Embed Size (px)
Citation preview

CURRENT KNOWLEDGE OF LARVAE OF SCULPINS (PISCES:
COTTIDAE AND ALLIES) IN NORTHEAST PACIFIC GENERAWITH NOTES ON INTERGENERIC RELATIONSHIPS 1
SALLY L. RICHARDSON 2
ABSTRACf
Current knowledge of cottid larvae in northeast Pacific genera is summarized. Larvae are known forrepresentatives of25 of the 40 genera reported from Baja California to the Aleutian Islands althoughtwo genera, Gymnocanthus and lcelus, are represented only by species which live in other areas asadults. Included are illustrations oflarvae of29 species representing the 25 genera plus one potentiallynew northeast Pacific genus, identified only as "Cottoid Type A."
The larvae exhibit a wide diversity of form. Based on shared larval characters, including spinepatterns, body shape, and pigmentation, 6 phenetically derived groups of genera are apparent withinthe 25 genera for which representatives are considered: 1) A rtedius , Clinocottus, Oligocottus,Orthonopias; 2) Paricelinus, Triglops, lcelus, Chitonotus, Icelinus; 3) Dasycottus, Psychrolutes,Gilbertidia, ?Malacocottus, "Cottoid Type A"; 4) Scorpaenichthys, Hemilepidotus; 5) Blepsias,Nautichthys; 6) Leptocottus, Cottus. Six genera do not fit with any group: Enophrys, Gymnocanthus,Myoxocephalus, Radulinus, Rhamphocottus, Hemitripterus.
Ifthese preliminary larval groupings reflect relationship, as evidence indicates, they tend to supporta number of previously implied relationships within the cottids, but there are some importantdifferences. These include the distinctiveness of the Artedius (Group 1) line; the separation ofArtedius and Icelus, once considered closely related; the relationship of Paricelinus, generallyconsidered a primitive and rather distinct form, with other members of Group 2; the apparentrelationship of lcelus to other genera in Group 2 and its questionable placement in a separate family;the distinctiveness of Radulinus, previously considered to be related to Chitonotus and Icelirms.
The Cottidae, which in this paper are consideredbroadly to include sculpinlike fishes of Cottidae,Icelidae, Cottocomephoridae, Comephoridae, Norlllanichthyidae, Cottunculidae, and Psychrolutidae of the suborder Cottoidei of Greenwood et al.(1966), comprises a diverse group oftemperate andboreal fishes. Nelson (1976) estimated that thegroup may contain over 350 species, three-fourthslllarine, in about 86 genera. They are generallyCoastal fishes inhabiting all oceans but the Indian.Greatest species diversity occurs in the NorthPacific. The systematics of the group are not wellunderstood (Quast 1965; Nelson 1976).
Until recently, larvae of relatively few cottids?ad been described. They were a difficult group toIdentify in ichthyoplankton collections, particularly in the northeast Pacific where 40 genera arereported to occur between Baja California and the
--;:-----''!-'his paper was presented at the Second International Sym-
f1~um on The Early Life History of Fish (sponsored by ICES,197 ,ICNAF, IAOB, SCOR) held at Woods Hole, Mass., 2-5 April
P b91: An abstract of the paper appeared in the symposium
~ 'cation.S <?ulf Coast Research Laboratory, East Beach Drive, Ocean
Prtngs, MS 39564.
M------,----FtsnusCript accepted August 1980.
HERY BULLETIN: VOL. 79, NO.1, 1981.
Aleutian Islands (Table 1). With the recent workby Richardson and Washington (1980), larvae arenow known for representatives of 25 of these 40cottid genera, although two genera, Icelus andGymnocanthus, are represented only by larvae ofspecies that live in other areas as adults.
The purpose of this paper is twofold. It presentsfor the first time a summary of important cottidlarval characters (those characters occurring onlyduring the larval period and most useful in identifying and distinguishing species) based on thelarvae of these 25 northeast Pacific genera. (Larvae of these genera that are known for speciesinhabiting other areas as adults are also considered.) This knowledge, which is a necessaryprerequisite for systematic studies using larvae, ispresented to provide a foundation to which futurework on cottid larvae can be compared and uponwhich it can be expanded as more larvae becomeknown. The paper also presents a preliminaryexamination of generic groupings within thesenortheast Pacific cottid genera based on sharedlarval characters, Le., similarity. These pheneticgroupings, even though preliminary, are helpful
103

FISHERY BULLETIN: VOL. 79, NO. I
TABLE I.-List of cottid genera occuring in the northeast Pacific Ocean between Baja California and the Aleutian Islands based onHowe and Richardson (text footnote 3) with a summary of illustrations (accessible to author) oflarvae known for those genera worldwide. Lengths (millimeters) oflarvae are reported as they appeared in the literature: NL = notochord length; 8L = standard length;TL = total length; mm = no length definition was given.
Genus and species
Alfediel/us Altedius harringtoniA. laferalis
Alfedius Type 2Ascel/chthys Asemichthys Biepsias ci"hosus
Chitono/us pugetensis
Clinocottus acuticepsC. analis
C. reca/vusCottus asper
Dasycottus setigerEnophrys bison
E. bubalis'
E. liI/jeborgi'
Eurymen-GI/bart/dia sigalutes
Gymnocanthus herzenstainl'G. tricuspis 1
G. ventralis 1
Heml/epidotus gilberti'
H. heml/epidotus
H.jordani
H.papilioH. spinosus
H. zapusHemitripterus americanus'
H. viliosus·
leal/nus spp.·tee/us bicornis'
Jordania Laiocottus Leptocottus armatus
Ma/acocottus 1M. zonurus Type 1
Myoxocephalus aenaeus'
M. octodecemsplnosus'
M. polyacanfhocephalusM. quadricornis 1 (marine form)
104
Reference
Richardson and Washington 1980Budd 1940Marliave 1975Richardson and Washington 1980
Blackburn 1973Marliave 1975Richardson and Washington 1980Misitano 1980Richardson and Washington 1980Eigenmann 1892Budd 1940Morris 1951Stein 1972Richardson and Washington 1980Blackburn 1973Blackburn 1973Marliave 1975Misitano 1978Richardson and Washington 1980Cunningham 1891"Mcintosh and Masterman 1897"Ehrenbaum 1904"Ehrenbaum 1905-9Russell 1976'Bruun 1925'Rass 1949'Russell 19766
Blackburn 1973Marliave 1975Kyushln 1970Koefoed 1907Rass 1949Khan 1972Ehranbaum 1905-9Gorbunova 1964Hattori 1964Gorbunova 1964Peden 1978Richardson and Washington 1980Gorbunova 1964Peden 1978Gorbunova 1964Follett 1952Peden 1978Richardson and Washington 1980Pedan 1978Warfel and Merriman 1944Khan 1972Fuiman 1976Kyushln 1968Okiyama and sando 1976Richardson and Washington 1980Ehrenbaum 1905-9'·Rass 1949
Jones 1962Blackburn 1973Marllave 1975WMe1977Richardson and Washington 1980Richardson and Bond 11
Richardson unpubl. dataPerlmutter 1939Khan 1972Lund and Marcy 1975Colton and Marak 1969Khan 1972Blackburn 1973Zvjagina 1963Khan 1972Khan and Faber 1974
Sizes illustrated
3.0,4.7,6.9 mm NL, 7,3, 9.3,13.6 mm SL4,1 mm SL4,8,11,14 mm TL3.0,4,7,6,0 mm NL, 7,2, 9.9,11.8 mm SL
12.2mm SL10,14,19,15.5 mm TL3.0,6.3 mm NL, 8.5, 11.5, 15.4, 16.6 mm SL3.0, 4.8 mm SL3.7,3.9,6.9 mm NL, 7.6,10.4,13.8,16.5 mm SLca.4mmca.4mm4.6,5.0,7.6,8.3,9.9,10.8,18.0,24.3 mm TL5.5,9.0,10.8 mm TL5.2 mm NL, 8.2, 9.9 mm SL7.4 mm SL7.5mmSL10 mm TL5.0,5.4, 5.8, 6,7, 7.1,7.6 mm SL4.8, 7.0 mm NL, 9.1 mm SL5.7mmLarva (size not given)5.8,10,11 mm5.8,10,11 mm4.5,5.7,6.4,9.5 mm5.6,6.8,8.7 mm TL6.8mm4.08,4.2,5.7,7.0 mm
7.9,9.5 mm SL7,13,15,25,34 mm TL5.79,6.59,7.55 mm10.7,12.7,15.5 mm9mm12.2,13.9,15.9 mm TL15,18 mm7.5,11.4,17.5 mm7.1,11.6,19.2,24.8,32.5 mm7.25,10.5 mmca.20mm SL5.8,5.9,9.1 mm NL, 10.7, 11.5, 19.0 mm SL6.4,10.7,13.0 mmca. 20 mm SL10.7,13.7mm12,21 mm SLca. 20 mm SL5.0, 6.6, 8.9 mm NL, 11.0; "11.8, 19.0 mm SLca.20mmSLca,12mm11.7,' 14.5, 18.8 mm TL12.6,15,5,20 mm TL14.78,15.57,16,52 mm SL11.6,14.4,17.4,20.0 'llm3.3,8.6 mm NL, 10.9, 13.8, 15.2, 16.5, 12.5,16.6 mm SL25mm12.3mm
ca. 4 mm7,6,8.3,12.0 mm SL8,12,13mmTL4.9mmNL5.1,8.1 mm NL, 11,1 mm SL7.0,9.8, 14.2,24.0 mm SL6.6,7.0,8.8,9.8,10.4,14.2,24.0 mm SL6mm5.0,7.1,9.7, 11.8 mm TL5.4,6,1,6,8,7.5,8.5,9.2 mm TL6.8,8.5,10.5,15.2 mm TL7,0,9,5,10.7, 12.5, 14.5 mm TL7.7,10.7 mm SL12.3, 12.8, 13.6, 14.5, 16.2,32 mm12.8,14.5,17.0 mm TL12,8,14.4,17.0 mm TL

RICHARDSON: CURRENT KNOWLEDGE OF SCULPIN LARVAE
TABLE I.-Continued.
'Probably Myoxocephalus scorpius (Laroche textlootnote 6) .'Species occurs in northeast Pacific but larvae described from other areas.'Not identified below genus level.
'0As Centriderm/chthys hamatus ."Textlootnote 5."As Cottus scorpius.
Genus and species
M. scorpius'
Nautichthys oculo/asciatus
Otigocottus maculosus
O. snyderiOrthonoplas triacisPar/cetinus hoptiticusPhal/ocottus -Porocottus -Psychrolutes paradoxus
Radul/nus asprel/usR. boleo/dasRhamphocottus richardsonl
Scorpaenichthys marmoratus
S/gm/stes-Ste/gistrum -Sternias -SUag/cottus -Synch/rus _Thyr/s/cus _Trlglops murray/'T. p/ngall'
Tr/g/ops sp.
Zastlce/us _
-;Not northaast Pacific species.•As Cottus bubel/so,As Tauru/us bubelis.,As Cottus /II/jaborg/.,As Acanthocottus /II/jeborgi.As Taurulus /ll/jeborg/.
Reference
Mcintosh and Prince 1890"Mcintosh and Masterman 1897"Ehrenbaum 1904"Ehrenbaum 1905-9Koefoed 1907Rass 1949Bigelow and Schroeder 1953Khan 1972Russell 1976Blackburn 1973Marliave 1975Richardson and Washington 1980Stein 1972Stein 1973Stein 1972Bolin 1941Richardson and Washington 1980
Blackburn 1973Marliave 1975Richardson and Washington 1980Richardson and Washington 1980Blackburn 1973Marliave 1975Richardson and Washington 1980O'Connell 1953Richardson and Washington 1980
Khan 1972Ehrenbaum 1905-9Koefoed 1907Rass 1949Blackburn 1973Richardson and Washington 1980
Sizes illustrated
Larva (size not given)Larva (size not given)8.24,8.16,10,18 mm8.2,10,18 mm9.5mm7.9,9.3 mm8.2,10,18mm7.6,8.5,10.4,14.0,17.4 mm TL7.5,9.5,10,14 mm7.5,13 mm SL9.5,13,17,26 mm TL11.7 mm NL, 16.6 mm SL4.6-5.2,6.0,6.6, 9.2 mm TL4.6-5.2,6.0,6.6,9.2 mm TL4.5-4.75,5.5 mm TLca. 3-4 mmSL5.6,6.2 mm NL, 13.8, 18.6,25.6 mm SL
10.3mm SL10.5,13,14,13 mm TL4.7,7.9,9.6,10.9 mm NL, 12.6, 14.4 mm SL8.7 mm NL6.7,10 mm SL10,11.5,15 mm TL8.4 mm NL, 10.6, 11.7 mm SL5.85,6.26, 10, 17, 30, 48 mm5.3,7.5,8.6 mm NL, 8.7,10.4,13.8 mm SL
8.4, 11.6, 18.9, 23.4 mm TL18mm13,16.5,22 mm10mm8.3, 12mm SL6.9 mm NL, 15.4 mm SL
in reducing taxonomic problems. The potentialusefulness of the larval groups in providing insights into systematic relationships and evolu~ionary trends within this difficult group offishesIS also discussed. The use of larval forms offishes to elucidate systematic relationships hasbeen demonstrated in a number of groups, e.g.,ceratioids (Bertlesen 1941), myctophids (Moserand Ahlstrom 1970, 1972, 1974), gonostomatids(Ahlstrom 1974), scombroids (Okiyama andDeYanagi 1978), and serranids (Kendall 1979).
LARVAL CHARACTERS OF COTTIDS
This is a summary of important larval characters in cottids, i.e., those characters occurring only?uring the larval period which can be of most useIn distinguishing species. These characters include preopercular spine pattern, body shape, and
pigmentation. Although meristic characters maybe of prime utility in identifying cottid larvae,they persist in adults and are not considered trulylarval. Meristic characters for northeast Pacificcottids have been discussed by Richardson andWashington (1980) and Howe and Richardson.3
The purpose of this summary is to point out thekinds of larval characters that are useful foridentification and the spectrum in which thosecharacters may be exhibited since cottid larvaemanifest a wide diversity of form.
This summary is based on only the representatives of northeast Pacific cottid genera listed in
"Howe, K. D., and S. L. Richardson. 1978. Taxonomicreview and meristic variation in marine sculpins (Osteichthys:Cottidae) of the northeast Pacific Ocean. Final rep" NOAANMFS Contract No. 03-78-M02-120, 1 January 1978 to 30September 1978, 142 p. Northwest and Alaska Fisheries Center,National Marine Fisheries Service, NOAA, 2725 MontlakeBoulevard East, Seattle, WA 98112.
105

Table 1. Species from areas outside the northeastPacific are included when larvae are known because of the taxonomic information their larvaemay provide. Generic level designations are usedthroughout the text for continuity and emphasisalthough larvae of all species (number of speciesbased on the taxonomic status summary by Howeand Richardson footnote 3) in a genus may not beknown. In some cases the genera are monotypic(Chitonotus, Dasycottus, Gilbertidia, Leptocottus,Orthonopias, Paricelinus, Rhamphocottus, Scorpaenichthys) and thus larval characters of thegenus may readily be defined. At least somedevelopmental stages are known for all six speciesof Hemilepidotus, providing good generic leveldefinition. In some cases larvae ofa few, but not allspecies within a genus are known [Artedius (3species out of7); Gymnocanthus (3 of6); Hemitripterus (2 of 2 or 3); Myoxocephalus (5 of 18);Oligocottus (2 of4); Radulinus (2 of5); Triglops (3of 9)]. In those instances, constancy of larvalcharacters among species provides good indications of generic level definition. Larvae of Icelinusspp. have only been described at the generic levelas none of the eight species have yet been distinguished. For some genera, larvae are known foronly one of a few species: Blepsias (l of 2), Cottus(1 of 2 brackish water species), Icelus (l of 13),Malacocottus (1 of 5), Nautichthys (1 of 3),Psychrolutes (1 of2). In those, generic level definition may not be as precise; however, larvae of allspecies appear rather distinctive and thus may begood representatives oftheir genera. In the following summary those genera which provide the bestexamples of patterns are listed in parentheses.
Principal preopercular spines typically (18 of25genera) number 4 (Scorpaenichthys, Icelinus,Leptocottus, Enophrys) and may vary in degree ofdevelopment. Modifications of this basic patternmay occur (Myoxocephalus, ?Malacocottus) inwhich four main spines are present with one ortwo auxiliary spines. Another pattern consists ofmultiple preopercular spines, usually small, numbering up to ca. 25 (Artedius, Clinocottus). Sometimes only one spine is present (Rhamphocottus)or none (Psychrolutes, Gilbertidia). Spines inother regions of the head (particularly parietalnuchal, postocular, posttemporal-supracleithral,opercular) may also be important.
General body shape can range from ratherstubby and deep (Artedius, Enophrys) to moderately slender and elongate (lcelinus, Triglops) toglobose (?Malacocottus). The snout can be quite
106
FISHERY BULLETIN: VOL. 79, NO.1
rounded (Scorpaenichthys, Hemilepidotus) orpointed (lcelinus, Chitonotus). Snout to anuslength can be rather short, < 40% SL (standardlength) (Dasycottus), to moderately long, >60%SL (Rhamphocottus), although this can changewith development. The gut may appear tightlycompacted (Dasycottus) or be distinctively coiled(Cottus). The hindgut may trail somewhat belowthe body (Artedius, Clinocottus). Unusual gutdiverticula may be present (Artedius, Clinocottus). Pectoral fins may be noticeably elongated(Nautichthys) or fanlike early in development(Myoxocephalus).
Melanistic pigment patterns range from relatively unpigmented to heavily pigmented. Pigment may be variously present or absent over thehead, snout, cheek, jaws, cleithral base, throat.Pigment over the dorsolateral surface of the gutmay vary in intensity, ventrolateral extent, andpattern (e.g., bars, Leptocottus; distinct roundmelanophores, Enophrys). In some species theentire gut region is pigmented (Paricelinus). Theventral midline of the gut may have a distinct lineofmelanophores (Cottus, some Myoxocephalus) orbe unpigmented (Scorpaenichthys). The nape maybe distinctively pigmented (Artedius, Enophrys).The lateral body surface above the gut may beunpigmented (Chitonotus), have dorsolateral pigment not extending to the gut (Radulinus) or beentirely pigmented (Scorpaenichthys). In the tailregion posterior to the anus, pigment may beabsent (some Triglops, Dasycottus), present alongonly the ventral midline (Artedius, Chitonotus) ,present along only the ventral and dorsal midlines[Gymnocanthus, small (< 8 mm) Hemilepidotus],or present on the lateral body surface, sometimesin combination with a ventral midline series(Scorpaenichthys, Radulinus, Blepsias). Number, spacing, position, and shape of ventral midline melanophores are important as is the posterior extent of lateral pigment. Melanophoresmay variously appear along the caudal fin base(Paricelinus, Chitonotus). Pectoral fins are generally unpigmented, but some species have heavily pigmented fins (Psychrolutes, Gilbertidia) or apigment band along the fin margin (Nautichthys).
LARVAL COTTID GROUPS
Within the 25 cottid genera considered, 6 groupsof genera are apparent based on shared larvalcharacters, Le., similarity, and 6 genera do not fitinto any group (Table 2). Characters within each

RICHARDSON: CURRENT KNOWLEDGE OF SCULPIN LARVAE
TABLE 2.-Groupings of 25 cottid genera reported to occur in the northeast Pacific Ocean between Baja California and the AleutianIslands based on shared larval characters. Group characteristics were based on representative species for which larvae are known, aslisted in Table 1. Also included in the groupings is an unidentified larval type, "Cottoid Type P;' of Richardson and Washington (1980)~hich may represent a new genus.
~oup General characteristics Genera
Multiple preopercular spines, rounded snout, stubby shape, slightly trailing gut,sometimes with gut protrusions or diverticula
2 Four preopercular spines, pointed snout, moderately slender, postanal pigment whenpresent usually restricted to ventral midline
3 Four principal preopercular spines or none, rounded snout, often globose shapewith loose skin, pigmented pectoral fins
4 Four preopercular spines, rounded snout, relatively deep bodied, ca. 4-5 mm NL athatching, postanal pigment dorsally, ventrally, and laterally
5 Four preopercular spines not pronounced, rounded snout, relatively slender, post·anal pigment dorsally, ventrally, laterally, probably >7 mm NL at hatching,pectoral fins unpigmented or with pigment band near margin
6 Four preopercular spines, rounded snout, relalively slender, no additional headspines, postanal pigment restricted to ventral midline
~roUPed genera Enophrys, Gymnocanthus, Myoxocephalus, Radulinus, Rhamphocottus, Hemitripterus
Artedius, Clinocottus, Oligocottus,Orthonoplas
Paricel/nus, Trlglops, Icelus,Chitonotus, leel/nus
Dasyeottus, Psyehrolutes, Gllbertldia,?Malaeoeottus, Cottoid Type A (newgenus?)
Seorpaenlehthys, Hemi/epidotus
Blepsias, Nautiehthys
Leptoeottus, Cot/us
group and of each ungrouped genus are summarized to facilitate recognition and minimize taxonomic and identification problems involvingcottid larvae. These groupings are based on complete developmental series to the extent available,but only representative figures illustrating onepoint on a developmental continuum are pre~ented (Figures 1-9). The groupings are necessarlly preliminary because not all species in allgenera are known as larvae. The groups describedbelow are not arranged in any particular order.Generic designations are used as discussed in theprevious section,
Group 1
This is the tightest group among the 25 genera.InclUded are Artedius, Clinocottus, Oligocottus,and tentatively Orthonopias (Figures 1, 2). TheUnique multiple preopercular spine pattern distinguishes it from all other groups or genera.[Although a complete series of Orthonopias hasnot been described and the spine pattern is unknown, small larvae (Figure 2) are very similarto Artedius in form and pigment characteristicsand are tentatively included in this group.] ThestUbby body shape, rounded snout, and somewhattrailing gut are remarkably consistent within thegroup. Presently, identification to genus based onlarval characters is still difficult and in need ofbetter definition. Characters used to distinguishspecies (besides fin ray counts) include: number,Spacing, and shape of ventral midline melanoPhores; intensity of gut pigmentation; presence ofunUsual gut diverticula; total number ofpreopercUlar spines and position of largest spines; num-
ber of spines (e.g., none, two, cluster) in theparietal and posttemporal-supracleithral regions;presence or absence of pigment on the nape orhead.
Although the multiple preopercular spine pattern persists through the larval period, adultshave four preopercular spines with the lower threereduced and the upper variously modified. Remnants of the larval serrations have been observedonly in adult A. notospilotus (Howe4
). It is unclearwhich four spines of the larvae persist in adults.
Group 2
This is also a rather cohesive group (Figure 3)consisting of slender forms with pointed snoutsand four prominent preopercular spines [Paricelinus, Triglops, [celus (tentatively), Chitonotus,Icelinus]. This general body shape is remarkablysimilar among genera and is not found in anyother genera considered. All have a relativelyshort snout to anus distance. Postanal ventralmidline pigment is usually present (absent in onespecies of Triglops) with some additional melanophores along the caudal fin base. Dorsal midlinepigment is usually absent except for a few spots insome Icelinus and possibly a row in some latestage Triglops. Generic differences include degreeof gut pigmentation (e.g., darkest in Paricelinusand some Triglops), number and position of ventral midline melanophores, and degree of headspination (e.g., postocular spines in Paricelinusand Triglops).
4K, D, Howe, Ph.D. candidate, Department of Fisheries andWildlife, Oregon State University, Corvallis, OR 97331, pers.commun. September 1978.
107

FISHERY BULLETIN: VOL. 79. NO.1
c
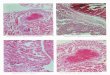
FIGURE I.-Larvae of A) Artedius harringtoni (7.3 mm SL), B) Artedius Type 2 (9.9 mm SL), C) Clinocottus acuticeps (7.6 mm SL).D) Oligocottus maculosus (9.2 mm TL) (A-C, Richardson and Washington 1980; D, Stein 1973).
108

RICHARDSON: CURRENT KNOWLEDGE OF SCULPIN LARVAE
A
B
;.\\\~\ .... "",\ '\~.\\'.. "'\.\
--- ~- .' -_. -.. ~ -.~~":--~";;'-:':.'.'-~.'"
(;>..<_..J~~~~~--""--- . .
FIGURE 2.-Larvae of A) Artedius harringtani (3.0 mm NL) and B) Orthanapias triacis (""4 mm) (A, Richardson and Washington1980; B, Bolin 1941).
The tentative placement of [celus with thisgroup is of interest as it has been considered toConstitute a distinct family, the Icelidae (Jordan1923; Greenwood et al. 1966) based on the presenceof scales in adults. Although larvae of [celus areknown to the author only from descriptions in theliterature (Table 1), they strongly resemble otherIllembers of this group in form and preopercularspine pattern. Inclusion of Paricelinus is also ofinterest as it has been considered to be a ratherdistinct and primitive form (Bolin 1947). It possesses five pelvic soft rays, the ancestral condition,Whereas the number ofsoft rays is reduced to threeor two in other members of the group.
Group 3
Group 3 (Figure 4) consists of the "psychrolutid"cottids [Dasycottus, Psychrolutes, Gilbertidia,?lvtalacocottus,S "Cottoid Type N' (new genus?)]
~entification of larvae is tentative pending resolution of:xonomic problems of adults at the generic level [see Howe andBlchardson footnote 3 and also Richardson, S. L., and C. E.Pon.d. 1978. 'I\vo unusual cottoid fishes from the northeast
aciflc. Unpubl. manuscr., 6 p. + 25 figs. (Available from senior~uthor.) (Paper presented at the American Society of Ichthyolo
sts and Herpetologists, 1978.»
often considered a separate family (Nelson 1976).["Cottoid Type N' may possibly be Psychrolutesphrictus but positive identification awaits additional specimens-see Discussion by Richardsonand Washington (1980). If it is P phrictus, larvalevidence indicates that the species is incorrectlyplaced and that a new northeast Pacific genus ofcottid is in need of description.] This group is notas cohesive as the two previous groups. The mostdistinctive character of Group 3 is the pattern ofpigmentation of the pectoral fin, a pattern notfound in any of the other genera considered. In all,at least the basal portion of the fin developspigment with the entire fin pigmented in Psychrolutes, Gilbertidia, and small « 9 mm SL) ?Malacocottus. Pigment on small (< 8 mm SL) Dasycottus is restricted to the inside surface ofthe pectoralfin base but later it develops distally on the outersurface. (The pigment band near the margin of theelongated pectoral fin of Nautichthys is a verydifferent pattern.) Only Dasycottus and ?Malacocottus develop four preopercular spines, thelatter genus with an accessory spine at the base ofthe second spine. All but the more slender Dasycottus have relatively rounded snouts and deepbodies. Both?Malacocottus and "Cottoid Type N'
109

FISHERY BULLETIN: VOL. 79, NO. I
B
FIGURE 3.-Larvae ofA) Paricelinus hopliticus (13.8 mm SL), B) Triglops sp. (15.4 mm SL), C) Icelus bicomis (25 mm), D) ChUanatuspugetensis (11.5 mm SL), E) Icelinus sp. (13.8 mm SL) (A, B, D, E, Richardson and Washington 1980; C, Ehrenbaum 1905·9).
110

RICHARDSON: CURRENT KNOWLEDGE OF SCULPIN LARVAE
------~~r-);' /
~..:::.3Y~'--- ~-J~~
c
.~
.:/~• Ii'
~.....
FIGURE 4.-Larvae of A) Dasycottus setiger (8 mm NL), B) Psychrolutes paradoxus (transfonning, 13 mm SL), C) Gilbertidia~ga~utes 03 mm SL), D) ?Malacocottus ZOIlUruS 00.4 mm SLl, E) Cottoid Type A (9.8 mm SL) (A-D, original illustrations;
,Richardson and Washington 1980).
111

develop a pronounced globose appearance with anouter bubble of skin. Small larvae of all formshave head and gut pigment and "Cottoid Type A"also has lateral pigment posterior to the anus.Some lateral pigment develops later in all butDasycottus. "Cottoid Type N' has unique "thumbtack" prickles covering the belly region. Thepelvic fins of ?Malacocottus and "Cottoid Type N'often appear to be withdrawn into pockets of skin.
Group 4
Group 4 (Figure 5) includes larvae withfour preopercular spines, conspicuously rounded
FISHERY BULLETIN: VOL. 79, NO. I
snouts, and relatively deep bodies with ratherheavy pigmentation, except at the smallest sizes(Scorpaenichthys, Hemilepidotus) , Differencesbetween genera include the longer gut and thepreanal fin fold of Scorpaenichthys and the increased head spination ofHemilepidotus (parietal,nuchal, postocular, posttemporal-supracleithral).Pigmentation is generally heavier in Scorpaenichthys than in Hemilepidotus of comparablesize. It is initially concentrated along the dorsaland ventral midlines, particularly in Hemilepidotus, filling in laterally with development. Larvae of both are neustonic.
The two genera in this group are certainly more
FIGURE 5.-Larvae of A) Scorpaenichthys m.armoratus (8.7 mm 3L), B) Hemilepidotus spinosus (11.0 mm 3L), C) H. hemilepidotus00.7 mm 3L) (A-C, Richardson and Washington 1980).
112

RICHARDSON: CURRENT KNOWLEDGE OF SCULPIN LARVAE
similar to each other than to any other cottidsconsidered, although Scorpaenichthys was givenfamilial status in the past (Jordan 1923; Tarenets1941).
Group 5
This group consists of two elongated, slenderbodied genera, Blepsias and Nautichthys (Figure6). Both hatch at a relatively large size, >7 mmNL (notochord length). Both have rounded snouts,relatively heavy pigmentation, and four preopercular spines that never become pronounced andsharp. Nautichthys (at least N. oculofasciatus)develops greatly elongated pectoral fins soon afterhatching, each of which develops a pigment bandnear its distal margin.
The genera Blepsias and Nautichthys havebeen placed in a separate family, Blepsiidae, inthe past (Jordan 1923).
Group 6
Group 6 (Figure 7) contains Leptocottus andCottus (as based on the brackish water species C.asper). They share several characters, includingthe relatively slender body, rounded snout, fourPreopercular spines, absence of other head spines,
A
ventral midline pigment along gut, and postanalpigment restricted to ventral midline. Both hatchand transform at similar sizes, ca. 3-4 mm NL andca. 10-12 mm SL, respectively. Leptocottus has aunique gut pigment pattern of bars, and Cottushas a distinctively coiled gut.
Ungrouped Genera
Enophrys (Figure 8) has four pronounced preopercular spines, rounded snout, deep stubbyshape, pigmented nape, and postanal pigmentonly along the ventral midline. It has a postocularspine and opercular spines, and a preanal fin fold.Melanophores over the gut are distinctively roundin shape and densely concentrated. This suite oflarval characters is not shared by any other genus.Enophrys bears some resemblance to Group 6(Leptocottus-Cottus) but differs too much to bepart of it. The deep body, bulging gut, and pigmented nape somewhat resemble Group 1 (Ar
tedius et a1.) but spine patterns differ drastically.Larvae of E. bubalis and E. lilljeborgi from theNorth Atlantic (Table 1) are extremely similar toE. bison from the northeast Pacific.
Gymnocanthus (Figure 8) apparently never develops pronounced preopercular spines, accordingto the literature (Table 1). Larvae of G. tricuspis
FIGURE G.-Larvae of A) Blepsias cirrhoslts (11 nun NL) and B) Nalttichthys ocltlofasciatlts (11.7 mm NL) (A, original illustration;B, Richardson and Washington 1980).
113

FISHERY BULLETIN: VOL. 79, NO. I
.,' .'
.> .~' ..,".~.
" .... "
.. :.......
". ". ,..'. '; ." .': :... ~
.:' ~; ..~.~.( ..~:..•:.;.:::~ :'~~"'::' •.~.-~
.!
A
B.: .: .... '
-------........4,. ~ ...
,,'" " . "
~-_..,..--J-
FIGURE 7.-Larvae of A) Leplocoltlls annalus (8.1 mm NL) and B) Coitus asped8.2 mm SL) (A, B, Richardson and Washington 1980).
develop three very small preopercular spines,visible only on stained specimens, and the spinesof 14-15 mm G. ventralis are barely perceptible.Gymnocanthus is relatively slender with a semipointed snout; it develops dorsal (at least G.tricuspis), ventral, and lateral midline pigment.Although it shares some characters with Myoxocephalus, grouping the two does not seem warranted. The spacing of the ventral series ofmidline melanophores somewhat resembles thesituation in Myoxocephalus. General shape ofspecies of the two genera is also similar althoughMyoxocephalus is much spinier. Some figuresindicate that Gymnocanthus has a preanal fin foldas do at least some Myoxocephalus.
Myoxocephalus (Figure 8) may have four to sixpreopercular spines, two of which may be auxiliary. Larvae are moderately slender with a semipointed snout. Two distinct larval forms (subgenera?) occur in the genus. Those with dense lateralpigment in the region posterior to the anus (M.polyacanthocephalus, M. scorpius) and those withonly ventral midline pigment postanally (M.aeneus, M. octodecemspinosus). Another relatively unpigmented species, M. quadricornis (marineform), develops a lateral midline pigment series.The spacing of the ventral midline melanophoresis similar in all species as is gut pigment. The fanshaped pectoral fin is obvious early in development (at least in some species) and a preanal fin
fold is present (in some or maybe all species). Thepattern of preopercular spination is unusual. Allspecies apparently have four principal spines butsome may have one or two posteriorly directedauxiliary spines anterior to and near the base ofthe second principal spine and/or an auxiliaryspine ventral and adjacent to the fourth principalspine (Laroche6
). Development of this unusualpattern has not been adequately described for anyspecies of Myoxocephalus and needs closer examination. Auxiliary preopercular spines are unknown in other genera except ?Malacocottus,which has an anteriorly directed accessory spineat the base of the second preopercular spine.
Radulinus (Figure 9) is a heavily pigmentedform with four preopercular spines (never pronounced), relatively pointed snout, slender body(R. boleoides is deeper bodied than R. asprellus),and long gut. It shares the characters of pointedsnout and slender body with Group 2 (ParicelinuSet al.) but differs too much (longer gut, heavierpigmentation, reduced prominence of preopercular spines) to be part of that group. It shares afew similarities with the dark species of Myoxocephalus, including a series of ventral midlinemelanophores on the tail beyond the lateral pigment (in larvae <8-9 mm NL) and a relatively
GJ. L. Laroche, Research Assistant, Gulf Coast ResearchLaboratory, East Beach Drive, Ocean Springs, MS 39564, pel's.cornmun. September 1978.
114

RICHARDSON: CURRENT KNOWLEDGE OF SCULPIN LARVAE
A
". ' ..
o .~~\ ~~.~\_--._)/.. / .;;.' ;y/. . ..
. I- , + ... . I I I 7
.. , ...-.",\,~~-~---~
~~G lIRE 8. _ Larvae of A) Enophrys bison (7.0 mm NL) l B) Gymnocanthus tricuspis (13.9 mm), C) Myoxocephalus aeneas (8.5 mm TL) I
D !vI. polyacanthocephalus (12 nun SL) (A, Richardson and Washington 1980; B, Khan 1972; C, Lund and Marcy 1975;, Original illustration).
115

FISHERY BULLETIN: VOL. 79. NO. I
A
FIGURE 9.-Larvae of A) Radulinus asprellus (10.9 nun NLl. B) Rhamphocottus richardsoni (8.4 nun NL). C) Hemitripterusuillosus (17.4 mm SL) (A, B. Richardson and Washington 1980; C. Okiyama and Sando 1976).
unpigmented area on the body above the abdominal cavity. Small larvae also resemble Scarpaenichthys except that they have a distinctlateral midline series of melanophores and soondevelop a pointed snout and more slender body.
Rhamphacattus (Figure 9) is one of the mostaberrant cottid forms. It develops only one preopercular spine, an unusual snout, a deep body,heavy pigmentation, and a preanal fin fold. Atsmall sizes, ca. 6-7 rom NL, it bears some resemblance to Scorpaenichthys in pigmentation and
116
shape, but it has a longer gut, more pigmentventrally along the head and gut, and a pigmentedpreanal fin fold. By 8-9 rom SL, the distinct shapeof R hamphocottus is obvious and spinelikeprickles develop over the body. The single specieshas been considered to represent a separate family (Jordan and Evermann 1898; Jordan 1923;Taranets 1941).
Hemitripterus (Figure 9) is also a heavily pigmented and distinct form. Based on the literature(Table 1), it has four preopercular spines, a moder-

RICHARDSON: CURRENT KNOWLEDGE OF SCULPIN LARVAE
ately pointed snout, relatively deep body, and longgut. Larvae are quite large, ca. 12-14 mm NL, athatching. The larvae of H. americanus from theAtlantic and H. villosus from Japan (Table 1) areVery similar. The heavy body pigment is a character shared with a number of apparently unrelated cottid genera. The genus was considered toConstitute a separate family, Hemitripteridae, byJordan (1923) and Taranets (1941).
DISCUSSION
The present state of cottid systematics is confused and the group and its allies are in need ofintensive study (Nelson 1976; Howe and RichardSon footnote 3). Family limits are not well defined(compare, e.g., Jordan 1923; Berg 1940; Taranets1941; Greenwood et al. 1966; Bailey et al. 1970;Nelson 1976). Some genera still need revision andPotential new species remain to be described (e.g.,Nelson 1977; Richardson and Washington 1980;liowe and Richardson footnote 3). Studies ofintergeneric relationships have been few (e.g.,Regan 1913; Taranets 1941; Bolin 1947; Watanabe1960) and these had many disagreements (Table3). Jordan and Evermann's (1898:1879-1800) comInent of North American Cottidae still has merit,"The family is an extremely varied one whichCannot be readily thrown into subordinate groups.~lrnost every species has an individuality ofIts own...."
Because of the confused state of cottid systematics it seems reasonable to consider whether thispreliminary summary of25 genera ofcottid larvaernay provide insight into intergeneric relationships within the group. Whether these phenetiCally derived larval groupings are indicative ofrelationships among cottid genera depends onWhether the groups actually possess a set ofshared, derived characters. Determination of derived states of larval characters is difficult whendealing with such a diverse group as the cottidsand their allies because the larvae ofmany speciesare still unknown and complete developmentalSeries have not been described for many other~pecies.Such determinations are further hindered1 the confused state of adult cottid systematics.t lthough an in depth analysis of derived charac-er states is beyond the scope of this study,
Consideration of several factors allows discussion~! the ?otential significance of, and possible relalonshlpS within, at least some of the larval cottid
groups described here. Larval characters such as
spine patterns, relative body form, and pigmentation have been used to demonstrate or clarifysystematic relationships in other groups offishes,e.g., scorpelarchids (Johnson 1974), gonostomatids(Ahlstrom 1974), myctophids (Moser and Ahlstrom 1974), myctophiforms (Okiyama 1974), marine teleosts in general (Ahlstrom and Moser1976), bothids (Futch 1977), scombroids (Okiyamaand Ueyanagi 1978), serranids (Kendall 1979). Inthese studies, similarity oflarval form has been in .remarkable agreement with relationships impliedfrom adult characters. Although larval charactershave not been used previously as indicatorsof relationship (i.e., based on synapomorphies)among cottids, it seems highly probable that atleast some of these characters would be as usefulin cottids as in other groups offishes. In addition,if the cottids were derived from an ancestral stockrelated to the Scorpaenidae, the most generalizedgroup in the Order Scorpaeniformes (Gill 1889;Taranets 1941; Bolin 1947) and if Bolin's (1947)ancestral cottid form is valid and Scorpaenichthysclosely resembles the primitive condition, thenprimitive or derived states of at least some larvalcharacters of cottids can be postulated. Primitivestates oflarval characters may include four strongpreopercular spines, relatively deep but compactbody, compact gut, lack of gut diverticula, possession of a preanal fin fold, rounded snout, relativelyshort pectoral fin. Derived character states mayinclude reduced size and/or numbers of preopercular spines or modification ofthe basic pattern offour, slender or globose body, trailing gut, presence of gut diverticula, no preanal fin fold, semipointed or pointed snout, elongated pectoral fin.Pigment patterns are more difficult to evaluate aspresumably they may possibly reflect responses tohabitat or may be more easily modified genetically than other morphological characters. This ideahas been generally discredited in other groupswhere larval characters have been used to implyrelationships (e.g., Ahlstrom 1974; Moser andAhlstrom 1974; Kendall 1979) as pigment patternshave substantiated findings based on other characters. Recent experiments on larvae ofthe zebrafish, Brachydanio rerio, (Milos and Dingle 1978)have demonstrated constancy in numerical regulation of melanophores which indicates larvalpigment patterns may not be as plastic as oncethought. Among the cottids, Scorpaenichthys isheavily pigmented but Enophrys, also consideredto be a relatively primitive form (Sandercock andWilimovsky 1968), is not. Heavy pigmentation
117

seems to be related to a neustonic habitat in some(e.g., Scorpaenichthys, Hemilepidotus) but notothers (e.g., Radulinus). Relative constancy ofpigment pattern (such as presence or absence oflateral pigment posterior to the anus) within agroup used in conjunction with other characters,however, may provide additional evidence forwithin-group relationships.
If this line of reasoning and these assumptionsare valid, then certain trends seem apparentwhich may be indicative of relationships. Group 1(Artedius et al.) appears to be a natural groupsharing a number of derived characters not present in any other group or genus (Le., multiplepreopercular spines, somewhat trailing gut, unusual gut diverticula, or at least bulging guts).A preanal fin fold is apparently absent and pigment pattern is relatively constant. The groupingagrees with findings of Taranets (1941), in part,and Bolin (1947), who considered the genera to beclosely related (Table 3). It seems to be a ratherspecialized group as Bolin (1947) implied, and,
FISHERY BULLETIN: VOL. 79, NO.1
based on the distinctiveness of larval characters,may warrant consideration at possibly the subfamilial level.
Group 2 (Paricelinus et al.) shares the derivedslender body form with pointed snout, and alsopossesses relative constancy of pigmentation, i.e.,no lateral pigment. Relationships among at leastsome ofthe genera in this group have been impliedpreviously (Table 3). The distinctiveness of larvalform within this group suggests a separate lineage; this group may warrant possible subfamilialstatus.
In Group 3, all but Dasycottus share a highlymodified larval form tending in degrees towardglobose. The constancy of the pigmented pectoralfin is unique among all groups or genera considered. With the possible exception ofDasycottus,the genera appear to bear at least some relationship to each other.
Group 4 is the most generalized in that a number ofprimitive character states are exhibited andrelationships cannot be assessed on given present
TABLE 3.-Intergeneric relationships of cottids as interpreted by A = Regan (1913), B = Taranets (1941), C = Bolin (1947), andD = Watanabe (1960). Included are only those 25 northeast Pacific genera for which larvae are known and discussed in this paper.Parentheses indicate a more distant relationship.
~
~., ~., ~~
.,::J ~
~., ~ €
'£ '" S ..'" ~ 0 .c:
'" ~ ~ l ~ ! s .c: ~
~.!!1 ~ g .~-., i:! ~ 8 al- s g. ~
.,~
g .9- 0~
::J .c:.!!1 0 § 0 !3 0 0 .c: <: .,<: '" ~
.c: <: .. .{;; '"0
0 ~ .g g 8 ~ ~.. g.
~ g S 8- "" ~l'l ~ §'
~ ~ !a .Q [ E E -'< Cl. .!!1~
::J ,g>~
.g ..~t3 <3 ,fi "" ~ ~
.. .. ~ ~ If .. .c: .gGenus a; c (!l (!l .Q .Q -.J 0 .r 0: 0:
ArtediusS/epsias'Chitonotus B.'(C)Ciinocottus B,'CCoitusDasyeottusEnophrys AGilbertidie' BGymnoeanthus'Hemilepidotus (C) (C)
Hemitripterus • (A),DJceUnus B,'(C) B,C (C)/ee/us B' B 0 A,BLeptoeottus B CMaJacocottus B,D BMyoxocepha/us BNautiehthys A,C,D (A),DOl/gocottus B,'C B,C A AOrthonop/es B,'C B,C B,CParieel/nusPsychrolutes B B BRadul/nus (C) C (C) CRhamphocottus 7 A ASeorpaeniehthys· A (C)Trig/ops' A A A
'Considered in family Ble~siidae by Taranets (1941). 'Considered in subfamily Gymnocanthinae by Watanabe (1960).'Arledius as Ruscarius, useeriops of Taranets (1941). ·Considered in family Hem,tripteridae by Taranets (1941).,Artedius induding Aryias, Allarledius. Astrolytes, 7 Considered in family Rhamphocottidae by Taranets (1941).
PeraslTo/rtes of Taranets (1941). 'Considered in order Notothenliformes by Taranets (1941).'Considered in subfamily Gilbertinae by Watanabe (1960). 'Considered in subfamily Triglopsinae by Watanabe (1960).
118

RICHARDSON: CURRENT KNOWLEDGE OF SCULPIN LARVAE
knowledge. Group 5 shares derived characterstates of a very elongate, slender body form andreduced prominence of preopercular spines; it alsoexhibits relative constancy in pigment patternSuggesting relationships between the genera. Therelatively large hatching size (>7 mm NL) oflarVae in Group 6 represents another specializationindicative of relationship; such a large size is notknown in any of the other genera except Hemitripterus whose larvae are ca. 12-14 mm NL athatching. Group 6 genera share a somewhatelongate form and constancy of pigmentation, Le.,lack oflateral pigment, although these charactersalone do not provide strong evidence of relationship. That the six ungrouped genera did not sharea set ofderived characters suggests that they bearno close relationship with one another.
In summary, this preliminary examination oflarval characters within 25 genera of cottids hasProvided some new insights into cottid systematics. Larval' evidence seems to support currentconcepts of generic limits in most instances (e.g.,Enophrys, Hemitripterus, Hemilepidotus) andhas indicated a potentially new northeast Pacificgenus represented by "Cottoid Type A." Larvalcharacters offer support for the distinctiveness ofsome genera (e.g., Rhamphocottus) and strongrelationships among others (e.g., the Artediusgroup). Some of the larval groupings discussedhere tend to support previously implied relationships within the cottids (compare Tables 2 and 3)but some important differences seem apparent(e.g., the distinctiveness of Group 1 demonstratedherein; the separation ofArtedius and Icelus, onceC?nsidered closely related (Jordan 1923); the relatIonship of Paricelinus, generally considered aPrimitive and rather distinct form (Bolin 1947;Sandercock and Wilimovsky 1968), with othermembers of Group 2; the apparent relationship of[celus to other genera in Group 2 and its questionable placement in a separate family (Jordan 1923;Greenwood et al. 1966); the distinctiveness of1?adulinus, previously considered related to Chitonotus and Icelinus (Bolin 1947)]. Because of theWide diversity of form among cottid larvae, theyoffer great potential for helping to clarify relationships and evolutionary trends within this difficultgroup of fishes. However, larvae of many speciesremain to be described (rearing may be the bestapproach), generic limits of larval characters~ust be defined, and developmental sequencesrncluding osteology need to be examined beforethat potential can be fully realized.
ACKNOWLEDGMENTS
Many who helped make this paper possible wereacknowledged by Richardson and Washington(1980). In addition, larvae for illustration wereprovided as follows: Dasycottus setiger, J. R. Dunn(Northwest and Alaska Fisheries Center, National Marine Fisheries Service, NOAA); Psychrolutesparadoxus, Gilbertidia sigalutes, J. Blackburn(Alaska Department of Fish and Game); ?Malacocottus zonurus, P. Wagner and G. Mueller (University of Alaska); Blepsias cirrhosus, Myoxocephalus polyacanthocephalus, A. Lamb (PacificEnvironment Institute, British Columbia) andC. Moffett (Bellingham, Wash.). B. Washingtonillustrated these specimens and provided technical assistance. N. Y. Khan granted permissionto reproduce a figure of Gymnocanthus tricuspisfrom his dissertation. J. L. Laroche (Oregon StateUniversity) provided information on preopercularspines and pigmentation on Myoxocephalus. Conversations on cottid systematics with K. Howe(Oregon State University) were particularly informative and stimulating. K. Howe and B. Washington read the manuscript and made helpfulcomments.
LITERATURE CITED
AHLSTROM, E. H.1974. The diverse patterns of metamorphosis in gonosto
matid fishes - an aid to classification. In J. H. S. Blaxter(editor), The early life history offish, p. 659-674. SpringerVerlag, N.Y.
AHLSTROM, E. H., AND H. G. MOSER.1976. Eggs and larvae offishes and their role in systematic
investigations and in fisheries. Rev. Trav. Inst. PechesMarit.40:379-398.
BAILEY, R. M., J. E. FITCH, E. S. HERALD, E. A. LACHNER, C. C.
LINDSEY, C. R. ROBINS, AND W B. SCOTT.1970. A list of common and scientific names of fishes from
the United States and Canada. 3d ed. Am. Fish. Soc.Spec. Publ. 6, 149 p.
BERG,L.S.1940. A classification offishes, both recent and fossil. Tr.
Zool. Inst. Akad. Nauk SSSR 5(2):87-517.BERTELSEN, E.
1941. The Ceratioid fishes. Ontogeny, taxonomy, distribution and biology. Dana-Rep., Carlsberg Found. 39,276 p.
BIGELOW, H. B., AND W C. SCHROEDER.
1953. Fishes of the Gulf of Maine. First Revision. U.S.Fish WildL Serv., Fish. Bull. 53, 577 p.
BLACKBURN, J. E.
1973. A survey of the abundance, distribution, and factorsaffecting distribution of ichthyoplankton in Skagit Bay.M.S. Thesis, Univ. Washington, Seattle,136 p.
BOLIN, R. L.1941. Embryonic and early larval stages of the cottid
119

fish Orthonopias triads Starks and Mann. StanfordIchthyol. Bull. 2:73-82.
1947. The evolution of the marine Cottidae of Californiawith a discussion of the genus as a systematic category.Stanford Ichthyo!. Bull. 3:153-168.
BRUUN,A.F.1925. On the development and distribution of the Norway
bullhead (Cottus lilljeborgi Collett). Cons. Perm. Int.Explor. Mer, PubI. Circonstance 88, 15 p.
BUDD, P. L.1940. Development of the eggs and early larvae of six
California fishes. Calif. Div. Fish Game, Fish Bull. 56,50 p.
COLTON, J. B., JR., AND R. R. MARAK.1969. Guide for identifying the common planktonic fish
eggs and larvae of continental shelf waters, Cape Sableto Block Island. Bur. Commer. Fish., BioI. Lab., WoodsHole, Mass., Lab. Ref. 69-9, 43 p.
CUNNINGHAM, J. T.1891. On some larval stages of fishes. J. Mar. BioI. Assoc.
U.K., New Ser., 2:68-74.EHRENBAUM, E.
1904. Eier und Larven von Fischen der deutschen Bucht.III. Fische mit festsitzenden Eiern.· Wiss. Meeresunters.Komm. wiss. Unters. Dtsch. Meere Kiel BioI. Anst.Helgo!. 3:127-200.
1905-9. Eier und Larven von Fischen. NordischesPlankton, Zool. Teil, 1:1-413.
EIGENMANN, C. H.1892. The fishes of San Diego, California. Proc. U.S.
Natl. Mus. 15:123-178.FOLLET'I', W. 1.
1952. Annotated list of fishes obtained by the CaliforniaAcademy of Sciences during six cruises of the U.S.S.Mulberry conductedby the United States Navy off centralCalifornia in 1949 and 1950. Proc. Calif. Acad. Sci.,Ser. 4, 27:399-432.
FUIMAN, L. A.1976. Notes on the early development of the sea raven,
Hemitripterusamericanus. Fish. Bull., U.s. 74:467-470.FUTCH,C.R.
1977. Larvae of Trichopsetta ventralis (Pisces: Bothiidae),with comments on intergeneric relationships within theBothidae. Bull. Mar. Sci. 27:740-757.
GILL,T.1889. On the classification of the mail-cheeked fishes.
Proc. U.S. Nat!. Mus. 11:567-592.GoRBUNOVA, N. N.
1964. Razmnozhenie i razvitie polucheshuiykh bychkov(Cottidae, Pisces). (Breeding and development ofhemilep.idotine sculpins (Cottidae, Pisces).) Tr. Inst. Okeanol.Akad. Nauk SSSR 73:235·252. [Translated from Russ.In T. S. Rass (editor), Fishes of the Pacific and IndianOceans, biology and distribution, p. 249-266. Isr. ProgramSci. Transl.,1966, 266 p.; avail. U.S. Dep. Commer., Natl.Tech. Inf. Serv., Springfield, Va., as 'IT 65-50120.)
GREENWOOD, P. H., D. E. ROSEN, S. H. WEITZMAN, ANDG.S.MYERS.
1966. Phyletic studies of teleostean fishes, with a pro·visional classification of living forms. Bull. Am. Mus.Nat. Rist. 131:341.455.
HAT'I'ORI, S.1964. Studies on fish larvae in the Kuroshio and adjacent
waters. [In Jpn., Eng!. synop.l Bull. Tokai Reg. Fish.Res. Lab. 40, 158 p.
120
FISHERY BULLETIN: VOL. 79, NO.1
JOHNSON, R. K.1974. A revision of the alepisauroid family Scopelarchidae
(Pisces: Myctophiformes). Fieldiana, Zool. 66, 249 p.JONES,A. C.
1962. The biology of the euryhaline fish Leptocottusarmatus armatus Girard (Cottidae). Univ. Calif. PubI.Zoo!' 67:321-368.
JORDAN, D. S.1923. A classification of fishes including families and
genera as far as known. Stanford Univ. Publ., Univ.Ser., BioI. Sci. 3:79-243.
JORDAN, D. S., AND B. W. EVERMANN.1898. The fishes of North and Middle America. Bull.
U.S. Natl. Mus. 47:1241-2183.KENDALL, A. w., JR.
1979. Morphological comparisons of North American seabass larvae (Pisces: Serranidae). U.S. Dep. Commer.,NOAA Tech. Rep. NMFS CIRC-428, 50 p.
KHAN,N. Y1972. Comparative morphology and ecology of the pelagic
larvae of nine Cottidae (Pisces) of the northwest Atlanticand St. Lawrence drainage. Ph.D. Thesis, Univ. Ottawa,234 p.
KHAN, N. Y., AND D. J. FABER.1974. A comparison of larvae of the deepwater and
fourhom sculpin, Myoxocephalus quadricornis L. fromNorth America. 1. Morphological development. In J.H. S. Blaxter (editor), The early life history of fish,p. 703-712. Springer.Verlag, N.Y.
KOEFOED,E.1907. Poissons. In L. P. R. d'Orleans, Croisiere ocean
ographique accompHe abord de Ill. Belgica dans Ill. Merdu Gronland 1905, p. 485-499. Charles Bulens, Brussels.
KYUSHIN,K.1968. The embryonic development and larval stages
of Hemitripterus villosus WaBas). BuB. Fac. Fish.,Hokkaido Univ. 18:277-289.
1970. Embryonic development and larvae of Gymnocanthus herzensteini Jordan and Starks. Jpn. J. Ichthyol.17:74-79.
LUND, W. A., JR., AND B. C. MARCY, JR.1975. Early development of the grubby, Myoxocephalus
aenaeus (MitchiJI). BioI. Bull. (Woods Hole) 149:373-383.MARLIAVE, J. B.
1975. The behavioral transformation from the planktoniclarval stage of some marine fishes reared in the laboratory. Ph.D. Thesis, Univ. British Columbia, Vancouver,231 p.
MCINTOSH, W C., AND A. T. MASTERMAN.1897. The life-histories of the British marine food-fishes.
C. J. Clay, Lond., 516 p.MCINTOSH, W C., AND E. E. PRINCE.
1890. On the development and life-histories of the teleostean food- and other fishes. Trans. R. Soc. Edinb.35:665-946.
MILOS, N., AND A. D. DINGLE.1978. Dynamics ofpigment pattern formation in the zebra
fish, Brachydanio reno. II. Liability of lateral line stripeformation and regulation of pattern defects. J. Exp.Zool. 205:217-224.
MISITANO, D. A.1978. Description of laboratory·reared larvae of the buf·
falo sculpin, Erwphrys bison Girard (Pisces: Cottidae).Copeia 1978:635-642.
1980. A record of internal fertilization in the roughback

RICHARDSON: CURRENT KNOWLEDGE OF SCULPIN LARVAE
sculpin Chitinotus pugetensis with descriptive notes oftheir early larvae. Copeia 1980:162-165.
MORRIS, R. W.1951. Early development of the cottid fish Clinocottus
recalvus (Greeley). Calif. Fish Game 37:281-300.MOSER, H. G., AND E. H. AHLSTROM.
1970. Development of lanternfishes (family Myctophidae)in the California Current. Part 1. Species with narroweyed larvae. Bull. Los Ang. Cty. Mus. Nat. Hist. Sci.7,145 p.
1972. Development of the lanternfish, Scopelopsis multipunctatus Brauer 1906, with a discussion of its phylogenetic posi tion in the family Myctophidae and its rolein a proposed mechanism for the evolution of photophorepatterns in lanternfishes. Fish. Bull., U.S. 70:541-564.
1974. Role of larval stages in systematic investigations ofmarine teleosts: The Myctophidae, a case study. Fish.Bull., U.S. 72:391-413.
NELSON, J. S.1976. Fishes of the world. Wiley, N.Y., 416 p.1977. Fishes of the southern hemisphere genus Neophryn
ichthys (Scorpaenifonnes: Cottoidei), with descriptions oftwo new species from New Zealand and Macquarie Island.J. R. Soc. N.z. 7:485-511.
O'CONNELL, C. P.1953. The life history of the cabezon Scorpaenichthys
marmoratus (Ayres). Calif. Dep. Fish Game, Fish Bull.93,76 p.
OKIYAMA,M.1974. The larval taxonomy of the primitive myctophifonn
fishes. In J. H. S. Blaxter (editor), The early life historyoffish, p. 609-621. Springer-Verlag, N.Y.
OKIYAMA, M., AND H. SANDO.1976. Early life history of the sea raven, Hemitripterus
villosus, (Hemitripterinae, Cottidae) in the Japan Sea.[In Jpn., Eng!. abstr.] Bull. Jpn. Sea Reg. Fish. Res.Lab. 27:1-10.
O!{JYAMA, M., AND S. UEYANAGI.1978. Interrelationships of scomhroid fishes: an aspect
from larval morphology. Bull. Far Seas Fish. Res. Lab.(Shimizu) 16:103-113.
PEDEN,A. E.1978. A systematic revision of the hemilepidotine fishes
(Cottidae). Syesis 11:11-49.PERLMUTI'ER, A.
1939. Section 1. An ecological survey of young fish andeggs identified from tow-net collections. In A biologicalsurvey of the salt waters of Long Island, 1938, Part II,p. 11-71. N.Y. Conserv. Dep., Supp!. 28th Annu. Rep.,1938, Salt-water Surv. 15.
QUAST,J. C. '1965. Osteological characteristics and affinities of the
hexagrammid fishes, with a synopsis. Proc. Calif. Acad.Sci., Ser. 4, 31:563-600.
RASS, T. S.1949. The composition of fish fauna of the Barents Sea
and the systematical characters of the fish eggs andlarvae. [In Russ.] Tr. Vses. Nauchno-Issled. Inst.Morsk. Rybn. Khoz. Okeanogr. 17:7-65.
REGAN, C. T.1913. The osteology and classification of the teleostean
fishes of the order Scleroparei. Ann. Mag. Nat. Hist.,,Ser. 8, 11:169-184.
RICHARDSON, S. L., AND B. B. WASHINGTON.1980. Guide to identification of some sculpin (Cottidae)
larvae from marine and brackish waters off Oregon andadjacent areas of the northeast Pacific. U.S. Dep.Commer., NOAA Tech. Rep. NMFS CIRC-430, 56 p.
RUSSELL, F. S.1976. The eggs and planktonic stages of British marine
fishes. Acad. Press, Lond., 524 p.
SANDERCOCK, F. K., AND N. J. WILIMOVSKY.1968. Revision of the cottid genus Enophrys. Copeia
1968:832-853.
STEIN, R.1972. Identification of some larval Pacific cottids. M.S.
Thesis, Humboldt State Coli., Arcata, Calif., 41 p.1973. Description of laboratory-reared larvae of Oligocot
tus maculosus Girard (Pisces:Cottidae). Copeia 1973:373-377.
TARANETS, A. Y.1941. K klassifikatsii iproiskhozhdenifbychkov semeystva
Cottidae. (On the classification and origin of the familyCottidae.) Izvestiya Akad. Nauk SSSR. Otd. Bio!. 1941(3):427-447. [Translated from Russ., Inst. Fish., Univ.B.C., Mus. Contrib. 5, 28 p.)
WARFEL, H. E., AND D. MERRIMAN.1944. The spawning habits, eggs, and larvae of the sea
raven, Hemitripterus americanus, in southern NewEngland. Copeia 1944:197-205.
WATANABE, M.1960. Fauna Japonica Cottidae (Pisces). Tokyo News
Service, 218 p.WHITE, W. A.
1977. Taxonomic composition, abundance, distributionand seasonality of fish eggs and larvae in Newport Bay,California. M.S. Thesis, California State Univ., fullerton, 107 p.
ZVJAGINA, O. A.1963. Materialy po razmnozheniyu i razvituju ryb morya
Laptevykh 2. Ledovitomorskaya rogatka, i 3. AziatskayaKoryushka. (Materials on the reproduction and development of fish of the Laptev Sea 2. Arctic sculpin and3. Asiatic smelt.) [In Russ., Eng!. summ.] Tr. Inst.Okeanol. 62:3-12. [Eng!. trans!. in Lib. Nat. Mus. Can.,Ottawa.]
121