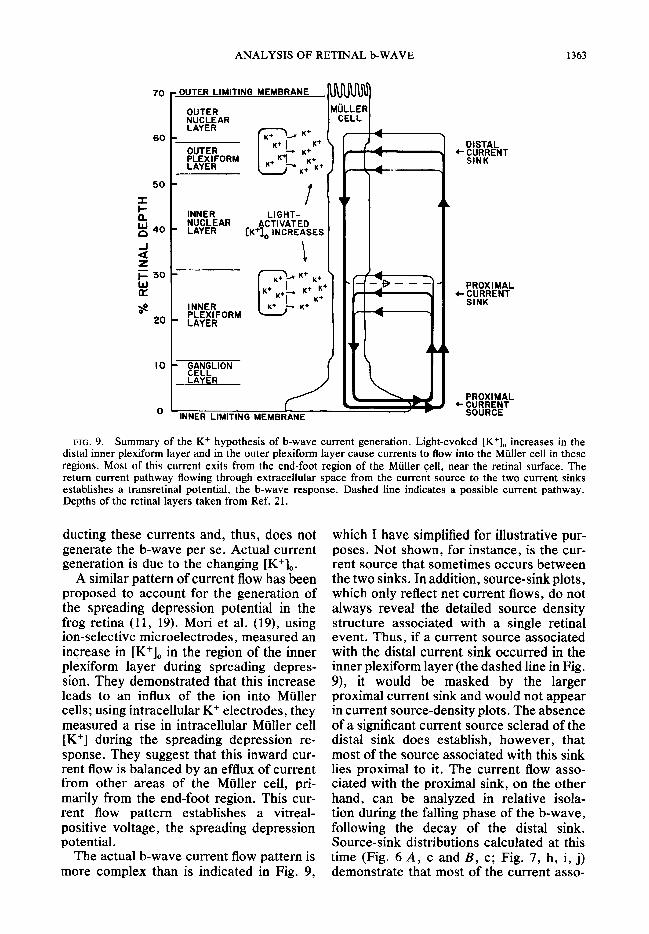
Figure 9 illustrates this situation diagram- matically . [K+], increases generate current flows into the Muller cells in both the prox- imal and distal retina. To balance this in- ward current flux, an equal amount of cur- rent must exit from other regions of the Mtil- ler cell. This outward current flow appears in regions where smaller increases or actual decreases in [K+], occur. Most current exits from the end-foot region of the Muller cell, at the inner limiting membrane. The return current, flowing through the extra- cellular space from the regions of outward current flux (sources) to regions of inward current flux (sinks), establishes the b-wave voltage. The Muller cell is passively con-
60
ANALYSIS OF RETINAL b-WAVE
1 OUTER LIMITING MEMBRANE
OUTER NUCLEAR LAYER
OUTER PLEXIFORM LAYER
K+ K+
K+ K+ K+
INNER LIGHT- NUCLEAR
’ LAYER ACTIVATED
[K+lo INCREASES
INNER PLEXIFORM
l LAYER
K+ A C K+ ’ K++
I K+ -
’ MYLIoN LAYER
DISTAL
+ &EENT
PROXIMAL
+ i%EENT
FIG. 9. Summary of the K+ hypothesis of b-wave current generation. Light-evoked [K+], increases in the distal inner plexiform layer and in the outer plexiform layer cause currents to flow into the Miiller cell in these regions. Most of this current exits from the end-foot region of the Muller cell, near the retinal surface. The return current pathway flowing through extracellular space from the current source to the two current sinks establishes a transretinal potential, the b-wave response. Dashed line indicates a possible current pathway. Depths of the retinal layers taken from Ref. 21.
ducting these currents and, thus, does not generate the b-wave per se. Actual current generation is due to the changing [K+],.
A similar pattern of current flow has been proposed to account for the generation of the spreading depression potential in the frog retina (11, 19). Mori et al. (19), using ion-selective microelectrodes, measured an increase in [K’], in the region of the inner plexiform layer during spreading depres- sion. They demonstrated that this increase leads to an influx of the ion into Miiller cells; using intracellular K+ electrodes, they measured a rise in intracellular Mtiller cell [K+] during the spreading depression re- sponse. They suggest that this inward cur- rent flow is balanced by an efflux of current from other areas of the Muller cell, pri- marily from the end-foot region. This cur- rent flow pattern establishes a vitreal- positive voltage, the spreading depression potential.
The actual b-wave current flow pattern is more complex than is indicated in Fig. 9,
which I have simplified for illustrative pur- poses. Not shown, for instance, is the cur- rent source that sometimes occurs between the two sinks. In addition, source-sink plots, which only reflect net current flows, do not always reveal the detailed source density structure associated with a single retinal event. Thus, if a current source associated with the distal current sink occurred in the inner plexiform layer (the dashed line in Fig. 9), it would be masked by the larger proximal current sink and would not appear in current source-density plots. The absence of a significant current source sclerad of the distal sink does establish, however, that most of the source associated with this sink lies proximal to it. The current flow asso- ciated with the proximal sink, on the other hand, can be analyzed in relative isola- tion during the falling phase of the b-wave, following the decay of the distal sink. Source-sink distributions calculated at this time (Fig. 6 A, c and B, c; Fig. 7, h, i, j) demonstrate that most of the current asso-

1364 E. A. NEWMAN
ciated with the proximal sink exits from the Muller cell end-foot region. Spatial properties
My results indicate that the two sinks of b-wave current are closely associated with the two light-evoked [K+], increases. The current source-density profiles presented here permit, for the first time, an accurate localization of the two b-wave sinks. The mean values of the experimental results in- dicate that the peaks of the proximal and distal sinks occur at approximately 26 and 58% retinal depths. These depths corre- spond to retinal locations in the distal por- tion of the inner plexiform layer (extending from 11 to 31% retinal depth) and in the outer plexiform layer (52 to 59% retinal depth). (Depth figures are mean values taken from histological sections of the frog retina, Ref. 21.)
Karwoski and Proenza (14) localized the source of the proximal [K+], increase to near the border of the inner plexiform and inner nuclear layers in mudpuppy. Kline et al. (15) localized the K+ source in skate slightly more proximally, in the distal region of the inner plexiform layer. In recent experiments, Dick and Miller (6) in mudpuppy and Kline et al. (15) in skate have also recorded light- evoked [K+], increases in the distal retina. Both works indicate that the distal K+ source lies in the outer plexiform layer. Thus, there is an excellent correspondence between the locations of the two K+ sources and posi- tions of the two b-wave current sinks. Temporal properties
The association between the b-wave cur- rent sinks and increased [K+], is further sup- ported by a comparison of their time courses. Kline et al. (15) and Dick and Miller (6) show that the distal increase in [K+], is more transient than is the proximal one. The two b-wave sinks have a similar time relation. As can be seen in Figs. 6, 7, and 8, the distal b-wave sink is more transient than is the proximal one. A report by Proenza and Freeman (25) also indicated that the distal b-wave sink has a faster time course than the proximal sink.
Figure 8 illustrates, in addition, that the proximal b-wave sink is initially greater in magnitude than the distal sink. This is con- sistent with the origin of the proximal [K+], increase, which is generally thought to be caused by depolarizing neurons in the inner
plexiform and inner nuclear layers, primarily amacrine and ganglion cells (14). Although the responses of these cells have a slightly longer latency than those of the horizontal and bipolar cells, they have a much faster rate of rise. This is clearly illustrated in Werblin and Dowling (26). Thus, the proxi- mal [K+], increase should have a faster rate of rise than the distal increase. The [K+], measurements of Kline et al. (15) and Dick and Miller (6), although not definitive, are consistent with this view.
The current source-density profiles in Figs. 3B, 5, 6, and 7 show a variety of sec- ondary sources and sinks of b-wave current located distal to, in between, and proximal to the two major current sinks. A small b-wave source is sometimes seen just sclerad of the distal sink, as noted by Faber (7). The differences between individual plots may be due, in part, to variations in the magnitude of K+ diffusion away from the two sites of K+ efflux (which are themselves diffuse K+ sources). When [I(+], increases are restricted to the immediate vicinity of the proximal and distal K+ sources, the locations of the two b-wave sinks will be similarly restricted. In this case, b-wave sources will be seen surrounding the two sinks (Fig. 5, 1.7 and 6.0). Conversely, when [K+], increases are more evenly distributed, one continuous b-wave sink will be seen to extend through- out much of the retina (Fig. 5, 4.3). The latter situation might be brought about by particularly damaging microelectrode pene- trations, which would facilitate K+ diffusion in a radial direction.
b-Wave summation model Proenza and Freeman (25) and Kline et al.
(15) have suggested that the distal b-wave sink is primarily associated with the rising phase of the response, while the proximal sink is related to its falling phase. Kline et al. (15) have proposed a specific model in which the decay of the b-wave is directly linked to the rise of the proximal sink. In their hy- pothesis, the distal sink generates a positive transretinal voltage, while the slower proximal sink generates a negative one.
My results do not support this hypothesis, at least for the case of the frog retina. As seen in Fig. 9, the dipoles established by the proximal and distal current sinks are oriented in the same direction. Thus, the currents associated with these two sinks
ANALYSIS OF RETINAL b-WAVE 1365
0.6
5 cs 0.4
%
!i 9 0.2
0.0
0 b-WAVE l SUM OF SINK VOLTAGES A PROXIMAL SINK VOLTAGE l DISTAL SINK VOLTAGE
I I I I I I 0 I 2 3
TIME (SECONDS)
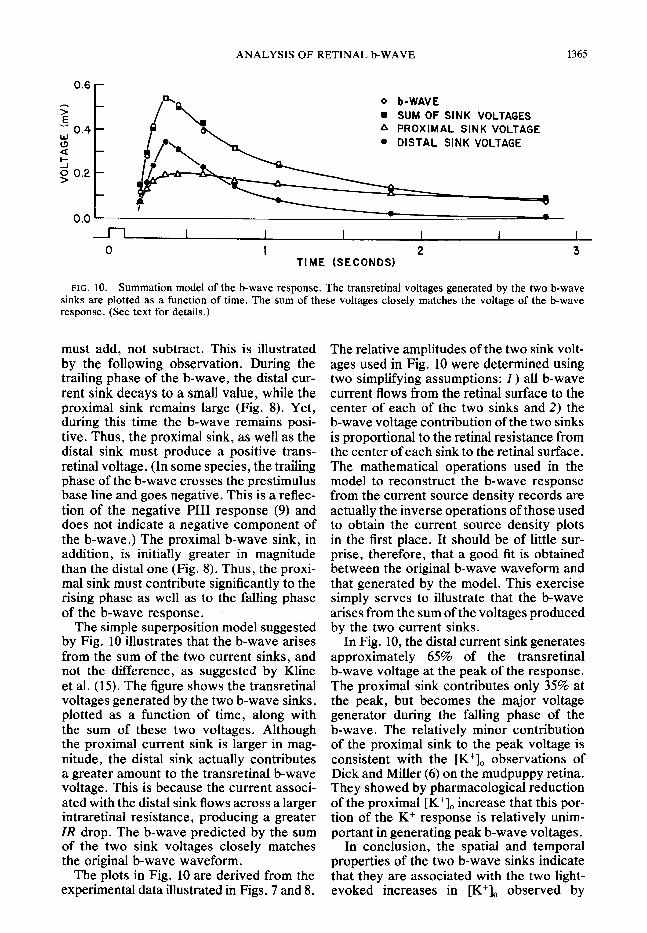
FIG. 10. Summation model of the b-wave response. The transretinal voltages generated by the two b-wave sinks are plotted as a function of time. The sum of these voltages closely matches the voltage of the b-wave response. (See text for details.)
must add, not subtract. This is illustrated The relative amplitudes of the two sink volt- by the following observation. During the ages used in Fig. 10 were determined using trailing phase of the b-wave, the distal cur- two simplifying assumptions: 1) all b-wave rent sink decays to a small value, while the current flows from the retinal surface to the proximal sink remains large (Fig. 8). Yet, center of each of the two sinks and 2) the during this time the b-wave remains posi- b-wave voltage contribution of the two sinks tive. Thus, the proximal sink, as well as the is proportional to the retinal resistance from distal sink must produce a positive trans- the center of each sink to the retinal surface. retinal voltage. (In some species, the trailing The mathematical operations used in the phase of the b-wave crosses the prestimulus model to reconstruct the b-wave response base line and goes negative. This is a reflec- from the current source density records are tion of the negative PI11 response (9) and actually the inverse operations of those used does not indicate a negative component of to obtain the current source density plots the b-wave.) The proximal b-wave sink, in in the first place. It should be of little sur- addition, is initially greater in magnitude prise, therefore, that a good fit is obtained than the distal one (Fig. 8). Thus, the proxi- between the original b-wave waveform and mal sink must contribute significantly to the that generated by the model. This exercise rising phase as well as to the falling phase simply serves to illustrate that the b-wave of the b-wave response. I arises from the sum of the voltages produced
The simple superposition model suggested by the two current sinks. by Fig. 10 illustrates that the b-wave arises from the sum of the two current sinks, and not the difference, as suggested by Kline et al. (15). The figure shows the transretinal voltages generated by the two b-wave sinks, plotted as a function of time, along with the sum of these two voltages. Although the proximal current sink is larger in mag- nitude, the distal sink actually contributes a greater amount to the transretinal b-wave voltage. This is because the current associ- ated with the distal sink flows across a larger intraretinal resistance, producing a greater IR drop. The b-wave predicted by the sum of the two sink voltages closely matches the original b-wave waveform.
In Fig. 10, the distal current sink generates approximately 65% of the transretinal b-wave voltage at the peak of the response. The proximal sink contributes only 35% at the peak, but becomes the major voltage generator during the falling phase of the b-wave. The relatively minor contribution of the proximal sink to the peak voltage is consistent with the [K+], observations of Dick and Miller (6) on the mudpuppy retina. They showed by pharmacological reduction of the proximal [K+], increase that this por- tion of the K+ response is relatively unim- portant in generating peak b-wave voltages.
The plots in Fig. 10 are derived from the experimental data illustrated in Figs. 7 and 8.
In conclusion, the spatial and temporal properties of the two b-wave sinks indicate that they are associated with the two light- evoked increases in [K+], observed by

1366 E. A. NEWMAN
others. This correspondence supports the cussions during the course of this work and for their hypothesis that b-wave currents are gen- comments On the text* erated by changes in [K+],. The currents This research was supported in part by the National
Institutes of Health Grants TO1 EY-00090 and T32 of both the proximal and distal sinks con- EY-07028, and in part by a grant from Bell Telephone tribute positively to the transretinal poten- Laboratories, Inc. tial, summing to give the b-wave response. Present address of author: Eye Research Institute The distribution of these currents indicates of Retina Foundation, 20 Staniford St., Boston, MA
a current pathway through the Muller cells. 02114. Please send reprint requests to the author at his present address.
ACKNOWLEDGMENTS
I thank Jerome Lettvin, Stephen Raymond, Peter Received 7 March Hartline, and Janice Gepner for their many helpful dis- December 1979.
REFERENCES
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
BRINDLEY, G. S. The passive electrical properties of the frog’s retina, choroid and sclera for radial fields and currents. J. Physiol. London 134: 339- 352, 1956. BRINDLEY, G. S. AND HAMASAKI, D. I. The prop- erties and nature of the R-membrane of the frog’s eye. J. Physiol. London 167: 599-606, 1963. BURKHARDT, D. A. Proximal negative response of the frog retina. J. Neurophysiol. 33: 405-420, 1970.
COHEN, A. I. A possible cytological basis for the ‘R’ membrane in the vertebrate eye. Nature Lon- don 205: 1222-1223, 1965. DICK, E. Light- and Dark-Dependent Extracellular F-Activity Modulations in the Vertebrate Retina; Origins of Electroretinographic Components (Ph.D. Thesis). Buffalo: State University of New York, 1979.
DICK, E. AND MILLER, R. F. Light-evoked potas- sium activity in mudpuppy retina: its relation to the b-wave of the electroretinogram. Bruin Res. 154: 388-394, 1978.
FABER, D. S. Analysis of Slow Transretinal Po- tentials in Response to Light (Ph.D. Thesis). Buffalo: State University of New York, 1969. FREEMAN, J. A. AND NICHOLSON, C. Experimental optimization of current source-density technique for anuran cerebellum. J. Neurophysiol. 38: 369- 382, 1975. GRANIT, R. The components of the retinal action potential in mammals and their relation to the dis- charge in the optic nerve. J. Physiol. London 77: 207-239, 1933. GREEN, D. G., DOWLING, J. E., SIEGEL, I. M., AND RIPPS, H. Retinal mechanisms of visual adapta- tion in the skate. J. Gen. Physiol. 65: 483-502, 1975. HANAWA, I., KUGE, K., AND MATSUMURA, K. Mechanism of the slow depressive potential pro- duction in the isolated frog retina. Jpn. J. Physiol. 18: 59-70, 1968,
HOWLAND, B., LETTVIN, J. Y., MCCULLOCH, W. S., PITTS, W., AND WALL, P. D. Reflex in- hibition by dorsal root interaction. J. Neurophysiol. 18: 1-17, 1955.
KARWOSKI, C. J. AND PROENZA, L. M. Relation- ship between Mtiller cell responses, a local trans- retinal potential, and potassium flux. J. Neuro-
1979; accepted in final form 17
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
KARWOSKI, C. J. AND PROENZA, L. M. Light- evoked changes in extracellular potassium concen- tration in mudpuppy retina. Bruin Res. 142: 515- 530, 1978. KLINE, R. P., RIPPS, H., AND DOWLING, J. E. The generation of b-wave currents in the skate retina. Proc. Natl. Acad. Sci. USA 75: 5727-5731, 1978. KUFFLER, S. W. Neuroglial cells: physiological properties and a potassium mediated effect of neuronal activity on the glial membrane potential. Proc. R. Sot. London Ser. B 168: 1-21, 1967. MILLER, R. F. Role of K+ in generation of the b-wave of the electroretinogram. J. Neurophysiol. 36: 28-38, 1973. MILLER, R. F. AND DOWLING, J. E. Intracellular responses of the Mtiller (glial) cells of the mud- puppy retina: their relation to b-wave of the elec- troretinogram. J. Neurophysiol. 33: 323-341,197O. MORI, S., MILLER, W. H., AND TOMITA, T. Mtiller cell function during spreading depression in the frog retina. Proc. Natl. Acad. Sci. USA 73: 1351- 1354, 1976. NEWMAN, E. A. The role of Mtiller cells in the generation of the retinal b-wave response: a source density analysis. Sot. Neurosci. Abstr. 8: 639, 1978.
NEWMAN, E. A. B-wave currents in the frog retina. Vision Res. 19: 227-234, 1979. NEWMAN, E. A. AND LETTVIN, J. Y. Relation of the e-wave to ganglion cell activity and rod re- sponses in the frog. Vision Res. 18: 1181- 1188, 1978.
NICHOLSON, C. AND FREEMAN, J. A. Theory of current source-density analysis and determination of conductivity tensor for anuran cerebellum. J. Neurophysiol. 38: 356-368, 1975. OAKLEY, B. II AND GREEN, D. G. Correlation of light-induced changes in retinal extracellular potas- sium concentration with c-wave of the electro- retinogram. J. Neurophysiol. 39: 1117- 1133, 1976. PROENZA, L. M. AND FREEMAN, J. A. Light-evoked extracellular potentials of the Necturus retina: current source density analysis of the electro- retinographic b-wave and the proximal negative response. Sot. Neurosci. Abstr. 5: 104, 1975. WERBLIN, F. S. AND DOWLING, J. E. Organization of the retina of the mudpuppy Necturus maculo- sus . II. Intracellular recordings. J. Neurophysiol.
physiol. 40: 244-259, 1977. 32: 339-355, 1969.





























