Embed Size (px)
Citation preview

DOI: 10.1126/science.1180310 , 1668 (2009); 326Science
et al.Xiao Tao,Resolution
Channel Kir2.2 at 3.1 Å+Inward-Rectifier KCrystal Structure of the Eukaryotic Strong
This copy is for your personal, non-commercial use only.
. clicking herecolleagues, clients, or customers by , you can order high-quality copies for yourIf you wish to distribute this article to others
. herefollowing the guidelines can be obtained byPermission to republish or repurpose articles or portions of articles
(this information is current as of July 13, 2010 ):The following resources related to this article are available online at www.sciencemag.org
http://www.sciencemag.org/cgi/content/full/326/5960/1668version of this article at:
including high-resolution figures, can be found in the onlineUpdated information and services,
http://www.sciencemag.org/cgi/content/full/326/5960/1668/DC1 can be found at: Supporting Online Material
http://www.sciencemag.org/cgi/content/full/326/5960/1668#otherarticles, 30 of which can be accessed for free: cites 67 articlesThis article
5 article(s) on the ISI Web of Science. cited byThis article has been
http://www.sciencemag.org/cgi/content/full/326/5960/1668#otherarticles 3 articles hosted by HighWire Press; see: cited byThis article has been
http://www.sciencemag.org/cgi/collection/biochemBiochemistry
: subject collectionsThis article appears in the following
registered trademark of AAAS. is aScience2009 by the American Association for the Advancement of Science; all rights reserved. The title
CopyrightAmerican Association for the Advancement of Science, 1200 New York Avenue NW, Washington, DC 20005. (print ISSN 0036-8075; online ISSN 1095-9203) is published weekly, except the last week in December, by theScience
on
July
13,
201
0 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

Crystal Structure of the EukaryoticStrong Inward-Rectifier K+ ChannelKir2.2 at 3.1 Å ResolutionXiao Tao, Jose L. Avalos, Jiayun Chen, Roderick MacKinnon*
Inward-rectifier potassium (K+) channels conduct K+ ions most efficiently in one direction, into thecell. Kir2 channels control the resting membrane voltage in many electrically excitable cells, andheritable mutations cause periodic paralysis and cardiac arrhythmia. We present the crystalstructure of Kir2.2 from chicken, which, excluding the unstructured amino and carboxyl termini, is90% identical to human Kir2.2. Crystals containing rubidium (Rb+), strontium (Sr2+), andeuropium (Eu3+) reveal binding sites along the ion conduction pathway that are both conductiveand inhibitory. The sites correlate with extensive electrophysiological data and provide astructural basis for understanding rectification. The channel’s extracellular surface, with largestructured turrets and an unusual selectivity filter entryway, might explain the relative insensitivityof eukaryotic inward rectifiers to toxins. These same surface features also suggest a possibleapproach to the development of inhibitory agents specific to each member of the inward-rectifierK+ channel family.
In 1949 Bernard Katz introduced the term“anomalous rectification” to distinguish theK+ currents he observed in frog skeletal muscle
from the “delayed rectification” K+ currents ofthe squid axon action potential (1, 2). Today weknow that “delayed rectifiers” are a subset of thelarge family of voltage-dependent K+ (Kv) chan-nels, whereas “anomalous rectifiers” are membersof a different family of channels more commonlyknown as inward-rectifier K+ (Kir) channels (3).The name “inward rectifier” refers to a fundamen-tal ion-conduction property exhibited to a greateror lesser degree by all members of the family:Given an equal but opposite electrochemical driv-ing force, K+ conductance into the cell far ex-ceeds conductance out of the cell. Thus, Kirchannels are analogous to one-way conductors,or diodes, in solid-state electronic devices.
Electrophysiological experiments have shownthat inward rectification is a consequence of voltage-dependent pore blockage by intracellular multi-valent cations, especially Mg2+ and polyamines(4–8). At internal negative (hyperpolarizing) mem-brane voltages, the blocking ions are cleared fromthe pore so that K+ conducts, whereas at internalpositive (depolarizing) membrane voltages theblocking ions are driven into the pore from thecytoplasm so that K+ conduction is blocked. Asa result, Kir channels are conductive when anexcitable cell is at rest and nonconductive dur-ing excitation. This property is thought to fosterenergy efficiency because it enables Kir channelsto regulate the resting membrane potential, butnot dissipate the K+ gradient during an actionpotential (3).
A central mechanistic question is, why areKir channels blocked by intracellular multi-
valent cations? Mutational studies have identi-fied several amino acids that confer sensitivityto blocking ions (9–19), but a structural descrip-tion of these sites has remained elusive. Struc-tures of prokaryotic Kir channels, because oftheir low sequence similarity to eukaryotic Kirchannels, do not contain the specific amino acidsthat are known to underlie blockage and rectifi-cation (20, 21).
Another long-standing puzzle in eukaryoticKir channel studies is their relative insensitivityto natural toxins that typically inhibit other K+
channels (22–24). Snake, spider, and scorpionvenoms, for example, contain numerous toxinsagainst various Kv channels and Ca2+-activatedK+ channels (25–27). By contrast, Kir channeltoxins are rare, and no specific toxins againstKir2 channels have been discovered.
Eukaryotic Kir channels as a molecularfamily. The eukaryotic Kir channels contain sev-eral amino acid sequence motifs and conservedamino acids that are essential to their functionalproperties (Fig. 1). For example, in most otherK+ channels the selectivity filter comprises the“canonical” filter sequence TXGYGDX, whereX represents an aliphatic amino acid (Fig. 1).The corresponding sequence in eukaryotic Kirchannels is TXGYGFR, with F sometimes re-placed by another amino acid. In light of thestructural importance of DX in the canonicalsequence, the amino acids FR signify a markedvariation on the filter sequence. Eukaryotic Kirchannels also contain an absolutely conservedpair of cysteine residues flanking the pore re-gion, which is the reentrant peptide segmentthat forms the pore helix and selectivity filterof K+ channels. Between the outer helix (thefirst transmembrane segment) and pore regionthe “turret,” though varied among inward rec-tifiers, contains the sequence HGDL that couldbe considered a signature of eukaryotic Kir chan-nels. Finally, through extensive studies combin-
ing electrophysiology and mutagenesis, severalacidic amino acids (D and E) are known to becritical to inward rectification (9–19), and motifscontaining basic amino acids (e.g., PKKR) arecritical to phosphatidylinositol 4,5-bisphosphate(PIP2) activation of Kir channels (28–35). Thesepositions are highlighted on the sequences inFig. 1.
The Kir2.2 channel from chicken is 90%identical to the human ortholog (excluding theN and C termini) and contains all of the se-quence characteristics of a strong inward recti-fier (36). Figure S1 shows that the chicken Kir2.2channel expressed in Xenopus oocytes indeedfunctions as a strong rectifier. In oocyte two-electrode voltage-clamp recordings with 98 mMKCl in the bath solution, inward currents aremuch larger than outward currents (fig. S1B).In on-cell and excised gigaseal patch recordings,channel activity is observed at hyperpolarizing(negative internal) membrane voltages but notat depolarizing (positive internal) voltages (fig.S1C). The single-channel conductance measurednear −80 mV is ~40 pS, which is very similar tothe values reported for the guinea pig and mouseKir2.2 channels (37, 38) (fig. S1D, inset). Thesharp transition between channel conductanceand nonconductance as a function of membranevoltage is characteristic of a strong rectifier (36).Upon patch excision from the oocyte surface,some outward current is observed at voltagesslightly positive to the reversal potential becausethe concentration of intracellular blockers is de-creased (fig. S1C, blue trace). However, the currentstill decreases with further depolarization (nega-tive conductance) as channels become blockedin a voltage-dependent manner; this behaviorreflects the inherent difficulty in washing awaytrace yet still active concentrations of polyaminemolecules due to their very high affinity for thepore in strong rectifiers (39, 40). Several minutesafter patch excision, the currents decrease (fig.S1C, red trace). This “run-down” reflects alteredchannel regulation mediated by kinases, phos-phatases, and lipid signaling (34, 36, 41, 42).
To obtain diffracting crystals, we removedthe intrinsically disordered N- and C-terminal re-gions. The electrophysiological recordings shownin fig. S1 were made using a similar constructwith N- and C-terminal truncations, confirmingthat the crystal structure corresponds to a function-al channel unit with strong rectifying properties.The Kir2.2 model, consisting of the cytoplasmicdomain and transmembrane channel, was refinedat 3.1 Å to a free R-factor of 0.27. A ribbondiagram in stereo shows the transmembrane pore(above) and the cytoplasmic pore (below) (Fig.2A). Lateral openings between the transmem-brane and cytoplasmic pores, at the level of thelipid membrane headgroup layer, contain manyarginine and lysine residues. The high density ofpositive charges makes it unlikely that K+ ionswould pass through these openings (fig. S2). Thestructure is therefore consistent with mutagen-esis studies, which support the conclusion that
Laboratory of Molecular Neurobiology and Biophysics, RockefellerUniversity, Howard Hughes Medical Institute, 1230 York Avenue,New York, NY 10065, USA.
*To whom correspondence should be addressed. E-mail:[email protected]
18 DECEMBER 2009 VOL 326 SCIENCE www.sciencemag.org1668
RESEARCH ARTICLES
on
July
13,
201
0 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

the ion pathway extends across the full length ofthe transmembrane and cytoplasmic pores (9–19).The overall architecture is similar to that of pro-karyotic Kir channels but with a notable differ-ence: The Kir2.2 channel contains prominent,highly structured turrets on the extracellular faceof the channel. These surround, as if to protect,the pore entryway.
The selectivity filter. At a detailed struc-tural level, Kir2.2 is quite different from pro-karyotic Kir channels owing to minimal (<20%)sequence conservation. The cysteine pair that isabsolutely conserved among eukaryotic Kir chan-nels creates a circularized pore region throughcovalent linkage of the segment preceding thepore helix (C123) to the segment following theselectivity filter (C155) (Fig. 2B). The existenceof a disulfide bond was correctly predicted onthe basis of mutagenesis studies: Mutation ofthe corresponding cysteines in Kir2.1 led to theabsence of currents even though expressed pro-tein was detectable by Western blot analysis(43, 44). Application of 10 mM dithiothreitol
(DTT) or reduced glutathione to the outside ofcells expressing the wild-type channels did notaffect currents. From these two observations itwas concluded that a disulfide bridge must beessential for proper folding, but apparently notfor function (43, 44). The structure provides analternative interpretation. The disulfide bridge isburied beneath the protein surface at the level ofthe membrane interface. Furthermore, the Kir2.2channel was purified and crystallized in the pres-ence of 20 mM DTT and 3 mM TCEP [(tris(2-carboxyethyl)phosphine)], and yet the disulfidebridge remained intact. It is therefore possiblethat the disulfide bridge remains intact upon ex-posure to moderate concentrations of DTT andthat the bridge may be important for channelfunction.
The pore region is further stapled togetherby an ionized hydrogen bond between R149in the filter sequence TXGYGFR and E139(Fig. 2, B and C). The Glu O-e to Arg N-hdistance is 2.4 Å, compatible with an energet-ically strong interaction. Mutations altering
this interaction are known to alter channel func-tion (45, 46). On the basis of studies with con-catenated subunits, the salt bridge was thoughtto be intersubunit, but the crystal structure showsthat this interaction ties together two segments ofthe pore region within a single subunit (46).
Despite the presence of substantially differ-ent protein contacts surrounding the selectivityfilter, the main-chain structure of the filter inKir2.2 is the same as in other K+ channels (47).For example, the main-chain root mean squaredeviation between Kv1.2 and Kir2.2 is 0.4 Å,which is within the margin of certainty to dis-criminate atomic positions with 3.1 Å diffrac-tion data (Fig. 2, D and E) (20, 48, 49). Onestructural difference near the filter could possi-bly account for important pharmacological dif-ferences between Kir and other K+ channels. Inthe canonical filter sequence, the Asp (D) res-idue in the filter sequence is buried, creating aflat surface surrounding the filter opening. Bycontrast, in Kir channels, the Phe (F) residue atthe corresponding position projects directly into
Fig. 1. Key residues in eukaryotic Kir channels. Sequence alignment ofchicken Kir2.2 (GI: 118097849), human Kir2.2 (GI: 23110982), human Kir2.1(GI: 8132301), human Kir1.1 (GI: 1352479), human Kir3.1 (GI: 1352482),human Kir3.4 (GI: 1352484), human Kir6.1 (GI: 2493600), human Kir7.1(GI: 3150184), KirBac1.1 (GI: 33357898), KcsA (GI: 39654804), and ratKv1.2 (GI: 73536156). For all the Kir sequences, only the core regioncorresponding to the expressed protein and atomic structure of Kir2.2 isincluded in the alignment. For Kv1.2, only the transmembrane poreregion is shown. Secondary-structure elements are indicated above the
sequences, and the turret is colored orange. Residues discussed in thetext are highlighted in red (acidic residues), green (two disulfide-bondedcysteines), cyan (the inner helix bundle activation gate), purple (conservedresidues among the turrets of eukaryotic Kir channels), orange (theselectivity filter and E139), and yellow (critical residues for channel-PIP2interactions). Abbreviations for the amino acid residues are as follows: A,Ala; C, Cys; D, Asp; E, Glu; F, Phe; G, Gly; H, His; I, Ile; K, Lys; L, Leu; M,Met; N, Asn; P, Pro; Q, Gln; R, Arg; S, Ser; T, Thr; V, Val; W, Trp; andY, Tyr.
www.sciencemag.org SCIENCE VOL 326 18 DECEMBER 2009 1669
RESEARCH ARTICLES
on
July
13,
201
0 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
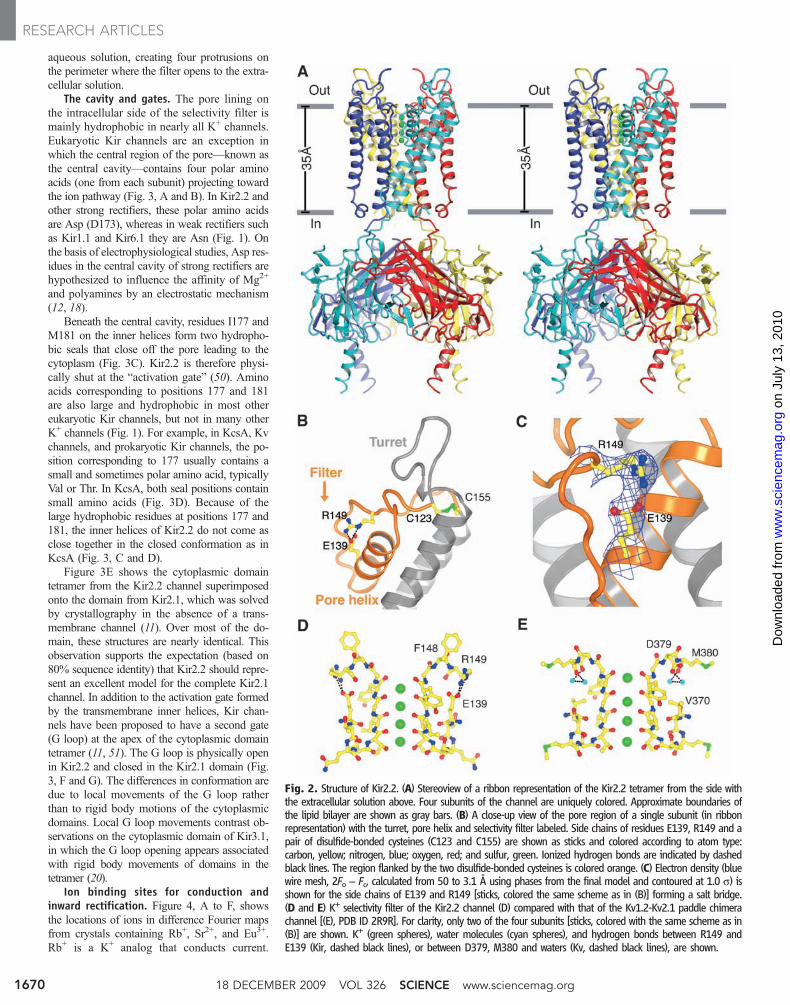
aqueous solution, creating four protrusions onthe perimeter where the filter opens to the extra-cellular solution.
The cavity and gates. The pore lining onthe intracellular side of the selectivity filter ismainly hydrophobic in nearly all K+ channels.Eukaryotic Kir channels are an exception inwhich the central region of the pore—known asthe central cavity—contains four polar aminoacids (one from each subunit) projecting towardthe ion pathway (Fig. 3, A and B). In Kir2.2 andother strong rectifiers, these polar amino acidsare Asp (D173), whereas in weak rectifiers suchas Kir1.1 and Kir6.1 they are Asn (Fig. 1). Onthe basis of electrophysiological studies, Asp res-idues in the central cavity of strong rectifiers arehypothesized to influence the affinity of Mg2+
and polyamines by an electrostatic mechanism(12, 18).
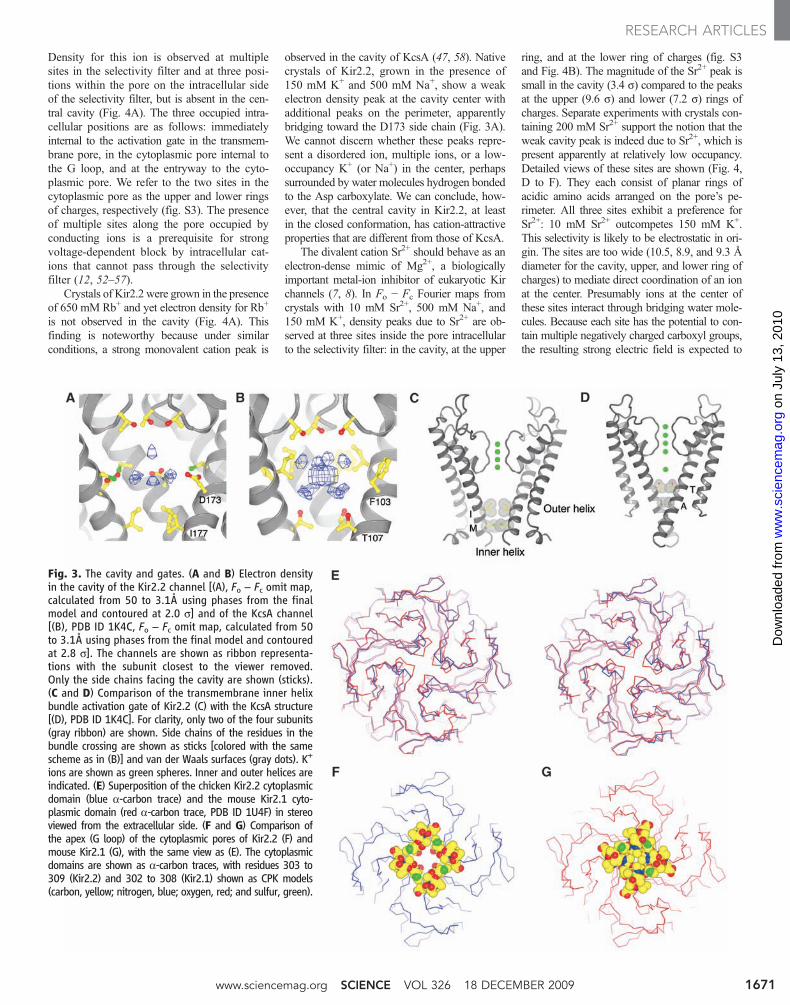
Beneath the central cavity, residues I177 andM181 on the inner helices form two hydropho-bic seals that close off the pore leading to thecytoplasm (Fig. 3C). Kir2.2 is therefore physi-cally shut at the “activation gate” (50). Aminoacids corresponding to positions 177 and 181are also large and hydrophobic in most othereukaryotic Kir channels, but not in many otherK+ channels (Fig. 1). For example, in KcsA, Kvchannels, and prokaryotic Kir channels, the po-sition corresponding to 177 usually contains asmall and sometimes polar amino acid, typicallyVal or Thr. In KcsA, both seal positions containsmall amino acids (Fig. 3D). Because of thelarge hydrophobic residues at positions 177 and181, the inner helices of Kir2.2 do not come asclose together in the closed conformation as inKcsA (Fig. 3, C and D).
Figure 3E shows the cytoplasmic domaintetramer from the Kir2.2 channel superimposedonto the domain from Kir2.1, which was solvedby crystallography in the absence of a trans-membrane channel (11). Over most of the do-main, these structures are nearly identical. Thisobservation supports the expectation (based on80% sequence identity) that Kir2.2 should repre-sent an excellent model for the complete Kir2.1channel. In addition to the activation gate formedby the transmembrane inner helices, Kir chan-nels have been proposed to have a second gate(G loop) at the apex of the cytoplasmic domaintetramer (11, 51). The G loop is physically openin Kir2.2 and closed in the Kir2.1 domain (Fig.3, F and G). The differences in conformation aredue to local movements of the G loop ratherthan to rigid body motions of the cytoplasmicdomains. Local G loop movements contrast ob-servations on the cytoplasmic domain of Kir3.1,in which the G loop opening appears associatedwith rigid body movements of domains in thetetramer (20).
Ion binding sites for conduction andinward rectification. Figure 4, A to F, showsthe locations of ions in difference Fourier mapsfrom crystals containing Rb+, Sr2+, and Eu3+.Rb+ is a K+ analog that conducts current.
Fig. 2. Structure of Kir2.2. (A) Stereoview of a ribbon representation of the Kir2.2 tetramer from the side withthe extracellular solution above. Four subunits of the channel are uniquely colored. Approximate boundaries ofthe lipid bilayer are shown as gray bars. (B) A close-up view of the pore region of a single subunit (in ribbonrepresentation) with the turret, pore helix and selectivity filter labeled. Side chains of residues E139, R149 and apair of disulfide-bonded cysteines (C123 and C155) are shown as sticks and colored according to atom type:carbon, yellow; nitrogen, blue; oxygen, red; and sulfur, green. Ionized hydrogen bonds are indicated by dashedblack lines. The region flanked by the two disulfide-bonded cysteines is colored orange. (C) Electron density (bluewire mesh, 2Fo − Fc, calculated from 50 to 3.1 Å using phases from the final model and contoured at 1.0 s) isshown for the side chains of E139 and R149 [sticks, colored the same scheme as in (B)] forming a salt bridge.(D and E) K+ selectivity filter of the Kir2.2 channel (D) compared with that of the Kv1.2-Kv2.1 paddle chimerachannel [(E), PDB ID 2R9R]. For clarity, only two of the four subunits [sticks, colored with the same scheme as in(B)] are shown. K+ (green spheres), water molecules (cyan spheres), and hydrogen bonds between R149 andE139 (Kir, dashed black lines), or between D379, M380 and waters (Kv, dashed black lines), are shown.
18 DECEMBER 2009 VOL 326 SCIENCE www.sciencemag.org1670
RESEARCH ARTICLES
on
July
13,
201
0 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
Density for this ion is observed at multiplesites in the selectivity filter and at three posi-tions within the pore on the intracellular sideof the selectivity filter, but is absent in the cen-tral cavity (Fig. 4A). The three occupied intra-cellular positions are as follows: immediatelyinternal to the activation gate in the transmem-brane pore, in the cytoplasmic pore internal tothe G loop, and at the entryway to the cyto-plasmic pore. We refer to the two sites in thecytoplasmic pore as the upper and lower ringsof charges, respectively (fig. S3). The presenceof multiple sites along the pore occupied byconducting ions is a prerequisite for strongvoltage-dependent block by intracellular cat-ions that cannot pass through the selectivityfilter (12, 52–57).
Crystals of Kir2.2 were grown in the presenceof 650 mM Rb+ and yet electron density for Rb+
is not observed in the cavity (Fig. 4A). Thisfinding is noteworthy because under similarconditions, a strong monovalent cation peak is
observed in the cavity of KcsA (47, 58). Nativecrystals of Kir2.2, grown in the presence of150 mM K+ and 500 mM Na+, show a weakelectron density peak at the cavity center withadditional peaks on the perimeter, apparentlybridging toward the D173 side chain (Fig. 3A).We cannot discern whether these peaks repre-sent a disordered ion, multiple ions, or a low-occupancy K+ (or Na+) in the center, perhapssurrounded by water molecules hydrogen bondedto the Asp carboxylate. We can conclude, how-ever, that the central cavity in Kir2.2, at leastin the closed conformation, has cation-attractiveproperties that are different from those of KcsA.
The divalent cation Sr2+ should behave as anelectron-dense mimic of Mg2+, a biologicallyimportant metal-ion inhibitor of eukaryotic Kirchannels (7, 8). In Fo − Fc Fourier maps fromcrystals with 10 mM Sr2+, 500 mM Na+, and150 mM K+, density peaks due to Sr2+ are ob-served at three sites inside the pore intracellularto the selectivity filter: in the cavity, at the upper
ring, and at the lower ring of charges (fig. S3and Fig. 4B). The magnitude of the Sr2+ peak issmall in the cavity (3.4 s) compared to the peaksat the upper (9.6 s) and lower (7.2 s) rings ofcharges. Separate experiments with crystals con-taining 200 mM Sr2+ support the notion that theweak cavity peak is indeed due to Sr2+, which ispresent apparently at relatively low occupancy.Detailed views of these sites are shown (Fig. 4,D to F). They each consist of planar rings ofacidic amino acids arranged on the pore’s pe-rimeter. All three sites exhibit a preference forSr2+: 10 mM Sr2+ outcompetes 150 mM K+.This selectivity is likely to be electrostatic in ori-gin. The sites are too wide (10.5, 8.9, and 9.3 Ådiameter for the cavity, upper, and lower ring ofcharges) to mediate direct coordination of an ionat the center. Presumably ions at the center ofthese sites interact through bridging water mole-cules. Because each site has the potential to con-tain multiple negatively charged carboxyl groups,the resulting strong electric field is expected to
Fig. 3. The cavity and gates. (A and B) Electron densityin the cavity of the Kir2.2 channel [(A), Fo − Fc omit map,calculated from 50 to 3.1Å using phases from the finalmodel and contoured at 2.0 s] and of the KcsA channel[(B), PDB ID 1K4C, Fo − Fc omit map, calculated from 50to 3.1Å using phases from the final model and contouredat 2.8 s]. The channels are shown as ribbon representa-tions with the subunit closest to the viewer removed.Only the side chains facing the cavity are shown (sticks).(C and D) Comparison of the transmembrane inner helixbundle activation gate of Kir2.2 (C) with the KcsA structure[(D), PDB ID 1K4C]. For clarity, only two of the four subunits(gray ribbon) are shown. Side chains of the residues in thebundle crossing are shown as sticks [colored with the samescheme as in (B)] and van der Waals surfaces (gray dots). K+
ions are shown as green spheres. Inner and outer helices areindicated. (E) Superposition of the chicken Kir2.2 cytoplasmicdomain (blue a-carbon trace) and the mouse Kir2.1 cyto-plasmic domain (red a-carbon trace, PDB ID 1U4F) in stereoviewed from the extracellular side. (F and G) Comparison ofthe apex (G loop) of the cytoplasmic pores of Kir2.2 (F) andmouse Kir2.1 (G), with the same view as (E). The cytoplasmicdomains are shown as a-carbon traces, with residues 303 to309 (Kir2.2) and 302 to 308 (Kir2.1) shown as CPK models(carbon, yellow; nitrogen, blue; oxygen, red; and sulfur, green).
www.sciencemag.org SCIENCE VOL 326 18 DECEMBER 2009 1671
RESEARCH ARTICLES
on
July
13,
201
0 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
create a good match for a multivalent cation.Crystals containing the lanthanide Eu3+, whichwe assume to be trivalent (59), provide supportfor this hypothesis. An anomalous differenceFourier map shows that Eu3+ binds at only onesite, the upper ring of charges (Fig. 4C). Thissite appears to be more electronegative than theothers because it contains two concentric ringsof acidic amino acids, E225 and E300.
Mutagenesis studies have identified severalamino acids that, when mutated, affect the affin-ity of Mg2+ and polyamines in strong rectifiers.D173 in the cavity, E225 and E300 forming the
upper ring of charges, and D256 forming thelower ring of charges are among those known tobe important (9–19). The weak Sr2+ peak in thecavity might seem incompatible with the largeinfluence that mutations of the cavity Asp (D173)have on Mg2+ affinity. However, the channel inthe crystal is not in an applied electric field: In anelectric field imposed by a depolarized (positiveinside) membrane, we expect that the distributionof blocker occupancies among the multiple siteswill change. Specifically, we expect the blockingcations to be driven deeper into the pore towardthe cavity. In correlating the crystallographic with
electrophysiological data, it is most notable thatthe amino acids forming the Sr2+ sites in the crystalare the same amino acids that are known to affectblockage and rectification in electrophysiologyexperiments (36). Beyond providing a structuralbasis with which to explain past electrophysio-logical studies, the Kir2.2 structure also suggestsmany new experiments. For example, most studieson the mechanism of rectification have focusedon electrostatic interactions between the positivelycharged blocker and negatively charged groupson the protein. But hydrophobic interactionsbetween methylene groups of polyamine mole-
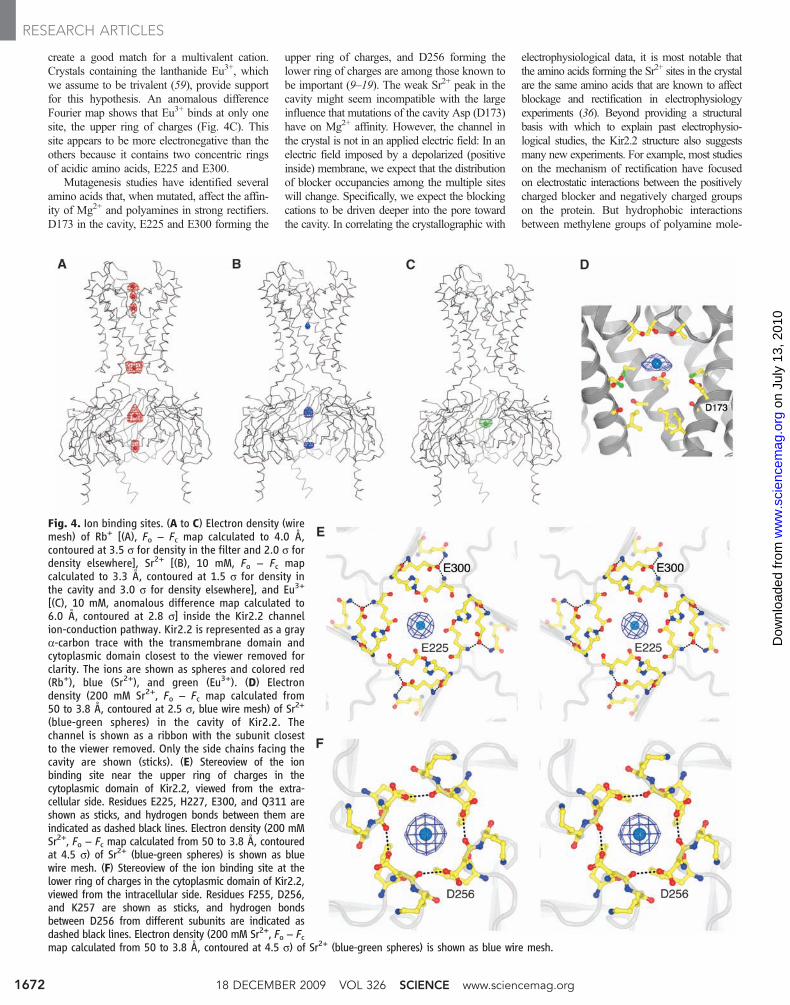
Fig. 4. Ion binding sites. (A to C) Electron density (wiremesh) of Rb+ [(A), Fo − Fc map calculated to 4.0 Å,contoured at 3.5 s for density in the filter and 2.0 s fordensity elsewhere], Sr2+ [(B), 10 mM, Fo − Fc mapcalculated to 3.3 Å, contoured at 1.5 s for density inthe cavity and 3.0 s for density elsewhere], and Eu3+
[(C), 10 mM, anomalous difference map calculated to6.0 Å, contoured at 2.8 s] inside the Kir2.2 channelion-conduction pathway. Kir2.2 is represented as a graya-carbon trace with the transmembrane domain andcytoplasmic domain closest to the viewer removed forclarity. The ions are shown as spheres and colored red(Rb+), blue (Sr2+), and green (Eu3+). (D) Electrondensity (200 mM Sr2+, Fo − Fc map calculated from50 to 3.8 Å, contoured at 2.5 s, blue wire mesh) of Sr2+
(blue-green spheres) in the cavity of Kir2.2. Thechannel is shown as a ribbon with the subunit closestto the viewer removed. Only the side chains facing thecavity are shown (sticks). (E) Stereoview of the ionbinding site near the upper ring of charges in thecytoplasmic domain of Kir2.2, viewed from the extra-cellular side. Residues E225, H227, E300, and Q311 areshown as sticks, and hydrogen bonds between them areindicated as dashed black lines. Electron density (200 mMSr2+, Fo − Fc map calculated from 50 to 3.8 Å, contouredat 4.5 s) of Sr2+ (blue-green spheres) is shown as bluewire mesh. (F) Stereoview of the ion binding site at thelower ring of charges in the cytoplasmic domain of Kir2.2,viewed from the intracellular side. Residues F255, D256,and K257 are shown as sticks, and hydrogen bondsbetween D256 from different subunits are indicated asdashed black lines. Electron density (200 mM Sr2+, Fo − Fcmap calculated from 50 to 3.8 Å, contoured at 4.5 s) of Sr2+ (blue-green spheres) is shown as blue wire mesh.
18 DECEMBER 2009 VOL 326 SCIENCE www.sciencemag.org1672
RESEARCH ARTICLES
on
July
13,
201
0 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
cules and hydrophobic residues in the channelmay be important. In particular, we might antic-ipate that when the pore opens, polyamines couldinteract strongly with the large hydrophobic aminoacids at positions 177 and 181 when the leadingamino group of the polyamine reaches into thecentral cavity (Fig. 3C) (54).
Since the earliest investigations of stronginward rectifiers, two important properties havebeen noted: a sharp transition from a conductivestate to a nonconductive (blocked) state over avery narrow voltage range, and a dependence ofthe transition on the extracellular K+ concentra-tion (60–63). Specifically, the voltage at whichthe transition occurs shifts to more depolarizingvalues as extracellular K+ concentration is in-creased. Both properties, the sharp transition(i.e., strong voltage dependence) and its de-pendence on extracellular K+, have been attri-buted to the simple notion that conducting ionsand blocking ions compete for sites in the pore(12, 52–57, 64–66). The crystallographic datapresented here support this conclusion. We ob-serve in the crystal Rb+ binding at the samesites that can bind multivalent blocking ions.Therefore, a high extracellular K+ (or Rb+) con-centration should favor occupation of the sitesby conducting ions, and a more depolarizingvoltage should be required to drive blockingions into the pore from the cytoplasm to replacethe conducting ions. Moreover, as blocking ionsenter the pore from the intracellular side, thedisplaced conducting ions must move throughthe selectivity filter to the extracellular side; thatis, movements of blocking and conducting ionsmust be coupled. Such coupling would haveenergetic consequences because movement ofan ion across the membrane voltage differenceconstitutes work. Hence, a blocking ion enteringthe pore will exhibit a voltage dependence thatresults from a combination of its own chargeand the charge of the displaced ions. This can bethe origin of strong voltage-dependent block,which can be the origin of a biologically im-portant property of strong rectifiers—their diode
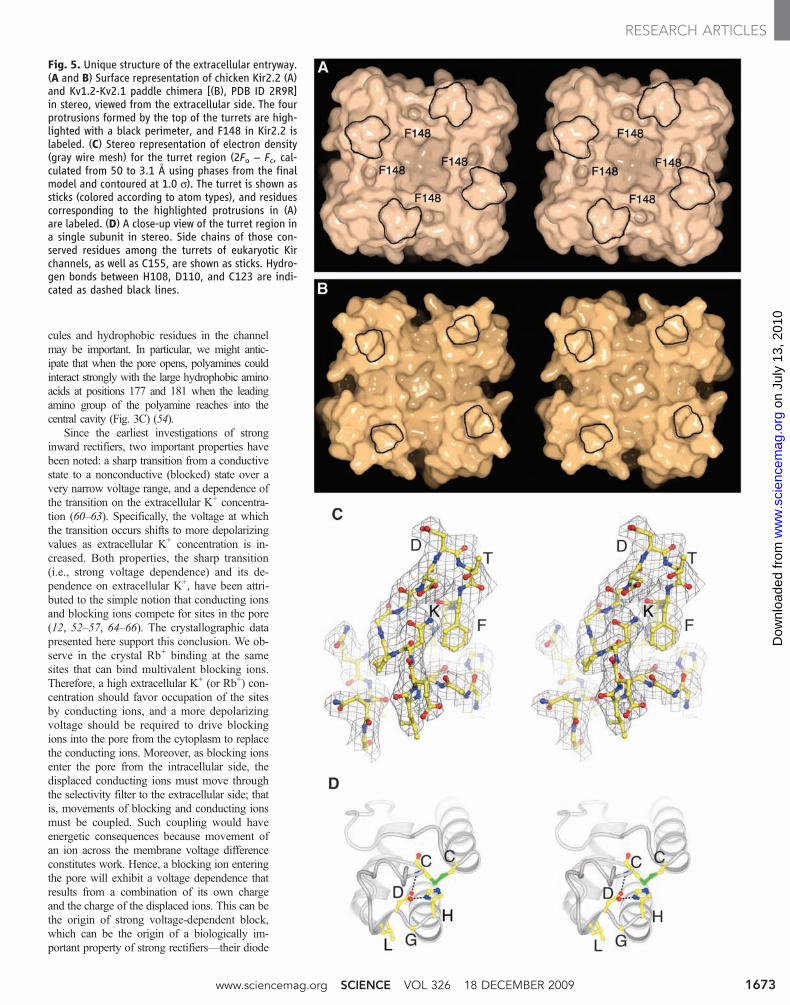
Fig. 5. Unique structure of the extracellular entryway.(A and B) Surface representation of chicken Kir2.2 (A)and Kv1.2-Kv2.1 paddle chimera [(B), PDB ID 2R9R]in stereo, viewed from the extracellular side. The fourprotrusions formed by the top of the turrets are high-lighted with a black perimeter, and F148 in Kir2.2 islabeled. (C) Stereo representation of electron density(gray wire mesh) for the turret region (2Fo − Fc, cal-culated from 50 to 3.1 Å using phases from the finalmodel and contoured at 1.0 s). The turret is shown assticks (colored according to atom types), and residuescorresponding to the highlighted protrusions in (A)are labeled. (D) A close-up view of the turret region ina single subunit in stereo. Side chains of those con-served residues among the turrets of eukaryotic Kirchannels, as well as C155, are shown as sticks. Hydro-gen bonds between H108, D110, and C123 are indi-cated as dashed black lines.
www.sciencemag.org SCIENCE VOL 326 18 DECEMBER 2009 1673
RESEARCH ARTICLES
on
July
13,
201
0 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

property of a sharp transition from a conductiveto a nonconductive state as a function of mem-brane voltage (12, 52–55, 64).
The extracellular pore entryway and phar-macology of Kir channels. Two aspects of thestructure may account for the relative insensitiv-ity of eukaryotic Kir channels, especially mem-bers of the Kir2 subfamily, to K+ channel toxins(22–24). The turrets in Kir2.2 are larger andcome closer together, constricting the pore entry-way compared to Kv1.2; and F148 in the se-quence TXGYGFR creates four protrusions onthe surface at the pore opening (Fig. 5, A andB). Thus, in Kv channels, the entryway is widerand the pore opens onto intersecting grooveswith a flat base, which form the docking surfacefor pore-blocking scorpion toxins (Fig. 5B). InKir2.2, the entryway is constricted and the groovesare absent (Fig. 5A).
Though the shape of the eukaryotic Kir chan-nel pore entryway might offer fewer opportuni-ties for inhibitory protein-protein interactions,inhibition might occur by a somewhat differentstrategy. Inhibitors of Kir1.1 and Kir3.4 channelshave been identified. A bee venom toxin, tertiapin,inhibits both of these channels (22). At 21 aminoacids in length, tertiapin is smaller than mostother venom toxins, so it might fit between theturrets more effectively. Alternatively, the turretsthemselves might form the binding site for tertiapin(67–69). At 57 amino acids, d-dendrotoxin fromthe green mamba snake is rather large, and yetit inhibits Kir1.1 channels (23). Compared totertiapin, less is known about the binding siteon the channel for d-dendrotoxin, but one aspectof its inhibition is intriguing: The blocked statereduces single-channel conductance to about 10%rather than inhibiting it completely. d-Dendrotoxinmost likely binds to the turrets but is too large tofit tightly over the pore, which would imply thatbinding to the turret may be sufficient to alter thechannel’s function.
The idea that binding to the turrets couldalter function is not surprising when one con-siders that the turret in Kir2.2 is not a loop, butforms a highly ordered structure (Fig. 5C). Thebase of the turret is formed and pinned togetherby the HGDL sequence, which with only minorvariation is found in all eukaryotic Kir channels(Figs. 1 and 5D). H108 stabilizes D110 througha hydrogen bond. The Asp (D) itself is hy-drogen bonded to the amide nitrogen of C123,which effectively holds the two ends of the tur-ret together. L111 projects from the surface of ashort 310 helix into the protein interior to makestabilizing hydrophobic interactions. Thus, theturrets are structurally important elements ofthe channel. Between the sequence HGDL andthe first Cys of the disulfide bridge, the turretsequence is highly variable among Kir channelsubtypes. The Kir2.1 channel becomes sensitiveto tertiapin if the variable sequence is mutated tobe Kir3.4-like (68). Therefore, the turrets appearto be structures through which specific inhibitionof Kir channel subtypes might be achievable
through directed evolution of specific proteinbinding partners.
Summary. This paper presents the atomicstructure of a eukaryotic Kir channel, Kir2.2, astrong inward rectifier. The sequence TXGYGFRgives rise to a K+ selectivity filter stabilized bydisulfide bridges and salt bridges that distinguisheukaryotic Kir channels. Multiple ion bindingsites on the intracellular side of the selectivityfilter can be occupied by conducting ions butexhibit higher affinity for multivalent blockingions. Thus, blocking ions entering from the cy-toplasm must displace conducting ions throughthe pore. This situation is expected to give riseto strong voltage-dependent block and diode-like conduction properties. Structural features ofthe extracellular pore entryway offer an expla-nation for the relative insensitivity of Kir chan-nels to venomous toxins and a possible approachto the development of selective Kir channelinhibitors.
References and Notes1. A. L. Hodgkin, A. F. Huxley, B. Katz, Arch. Sci. Physiol.
(Paris) 3, 129 (1949).2. B. Katz, Arch. Sci. Physiol. (Paris) 3, 285 (1949).3. B. Hille, Ion Channels of Excitable Membranes (Sinauer,
Sunderland, MA, 2001).4. B. Fakler et al., Cell 80, 149 (1995).5. A. N. Lopatin, E. N. Makhina, C. G. Nichols, Nature 372,
366 (1994).6. M. Horie, H. Irisawa, A. Noma, J. Physiol. 387, 251 (1987).7. H. Matsuda, A. Saigusa, H. Irisawa, Nature 325, 156
(1987).8. C. A. Vandenberg, Proc. Natl. Acad. Sci. U.S.A. 84, 2560
(1987).9. H. T. Kurata, W. W. Cheng, C. Arrabit, P. A. Slesinger,
C. G. Nichols, J. Gen. Physiol. 130, 145 (2007).10. Y. Fujiwara, Y. Kubo, J. Gen. Physiol. 127, 401 (2006).11. S. Pegan et al., Nat. Neurosci. 8, 279 (2005).12. D. Guo, Z. Lu, J. Gen. Physiol. 122, 485 (2003).13. Y. Kubo, Y. Murata, J. Physiol. 531, 645 (2001).14. J. Yang, Y. N. Jan, L. Y. Jan, Neuron 14, 1047 (1995).15. M. Taglialatela, E. Ficker, B. A. Wible, A. M. Brown,
EMBO J. 14, 5532 (1995).16. B. A. Wible, M. Taglialatela, E. Ficker, A. M. Brown,
Nature 371, 246 (1994).17. P. R. Stanfield et al., J. Physiol. 478, 1 (1994).18. Z. Lu, R. MacKinnon, Nature 371, 243 (1994).19. B. Fakler et al., FEBS Lett. 356, 199 (1994).20. M. Nishida, M. Cadene, B. T. Chait, R. MacKinnon,
EMBO J. 26, 4005 (2007).21. A. Kuo et al., Science 300, 1922 (2003).22. W. Jin, Z. Lu, Biochemistry 37, 13291 (1998).23. J. P. Imredy, C. Chen, R. MacKinnon, Biochemistry 37,
14867 (1998).24. Z. Lu, R. MacKinnon, Biochemistry 36, 6936 (1997).25. A. L. Harvey, B. Robertson, Curr. Med. Chem. 11, 3065
(2004).26. K. J. Swartz, R. MacKinnon, Neuron 15, 941 (1995).27. M. L. Garcia, M. Hanner, H. G. Knaus, R. Slaughter,
G. J. Kaczorowski, Methods Enzymol. 294, 624 (1999).28. D. Schulze, T. Krauter, H. Fritzenschaft, M. Soom,
T. Baukrowitz, J. Biol. Chem. 278, 10500 (2003).29. W. Z. Zeng, H. H. Liou, U. M. Krishna, J. R. Falck,
C. L. Huang, Am. J. Physiol. Renal Physiol. 282, F826(2002).
30. C. M. Lopes et al., Neuron 34, 933 (2002).31. M. Soom et al., FEBS Lett. 490, 49 (2001).32. S. L. Shyng, C. A. Cukras, J. Harwood, C. G. Nichols,
J. Gen. Physiol. 116, 599 (2000).33. H. Zhang, C. He, X. Yan, T. Mirshahi, D. E. Logothetis,
Nat. Cell Biol. 1, 183 (1999).34. C. L. Huang, S. Feng, D. W. Hilgemann, Nature 391, 803
(1998).
35. Z. Fan, J. C. Makielski, J. Biol. Chem. 272, 5388 (1997).36. P. R. Stanfield, S. Nakajima, Y. Nakajima, Rev. Physiol.
Biochem. Pharmacol. 145, 47 (2002).37. N. Takahashi et al., J. Biol. Chem. 269, 23274 (1994).38. G. X. Liu et al., J. Physiol. 532, 115 (2001).39. Z. Lu, Annu. Rev. Physiol. 66, 103 (2004).40. C. G. Nichols, A. N. Lopatin, Annu. Rev. Physiol. 59, 171
(1997).41. P. Henry, W. L. Pearson, C. G. Nichols, J. Physiol. 495,
681 (1996).42. B. Fakler, U. Brandle, E. Glowatzki, H. P. Zenner,
J. P. Ruppersberg, Neuron 13, 1413 (1994).43. H. C. Cho, R. G. Tsushima, T. T. Nguyen, H. R. Guy,
P. H. Backx, Biochemistry 39, 4649 (2000).44. M. L. Leyland, C. Dart, P. J. Spencer, M. J. Sutcliffe,
P. R. Stanfield, Pflugers Arch. 438, 778 (1999).45. K. M. Dibb et al., J. Biol. Chem. 278, 49537 (2003).46. J. Yang, M. Yu, Y. N. Jan, L. Y. Jan, Proc. Natl. Acad. Sci.
U.S.A. 94, 1568 (1997).47. Y. Zhou, J. H. Morais-Cabral, A. Kaufman, R. MacKinnon,
Nature 414, 43 (2001).48. S. B. Long, X. Tao, E. B. Campbell, R. MacKinnon, Nature
450, 376 (2007).49. S. B. Long, E. B. Campbell, R. Mackinnon, Science 309,
897 (2005).50. Y. Jiang et al., Nature 417, 523 (2002).51. S. Pegan, C. Arrabit, P. A. Slesinger, S. Choe,
Biochemistry 45, 8599 (2006).52. H. G. Shin, Z. Lu, J. Gen. Physiol. 125, 413 (2005).53. H. G. Shin, Y. Xu, Z. Lu, J. Gen. Physiol. 126, 123 (2005).54. D. Guo, Y. Ramu, A. M. Klem, Z. Lu, J. Gen. Physiol. 121,
261 (2003).55. D. Guo, Z. Lu, J. Gen. Physiol. 115, 799 (2000).56. M. Spassova, Z. Lu, J. Gen. Physiol. 112, 211 (1998).57. W. L. Pearson, C. G. Nichols, J. Gen. Physiol. 112, 351
(1998).58. Y. Zhou, R. MacKinnon, Biochemistry 43, 4978 (2004).59. A. Cotton, G. Wilkinson, Advanced Inorganic Chemistry
(Wiley, New York, ed. 5, 1988).60. D. Noble, R. W. Tsien, J. Physiol. 195, 185 (1968).61. C. A. Leech, P. R. Stanfield, J. Physiol. 319, 295 (1981).62. S. Hagiwara, M. Yoshii, J. Physiol. 292, 251 (1979).63. A. L. Hodgkin, P. Horowicz, J. Physiol. 148, 127 (1959).64. D. Guo, Z. Lu, J. Gen. Physiol. 117, 395 (2001).65. D. Oliver, H. Hahn, C. Antz, J. P. Ruppersberg, B. Fakler,
Biophys. J. 74, 2318 (1998).66. M. Spassova, Z. Lu, J. Gen. Physiol. 114, 415 (1999).67. J. P. Felix et al., Biochemistry 45, 10129 (2006).68. Y. Ramu, A. M. Klem, Z. Lu, Biochemistry 43, 10701
(2004).69. W. Jin, A. M. Klem, J. H. Lewis, Z. Lu, Biochemistry 38,
14294 (1999).70. We thank P. Hoff and members of D. Gadsby’s
laboratory (Rockefeller University) for assistancewith oocyte preparation; R. Molday (University ofBritish Columbia) for providing the anti-1D4 tag cellline; K. R. Rajashankar and K. Perry at beamline24ID-C (Advanced Photon Source, Argonne NationalLaboratory) and H. Robinson at beamline X29(National Synchrotron Light Source, BrookhavenNational Laboratory) for assistance at the synchrotron;members of the MacKinnon laboratory for assistance;and A. Banerjee, J. Butterwick, M. Whorton, andJ. Chen (Purdue University) for comments on themanuscript. J.L.A. was a fellow of the Damon RunyonCancer Research Foundation. R.M. is an investigatorin the Howard Hughes Medical Institute. The x-raycrystallographic coordinates and structure factorfiles have been deposited in the Protein Data Bankwith accession ID 3JYC.
Supporting Online Materialwww.sciencemag.org/cgi/content/full/326/5960/1668/DC1Materials and MethodsFigs. S1 to S3Table S1References
7 August 2009; accepted 16 October 200910.1126/science.1180310
18 DECEMBER 2009 VOL 326 SCIENCE www.sciencemag.org1674
RESEARCH ARTICLES
on
July
13,
201
0 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from





























