Embed Size (px)
Citation preview

Cross-Species Colour Segmenting: A Novel Toolin Human Karyotype Analysis
S. Muller, P.C.M. O’Brien, M.A. Ferguson-Smith, and J. Wienberg*Department of Pathology, University of Cambridge, Cambridge, United Kingdom
Received 13 March 1998; Revision Received 13 August 1998; Accepted 15 September 1998
We used fluorescence in situ hybridization (FISH)with DNA probes derived from bivariate fluores-cence activated flow sorting of primate chromo-somes. In cases where human and primate karyo-types differ by chromosome rearrangements, reversepainting of primate probes resulted in a subregionaldelineation of the human homologous chromo-somes. Probes were used from two gibbon species(Hylobates concolor and H. syndactylus) which bothshowed highly rearranged karyotypes. Hybridiza-tion of human chromosomes with painting probesderived from both gibbons showed that, with theexception of human chromosomes 15, 18, 21, 22 andthe sex chromosomes, each chromosome was differ-entiated in at least two and up to six segments. Theseprobes have been used in the analysis of variouscases of constitutional chromosomal rearrange-
ments in human pathology including complex intra-chromosomal rearrangements. They were also usedin a multi colour format (colour segmenting) to differen-tiate the entire human karyotype into 81 homologouscoloured segments with probes derived from H. con-color, and 74 segments with probes derived from H.syndactylus. The addition of colours not only simplifieschromosome identification compared to the analysis ofclassical banding based on grey values, but coloursegmenting also provides simple coloured land-marks for further fine analysis by classical banding.Cytometry 33:445–452, 1998. r 1998 Wiley-Liss, Inc.
Key terms: cross-species colour segmenting; karyo-type analysis; chromosome colour segmenting; fluo-rescence in situ hybridization; image analysis; paint-ing probes
Fluorescence in situ hybridization (19), in particularchromosome painting with human chromosome specificprobes derived by fluorescence activated bivariate flowsorting (FACS), has contributed significantly to the progressin human karyotype analysis (3,12,20). Chromosome paint-ing has also been performed in multi colour formats bylabeling probes with different haptens in a simple Booleanfashion (16,17,22) or by including ratios of differentiallylabelled probes (18). However, these new strategies onlyallow identification of whole chromosomes and thereforeare limited to the analysis of chromosome translocationswithout further differentiation of chromosomal subre-gions. Intrachromosomal rearrangements (inversions, trans-positions) will escape analysis which would need furtherdelineation of chromosome subregions.
Subregional probes painting several coloured chromo-some bands have been established by microdissection (6)and ‘‘reverse painting’’ (2). Alternatively single landmarkscan be set by using cloned DNA (1,7,11,15,22) or probesderived from rearranged human/rodent somatic cell hy-brids (14). However, these probes are not yet available inequivalent quality and quantity to delineate the entire humankaryotype in different colours as compared to painting probes,where each member of the entire chromosome set can
now be simultaneously distinguished from another usingdifferent combinations of fluorescent labels (23,24).
Here we report on a set of subregional paints for humanchromosome analysis which were derived by FACS fromprimates and paint the entire human karyotype. Althoughgibbons (genus Hylobates) are closely related to humansand are classified with great apes and human in the sameprimate superfamily, their karyotypes (H. hoolock, 2n538;H. lar, 2n544; H. syndactylus, 2n550 and H. concolor,2n552) show extensive chromosomal reorganization whencompared to human chromosomes (8–10). The range ofconserved segments varies from 52 (H. lar) to 67 (H.concolor), and some of the rearranged patterns are sharedbetween species, indicating a common origin. Probesderived from these primates are not only useful as singlesubregional paints in human chromosome analysis but canalso delineate the entire human karyotype in about 100
Contract grant sponsor: Deutsche Forschungsgemeinschaft; Contractgrant sponsor: Medical Research Council.
*Correspondence to: J. Wienberg, Department of Pathology, Universityof Cambridge, Tennis Court Road, Cambridge CB2 1QP, United Kingdom.
E-mail: [email protected]
r 1998 Wiley-Liss, Inc. Cytometry 33:445–452 (1998)
painted segments in a multicolour format we refer to ascross-species colour segmenting (CSC-segmenting). Fur-ther, because repetitive sequences have diverged substan-tially during the evolution of humans and lesser apes,levels of non-specific hybridization are considerably re-duced (13), which allows hybridization in the absence ofcompetitor DNA. The hybridization pattern is robust andhighly reproducible since boundaries are defined by evolu-tionary derived translocations.
MATERIALS AND METHODSDual laser flow sorting of gibbon chromosomes was as
described for other species (2,5,21). Chromosome specificprobes were generated by degenerate oligonucleotideprimed PCR (DOP-PCR) directly from the flow sortedchromosomes using PCR primers and amplification condi-tions as described (21,25). DNA derived from the primaryreaction and the same primers were used to label thechromosome paints with either biotin-dUTP (BoehringerMannheim, Germany), FITC-dUTP (Vector Laboratories,Burlingame, CA, USA), or Cy3-dUTP (Amersham, Cardiff,UK). Gibbon probes were labelled with the three haptensin various combinations as shown in Table 1 in 50 µlstandard DOP-PCR reactions (21). 4 µl (containing approxi-mately 100ng) of each individual gibbon probe werepooled, ethanol precipitated and resuspended in 15 µl ofhybridisation mixture (50% formamide, 2xSSC, 10% dex-tran sulfate). In situ hybridization of painting probes tohuman and gibbon chromosomes was performed as previ-ously described (2,21). No Cot-1 DNA for suppression wasnecessary, neither for the hybridisations of gibbon probeson gibbon chromosomes or gibbon on human chromo-somes. Chromosome paints labelled with biotin weredetected with avidin-Cy-5 (Amersham).
Digital images were obtained using a cooled CCDcamera (Photometrics, NU200 series equipped with Ko-dak KAF 1400 CCD) coupled to a Zeiss Axiophot micro-
scope. Camera control and digital image acquisition (8-bitgrey scale) employed an Apple Macintosh computer. Theimages were processed as described (22). Photographs fromthe images were taken directly from the computer. At least tenmetaphases were evaluated for each painted chromosome todefine the areas hybridized by any particular probe.
RESULTSUsing FACS and subsequent DOP-PCR we have gener-
ated painting probes from Hylobates concolor and H.syndactylus chromosomes. Previous work has identifiedthe homologies of chromosomes and chromosome seg-ments between human and both gibbons using humanchromosome specific painting probes (9, 10). To identifythe content of each peak in the flow karyotype hapten-labelled probes derived from the individual peaks werehybridised to the DAPI banded chromosomes of the
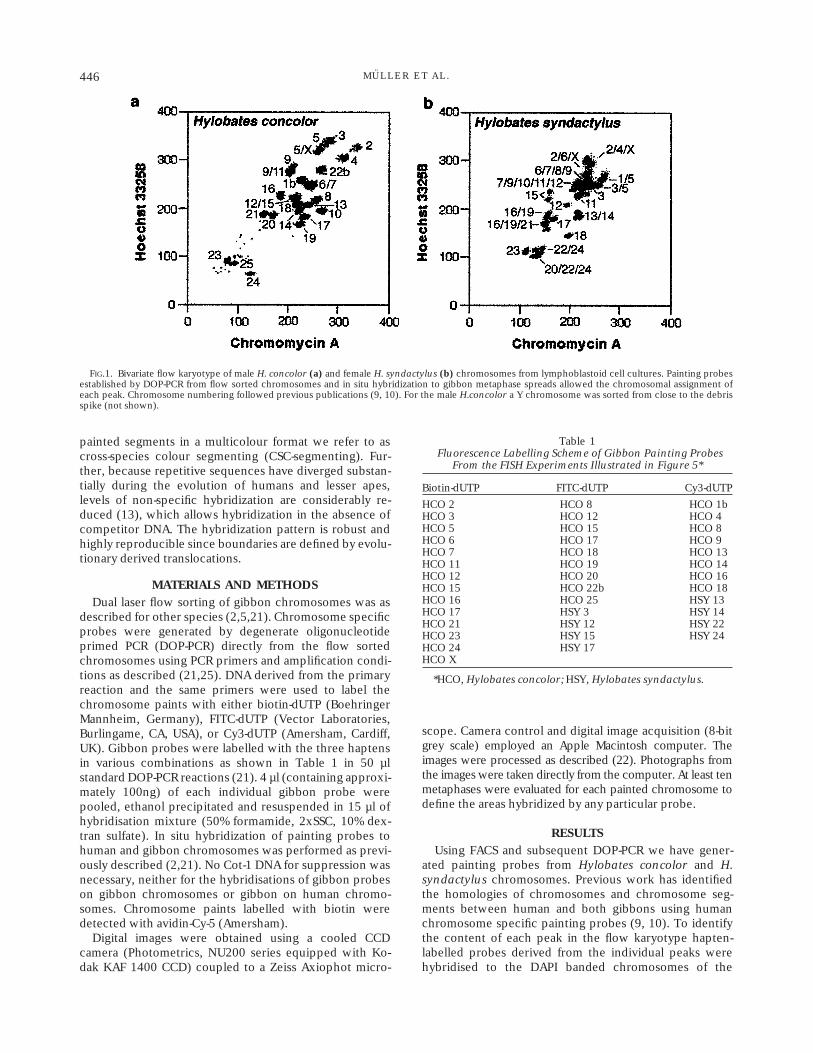
FIG.1. Bivariate flow karyotype of male H. concolor (a) and female H. syndactylus (b) chromosomes from lymphoblastoid cell cultures. Painting probesestablished by DOP-PCR from flow sorted chromosomes and in situ hybridization to gibbon metaphase spreads allowed the chromosomal assignment ofeach peak. Chromosome numbering followed previous publications (9, 10). For the male H.concolor a Y chromosome was sorted from close to the debrisspike (not shown).
Table 1Fluorescence Labelling Scheme of Gibbon Painting Probes
From the FISH Experiments Illustrated in Figure 5*
Biotin-dUTP FITC-dUTP Cy3-dUTP
HCO 2 HCO 8 HCO 1bHCO 3 HCO 12 HCO 4HCO 5 HCO 15 HCO 8HCO 6 HCO 17 HCO 9HCO 7 HCO 18 HCO 13HCO 11 HCO 19 HCO 14HCO 12 HCO 20 HCO 16HCO 15 HCO 22b HCO 18HCO 16 HCO 25 HSY 13HCO 17 HSY 3 HSY 14HCO 21 HSY 12 HSY 22HCO 23 HSY 15 HSY 24HCO 24 HSY 17HCO X
*HCO, Hylobates concolor; HSY, Hylobates syndactylus.
446 MULLER ET AL.
particular species. Figure 1 shows the flow karyotype ofthe two species. Many peaks contained single chromo-somes only, however, especially in H.syndactylus, a num-ber of chromosomes could not be resolved individuallyresulting in composite probes.
Hybridization of painting probes derived from bothgibbons to human chromosomes showed that, with theexception of human chromosomes 15, 18, 21, 22, and thesex chromosomes, each chromosome was differentiated inat least two and up to six subregions. Some humanchromosomes showed only poor subregional differentia-
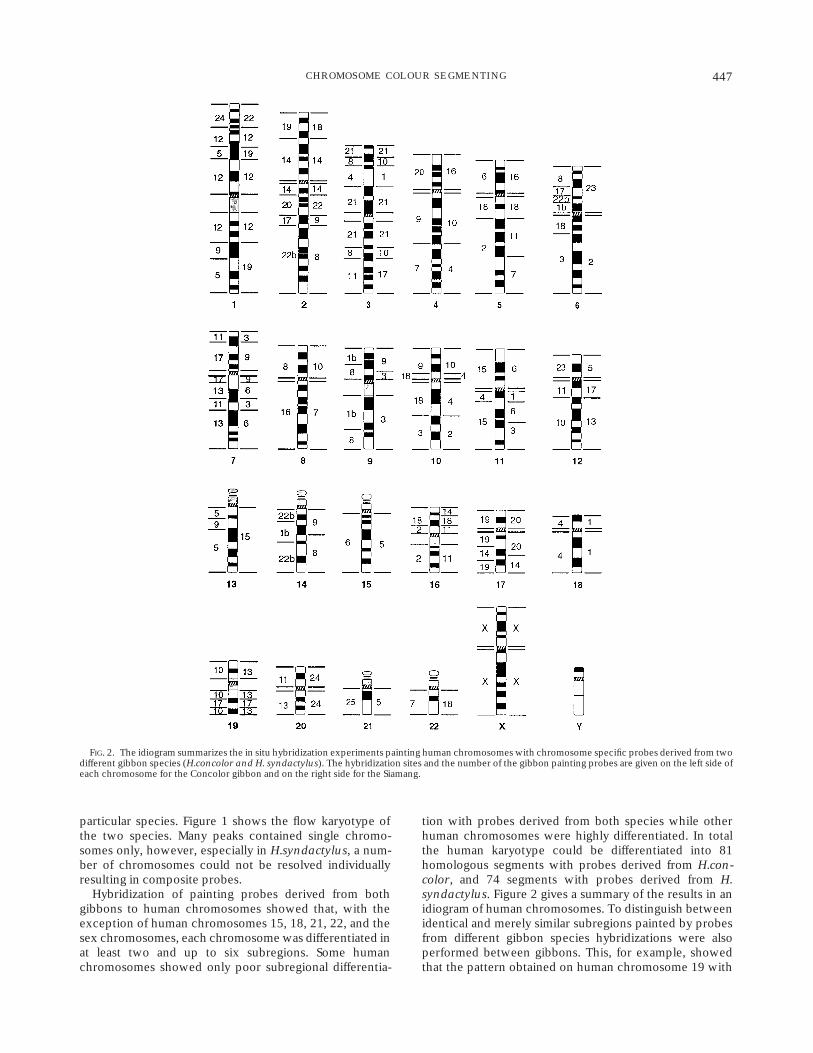
tion with probes derived from both species while otherhuman chromosomes were highly differentiated. In totalthe human karyotype could be differentiated into 81homologous segments with probes derived from H.con-color, and 74 segments with probes derived from H.syndactylus. Figure 2 gives a summary of the results in anidiogram of human chromosomes. To distinguish betweenidentical and merely similar subregions painted by probesfrom different gibbon species hybridizations were alsoperformed between gibbons. This, for example, showedthat the pattern obtained on human chromosome 19 with
FIG. 2. The idiogram summarizes the in situ hybridization experiments painting human chromosomes with chromosome specific probes derived from twodifferent gibbon species (H.concolor and H. syndactylus). The hybridization sites and the number of the gibbon painting probes are given on the left side ofeach chromosome for the Concolor gibbon and on the right side for the Siamang.
447CHROMOSOME COLOUR SEGMENTING
probes derived from both H.concolor and H.syndactyluswas identical (Fig. 2), whereas on human chromosome 17the hybridization between the two gibbons clearly indi-cated different rearrangements.
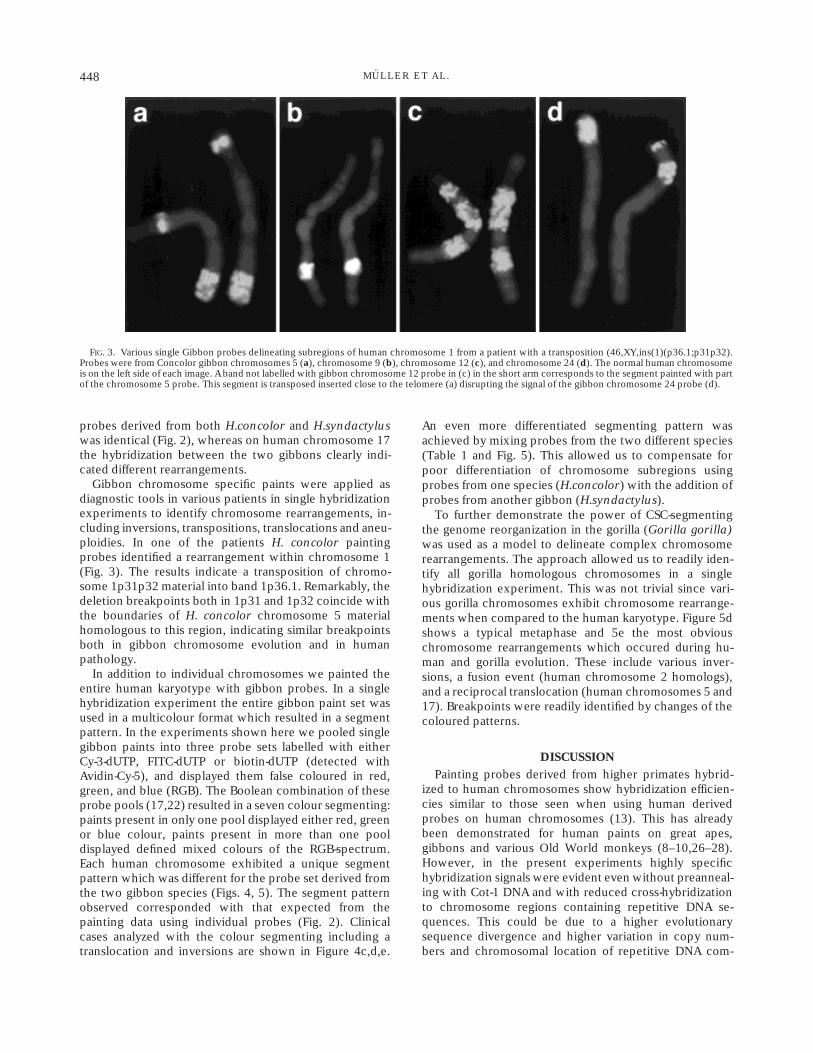
Gibbon chromosome specific paints were applied asdiagnostic tools in various patients in single hybridizationexperiments to identify chromosome rearrangements, in-cluding inversions, transpositions, translocations and aneu-ploidies. In one of the patients H. concolor paintingprobes identified a rearrangement within chromosome 1(Fig. 3). The results indicate a transposition of chromo-some 1p31p32 material into band 1p36.1. Remarkably, thedeletion breakpoints both in 1p31 and 1p32 coincide withthe boundaries of H. concolor chromosome 5 materialhomologous to this region, indicating similar breakpointsboth in gibbon chromosome evolution and in humanpathology.
In addition to individual chromosomes we painted theentire human karyotype with gibbon probes. In a singlehybridization experiment the entire gibbon paint set wasused in a multicolour format which resulted in a segmentpattern. In the experiments shown here we pooled singlegibbon paints into three probe sets labelled with eitherCy-3-dUTP, FITC-dUTP or biotin-dUTP (detected withAvidin-Cy-5), and displayed them false coloured in red,green, and blue (RGB). The Boolean combination of theseprobe pools (17,22) resulted in a seven colour segmenting:paints present in only one pool displayed either red, greenor blue colour, paints present in more than one pooldisplayed defined mixed colours of the RGB-spectrum.Each human chromosome exhibited a unique segmentpattern which was different for the probe set derived fromthe two gibbon species (Figs. 4, 5). The segment patternobserved corresponded with that expected from thepainting data using individual probes (Fig. 2). Clinicalcases analyzed with the colour segmenting including atranslocation and inversions are shown in Figure 4c,d,e.
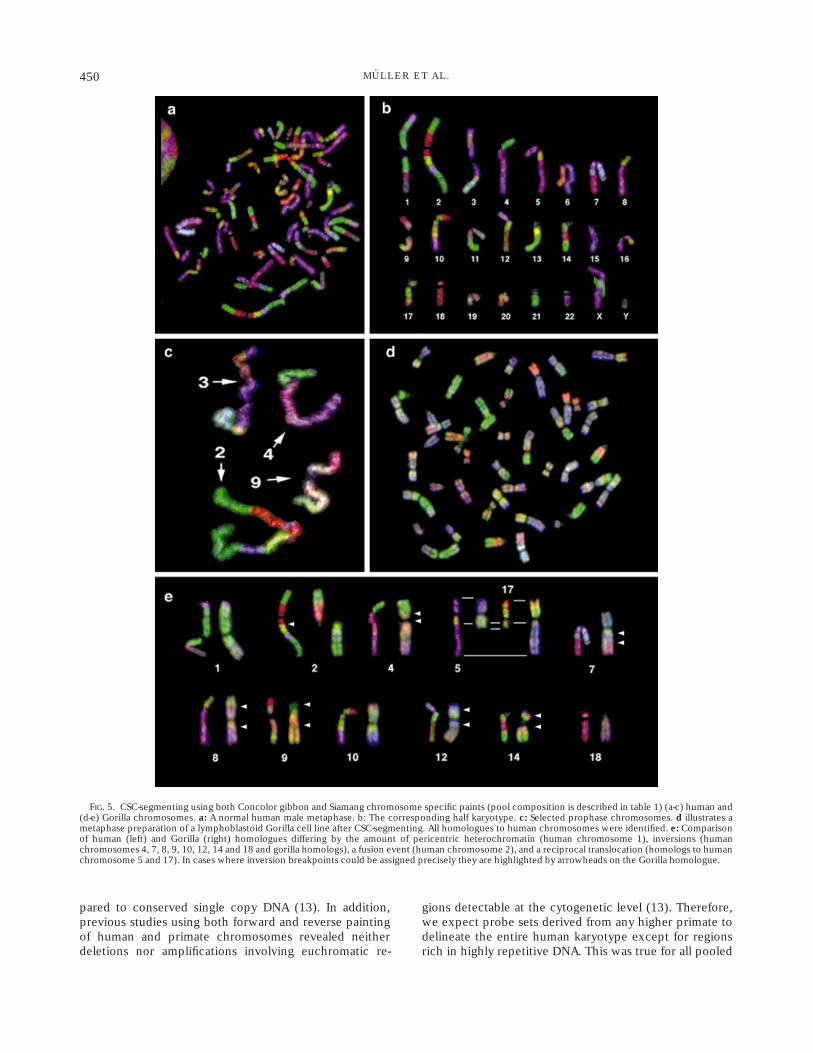
An even more differentiated segmenting pattern wasachieved by mixing probes from the two different species(Table 1 and Fig. 5). This allowed us to compensate forpoor differentiation of chromosome subregions usingprobes from one species (H.concolor) with the addition ofprobes from another gibbon (H.syndactylus).
To further demonstrate the power of CSC-segmentingthe genome reorganization in the gorilla (Gorilla gorilla)was used as a model to delineate complex chromosomerearrangements. The approach allowed us to readily iden-tify all gorilla homologous chromosomes in a singlehybridization experiment. This was not trivial since vari-ous gorilla chromosomes exhibit chromosome rearrange-ments when compared to the human karyotype. Figure 5dshows a typical metaphase and 5e the most obviouschromosome rearrangements which occured during hu-man and gorilla evolution. These include various inver-sions, a fusion event (human chromosome 2 homologs),and a reciprocal translocation (human chromosomes 5 and17). Breakpoints were readily identified by changes of thecoloured patterns.
DISCUSSION
Painting probes derived from higher primates hybrid-ized to human chromosomes show hybridization efficien-cies similar to those seen when using human derivedprobes on human chromosomes (13). This has alreadybeen demonstrated for human paints on great apes,gibbons and various Old World monkeys (8–10,26–28).However, in the present experiments highly specifichybridization signals were evident even without preanneal-ing with Cot-1 DNA and with reduced cross-hybridizationto chromosome regions containing repetitive DNA se-quences. This could be due to a higher evolutionarysequence divergence and higher variation in copy num-bers and chromosomal location of repetitive DNA com-
FIG. 3. Various single Gibbon probes delineating subregions of human chromosome 1 from a patient with a transposition (46,XY,ins(1)(p36.1;p31p32).Probes were from Concolor gibbon chromosomes 5 (a), chromosome 9 (b), chromosome 12 (c), and chromosome 24 (d). The normal human chromosomeis on the left side of each image. A band not labelled with gibbon chromosome 12 probe in (c) in the short arm corresponds to the segment painted with partof the chromosome 5 probe. This segment is transposed inserted close to the telomere (a) disrupting the signal of the gibbon chromosome 24 probe (d).
448 MULLER ET AL.
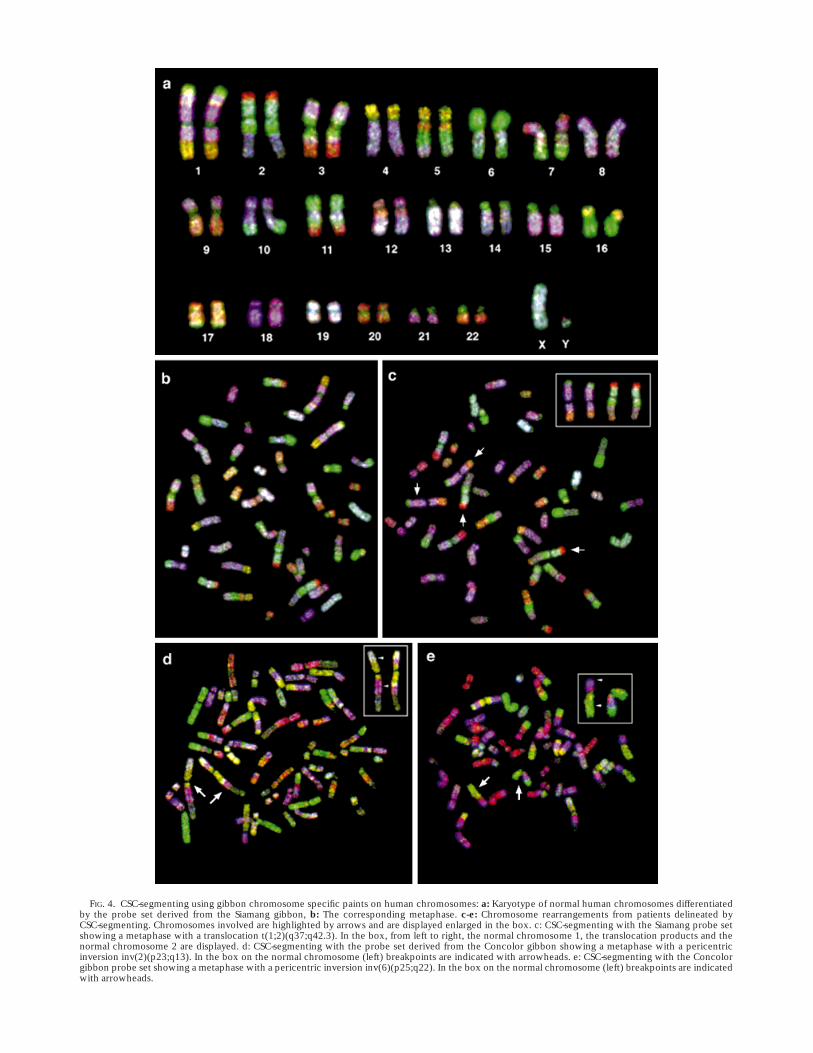
FIG. 4. CSC-segmenting using gibbon chromosome specific paints on human chromosomes: a: Karyotype of normal human chromosomes differentiatedby the probe set derived from the Siamang gibbon, b: The corresponding metaphase. c-e: Chromosome rearrangements from patients delineated byCSC-segmenting. Chromosomes involved are highlighted by arrows and are displayed enlarged in the box. c: CSC-segmenting with the Siamang probe setshowing a metaphase with a translocation t(1;2)(q37;q42.3). In the box, from left to right, the normal chromosome 1, the translocation products and thenormal chromosome 2 are displayed. d: CSC-segmenting with the probe set derived from the Concolor gibbon showing a metaphase with a pericentricinversion inv(2)(p23;q13). In the box on the normal chromosome (left) breakpoints are indicated with arrowheads. e: CSC-segmenting with the Concolorgibbon probe set showing a metaphase with a pericentric inversion inv(6)(p25;q22). In the box on the normal chromosome (left) breakpoints are indicatedwith arrowheads.
pared to conserved single copy DNA (13). In addition,previous studies using both forward and reverse paintingof human and primate chromosomes revealed neitherdeletions nor amplifications involving euchromatic re-
gions detectable at the cytogenetic level (13). Therefore,we expect probe sets derived from any higher primate todelineate the entire human karyotype except for regionsrich in highly repetitive DNA. This was true for all pooled
FIG. 5. CSC-segmenting using both Concolor gibbon and Siamang chromosome specific paints (pool composition is described in table 1) (a-c) human and(d-e) Gorilla chromosomes. a: A normal human male metaphase. b: The corresponding half karyotype. c: Selected prophase chromosomes. d illustrates ametaphase preparation of a lymphoblastoid Gorilla cell line after CSC-segmenting. All homologues to human chromosomes were identified. e: Comparisonof human (left) and Gorilla (right) homologues differing by the amount of pericentric heterochromatin (human chromosome 1), inversions (humanchromosomes 4, 7, 8, 9, 10, 12, 14 and 18 and gorilla homologs), a fusion event (human chromosome 2), and a reciprocal translocation (homologs to humanchromosome 5 and 17). In cases where inversion breakpoints could be assigned precisely they are highlighted by arrowheads on the Gorilla homologue.
450 MULLER ET AL.

gibbon probes which labelled the entire human genomeexcept for pericentromeric heterochromatin.
In case human and primate karyotypes differ by chromo-some rearrangements, reverse painting (2) should give asubregional delineation of the human homologous chromo-somes. Colour segmenting is based on the observation thatthe two gibbons used in the present experiments showextremely high numbers of chromosome rearrangements(8–10). In contrast to subregional probes generated bymicrodissection these probes have precise boundariessince they are defined by evolutionary derived transloca-tions.
In situ hybridisation of any differentially labelled DNAprobes will allow the design of colour segmenting orbanding appropriate to a particular application (7,11,14–18,22). This makes coloured patterns with molecularprobes entirely different from any classical chromosomebanding patterns which are, in general, either G-bandingor R-banding or variations thereof. However, our use ofcolour segmenting of the human genome is not to replaceclassical banding patterns but to complement and to assistin the identification of chromosome subregions which arefurther differentiated by DAPI G-banding.
The limits of colour segmenting to assist in the identifica-tion of chromosomal rearrangements have not yet beenfully explored. However, our data suggest that the additionof colours not only simplifies chromosome identificationcompared to the analysis of classical banding based ongray values, but segmenting also provides simple colouredlandmarks for further fine analysis by classical banding.This has been demonstrated in various cases of constitu-tional chromosomal rearrangements in human pathologyincluding a complex rearrangement (transposition) whereclassical banding analysis was ambiguous (Dr. L.Willatt,Addenbrooks Hospital, Cambridge, personal communica-tion).
Increasing resolution of segmenting can be achieved byprobes from lesser apes not yet incorporated into thepresent probe pools. Also various Old World monkeyderived probes, for example from African Green monkey(Cercopithecus aethiops) would contribute to the co-loured segmentation of the human karyotype. In contrastto gibbons, where translocations are predominant, Cerco-pithecus monkeys show numerous chromosome fissionswith karyotypes varying from 2n 5 48 to 72 (4). Thesespecies will certainly provide a set of chromosome armspecific probes for human karyotype analysis (Finelli et al.,manuscript in preparation.). The addition of other regionspecific probes to the probe pool (1,15) can also contrib-ute to a refined delineation of specific chromosomesegments. Therefore, we believe that the complexity ofthe segment pattern could be adjusted for any particularanalytical requirements.
The colour resolution of the segmenting pattern de-pends on the number of haptens involved. Using threehaptens simplifies the analysis of the segmenting patternsince the probes can be displayed in a simple red/green/blue (RGB) format accessible to most fluorescence micro-scopes and custom image processing computer software
(14,16,22). However, this arrangement does reduce thepossibility of identifying small telomeric translocations(Figure 4c) due to redundancy in the colour assignment.Recent publications successfully demonstrated that up tofive different labelling systems can be used in singlehybridization experiments to allow to display more than24 different chromosomes by ‘‘combinatorial labelling’’(23,24). In addition, various reports have demonstratedthat ‘‘ratio labelling’’ (17,18) can significantly increase thecolour information in FISH experiments. Similar strategiescan also be applied to colour segmenting. Each primatepaint would then produce segments of distinct colours onhuman chromosomes and would identify any chromo-some subregion on the molecular level in a single FISHexperiment.
CONCLUSIONSRecent multi-colour approaches using ‘‘spectral karyo-
typing’’ (SKY) and M-FISH allow to delineate the entirehuman karyotype in a single FISH experiment (23, 24). Incontrast to these techniques which identify inter-chromo-somal rearrangements, CSC-segmenting also allows todetect intra-chromosomal changes. This was demon-strated in our experiments both in clinical cytogeneticanalysis of translocations, inversions and a transposition aswell as in the comparative analysis of human and gorillametaphase chromosomes. In the gorilla CSC-segmentingidentified a translocation, chromosome fusion and variousperi- and paracentric inversions. Thus, when applied toclinical cytogenetics CSC-segmenting should provide ahighly valuable tool in the analysis of complex changes inkaryotypes as found in various tumors. This technique willbe complementary to classical chromosome banding andto molecular approaches like SKY and M-FISH whichfinally will allow to define each band of the humankaryotype by molecular probes.
ACKNOWLEDGMENTSWe thank Meena Bagga and Fengtang Yang for technical
support and the Kunming Cell Bank of the ChineseAcademy of Sciences and the Munich Zoo for providinggibbon cells. Clinical samples were kindly provided by Dr.L.Willatt, Addenbrookes Hospital, Cambridge.
LITERATURE CITED
1. Bray-Ward P, Menninger J, Lieman J, Desai T, Mokady N, Banks A,Ward DC: Integration of the cytogenetic, genetic, and physical mapsof the human genome by FISH mapping of CEPH YAC clones.Genomics 32:1–14, 1996.
2. Carter NP, Ferguson-Smith MA, Perryman MT, Telenius H, et al.:Reverse chromosome painting: A method for the rapid analysis ofaberrant chromosomes in clinical cytogenetics. J Mol Genet 29:299–307, 1992.
3. Collins C, Kuo WL, Segraves R, Pinkel D, Fuscoe J, Gray JW:Construction and characterization of plasmid libraries enriched insequences from single human chromosomes. Genomics 11: 997–1006, 1991.
4. Dutrillaux B: Chromosomal evolution in primates: tentative phylog-eny from Microcebus murinus (prosimian) to man. Hum Genet 48:251–314, 1979.
5. Ferguson-Smith MA: Genetic analysis by chromosome sorting andpainting: phylogenetic and diagnostic applications. Europ J HumGenet 5: 253–265, 1997.
451CHROMOSOME COLOUR SEGMENTING

6. Guan XY, Trent JM, Meltzer PS: Generation of band-specific paintingprobes from a single microdissected chromosome. Hum Mol Genet2:1117–1121, 1993.
7. Haaf T, Bray-Ward P: Region-specific YAC banding and paintingprobes for comparative genome mapping: implications for the evolu-tion of human chromosome 2. Chromosoma 104: 537–544, 1997.
8. Jauch A, Wienberg J, Stanyon R, Arnold N, Tofanelli S, Ishida T,Cremer T: Reconstruction of genomic rearrangements in great apesand gibbons by chromosome painting. Proc Natl Acad Sci (USA) 89:8611–8615, 1992.
9. Koehler U, Arnold N, Wienberg J, Tofanelli S, Stanyon R: Genomicreorganization in the concolor gibbon (Hylobates concolor) revealedby chromosome painting. Genomics 30: 287–292, 1995a.
10. Koehler U, Bigoni F, Wienberg J, Stanyon R, Genomic reorganizationand disrupted chromosomal synteny in the Siamang (Hylobatessyndactylus) revealed by fluorescence in situ hybridization. Am J PhysAnthropol 97: 37–47, 1995b.
11. Lengauer C, Speicher MR, Popp S, Jauch A, Taniwaki M, Nagaraja R,Riethman H, Donis-Keller H, D’Urso M, Schlessinger D, Cremer:Chromosomal bar codes produced by multicolour fluorescence in situhybridization with multiple YAC clones and whole chromosomepainting probes. Hum Mol Genet 2: 505–512, 1993.
12. Lichter P, Cremer T, Borden J, Manuelidis L, Ward DC: Delineation ofindividual human chromosomes in metaphase and interphase cells byin situ suppression hybridization using recombinant DNA libraries.Hum Genet 80: 224–234, 1988.
13. Muller S, O’Brien PCM, Ferguson-Smith MA, Wienberg J: A novelsource of highly specific chromosome painting probes derived fromprimate chromosomes for human karyotype analysis. Hum Genet 101:149–153, 1997.
14. Muller S, Rocchi M, Ferguson-Smith MA, Wienberg J: Towards amulticolour chromosome bar code for the entire human karyotype byfluorescence in situ hybridization. Hum Genet 100: 271–278,1997.
15. NIH and IMMC Collaboration. A complete set of human telomericprobes and their clinical application. Nature Genet 14: 86–89, 1996.
16. Nederlof PM, Robinson D, Abunesha R, Wiegant J, Hopman AHN,Tanke HJ, Raap AK: Three colour fluorescence in situ hybridizationfor the simultaneous detection of multiple nucleic acid sequences.Cytometry 10: 20–27, 1989.
17. Nederlof PM, Van der Flier S, Wiegant J, Raap AK, Tanke HJ, Ploem JS,van der Ploeg M: Multiple fluorescence in situ hybridization. Cytome-try 11: 126–131, 1990.
18. Nederlof PM, van der Fleier S, Wiegant J, Raap AK, Tanke HJ, Ploem JS,van der Ploeg M: Fluorescence ratio measurements of double-labelledprobes for multiple in situ hybridization by digital imaging micros-copy. Cytometry 13: 839–845, 1992.
19. Pinkel D, Straume T, Gray JW: Cytogenetic analysis using quantitative,high-sensitivity, fluorescence hybridization. Proc Natl Acad Sci (USA)83: 2934–2938, 1986.
20. Pinkel D, Landegent J, Collins C, Fuscoe J, Segraves R, Lucas J, Gray J:Fluorescence in situ hybridization with human chromosome specificlibraries: Detection of trisomy 21 and translocation of chromosome 4.Proc Natl Acad Sci USA 85: 9138–9142, 1988.
21. Rabbitts P, Impey H, Heppel-Parton A, Langford C, Tease N, Bailey D,Ferguson-Smith MA, Carter NP: Chromosome specific paints from ahigh resolution flow karyotype of the mouse. Nature Genet 9:369–375, 1995.
22. Ried T, Baldini A, Rand TC, Ward DC: Simultaneous visualization ofseven different DNA probes by in situ hybridization using combinato-rial fluorescence and digital imaging microscopy. Proc Natl Acad Sci(USA) 89: 1388–1392, 1992.
23. Schrock E, du Manoir S, Veldman T, Schoell B, Wienberg J, Ferguson-Smith MA, Ledbetter DH, Bar-Am I, Soenksen D, Garini Y, Ried T:Multicolour spectral karyotyping of human chromosomes. Science273: 494–497, 1996.
24. Speicher MR, Ballard GS, Ward DC:Karyotyping human chromosomesby combinatorial multi-fluor FISH. Nature Genet 12: 368–375, 1996.
25. Telenius H, Pelmear, A, Tunnacliffe A, Carter NP, Behmel A, Ferguson-Smith MA, Nordenskjold M, Pfragner R, Ponder B: Cytogeneticanalysis by chromosome painting using DOP-PCR amplified flow-sorted chromosomes. Genes Chromos Cancer 4: 257–263, 1992.
26. Wienberg J, Jauch A, Stanyon R, Cremer T: Molecular cytotaxonomyof primates by chromosomal in situ suppression hybridization.Genomics 8: 347–350, 1990.
27. Wienberg J, Stanyon R, Jauch A, Cremer T: Homologies in human andMacaca fuscata chromosomes revealed by in situ suppression hybrid-ization with human chromosome specific DNA libraries. Chromo-soma 101: 265–270, 1992.
28. Wienberg J, Stanyon R: Comparative painting of mammalian chromo-somes. Curr Opin Genet Develop 7:784–791, 1997.
452 MULLER ET AL.