Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY, Apr. 2007, p. 3583–3596 Vol. 81, No. 70022-538X/07/$08.00�0 doi:10.1128/JVI.02306-06Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Crystal Structure of Poliovirus 3CD Protein: Virally Encoded Proteaseand Precursor to the RNA-Dependent RNA Polymerase�
Laura L. Marcotte,1 Amanda B. Wass,1 David W. Gohara,1† Harsh B. Pathak,2‡ Jamie J. Arnold,2David J. Filman,1 Craig E. Cameron,2 and James M. Hogle1*
Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, Boston, Massachusetts 02115,1 andDepartment of Biochemistry and Molecular Biology, Pennsylvania State University, University Park, Pennsylvania 168022
Received 20 October 2006/Accepted 19 January 2007
Poliovirus 3CD is a multifunctional protein that serves as a precursor to the protease 3Cpro and the viralpolymerase 3Dpol and also plays a role in the control of viral replication. Although 3CD is a fully functionalprotease, it lacks polymerase activity. We have solved the crystal structures of 3CD at a 3.4-Å resolution andthe G64S fidelity mutant of 3Dpol at a 3.0-Å resolution. In the 3CD structure, the 3C and 3D domains are joinedby a poorly ordered polypeptide linker, possibly to facilitate its cleavage, in an arrangement that precludesintramolecular proteolysis. The polymerase active site is intact in both the 3CD and the 3Dpol G64S structures,despite the disruption of a network proposed to position key residues in the active site. Therefore, changes inmolecular flexibility may be responsible for the differences in fidelity and polymerase activities. Extensivepacking contacts between symmetry-related 3CD molecules and the approach of the 3C domain’s N terminusto the VPg binding site suggest how 3Dpol makes biologically relevant interactions with the 3C, 3CD, and 3BCDproteins that control the uridylylation of VPg during the initiation of viral replication. Indeed, mutationsdesigned to disrupt these interfaces have pronounced effects on the uridylylation reaction in vitro.
Poliovirus (PV), a member of the Picornaviridae family ofRNA viruses, must simultaneously perform many tasks inside ahost cell for efficient and successful viral replication, and onlya small number of gene products are responsible for theseprocesses. The virus produces a single polyprotein that iscleaved by virally encoded proteases. Many of the viral pro-teins, including precursor proteins, play multiple roles in viralreplication.
The viral protein 3CD is a multifunctional precursor to thepoliovirus protease 3Cpro and the RNA-dependent RNApolymerase 3Dpol. The 3CD molecule retains its ability tofunction as a protease but lacks polymerase activity (20). Itsproteolytic functions include cleavages, resulting in the pro-duction of structural proteins VP0, VP1, and VP3 and non-structural proteins 3AB, 3CD, 3Cpro, and 3Dpol. In many ofthese functions, 3CD serves as a better protease than 3Cpro,suggesting that the 3D region of 3CD contributes to that ac-tivity (34).
3CD is also a crucial component of the viral replicationcomplex. The 5�-terminal region of the poliovirus genomeadopts a cloverleaf structure to which 3CD can bind in thepresence of the viral protein 3AB or the cellular proteinPCBP2 (2, 35, 49). Both forms of this ribonucleoprotein com-plex appear to play important roles in inducing RNA synthesis(49). The initiation of RNA synthesis is primed by the addition
of two uridines to the 22-amino-acid protein primer VPg (3B)at the side-chain hydroxyl of Tyr3. Positive-strand synthesis isthought to use an RNA hairpin structure as its template withinthe PV genome (37, 42). One model suggests that this cis-acting replication signal in the 2C-noncoding region of the PVgenome, cre(2C), binds to 3CD and forms a multicomponentcomplex, with 3Dpol bound to VPg (37). Uridylylated VPg isthen transferred to the 3� end of viral RNA where it is ex-tended by the polymerase to produce VPg-linked RNA. Arecently reported crystal structure of 3Dpol from foot-and-mouth-disease virus (FMDV) in complex with VPg shows howthe N terminus of VPg interacts with 3Dpol (12). Alternatively,in negative-strand synthesis, poly(A) may serve as a templatefor uridylylation, though a poly(A) template does not providemuch specificity and does not require 3CD for the uridylylationof VPg (38). Uridylylation of VPg is essential for viral repli-cation and, having no counterpart in host cell metabolism,represents an attractive therapeutic target.
Cleavage of 3CD at its Gln-Gly pair, either by 3C or by asecond molecule of 3CD, releases 3Cpro and 3Dpol. Crystalstructures of 3Cpro and 3Dpol have been determined (18, 29,48). 3Cpro, a cysteine protease, is comprised of two antiparallel,six-stranded, �-barrel domains, four helices, and a flexiblepolypeptide strand linking the two barrel domains. The shallowactive site cavity utilizes Cys147 as its nucleophile, with His40and Glu71 serving as the acid/base catalysts (44).
The initial crystal structure of poliovirus 3Dpol showed thatthe protein contains a fingers-palm-thumb arrangement com-mon in DNA and RNA polymerases (18). However, much ofthe fingers region was disordered due to steric clashes amongthe fingers regions of neighboring polymerase molecules. Thewild-type crystal-packing arrangement was stabilized by astrong intermolecular interaction named interface I. InterfaceI involves the insertion of Leu446 from the thumb of one
* Corresponding author. Mailing address: Department of BiologicalChemistry and Molecular Pharmacology, Harvard Medical School, 240Longwood Avenue, Boston, MA 02115. Phone: (617) 432-3918. Fax:(617) 432-4360. E-mail: [email protected].
† Present address: Center for Computational Biology, WashingtonUniversity School of Medicine, St. Louis, MO 63110.
‡ Present address: Department of Medical Oncology, Fox ChaseCancer Center, Philadelphia, PA 19111.
� Published ahead of print on 24 January 2007.
3583
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Nov
embe
r 20
21 b
y 18
7.87
.15.
130.

molecule into a hydrophobic pocket on the palm of anothermolecule, which is further stabilized by intermolecular saltbridges. More recently, a more accurate structure of the po-liovirus polymerase with the crystal-packing artifacts removedwas solved by mutating Leu446 and a residue involved in saltbridge formation, Arg455, to prevent the formation of in-terface I (48). In the crystal form that results from elimi-nating interface I interactions, the finger region of 3Dpol iswell ordered and wraps around to touch the thumb, forminga closed circle that is likely to be important for processiveRNA synthesis. The latter structure is most likely the bio-logically relevant one, as it closely resembles the structuresof other known viral RNA-dependent RNA polymerases (1,6, 8, 13, 25, 32).
The complete structure of 3Dpol shows the N terminus bur-ied in a pocket at the base of the fingers region. The N termi-nus participates in a network of hydrogen bonds that wasproposed to help in positioning the 3D Asp238 residue in theactive site (48). Asp238 selects for the 2�-OH group of incom-ing ribonucleoside triphosphates (rNTPs), and mutationsmade to this residue abolish polymerase activity (16). The lackof polymerase activity in poliovirus 3CD protein is thought tobe associated with the removal of the N terminus of 3Dpol fromits binding pocket, as modifications made to the N terminus of3Dpol disrupt its polymerase activity. The deletion of the first 6residues of 3Dpol or the transfer of 11 residues from the Cterminus of 3Cpro to the 3Dpol N terminus leads to a completeloss of activity, while the G1A mutation of 3Dpol shows atwofold reduction in polymerase activity (21, 43, 48).
For many years, 3CD has been a high-priority target forcrystallographic studies, but the tendency of the protein toaggregate at high concentrations has hampered these efforts.We have produced a construct with mutations made to inter-face I of 3Dpol, to the crystallographic dimer interface of 3Cpro,and to the 3Cpro active site and have used this construct tosolve the structure of 3CD to a 3.4-Å resolution. We have alsosolved the structure of the 3Dpol G64S mutant to a 3.0-Åresolution. The arrangement of residues in the polymeraseactive site of our construct is very similar to that in 3CD, 3Dpol,and the 3Dpol G64S fidelity mutant, with the exception of ahydrogen bond network involving the N terminus of 3D. Sincethe N-terminal residues of 3CD’s 3D domain are part of thecovalent linker with the 3C domain, the network is absent in3CD, but it appears to be modified relative to that of the wildtype in the 3Dpol G64S structure. These findings suggest thatchanges in molecular flexibility rather than large structuralrearrangements may be important determinants of polymeraseactivity and fidelity, as is sometimes observed in other poly-merases (19, 22, 23).
In the 3CD crystal structure, the 3D domain makes extensivecontacts with the 3C and 3D domains of neighboring mole-cules, and the N terminus of 3C lies close to the VPg bindingsite. This arrangement is consistent with a possible biologicalrole for these contacts in forming and regulating the VPguridylylation complex. Indeed, several mutations designed todisrupt these proposed interfaces have pronounced effects onthe uridylylation reaction, helping to explain how 3CD plays anessential regulatory role in the complex.
MATERIALS AND METHODS
Expression and purification of 3CD proteins. 3CD protein was expressedusing the ubiquitin fusion system developed and reported by Gohara et al. (17).BL21(DE3)/pCG1 cells were transformed with pET26Ub fusion plasmids con-taining the 3CD gene and plated on NZCYM agar enriched with kanamycin (25�g/ml), chloramphenicol (20 �g/ml), and 0.4% dextrose. The resulting colonieswere then used to inoculate 100 ml of NZCYM medium and grown overnight at30°C. This culture was used to seed 1 liter of NZCYM medium, inoculated to abeginning optical density at 600 nm (OD600) of 0.03, and grown at 37°C until theyreached an OD600 of 1. The cells were chilled to 25°C and induced with isopro-pyl-�-D-thiogalactopyranoside (IPTG) to a final concentration of 500 �M. Cellswere grown for an additional 4 h at 25°C, harvested, and pelleted. The cell pelletswere stored at �80°C. Frozen cell pellets were thawed on ice and resuspendedin lysis buffer (100 mM potassium phosphate [pH 8.0], 20% glycerol, 1 mMdithiothreitol [DTT], 0.5 mM EDTA, 60 �M ZnCl2, and protease inhibitortablets [Roche]) at a concentration of 5 ml lysis buffer per gram of cell pellet.Cells were homogenized and passed through a cell disruptor (Avestin). Poly-ethylenimine was added to the cell lysate at a concentration of 0.25% (vol/vol) toprecipitate nucleic acid. The lysate was stirred at 4°C for 30 min and thencentrifuged for 30 min at 17,500 rpm at 4°C. Granular ammonium sulfate wasadded slowly to a 40% saturation to the decanted supernatant. The solution wascentrifuged for 30 min at 17,500 rpm at 4°C. The resulting ammonium sulfatepellets were resuspended in buffer A (50 mM Tris [pH 8.0], 20% glycerol, 1 mMDTT, 0.1% NP-40, and 60 �M ZnCl2) and diluted to a final salt concentration of50 mM. The diluted sample was loaded onto a phosphocellulose column (ap-proximately 1 ml bed volume/20 mg total protein) at a flow rate of 1 ml/min. Thecolumn was washed to baseline with buffer A containing 25 mM NaCl and elutedwith a linear gradient from 25 to 350 mM NaCl in buffer A. Fractions containing3CD were pooled and again diluted to a salt concentration of 50 mM. Thediluted protein was loaded onto a Q-Sepharose column (approximately 1 ml bedvolume/120 mg total protein). The column was washed to baseline with 50 mMNaCl in buffer A and eluted with a linear gradient from 50 to 500 mM NaCl inbuffer A. Fractions containing 3CD were pooled and concentrated. The proteinwas loaded onto a HiLoad 26/60 Superdex 200 column equilibrated and elutedwith buffer A containing 350 mM NaCl in the absence of the detergent NP-40.The purified protein was concentrated to 10 mg/ml, aliquoted, and stored at�80°C.
The 3CD construct utilized for crystallization contains a series of mutationsdesigned to abrogate the activity of the protease, inhibit the formation of a 3Cpro
dimer, and disrupt interface I of 3Dpol. The wild-type protease is inactivated inthis construct by mutating the nucleophilic Cys147 to an alanine residue. In orderto disrupt interface I of 3Dpol, the 3Dpol residues Leu446 (3CD residue Leu629)and Arg455 (3CD residue Arg638) were mutated to aspartic acid residues(L446D and R455D, respectively). The interface mutations resulted in encodinga protein that could be concentrated to �15 mg/ml and was less prone toprecipitation than the wild-type 3CD protein. This 3CD protein was crystallized,apparently, in the same space group as the structure we report here. However,both the appearance and the diffraction of these crystals were poor. 3Cpro formsa dimer in the 3Cpro crystal structure that is stabilized by a number of intermo-lecular contacts between the two molecules, though there is no evidence that3Cpro forms a dimer in vivo (29). To prevent any tendency of 3Cpro to dimerize,residues Glu55, Asp58, and Glu63 were mutated to alanines, whereupon thehighest quality crystals were obtained.
Expression and purification of the 3Dpol G64S mutant. The 3Dpol G64Sconstruct utilized for crystallization contains the same interface I mutationsmade in the 3CD construct (L446D and R455D). Gly64 within the fingersdomain was mutated to a serine residue. 3Dpol protein was expressed in the samemanner as 3CD and purified similarly to 3CD, with some notable differences.Ammonium sulfate was added to a 60% saturation following polyethylenimineextraction. Phosphocellulose and Q-Sepharose steps followed. The protein wasthen loaded onto a HiLoad 26/60 Superdex 200 column equilibrated and elutedwith buffer containing 200 mM NaCl, 5 mM Tris (pH 7.5), 0.1 mM EDTA, and2 mM DTT. The purified protein was concentrated to 10 mg/ml.
Crystallization and structure solution. Crystals were grown using hangingdrop vapor diffusion at 20°C, mixed 1:1 with a well solution. Crystals of 3CD grewover 2 to 3 days over a well solution containing 2 M ammonium sulfate, 0.1 MHEPES (pH 7.0), and 0.3% Jeffamine M600 (pH 7.0). These crystals weretransferred to a corresponding precipitant solution containing 20% glycerol(vol/vol) and soaked in this solution overnight prior to flash freezing in a nitrogenstream. Crystals of 3Dpol with G64S grew overnight over a well solution contain-ing 2 M sodium acetate and 0.1 M sodium cacodylate (pH 6.8), similar tocrystallization conditions used to solve the complete structure of 3Dpol (48).
3584 MARCOTTE ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Nov
embe
r 20
21 b
y 18
7.87
.15.
130.

These crystals were transferred to a cryoprotectant containing the crystallizationsolution with 20% glycerol and were flash frozen in a nitrogen stream. In bothcases, diffraction data were collected at the 19-ID beamline at the AdvancedPhoton Source (Argonne, IL). Data were integrated, merged, and scaled usingDENZO and SCALEPACK (33) programs. Structures were solved by molecularreplacement using PHASER (28). The search models were the structures ofPV-3Cpro (PDB code 1LIN) and PV-3Dpol (PDB code 1RA6) with solvent andkey residues omitted. The model of 3CD was built using Coot (11) and refinedwith REFMAC5 (30).
VPg uridylylation assay. Full-length cre (bases 3702 to 3763 of the humanpoliovirus 1 Mahoney strain, EMBL accession ID V01149.1) was synthesizedusing a MEGAshortscript kit (Ambion) and then gel purified, extracted withphenol-chloroform, lyophilized, and resuspended in water prior to use. VPgpeptide (poliovirus protein 3B, Swiss-Prot accession ID P03300) was synthesizedby Alpha Diagnostics. 3Dpol and 3CD proteins were purified as described above,except that purification was stopped after the Q-Sepharose step and Zn2� wasomitted from buffers used in that purification step. 3Dpol and 3CD mutants wereconstructed using wild-type and inactive protease (C147G) plasmid backgrounds,respectively, obtained from the Cameron laboratory. Mutagenesis was carriedout using a QuikChange II kit (Stratagene), and mutants were verified by DNAsequencing.
Uridylylation of VPg, using cre as a template (with or without 3CD as astimulatory factor), was assayed as previously described (36), except that theconcentration of 3Dpol was increased to 2 �M and reactions were run from 1 to4 h at either 30 or 37°C. Wild-type and mutant proteins 3Dpol and 3CD were usedin various combinations to see if the mutations affected the quantity of uridyly-lated VPg that was produced. Reaction products were analyzed by Tris-tricine gelelectrophoresis. The gels were dried and subsequently analyzed by phosphor-imager, quantifying free and VPg-bound radiolabel by using Quantity One soft-ware (Bio-Rad).
Mathematical treatment of the uridylylation data. The first objective was tomeasure and normalize the amount of uridylylation activity in each sample. Theamount of labeled VPg produced in each experiment was first normalized tocorrect for the decay of the label by dividing the signal of the VPg-bound bandby the total of the bound and unbound radioactivity in the lane, then correctedfor exposure time, and finally averaged over as many replicates as were available.To correct for substrate depletion (which was usually insignificant), the fractionbound was replaced by the negative of the natural logarithm of the unbound fraction.To measure the unstimulated controls accurately, they were determined in triplicate,with bound radioactivity exposed 12 times longer than the usual period.
The final objective was to determine the degree of stimulation, specifically howmuch more uridylylation is produced by a 3Dpol mutant (or wild-type 3Dpol) anda 3CD mutant (or wild-type 3CD) together, relative to the level produced by thatparticular 3Dpol mutant (or wild-type 3Dpol) alone. Thus, values were normalizedto correct for possible variations among 3Dpol preparations by dividing by thelevel of the corresponding “unstimulated” control that lacked 3CD. Each valuelisted in Table 3, therefore, represents a degree of stimulation.
Coordinate accession numbers. The refined coordinates and data for 3CD and3Dpol are available from the Protein Data Bank (PDB entries 2IJD and 2IJF,respectively).
RESULTS AND DISCUSSION
Structure determination and refinement of 3CD and 3Dpol
G64S mutants. 3CD crystallizes in space group C2221, with twomolecules per asymmetric unit and a solvent content of 80.5%.X-ray diffraction data were collected to 3.4-Å resolution (Table1), and a molecular replacement solution was found usingknown 3Cpro and 3Dpol crystal structures as input models.Each molecule of 3CD includes all 644 amino acid residuesand 10 proposed ion-binding sites. At the beginning of struc-ture determination, the structure was built to fit global omitmaps, and noise was routinely added to the atomic coordinatesat the beginning of each refinement cycle to prevent overfittingof the data. During refinement, missing and weakly orderedparts of the model were repeatedly rebuilt into omit maps (30).Twofold noncrystallographic symmetry (NCS) restraints wereapplied. Initially, strong NCS restraints were applied to themain-chain atoms and moderate restraints to the side chains.
Later in the refinement, only weak NCS restraints were ap-plied, as this optimized the Rfree value. It should be noted thatthe twofold noncrystallographic symmetry roughly doubles theobservation-to-parameter ratio and provides an internal checkof the accuracy of the result. Translation-libration-screw (TLS)parameters were imposed, treating the two 3C and two 3Ddomains in the asymmetric unit as independent rigid bodies. Inthe final stages of refinement, individual temperature factorswere included, giving R and Rfree values of 20.0 and 23.1,respectively, with good geometry (Table 1).
The 3Dpol G64S construct crystallizes in space group P65.The G64S structure was solved to 3.0-Å resolution by molec-ular replacement, using the known 3Dpol crystal structure as aninput model. There is one molecule of 3Dpol in the asymmetricunit containing all 461 amino acid residues of the polymerase.The mutated residues and several surrounding residues wereinitially excluded from the refinement of the molecular re-placement solution to eliminate bias. TLS parameters wereincluded, and restrained individual temperature factors wereincluded only during the final stages of refinement. The final Rand Rfree values for 3Dpol G64S were 21.1 and 24.4, respec-tively, with good geometry (Table 1). Omit electron densitymaps of each structure were of high quality and were readilyinterpretable (Fig. 1b and 2).
Structural overview of 3CD. The 3CD molecule consists oftwo domains, the 3C domain and the 3D domain (Fig. 1a).These two domains do not make any direct contact with oneanother but are instead separated by a seven-residue polypep-tide linker (residues 180 to 186). The crystal lattice is formedby a network of noncrystallographic- and crystallographic-sym-metry contacts.
The 3C and 3D component domains are similar in structureto 3Cpro and 3Dpol, respectively. Like 3Cpro, the 3C domain of3CD contains two �-barrel domains and a number of helicesand loop regions. The only significant structural differencesobserved between 3Cpro and 3CD were associated with crystalpacking interactions or were located in the flexible N termini
TABLE 1. 3CD and 3Dpol data collection and refinement statistics
Data set 3CD 3Dpol G64S mutant
Space group C2221 P65Unit cell a � 208.5, b � 230.4,
c � 151.0a � b � 126.3,
c � 113.5Resolution limits (Å) 20–3.4 (3.58–3.40)a 20–3.0 (3.11–3.00)a
Total observations 249,335 110,896Unique reflections 49,539 20,317Redundancy 5.0 (4.8)a 6.0 (4.7)a
Completeness (%) 98 (98)a 99 (98)a
Rmerge (%)b 7.3 (38.7)a 8.5 (32.4)a
Rcrystc 20.0 21.1
Rfreec 23.1 24.4
No. of protein atoms 10,086 3,695No. of ions 10 0No. of waters 8 0rmsd bond length (Å) 0.011 0.010rmsd bond angle (°) 1.88 1.55
a Values in parentheses are those shown for the highest resolution shell.b Rmerge � ¥ Ih � �Ih/¥Ih over all h, where Ih is the intensity of reflection h.c Rcryst and Rfree � ¥�Fo � Fc�/¥ Fo , where Fo and Fc are observed and
calculated amplitudes, respectively. Rfree was calculated using 5% of data thatwas excluded from the refinement.
VOL. 81, 2007 CRYSTAL STRUCTURE OF POLIOVIRUS 3CD PROTEIN 3585
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Nov
embe
r 20
21 b
y 18
7.87
.15.
130.
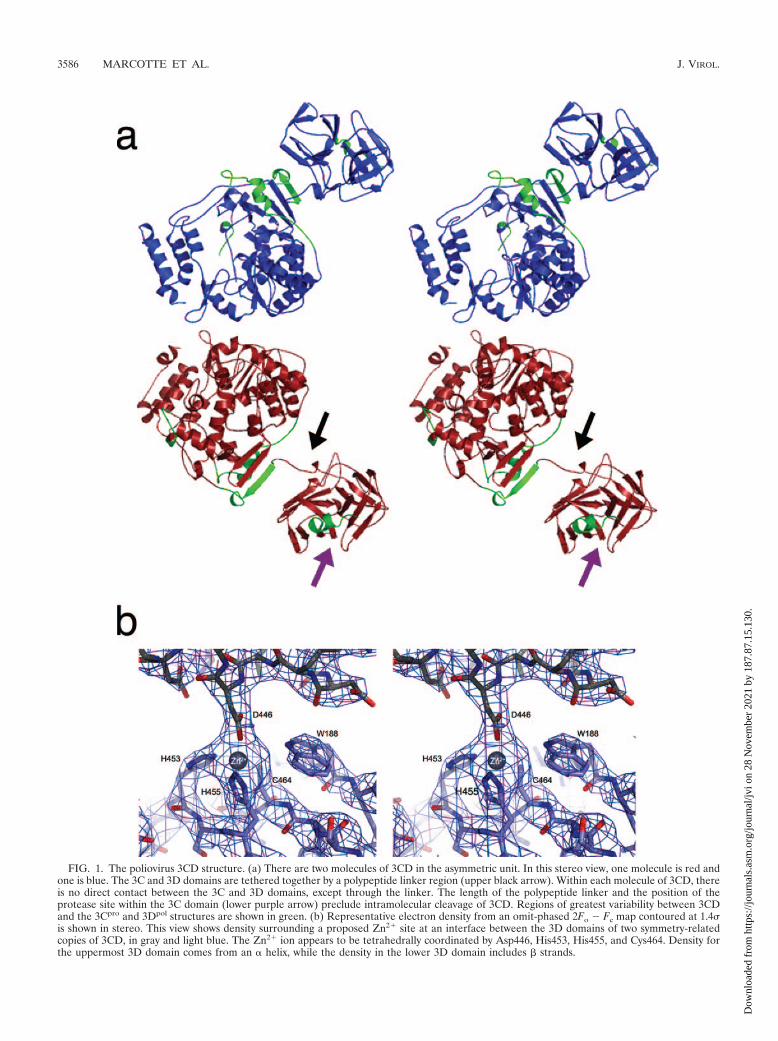
FIG. 1. The poliovirus 3CD structure. (a) There are two molecules of 3CD in the asymmetric unit. In this stereo view, one molecule is red andone is blue. The 3C and 3D domains are tethered together by a polypeptide linker region (upper black arrow). Within each molecule of 3CD, thereis no direct contact between the 3C and 3D domains, except through the linker. The length of the polypeptide linker and the position of theprotease site within the 3C domain (lower purple arrow) preclude intramolecular cleavage of 3CD. Regions of greatest variability between 3CDand the 3Cpro and 3Dpol structures are shown in green. (b) Representative electron density from an omit-phased 2Fo � Fc map contoured at 1.4is shown in stereo. This view shows density surrounding a proposed Zn2� site at an interface between the 3D domains of two symmetry-relatedcopies of 3CD, in gray and light blue. The Zn2� ion appears to be tetrahedrally coordinated by Asp446, His453, His455, and Cys464. Density forthe uppermost 3D domain comes from an � helix, while the density in the lower 3D domain includes � strands.
3586 MARCOTTE ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Nov
embe
r 20
21 b
y 18
7.87
.15.
130.

and loop regions (Fig. 1a). Main-chain atoms in the two NCS-related 3C domains (residues 1 to 180) can be superimposedwith a root mean square displacement (rmsd) of 0.29 Å, and3Cpro (ordered in residues 4 to 180) shows a 0.60-Å differencewith either copy.
The 3D domain of 3CD adopts the same fingers-palm-thumb structure shared by other RNA-dependent RNA poly-merases. The two NCS-related 3D domains (residues 184 to644) are very similar to each other, having an rms difference ofonly 0.37 Å along the main chain, and to the 3Dpol domain(with differences of 0.49-Å and 0.67-Å rms, respectively). Thislevel of difference between the coordinates provides an esti-mate for the uncertainty in atomic position, combined with theamount of local structural variability. Though the estimate oferror that comes from molecular replacement may be an un-derestimate, it does provide a way to distinguish between bet-ter ordered or structurally conserved parts of the molecule.Not unexpectedly, the N-terminal residues of the 3D domain
that comprise part of the polypeptide linker differ in positionand conformation from the corresponding residues of 3Dpol,where the cleaved N terminus is inserted into a binding pocketand extensively hydrogen bonded. Surprisingly, only the firstthree residues of the 3D domain are arranged in a qualitativelydifferent way. Smaller but statistically significant changes alsoinvolve residues 190 to 198 of 3CD (3Dpol residues 7 to 15),possibly due to the D-D� dimerization of 3D that is seen in the3CD structure, wherein the Arg7 side chain appears to pull itsmain-chain strand out of position while binding to Asp260 inthe opposite monomer. Additional positional differences occurwithin the fingers domain of the polymerase (3CD residues 226to 251; 3Dpol residues 43 to 68), where several side chains,including Phe242 and Tyr245 (3Dpol residues Phe59 andTyr62), have shifted by more than 2 Å. A third area of vari-ability lies within the pinky domain of 3Dpol (3CD residues 312to 319; 3Dpol residues 129 to 136). The maximum differences inthese regions range from 1.6 to 2.0 Å.
FIG. 3. Structure of the polypeptide linkers between the 3C and 3D domains of the two molecules of 3CD in the asymmetric unit. An omit mapcontoured at 1.4 shows weak density in the linker regions, though the overall path of the backbone is clear. The linker regions of the two 3CDmolecules in the asymmetric unit differ in structure, confirming that the linker is only weakly ordered. The two linkers are shown in similarorientations, with the 3C domain on the left and the 3D domain on the right.
FIG. 2. Hydrogen-bonding interactions around the N terminus of 3Dpol G64S, shown in stereo. An omit-phased 2Fo � Fc map contoured at1.4 shows density including the G64S mutation. Ser64 forms a hydrogen bond with Glu2, which in turn interacts with Gln4 of the N terminus.Gly1 maintains contacts with Ala239. Additional hydrogen bonding interactions are formed between the backbone of the N terminus and thebackbone of a neighboring � strand.
VOL. 81, 2007 CRYSTAL STRUCTURE OF POLIOVIRUS 3CD PROTEIN 3587
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Nov
embe
r 20
21 b
y 18
7.87
.15.
130.
3588
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Nov
embe
r 20
21 b
y 18
7.87
.15.
130.

The electron density of the polypeptide linker between the3C and 3D domains of 3CD is weak and differs significantlybetween the two NCS-related copies of the linker (Fig. 3). Thepresence of a visible electron density along the main chainimplies that the linker is not entirely disordered. Nevertheless,the existence of at least two distinctly different conformationssuggests that they are likely to be similar in energy and that thebarrier to their interconversions is likely to be low. The flexi-bility of the linker and its accessibility to the protease activesite may be relevant to the timing of 3CD cleavage (see below).Although the residues comprising this region were modeledinto the density as accurately as possible, it was not possible tofind an entirely satisfactory low-energy conformation for eithercopy of the linker. This, together with the lack of strong densityin this region, suggests that the linker is not well ordered.Indeed, the presence of a flexible linker may be necessary forthe proteolytic cleavage of 3CD into 3Cpro and 3Dpol, since astable association of the linker with either domain might makethe cleavage site inaccessible. Contrary to a previous sugges-tion, the linker seen in the 3CD structure is not long enoughfor the cleavage of 3CD at its intramolecular Gln-Gly site to beautocatalytic (7). After cleavage has taken place, the N-termi-nal residues of 3Dpol have a stable, well-characterized bindingsite, but the final three residues at the C terminus of 3Cpro
become disordered (29).Formation of interfaces between 3CD molecules. The sym-
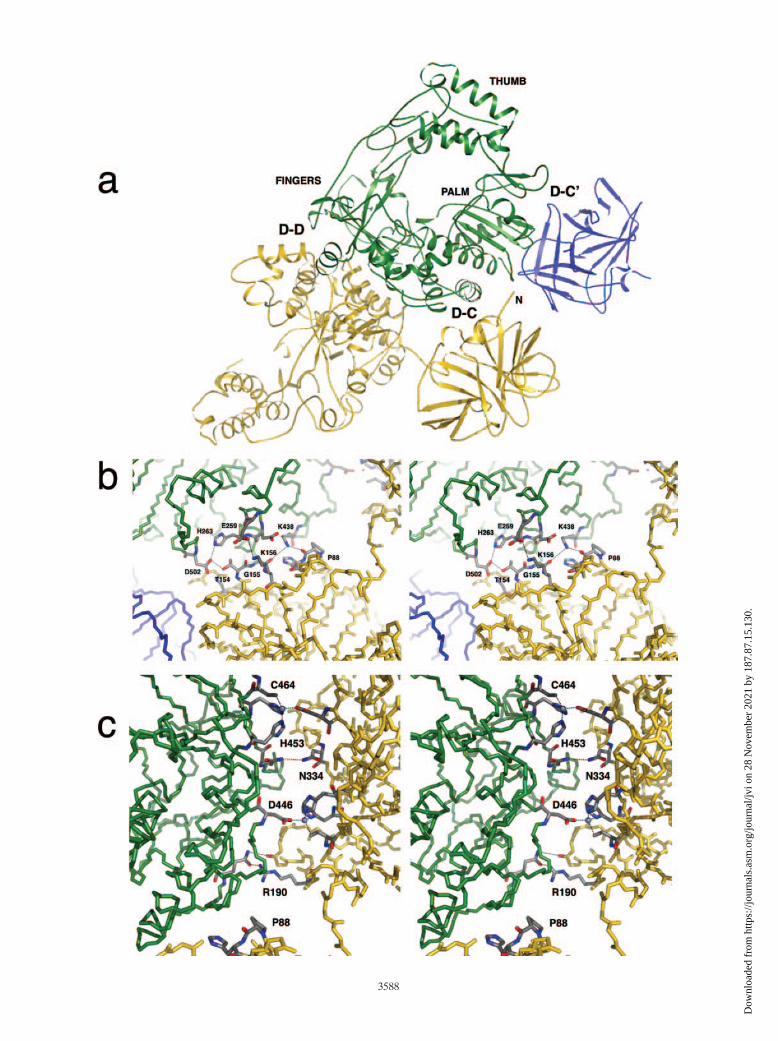
metry-related copies of 3CD form a number of crystal-packingcontacts, some of them quite extensive. One notable interfaceis formed between the 3C domain of one 3CD molecule andthe 3D domain of a twofold symmetry-related 3CD molecule(interface D-C) (Fig. 4a and b). This interface, involving theback of the 3D palm, appears to be stabilized by interactionsinvolving several 3CD residues (Table 2 and Fig. 4b), withLys438, Lys156, and Thr154 playing key roles. A second inter-face, D-D, is formed between the twofold symmetry-related 3Ddomains, mostly involving the back of the fingers region (Table2 and Fig. 4c), including interactions involving Arg190 andAsn334. Interface D-D is further stabilized by the binding of acation, possibly Zn2�. There is a strong peak in the electrondensity, with four tetrahedrally arranged ligands surroundingthis peak (Fig. 1b). This potential Zn2� atom coordinatesHis453 (3Dpol residue His270), His455 (3Dpol residue His272),and Cys464 (3Dpol residue Cys281) from one 3CD moleculeand Asp446 (3Dpol residue Asp263) from the other 3CD mol-ecule. Interestingly, the residues that appear to be involved inthe stabilization of these interfaces are conserved among pi-cornaviruses, and a role for Zn2� in the modulation of coop-erative polymerase activity has been noted (21).
A possible biological role for these interfaces (discussedbelow) is consistent with their size, which is large relative tothat of most ordinary crystal-packing contacts. Each of the D-C
and D-D contacts buries about 500 Å2 of solvent-accessiblesurface, for a total of about 1,000 Å2. However, their shapecomplementarity statistics are only 0.51 and 0.43, respectively(24). These low values reflect the possibility that most of thebinding across these interfaces is mediated by salt bridges andpolar interactions between flexible residues (Fig. 4b and c) andis consistent with the contacts being formed transiently.
Structural overview of 3Dpol G64S. A mutation of 3Dpol
Gly64 to a serine (G64S) in the fingers domain results in apolymerase with increased fidelity, allowing the polymeraseto discriminate against ribavirin, a nucleoside analog (9, 39).Though the G64S mutant supports a lower overall rate ofspontaneous mutation in the virus, this comes at the expense ofmaking 3Dpol threefold less efficient in nucleotide incorpora-tion (4). In the structure of 3Dpol, Gly1 forms two hydrogenbonds with the main-chain N and O of Gly64 (48). At the sametime, the alpha-amino group of Gly1 hydrogen bonds toAla239 and Leu241, helping to position these residues forproper nucleotide selection. The overall crystal structure of theG64S mutant is similar to that of the wild-type 3Dpol, with anrmsd of only 0.28 Å along the main chain. The arrangement ofresidues involved in the Gly1 hydrogen-bonding network, par-ticularly in the vicinity of the G64S mutation, is also similar,except that the Ser64 side chain is able to form an additionalhydrogen bond to the side chain of the carboxylate group ofGlu2 and thereby promotes the simultaneous hydrogen bond-ing of Glu2 to Gln4 (Fig. 2). Because no large positional shiftsare evident in the apoenzyme, it is possible that the mutation
FIG. 4. Key interfaces formed by 3CD-3CD interactions. (a) The three labeled interfaces are formed between symmetry-related molecules of3CD. Interfaces D-C and D-C� are each formed between the 3C domain of one molecule and the 3D domain of another. Interface D-D is formedbetween two 3D domains that are related by twofold crystallographic symmetry. Green, yellow, and blue ribbons come from three different 3CDmolecules. The thumb, palm, and fingers region of the green molecule are labeled. The N terminus of the yellow 3C domain is indicated. (b)Interface D-C is shown in stereo. Key residues and hydrogen bonds are indicated. (c) A portion of interface D-D is shown in stereo, with somekey residues and hydrogen bonds indicated. Two symmetry-related copies of the putative Zn2� coordination site are present, with Zn2� ionsindicated as gray spheres. Pro88 from interface D-C is included in both panel b and panel c as an aid to orientation.
TABLE 2. Residues forming interfaces between symmetry-related3CD molecules
Molecule (residue no.), chain location (side or main) Notes anddistance (Å)C D D�a
His89, sidec Lys438 (255), mainb 2.8Pro88, maind Lys438 (255), sideb 2.4Gly155b Lys438 (255), sideb 3.5Lys156c Glu259 (76), sidec 3.3Lys156c Asp262 (79), sidec PotentialThr154c Asp502 (319), sideb 2.7Thr154c His263 (80), sideb Potential
Gln277 (94)c Arg190 (7)d 3.1Asp443 (260)c Arg190 (7)d 3.5His453 (270)c Asp446 (263)c Putative Zn2�
His455 (272)c Asp446 (263)c Putative Zn2�
Cys464 (281)c Asp446 (263)c Putative Zn2�
Asn334 (151)c Asn334 (151)c 3.2
a Note that molecules D and D� are related by a crystallographic twofold axisso that each of the listed interactions occurs twice in the D-D interface.
b Conserved among polioviruses, enteroviruses, coxsackieviruses, echoviruses,and certain rhinoviruses (A. Palmenberg, personal communication).
c Conserved among polioviruses.d Not conserved.
VOL. 81, 2007 CRYSTAL STRUCTURE OF POLIOVIRUS 3CD PROTEIN 3589
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Nov
embe
r 20
21 b
y 18
7.87
.15.
130.
affects fidelity and efficiency primarily by helping to lock the3Dpol N terminus and its surroundings into a more stablearrangement.
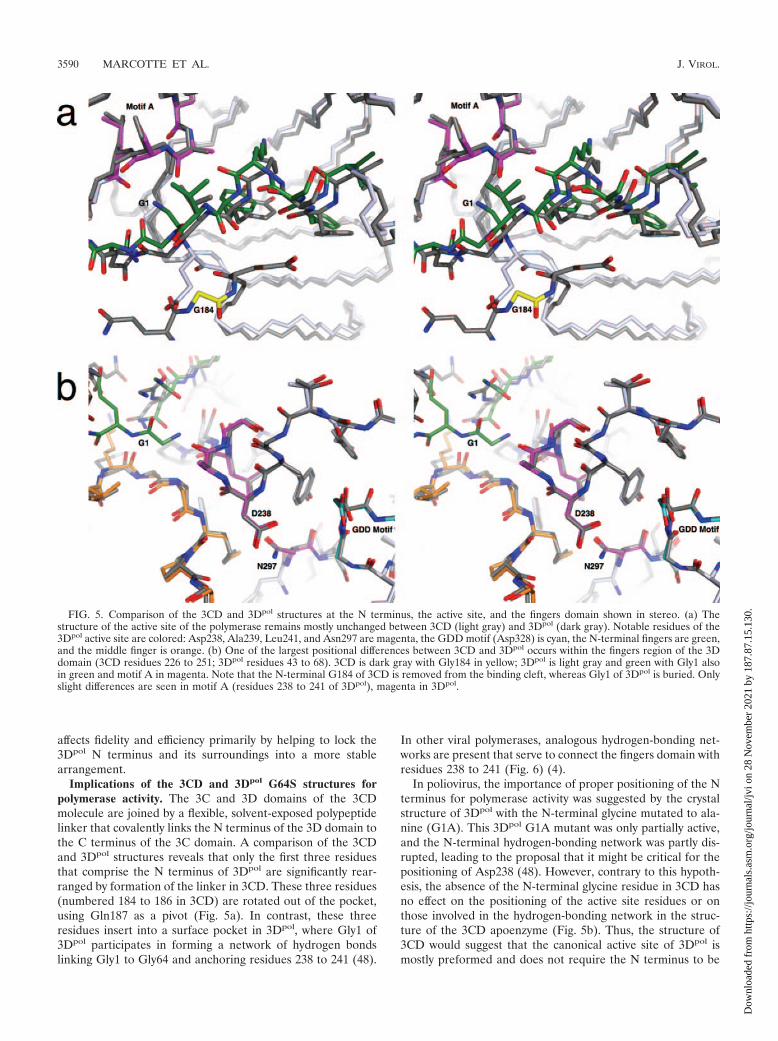
Implications of the 3CD and 3Dpol G64S structures forpolymerase activity. The 3C and 3D domains of the 3CDmolecule are joined by a flexible, solvent-exposed polypeptidelinker that covalently links the N terminus of the 3D domain tothe C terminus of the 3C domain. A comparison of the 3CDand 3Dpol structures reveals that only the first three residuesthat comprise the N terminus of 3Dpol are significantly rear-ranged by formation of the linker in 3CD. These three residues(numbered 184 to 186 in 3CD) are rotated out of the pocket,using Gln187 as a pivot (Fig. 5a). In contrast, these threeresidues insert into a surface pocket in 3Dpol, where Gly1 of3Dpol participates in forming a network of hydrogen bondslinking Gly1 to Gly64 and anchoring residues 238 to 241 (48).
In other viral polymerases, analogous hydrogen-bonding net-works are present that serve to connect the fingers domain withresidues 238 to 241 (Fig. 6) (4).
In poliovirus, the importance of proper positioning of the Nterminus for polymerase activity was suggested by the crystalstructure of 3Dpol with the N-terminal glycine mutated to ala-nine (G1A). This 3Dpol G1A mutant was only partially active,and the N-terminal hydrogen-bonding network was partly dis-rupted, leading to the proposal that it might be critical for thepositioning of Asp238 (48). However, contrary to this hypoth-esis, the absence of the N-terminal glycine residue in 3CD hasno effect on the positioning of the active site residues or onthose involved in the hydrogen-bonding network in the struc-ture of the 3CD apoenzyme (Fig. 5b). Thus, the structure of3CD would suggest that the canonical active site of 3Dpol ismostly preformed and does not require the N terminus to be
FIG. 5. Comparison of the 3CD and 3Dpol structures at the N terminus, the active site, and the fingers domain shown in stereo. (a) Thestructure of the active site of the polymerase remains mostly unchanged between 3CD (light gray) and 3Dpol (dark gray). Notable residues of the3Dpol active site are colored: Asp238, Ala239, Leu241, and Asn297 are magenta, the GDD motif (Asp328) is cyan, the N-terminal fingers are green,and the middle finger is orange. (b) One of the largest positional differences between 3CD and 3Dpol occurs within the fingers region of the 3Ddomain (3CD residues 226 to 251; 3Dpol residues 43 to 68). 3CD is dark gray with Gly184 in yellow; 3Dpol is light gray and green with Gly1 alsoin green and motif A in magenta. Note that the N-terminal G184 of 3CD is removed from the binding cleft, whereas Gly1 of 3Dpol is buried. Onlyslight differences are seen in motif A (residues 238 to 241 of 3Dpol), magenta in 3Dpol.
3590 MARCOTTE ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Nov
embe
r 20
21 b
y 18
7.87
.15.
130.

buried in order to achieve the correct conformation for initialNTP binding. Although it has been speculated that the post-cleavage insertion of the 3Dpol N terminus into its binding cleftmight shift active site residues from nonproductive to produc-tive positions, we cannot rule out the possibility that such ashift occurs in the elongation complex when 3Dpol binds itssubstrates. However, in our current snapshot of the apoenzymestructure, no such structural difference is evident. Indeed, theobservation that 3CD of FMDV shows some polymerase ac-tivity suggests that the specific contacts made by Gly1 areunlikely to be essential for catalysis (31). In addition, a gluta-thione S-transferase tag can be added to the N terminus ofhuman rhinovirus 2 polymerase without significantly affectingits activity (14, 15).
Because the structure of the active site seems to retain afunctional conformation, we looked for structural variability inother regions of the polymerase to see what differences mightbe associated with the lack of polymerase activity in 3CD.Aside from the polypeptide linker connecting the 3C and 3Ddomains of 3CD, the largest differences in the 3D domainoccurred in three areas of the molecule that are remote fromthe active site. One difference involved a main-chain shift at
the D-D interface, possibly to accommodate the salt linkage ofArg7 and Asp260 between the two domains. A second main-chain shift occurred in the pinky region of the polymerase,which was reported to have a relatively poor electron density inthe 3Dpol crystal structure (48). A structural alignment of the3Dpol active site with the active site of bacteriophage T7 RNApolymerase indicated that this region of the pinky domain(residues 124 to 149) is analogous to the T7 specificity loop(48). The T7 specificity loop, which includes several conservedarginines and lysines in both T7 and 3Dpol, specifically recog-nizes the promoter and also interacts with the nascent tran-script, possibly stabilizing the elongation complex (47). Afterclustered charged-to-alanine mutation of the correspondingresidues in poliovirus, transfection into HeLa cells failed toyield viable virus (10).
A possibly more relevant shift occurs in a stretch of residuesin the fingers domain of 3D (3CD residues 226 to 251; 3Dpol
residues 43 to 68). This area displays significant variabilitybetween the 3CD and the 3Dpol structures, with the main-chainatoms of 3CD shifting up to �1.7 Å compared to those of thestructure of 3Dpol (Fig. 5a). In 3Dpol, the functions of severalresidues in this region are well characterized, and their muta-
FIG. 6. Hydrogen-bonded linkage between amino-terminal residues (blue), active site residues (red and yellow), and the fingers domain (gray)in several viral RNA-dependent RNA polymerases (RdRps). The RdRps of poliovirus, FMDV and HRV14, form hydrogen-bonding networks,connecting the fingers domain with the N terminus and active site residues. These interactions are absent in PV 3CD. Structural motif A is red;structural motif C is yellow (18).
VOL. 81, 2007 CRYSTAL STRUCTURE OF POLIOVIRUS 3CD PROTEIN 3591
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Nov
embe
r 20
21 b
y 18
7.87
.15.
130.

tions have detrimental effects on polymerase activity. The3Dpol residue Lys61 appears to be critical, as its mutationeliminates the binding of GTP and polymerase activity (41).Simultaneous charged-to-alanine mutation of 3Dpol residueLys51-Asp53 or Asp53-Glu55-Glu56 prevents the accumula-tion of viral RNA (40). Removal of the first 68 amino acidresidues of 3Dpol also inactivates the polymerase (48). Con-ceivably, residues 43 to 68 of 3Dpol may serve as an allostericswitch for polymerase activity that is controlled by the presenceor absence of the 3Dpol N terminus.
In a number of viral polymerases, the N terminus and theactive site are stabilized via hydrogen bonding networks involv-ing the fingers domain, a conservation suggesting that thisbinding scheme could be relevant for polymerase activity (Fig.6) (3, 4, 13). It may also be possible that insertion of the 3D Nterminus in its binding pocket serves to modulate activity byregulating molecular flexibility. Indeed, there is precedent byother polymerases for fidelity mutations to occur distantly fromthe active site, which operate by making their polymerases lessflexible (23). We can certainly envision an activity spectrumwherein the molecule that is too inflexible (e.g., G64S) is lessefficient than that in the wild type; the molecule that is moreflexible (e.g., G1A, due to an imperfect fit) is poorer than thatin the wild type; and the molecule that is much too flexible (i.e.,3CD or 3D missing its N-terminal residues) is incapable ofsupporting viral replication.
Implications for the uridylylation of VPg. Both 3CD and3Dpol contribute to the initiation of poliovirus RNA replica-tion. This requires the uridylylation of VPg, wherein two copiesof UMP are linked in succession to the hydroxyl group of Tyr3via phosphodiester bonds (38). Uridylylation is catalyzed by3Dpol using an RNA template, either from the 3�-poly(A) tailor from cre(2C). Although uridylylation requires only 3D, VPg,and Mg2�, the uridylylation templated by cre(2C) is stimulated20-fold by the addition of 3CD or 10-fold by the addition of3Cpro (36, 37) (Fig. 7). Furthermore, the uncleaved precursor3BC is a ninefold better substrate for uridylylation than 3Balone. The uncleaved precursor 3BCD is also a substrate foruridylylation, albeit a poorer one than 3B or 3BC. Collectively,these observations suggest that domains 3B, 3C, and 3D eachhave distinct, nonoverlapping binding sites in the biologicallyrelevant uridylylation complex. They also imply that the bind-ing sites must be separated by distances short enough for theavailable linkers to connect them all together.
Three models for uridylylation have recently been published.Two of the models, based on mutation and computation studies,assume that VPg and the VPg precursor 3AB bind to the samesurface on 3Dpol and conclude that VPg enters the polymeraseactive site from the back of the polymerase molecule (45, 46). Incontrast, structural evidence from the FMDV 3Dpol-VPg complexsuggests an alternative arrangement, since the last ordered resi-dues at the carboxyl end of VPg are seen to bind to the front faceof 3Dpol. The crystal structure of 3CD supports the latter model.The 3CD structure appears to be relevant to the uridylylationreaction, as it suggests how the components of the 3CD-stimu-lated uridylylation reaction may be arranged in the complex.
VPg binding to FMDV 3Dpol is likely to be highly analogousto the binding in poliovirus 3Dpol, particularly because theFMDV enzyme can use poliovirus VPg as its substrate (31).Additionally, mutations made to PV 3Dpol residues Tyr326,
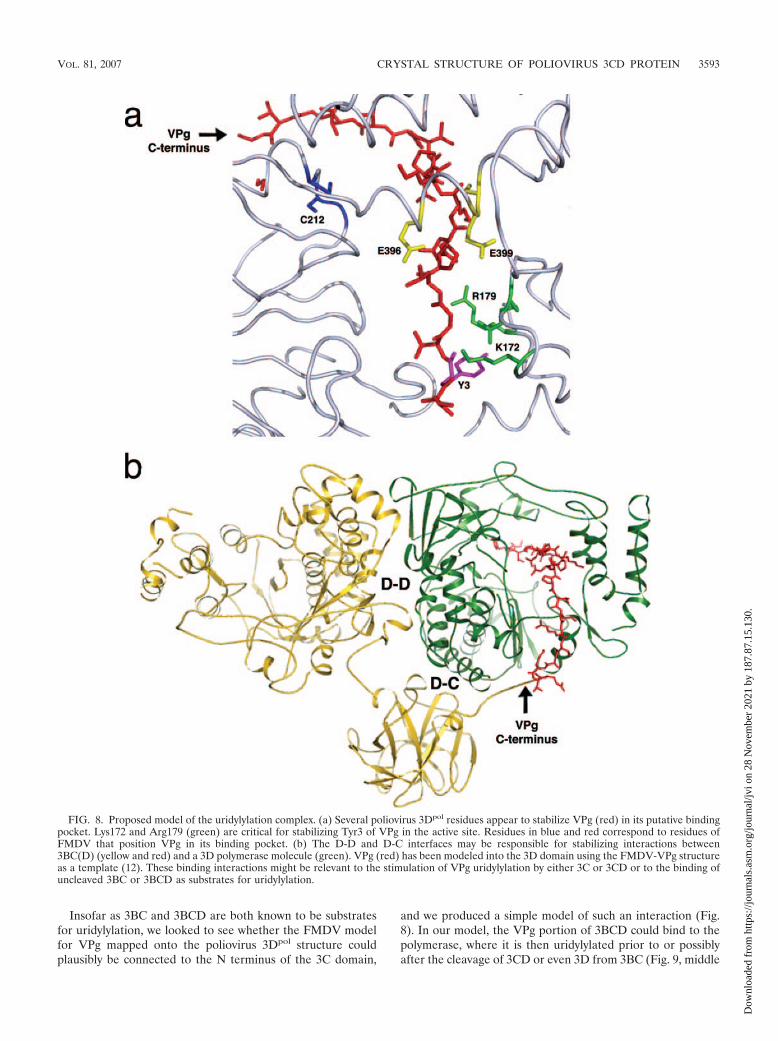
Asp358, and Lys359, which correspond to FMDV residuesactively involved in VPg binding, result in an almost completeloss of uridylylation activity (26, 27). In FMDV, the VPgprimer appears to enter from the front of the molecule andoccupies regions of the polymerase that bind to the templateand primer RNA (13). The N terminus of VPg binds a portionof the primer’s binding site; the C-terminal end of VPg looks asif it would follow along the path of the exiting RNA duplexproduct, though only the first 15 residues of VPg are actuallyvisible (12). Conserved residues in the fingers and thumb do-mains of the polymerase were identified as being responsiblefor stabilizing VPg in its binding cavity (Fig. 8a).
In the 3CD crystal structure, each 3D domain forms exten-sive interfaces with the 3C and 3D domains of a symmetry-related 3CD molecule (Fig. 4a). Several residues, some con-served, appear to be stabilizing these interfaces (Table 2 andFig. 4b and c). Notably, the N terminus of one 3C domain islocated close to the VPg binding site of an adjacent 3D domainof another 3CD molecule, as VPg was seen in the FMDVpolymerase complex (12). Assuming that these interfaces arerelevant to the 3CD-stimulated uridylylation reaction, it is onlypossible to connect the VPg binding site to the N terminus of3C by linking across the front face of 3Dpol (Fig. 8b). As in theFMDV structure, VPg would exit the polymerase on the frontof 3Dpol, and there would be no way for a connection on theback face to reach any 3C N terminus.
FIG. 7. 3BCD and its cleavage products: substrates and stimulatoryfactors for uridylylation. Large ovals represent the viral protein do-mains; wavy lines represent polypeptide chains. The cre RNA is pre-dicted to have a stem-loop structure and is shown in gray. The purpleand yellow rectangles represent the D-D and D-C interfaces, respec-tively, as they are seen in the 3CD crystal structure (shown in Fig. 5).The binding of additional copies of the 3C and/or 3D domains and theinvolvement of additional interfaces cannot be ruled out.
3592 MARCOTTE ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Nov
embe
r 20
21 b
y 18
7.87
.15.
130.
Insofar as 3BC and 3BCD are both known to be substratesfor uridylylation, we looked to see whether the FMDV modelfor VPg mapped onto the poliovirus 3Dpol structure couldplausibly be connected to the N terminus of the 3C domain,
and we produced a simple model of such an interaction (Fig.8). In our model, the VPg portion of 3BCD could bind to thepolymerase, where it is then uridylylated prior to or possiblyafter the cleavage of 3CD or even 3D from 3BC (Fig. 9, middle
FIG. 8. Proposed model of the uridylylation complex. (a) Several poliovirus 3Dpol residues appear to stabilize VPg (red) in its putative bindingpocket. Lys172 and Arg179 (green) are critical for stabilizing Tyr3 of VPg in the active site. Residues in blue and red correspond to residues ofFMDV that position VPg in its binding pocket. (b) The D-D and D-C interfaces may be responsible for stabilizing interactions between3BC(D) (yellow and red) and a 3D polymerase molecule (green). VPg (red) has been modeled into the 3D domain using the FMDV-VPg structureas a template (12). These binding interactions might be relevant to the stimulation of VPg uridylylation by either 3C or 3CD or to the binding ofuncleaved 3BC or 3BCD as substrates for uridylylation.
VOL. 81, 2007 CRYSTAL STRUCTURE OF POLIOVIRUS 3CD PROTEIN 3593
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Nov
embe
r 20
21 b
y 18
7.87
.15.
130.

panel). The 3BCD molecule has been shown to be inefficientlyuridylylated, though 3BC is a much better substrate for uri-dylylation than VPg alone. These biochemical results may suggestthat the 3D region is preferentially cleaved prior to uridylyla-tion to accommodate other components of the uridylylationreaction. Additionally, the observation that 3B yields a fasterturnover of uridylylated product than 3BCD can possibly beexplained by the difficulty that uridylylated 3BCD may have indissociating from the uridylylation complex. Although we havenoticed that an alternative pathway could connect VPg to theN terminus of a different 3CD molecule via the D-C� interface(Fig. 4a), we do not believe that this 3CD molecule is relevantto the uridylylation reaction: its 3C domain makes less exten-sive contacts than the D-C interface, and its 3D domain makesno contacts at all.
Importantly, the 3BCD model showing how 3BCD mightinteract with 3Dpol makes testable predictions about the natureof the enzyme-substrate complexes (3D-3B, 3D-3BC, and 3D-3BCD) and about the complexes wherein uridylylation is stim-ulated by 3C or 3CD binding (3D-3B-3C and 3D-3B-3CD).Therefore, we have used the 3CD structure and the 3BCDmodel to identify residues involved in binding at the interfaceregions (Table 2 and Fig. 4), and mutations have been made to
several of these residues. Uridylylation reactions using these3CD and 3Dpol mutant proteins were used to assess how wellthe uridylylation reaction could be stimulated in vitro (Table3). In general, interface mutations made to 3Dpol decreased3CD-stimulated uridylylation of VPg, whereas mutations madeto 3CD led to an increase in stimulation. The finding thaturidylylation is consistently affected lends support to our hy-pothesis that the 3D-3CD contacts identified in the crystalstructure are biologically relevant ones. When 3Dpol is mu-tated, stimulation is reduced, as would be expected if the mu-tations reduced the extent of the contacts with 3CD. On theother hand, attempts to mutate the interfaces from the 3CDside were less predictable, since equilibria involving the inter-actions of 3CD molecules with one another would also beaffected, perhaps even more strongly (Fig. 9, top panel). Themost dramatic effect of a tested mutation was the completeinability of the 3CD K156A mutant at interface D-C to stim-ulate uridylylation (Table 3). A double mutant that includesK156A showed the same behavior. Previously, it was alsofound that a 3CD K156E mutant was incapable of forminganother regulatory ribonucleoprotein complex that includedthe 5� cloverleaf RNA of poliovirus and that mutations madeto other residues surrounding K156 in the 3CD structure abol-ished RNA replication in vivo (2).
Roles of other protein-protein interfaces in the uridylylationcomplex. The 3CD protein is poorly behaved at high concen-trations. It was difficult to crystallize 3CD due partly to multi-ple interactions between its component domains. More thanone set of these interactions is likely to be biologically signif-icant, affecting 3CD’s roles in uridylylation, RNA elongation,and the regulation of transcription and RNA polymerization.Additionally, interactions may vary, depending on the contextof higher-order complexes. To further understand what thevarious multicomponent complexes may look like, it may befruitful to ask which of the interfaces seen in this and otherstudies can physically coexist with one another without pro-ducing steric clashes.
Previously, much attention has been focused on the head-to-tail stacking of 3Dpol molecules (interface I) that was dis-covered by Hansen et al. (18) in the 3Dpol crystal structure and
FIG. 9. How the structure of the uridylylation complex might affectbiological function. Large ovals represent the viral protein domains;wavy lines represent polypeptide chains. The cre RNA is predicted tohave a stem-loop structure and is shown in gray. The purple and yellowrectangles represent the D-D and D-C interfaces, respectively, as theyare seen in the 3CD crystal structure. Aggregation of 3CD may com-pete with the formation of the uridylylation complex (top panel). 3CDmight dissociate either intact or after it has been cleaved (middlepanel). Interfaces D-D and D-C can coexist with interface I (redrectangle, bottom panel, left); however, the formation of interfaceD-C� (white rectangle) would be in steric conflict with interface I(bottom panel, right). Interface II (not shown) would conflict withinterface D-C.
TABLE 3. Effect of the 3CD-3D interface on the ratio between3CD-stimulated cre-dependent uridylylation of VPg
and unstimulated uridylylationa
3CD WT ormutants 3D WT
3Dpol mutant proteins (residue no.)d
R7A(190)
D260A(443)
D263A(446)
D260AD263A
D319A(502)c
3CD WT 36.3 9.8 5.9 6.9 6.1 5.0T154Ac 47.3 16.6 11.9 9.7 7.1 7.4K156Ab,c 2.2 0.8 1.1 1.5 0.5 0.5K156A T154Ab,c 1.4 2.7 3.5 2.5 2.1 2.4R190Ad 148.1 78.6 29.8 37.4 24.7 11.2D446Ad 153.8 77.6 41.1 46.4 26.1 18.4D443A D446Ad 138.0 82.7 51.7 50.2 50.8 25.0
a Each value represents the ratio of 3CD-stimulated uridylylation to unstimu-lated uridylylation (see Materials and Methods). WT, wild type.
b Bands too weak to measure reliably were observed in the K156A mutant andin the 3CD K156A T154A double mutant.
c Residue located at interface D-C.d Residue located at interface D-D.
3594 MARCOTTE ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Nov
embe
r 20
21 b
y 18
7.87
.15.
130.

which is seen by electron microscopy (26) at high concentra-tions. Here, residues from the back of the thumb of one 3Dpol
molecule (residues Leu446, Pro448, Thr452, Arg455, andArg456) interact with residues from the back of the palm of asecond 3Dpol molecule (including residues Tyr313, Asp339,Ser341, Leu342, Gln345, and Asp349). Although this interac-tion is not conserved among picornaviruses (which argues thatits role may not be crucial), the obvious strength of the inter-action does suggest a possible biological role, as interface I haspreviously been proposed to be involved in elongation (21) andin uridylylation (5, 36). In this report, we have provided addi-tional pieces to the puzzle, showing that the mutations ofconserved residues in the D-D and D-C interfaces affect thestimulation of uridylylation and, in particular, that mutationson the 3Dpol side reduce this stimulation (Table 3).
To better understand what structure(s) of the uridylylationcomplex is possible and as a preliminary step to modeling thecomplex (A. B. Wass et al., manuscript in preparation), weassessed whether interfaces I, D-D, and D-C can coexist simul-taneously within a single molecular complex. Indeed, they can(Fig. 9, bottom panel, left). When a vertical column of head-to-tail 3Dpol domains is modeled, corresponding to the crystalstructure (PDB entry 1RDR) described by Hansen et al. (18),and a 3D domain (here representing 3Dpol) from the 3CDcrystal structure is superimposed by a least-squares fit, the 3Cand 3D domains of the twofold-related 3CD molecule wouldproject laterally from the side of the column. Thus, a central3Dpol molecule can be decorated simultaneously by two copiesof 3Dpol (each bound across interface I) and one copy of 3CD(bound to interfaces D-D and D-C) without any of these ad-ditional molecules clashing with one another. The closest ap-proach between the decorations is 9 Å, measured main chainto main chain, which would preclude steric hindrance, thoughone favorable side-chain interaction can be modeled.
A role should also be considered for the D-C� interface (Fig.4a). Here, a second copy of 3C is bound whose N terminus isclose enough to the neighboring 3Dpol active site for 3BC’s roleas a substrate to be modeled. However, we argue against thelikelihood of its participation in the uridylylation complex be-cause its 3D domain is not located close enough to participatein a 3BCD complex with 3Dpol. Furthermore, its Lys156 andThr154 residues are located too distant from 3Dpol to accountfor their dramatic effect on uridylylation (Table 3). The exis-tence of interface I may also be relevant because its formationwould inhibit 3C binding at the D-C� interface and vice versa,though the other copy of interface I (involving the thumb of3Dpol) would be unaffected (Fig. 9, bottom panel, right). Re-gardless of its biological relevance, the ability of 3C to interactwith residues from the back of the palm (3Dpol residues Tyr313and Lys314) might complicate the interpretation of previousmutation studies of interface I (5, 10). Nothing in the presentcrystal structure supports the prior hypothesis that the back ofthe thumb is involved in 3C binding at some point (36), but thisinteraction cannot be ruled out.
Finally, the formation of interface II between 3Dpol mole-cules, as described by Hansen et al. (18), is sterically incom-patible with the formation of interface D-C, though the keybinding interactions do not involve identical residues. Centralto interface II is Asp89 (3CD residue 272), which is involved indivalent cation binding, while Lys255 (3CD residue 438) lies
peripherally in the interface and does not bind directly. Incontrast, in the 3CD structure, Asp89 (residue 272) makes noobvious contacts, while Lys255 (residue 438) is central to form-ing the D-C interface (Fig. 4b). Indeed, it forms a hydrogenbond to the backbone oxygen of Gly155, and mutations in thatvicinity have a pronounced effect on uridylylation in vitro (Ta-ble 3). Mutation studies of charged residues in 3Dpol (10) haveconfirmed the likely importance of Lys255 (residue 438) forvirus viability (at least in combination with Glu254) and thelikely unimportance of Asp89 (residue 272) and interface II.The conservation of Lys255 (residue 438) among the poliovi-ruses, enteroviruses, echoviruses, coxsackieviruses, and certainrhinoviruses is probably relevant.
The poliovirus polymerase facilitates many critical functionsof the virus. However, its activity must be regulated. Regula-tion is provided in the form of 3CD, a precursor with nopolymerase activity. The crystal structure of 3CD reveals howthe two domains of the protein are tethered together, helps todefine the structural requirements for an active polymerase,and provides a model for the uridylylation of VPg. Uridylyla-tion and subsequent replication require the binding of a host ofmacromolecular components to form large replication com-plexes. As the structures of the individual components of thereplication complex become known, our incomplete picture ofcatalysis and regulation steadily improves. Our structure of3CD provides a more complete understanding of how 3CD and3Dpol function in viral replication and provides testable modelsfor future experiments.
ACKNOWLEDGMENTS
We thank Piotr Sliz for assistance with data collection and membersof the Hogle and Cameron laboratories for useful advice and criticalreading of the manuscript. We thank the staff members of ArgonneAdvance Photon Source and of the Brookhaven National SynchrotronLight Source for their assistance in data collection. We also thankTodd Parsley and Bert Semler for their previous work with 3CD andgenerous contribution of reagents and protocols over the years.
This work was supported by NIH grant AI20566 (to J.M.H.) andNIH/NIAID grant AI053531 (to C.E.C.).
REFERENCES
1. Ago, H., T. Adachi, A. Yoshida, M. Yamamoto, N. Habuka, K. Yatsunami,and M. Miyano. 1999. Crystal structure of the RNA-dependent RNA poly-merase of hepatitis C virus. Structure 7:1417–1426.
2. Andino, R., G. E. Rieckhof, P. L. Achacoso, and D. Baltimore. 1993. Polio-virus RNA synthesis utilizes an RNP complex formed around the 5�-end ofviral RNA. EMBO J. 12:3587–3598.
3. Appleby, T. C., H. Luecke, J. H. Shim, J. Z. Wu, I. W. Cheney, W. Zhong, L.Vogeley, Z. Hong, and N. Yao. 2005. Crystal structure of complete rhinovirusRNA polymerase suggests front loading of protein primer. J. Virol. 79:277–288.
4. Arnold, J. J., M. Vignuzzi, J. K. Stone, R. Andino, and C. E. Cameron. 2005.Remote site control of an active site fidelity checkpoint in a viral RNA-dependent RNA polymerase. J. Biol. Chem. 280:25706–25716.
5. Boerner, J. E., J. M. Lyle, S. Daijogo, B. L. Semler, S. C. Schultz, K.Kirkegaard, and O. C. Richards. 2005. Allosteric effects of ligands andmutations on poliovirus RNA-dependent RNA polymerase. J. Virol. 79:7803–7811.
6. Bressanelli, S., L. Tomei, F. A. Rey, and R. De Francesco. 2002. Structuralanalysis of the hepatitis C virus RNA polymerase in complex with ribonucle-otides. J. Virol. 76:3482–3492.
7. Burns, C. C., M. A. Lawson, B. L. Semler, and E. Ehrenfeld. 1989. Effects ofmutations in poliovirus 3Dpol on RNA polymerase activity and on polypro-tein cleavage. J. Virol. 63:4866–4874.
8. Choi, K. H., J. M. Groarke, D. C. Young, R. J. Kuhn, J. L. Smith, D. C.Pevear, and M. G. Rossmann. 2004. The structure of the RNA-dependentRNA polymerase from bovine viral diarrhea virus establishes the role ofGTP in de novo initiation. Proc. Natl. Acad. Sci. USA 101:4425–4430.
9. Crotty, S., M. C. Saleh, L. Gitlin, O. Beske, and R. Andino. 2004. Thepoliovirus replication machinery can escape inhibition by an antiviral drugthat targets a host cell protein. J. Virol. 78:3378–3386.
VOL. 81, 2007 CRYSTAL STRUCTURE OF POLIOVIRUS 3CD PROTEIN 3595
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Nov
embe
r 20
21 b
y 18
7.87
.15.
130.

10. Diamond, S. E., and K. Kirkegaard. 1994. Clustered charged-to-alaninemutagenesis of poliovirus RNA-dependent RNA polymerase yields multipletemperature-sensitive mutants defective in RNA synthesis. J. Virol. 68:863–876.
11. Emsley, P., and K. Cowtan. 2004. Coot: model-building tools for moleculargraphics. Acta Crystallogr. Sect. D 60:2126–2132.
12. Ferrer-Orta, C., A. Arias, R. Agudo, R. Perez-Luque, C. Escarmis, E. Domingo,and N. Verdaguer. 2006. The structure of a protein primer-polymerase complexin the initiation of genome replication. EMBO J. 25:880–888.
13. Ferrer-Orta, C., A. Arias, R. Perez-Luque, C. Escarmis, E. Domingo, and N.Verdaguer. 2004. Structure of foot-and-mouth disease virus RNA-dependentRNA polymerase and its complex with a template-primer RNA. J. Biol.Chem. 279:47212–47221.
14. Gerber, K., E. Wimmer, and A. V. Paul. 2001. Biochemical and geneticstudies of the initiation of human rhinovirus 2 RNA replication: identifica-tion of a cis-replicating element in the coding sequence of 2Apro. J. Virol.75:10979–10990.
15. Gerber, K., E. Wimmer, and A. V. Paul. 2001. Biochemical and geneticstudies of the initiation of human rhinovirus 2 RNA replication: purificationand enzymatic analysis of the RNA-dependent RNA polymerase 3Dpol.J. Virol. 75:10969–10978.
16. Gohara, D. W., S. Crotty, J. J. Arnold, J. D. Yoder, R. Andino, and C. E.Cameron. 2000. Poliovirus RNA-dependent RNA polymerase (3Dpol):structural, biochemical, and biological analysis of conserved structural motifsA and B. J. Biol. Chem. 275:25523–25532.
17. Gohara, D. W., C. S. Ha, S. Kumar, B. Ghosh, J. J. Arnold, T. J. Wisniewski,and C. E. Cameron. 1999. Production of “authentic” poliovirus RNA-de-pendent RNA polymerase (3D(pol)) by ubiquitin-protease-mediated cleav-age in Escherichia coli. Protein Expr. Purif. 17:128–138.
18. Hansen, J. L., A. M. Long, and S. C. Schultz. 1997. Structure of the RNA-dependent RNA polymerase of poliovirus. Structure 5:1109–1122.
19. Harris, D., N. Kaushik, P. K. Pandey, P. N. Yadav, and V. N. Pandey. 1998.Functional analysis of amino acid residues constituting the dNTP bindingpocket of HIV-1 reverse transcriptase. J. Biol. Chem. 273:33624–33634.
20. Harris, K. S., S. R. Reddigari, M. J. Nicklin, T. Hammerle, and E. Wimmer.1992. Purification and characterization of poliovirus polypeptide 3CD, aproteinase and a precursor for RNA polymerase. J. Virol. 66:7481–7489.
21. Hobson, S. D., E. S. Rosenblum, O. C. Richards, K. Richmond, K. Kirkegaard,and S. C. Schultz. 2001. Oligomeric structures of poliovirus polymerase areimportant for function. EMBO J. 20:1153–1163.
22. Kim, T. W., L. G. Brieba, T. Ellenberger, and E. T. Kool. 2006. Functionalevidence for a small and rigid active site in a high fidelity DNA polymerase:probing T7 DNA polymerase with variably sized base pairs. J. Biol. Chem.281:2289–2295.
23. Kool, E. T. 2002. Active site tightness and substrate fit in DNA replication.Annu. Rev. Biochem. 71:191–219.
24. Lawrence, M. C., and P. M. Colman. 1993. Shape complementarity at pro-tein/protein interfaces. J. Mol. Biol. 234:946–950.
25. Love, R. A., K. A. Maegley, X. Yu, R. A. Ferre, L. K. Lingardo, W. Diehl, H. E.Parge, P. S. Dragovich, and S. A. Fuhrman. 2004. The crystal structure of theRNA-dependent RNA polymerase from human rhinovirus: a dual functiontarget for common cold antiviral therapy. Structure 12:1533–1544.
26. Lyle, J. M., E. Bullitt, K. Bienz, and K. Kirkegaard. 2002. Visualization andfunctional analysis of RNA-dependent RNA polymerase lattices. Science296:2218–2222.
27. Lyle, J. M., A. Clewell, K. Richmond, O. C. Richards, D. A. Hope, S. C.Schultz, and K. Kirkegaard. 2002. Similar structural basis for membranelocalization and protein priming by an RNA-dependent RNA polymerase.J. Biol. Chem. 277:16324–16331.
28. McCoy, A. J., R. W. Grosse-Kunstleve, L. C. Storoni, and R. J. Read. 2005.Likelihood-enhanced fast translation functions. Acta Crystallogr. Sect. D61:458–464.
29. Mosimann, S. C., M. M. Cherney, S. Sia, S. Plotch, and M. N. James. 1997.Refined X-ray crystallographic structure of the poliovirus 3C gene product.J. Mol. Biol. 273:1032–1047.
30. Murshudov, G. N., A. A. Vagin, and E. J. Dodson. 1997. Refinement ofmacromolecular structures by the maximum-likelihood method. Acta Crys-tallogr. Sect. D 53:240–255.
31. Nayak, A., I. G. Goodfellow, and G. J. Belsham. 2005. Factors required forthe uridylylation of the foot-and-mouth disease virus 3B1, 3B2, and 3B3peptides by the RNA-dependent RNA polymerase (3Dpol) in vitro. J. Virol.79:7698–7706.
32. Ng, K. K., M. M. Cherney, A. L. Vazquez, A. Machin, J. M. Alonso, F. Parra,and M. N. James. 2002. Crystal structures of active and inactive conforma-tions of a caliciviral RNA-dependent RNA polymerase. J. Biol. Chem. 277:1381–1387.
33. Otwinowski, Z., and W. Minor. 1997. Processing of X-ray diffraction datacollected in oscillation mode, p. 307–326. In J. C. W. Carter and R. M. Sweet(ed.), Methods in enzymology, vol. 276. Academic Press, New York, NY.
34. Parsley, T. B., C. T. Cornell, and B. L. Semler. 1999. Modulation of the RNAbinding and protein processing activities of poliovirus polypeptide 3CD bythe viral RNA polymerase domain. J. Biol. Chem. 274:12867–12876.
35. Parsley, T. B., J. S. Towner, L. B. Blyn, E. Ehrenfeld, and B. L. Semler. 1997.Poly (rC) binding protein 2 forms a ternary complex with the 5�-terminalsequences of poliovirus RNA and the viral 3CD proteinase. RNA 3:1124–1134.
36. Pathak, H. B., S. K. Ghosh, A. W. Roberts, S. D. Sharma, J. D. Yoder, J. J.Arnold, D. W. Gohara, D. J. Barton, A. V. Paul, and C. E. Cameron. 2002.Structure-function relationships of the RNA-dependent RNA polymerasefrom poliovirus (3Dpol). A surface of the primary oligomerization domainfunctions in capsid precursor processing and VPg uridylylation. J. Biol.Chem. 277:31551–31562.
37. Paul, A. V., E. Rieder, D. W. Kim, J. H. van Boom, and E. Wimmer. 2000.Identification of an RNA hairpin in poliovirus RNA that serves as theprimary template in the in vitro uridylylation of VPg. J. Virol. 74:10359–10370.
38. Paul, A. V., J. H. van Boom, D. Filippov, and E. Wimmer. 1998. Protein-primed RNA synthesis by purified poliovirus RNA polymerase. Nature 393:280–284.
39. Pfeiffer, J. K., and K. Kirkegaard. 2003. A single mutation in poliovirusRNA-dependent RNA polymerase confers resistance to mutagenic nucleo-tide analogs via increased fidelity. Proc. Natl. Acad. Sci. USA 100:7289–7294.
40. Plotch, S. J., O. Palant, and Y. Gluzman. 1989. Purification and propertiesof poliovirus RNA polymerase expressed in Escherichia coli. J. Virol. 63:216–225.
41. Richards, O. C., S. Baker, and E. Ehrenfeld. 1996. Mutation of lysine resi-dues in the nucleotide binding segments of the poliovirus RNA-dependentRNA polymerase. J. Virol. 70:8564–8570.
42. Rieder, E., A. V. Paul, D. W. Kim, J. H. van Boom, and E. Wimmer. 2000.Genetic and biochemical studies of poliovirus cis-acting replication elementcre in relation to VPg uridylylation. J. Virol. 74:10371–10380.
43. Rothstein, M. A., O. C. Richards, C. Amin, and E. Ehrenfeld. 1988. Enzy-matic activity of poliovirus RNA polymerase synthesized in Escherichia colifrom viral cDNA. Virology 164:301–308.
44. Sarkany, Z., and L. Polgar. 2003. The unusual catalytic triad of poliovirusprotease 3C. Biochemistry 42:516–522.
45. Schein, C. H., N. Oezguen, D. E. Volk, R. Garimella, A. Paul, and W. Braun.2006. NMR structure of the viral peptide linked to the genome (VPg) ofpoliovirus. Peptides 27:1676–1684.
46. Tellez, A. B., S. Crowder, J. F. Spagnolo, A. A. Thompson, O. B. Peersen,D. L. Brutlag, and K. Kirkegaard. 2006. Nucleotide channel of RNA-de-pendent RNA polymerase used for intermolecular uridylylation of proteinprimer. J. Mol. Biol. 357:665–675.
47. Temiakov, D., P. E. Mentesana, K. Ma, A. Mustaev, S. Borukhov, and W. T.McAllister. 2000. The specificity loop of T7 RNA polymerase interacts firstwith the promoter and then with the elongating transcript, suggesting amechanism for promoter clearance. Proc. Natl. Acad. Sci. USA 97:14109–14114.
48. Thompson, A. A., and O. B. Peersen. 2004. Structural basis for proteolysis-dependent activation of the poliovirus RNA-dependent RNA polymerase.EMBO J. 23:3462–3471.
49. Xiang, W., K. S. Harris, L. Alexander, and E. Wimmer. 1995. Interactionbetween the 5�-terminal cloverleaf and 3AB/3CDpro of poliovirus is essentialfor RNA replication. J. Virol. 69:3658–3667.
3596 MARCOTTE ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 28
Nov
embe
r 20
21 b
y 18
7.87
.15.
130.





























