Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, andresearch funders in the common goal of maximizing access to critical research.
Crayfish occupancy and abundance in lakes of the Pacific Northwest, USAAuthor(s): Eric R. Larson and Julian D. OldenSource: Freshwater Science, 32(1):94-107. 2013.Published By: The Society for Freshwater ScienceDOI: http://dx.doi.org/10.1899/12-051.1URL: http://www.bioone.org/doi/full/10.1899/12-051.1
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, and environmentalsciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies,associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms ofUse, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights andpermissions requests should be directed to the individual publisher as copyright holder.

Crayfish occupancy and abundance in lakes of the PacificNorthwest, USA
Eric R. Larson1AND Julian D. Olden2
School of Aquatic and Fishery Sciences, University of Washington, Seattle, Washington 98195-5020 USA
Abstract. Crayfish are ecologically important in freshwater ecosystems, but few investigators haveevaluated landscape patterns of their prevalence and abundance in temperate lakes. We report results of acrayfish survey for 100 lakes in the Puget Sound lowlands of Washington State (USA) serving as a censusof the distributions of the signal crayfish Pacifastacus leniusculus and previously unknown populations ofnonnative crayfish, including the red swamp crawfish Procambarus clarkii. We modeled crayfish presenceand abundance with classification and regression trees based on multivariate ordinations of lake-, riparian-zone-, and watershed-scale predictors. We found 4 nonnative crayfish in 15% and P. leniusculus in 53% ofsurveyed lakes. The presence of P. leniusculus was best predicted by a lake-scale gradient of greater depth(present) to greater productivity (absent), but forested riparian zones with firm or rocky substrates alsosupported crayfish presence. No models adequately characterized patterns of P. leniusculus abundancemeasured as catch-per-unit effort from traps. Exclusion of data on predatory fish communities and lake pHand Ca may have impeded our ability to characterize patterns of crayfish abundance in the study lakes.Prevalence and abundance of P. leniusculus were low relative to what has been reported for native crayfishin other lake districts or for P. leniusculus in its Asian and European invasive ranges. We hypothesize thatthe erratic distribution of P. leniusculus in Puget Sound lakes may be attributable to challenges ofpostglacial colonization in this region and potentially to human introductions of this crayfish. Our studyexpands the geographic and taxonomic scope of inquiry into relationships between crayfish populationsand lakes at landscape scales and provides a baseline for monitoring and understanding crayfish in anincreasingly urban and invaded region of North America.
Key words: classification and regression trees, crayfish, invasive species, lakes, landscape scale,Pacifastacus leniusculus, Pacific Northwest, Puget Sound, Procambarus clarkii, urbanization.
Temperate lakes are valuable model systems forunderstanding the distribution and abundance oforganisms (Whittaker and Fairbanks 1958, Keddy1976, Browne 1981). Temperate lakes vary in spatialisolation and abiotic conditions, which influence theways organisms colonize, persist, and interact in thesehabitats (Jackson et al. 2001). However, few investi-gators have studied landscape patterns of occupancyand abundance of crayfish in temperate lakes (but seeCapelli and Magnuson 1983, Garvey et al. 2003, Puthand Allen 2004, Olden et al. 2011). Studies have beenlargely confined to a single lake district (Wisconsin,USA) and have had narrow taxonomic scope (Orco-nectes spp.). Crayfish have dispersal capacities, abiotictolerances, and biotic interactions that differ fromthose of zooplankton, aquatic insects, and fish, which
have been used more widely to investigate local andregional processes that structure lake communities(McPeek 1990, Shurin et al. 2000, Olden et al. 2001).Thus, crayfish may provide novel insights into lakepopulation and community dynamics at local andlandscape scales.
Patterns of crayfish occupancy and abundance inlakes are important for conservation and managementof aquatic ecosystems. Half of the crayfish native toNorth America require conservation attention (Tayloret al. 2007), and even some common crayfish speciesare in population decline (Edwards et al. 2009). Incontrast, other crayfish are invasive species withimpacts ranging from displacement of native crayfishto induction of stable-state shifts in aquatic ecosys-tems via foraging and burrowing behaviors (Lodgeet al. 2000, Matsuzaki et al. 2009). Baseline data oncrayfish occupancy and abundance in lakes arevaluable for monitoring the status of native popula-
1 E-mail addresses: [email protected] [email protected]
Freshwater Science, 2013, 32(1):94–107’ 2013 by The Society for Freshwater ScienceDOI: 10.1899/12-051.1Published online: 21 December 2012
94

tions and for identifying and understanding patternsof invasions (Edwards et al. 2009, Olden et al. 2011),but such baselines are exceedingly rare outside of afew well studied regions, such as the midwestern US.
We report results of a survey of crayfish occupancyand abundance in lakes of the Puget Sound lowlands,Washington State, USA. Our study was motivated bythe first discovery of an invasive crayfish populationin the Puget Sound region (Mueller 2001) and theneed to document and anticipate other crayfishinvasions in this lake district. The Puget Sound regionis home to the signal crayfish Pacifastacus leniusculus, amajor invasive species in Asia and Europe that ispoorly understood in its native Pacific Northwestrange (Lodge et al. 2000, Larson and Olden 2011). Oursurvey provides a baseline of P. leniusculus distribu-tion in an increasingly urban and invaded lake district(Francis and Schindler 2006, Larson and Olden 2008)and an understanding of local- and landscape-scalehabitat associations for this species that may bemeaningful to managers where P. leniusculus isinvasive. Our work expands the taxonomic andgeographic scope of inquiry into factors affectingcrayfish occupancy and abundance in temperatelakes.
Methods
Study region and species
The Puget Sound lowlands were carved by a lobe ofthe Cordilleran ice sheet, which retreated 14,000 to12,000 y before present and left a complex estuarinefjord fed by dozens of major and minor riverdrainages and several hundred freshwater lakes(Bortleson et al. 1976, Thorson 1980). The surfacegeology of this region is dominated by glacial till,outwash, and drift, with occasional peat deposits andbedrock intrusions of igneous, metamorphic, andsedimentary origins (WDNR 2010). Land cover priorto European settlement was mixed evergreen anddeciduous forest with scattered prairies, but sizeableportions of the Puget Sound lowlands have beenconverted to urban uses of varying intensities inassociation with cities like Seattle, Tacoma, and theirsuburbs (Alberti et al. 2007). Residential developmenton shorelines has affected Puget Sound lake structureand function (Francis and Schindler 2006, Larson et al.2011), and freshwater communities have been alteredby widespread introductions of nonnative speciesincluding many warmwater sport fish (WDFW 2005).
No previous investigator has studied crayfishdistributions in the Puget Sound lowlands. Pacifasta-cus leniusculus was reported from scattered PugetSound locations by the middle 20th century (Miller
1960, Larson et al. 2012). Pacifastacus leniusculus isnative to the Pacific Northwest region of NorthAmerica and may have colonized Puget Sound lakesduring and following glacial retreat as meltwatercaused Puget Sound to drain southward into theChehalis River glacial refugium (Thorson 1980).Postglacial inundation of Puget Sound lowlands withsalt water may have made northward colonizationdifficult until isostatic rebound raised this regionabove sea level. The east–west orientation of mostPuget Sound rivers further complicates northwardspread of obligate freshwater species (Thorson 1980,McPhail and Lindsey 1986).
A competing explanation for the presence of P.leniusculus in the Puget Sound region is introductionby humans from southern glacial refugia like theColumbia River, the hypothesized origin for invasivepopulations of this species in Japan and Californiaand an area where P. leniusculus has been harvestedcommercially since the late 19th century (Abrahams-son and Goldman 1970, Miller and Van Hyning 1970,Usio et al. 2007). Given the potential challenges ofpostglacial colonization and the success of P. lenius-culus as an invasive species elsewhere, we suggest thiscrayfish be considered cryptogenic (a species thatcannot be assigned definitively to native or human-induced origin; sensu Carlton 1996) in the PugetSound region. In particular, its presence at somelocations, such as lakes on islands in Puget Sound, isdifficult to explain independent of human introduc-tions. Reporting of less ambiguous crayfish invasionsin the Puget Sound region began in 2000, whenMueller (2001) found the red swamp crawfishProcambarus clarkii, a native of southern USA andnortheastern Mexico, established in an urban lakenear Seattle. We began our study to identify thecrayfish species present and to quantify distributionsand abundances relative to lake- and landscape-scaleattributes in the Puget Sound region.
Field sampling
We surveyed 100 lakes in the Puget Soundlowlands between 2007 and 2009 from mid-June toearly October of each year (Fig. 1; Appendix, availableonline at: http://dx.doi.org/10.1899/12-051.1.s1). Ateach lake, we set 20 cylindrical, wire-mesh crayfishtraps (0.42 m long 3 0.21 m diameter with two 60-mmopenings) baited with 120 ml volume of dry, fish-based dog food (Somers and Stechey 1986). Inpreliminary investigations at lakes with moderateabundances of P. leniusculus, catch-per-unit effort(CPUE; number of crayfish/trap) did not differamong traps baited with dry dog food and 2
2013] CRAYFISH IN PACIFIC NORTHWEST LAKES 95

commonly used crayfish baits: canned cat food andbeef liver (see Discussion). The dry dog food was notcontained in a bait bag or box, but did not dissolve ordissipate overnight and remained pungent upon traprecovery. Only in rare cases of extremely high crayfishCPUE was all bait in traps consumed. Our use of 20traps/lake was based on sampling effort in similarstudies (e.g., Capelli and Magnuson 1983, Roth et al.2007) (see Discussion for further comments ontrapping effort and interpretation of CPUE).
We deployed traps in 4 clusters of 5 traps dispersedthroughout the lake by cardinal directions (north,east, south, west). We spaced traps 10 to 30 m apart tomaintain independence based on reported crayfishresponse distances to bait (Abrahamsson and Gold-man 1970, Acosta and Perry 2000). We targetedtrapping at habitats expected to be preferred bycrayfish in the order: rocky substrates, large woodydebris or macrophytes, and open lake bed if preferredhabitats were absent (Capelli and Magnuson 1983,Garvey et al. 2003, Nystrom et al. 2006). We set trapsat depths between 0.5 and 6.0 m (mean = 1.6 m) and
recovered them the next day (mean = 21.4 h, range =
15–26 h). We identified all crayfish collected fromtraps to species and retained voucher specimensfrom each lake for confirmation of identifications andarchiving at a natural history museum. We measuredSecchi disk depth (m) at each lake and characterizedpublic access as an indicator of risk of introduction ofnonnative crayfish. Categories were: 1) boat access =
0 (none; 15% of lakes), 1 (car-top boat launch; 20%),or 2 (boat ramp; 65%); and 2) park = 0 (absent; 63%)or 1 (present; 37%). Parks usually were city parksand less frequently were county parks, state parks, orcampgrounds.
Data sources
The presence and abundance of organisms infreshwater ecosystems is influenced by factors oper-ating across spatial scales from local abiotic and bioticconditions to connectivity with other habitats to landuse in the contributing watershed (Jackson et al. 2001).In freshwater lakes, characteristics like maximum
FIG. 1. Lakes of the Puget Sound lowlands surveyed between 2007 and 2009 for crayfish: Pacifastacus leniusculus, Procambarusclarkii, other species (Orconectes sanbornii, Orconectes virilis, or Procambarus acutus), or no crayfish detected.
96 E. R. LARSON AND J. D. OLDEN [Volume 32
depth and favorable water chemistry influencecrayfish populations (Capelli and Magnuson 1983,Olden et al. 2011). Residential development inriparian zones reduces terrestrial subsidies of foodand habitat, such as large woody debris, to lakes(Francis and Schindler 2006, Larson et al. 2011).Human land use at the whole-watershed scale affectsfreshwater ecosystems by replacing natural vegeta-tion with pasture, cropland, or impervious surfaces,and degrades or destroys small streams that aremigration corridors for aquatic animals (Alberti et al.2007, Elmore and Kaushal 2008). Therefore, we soughtto evaluate determinants of crayfish presence andabundance in the Puget Sound region across 3 scales:lake, riparian zone, and watershed.
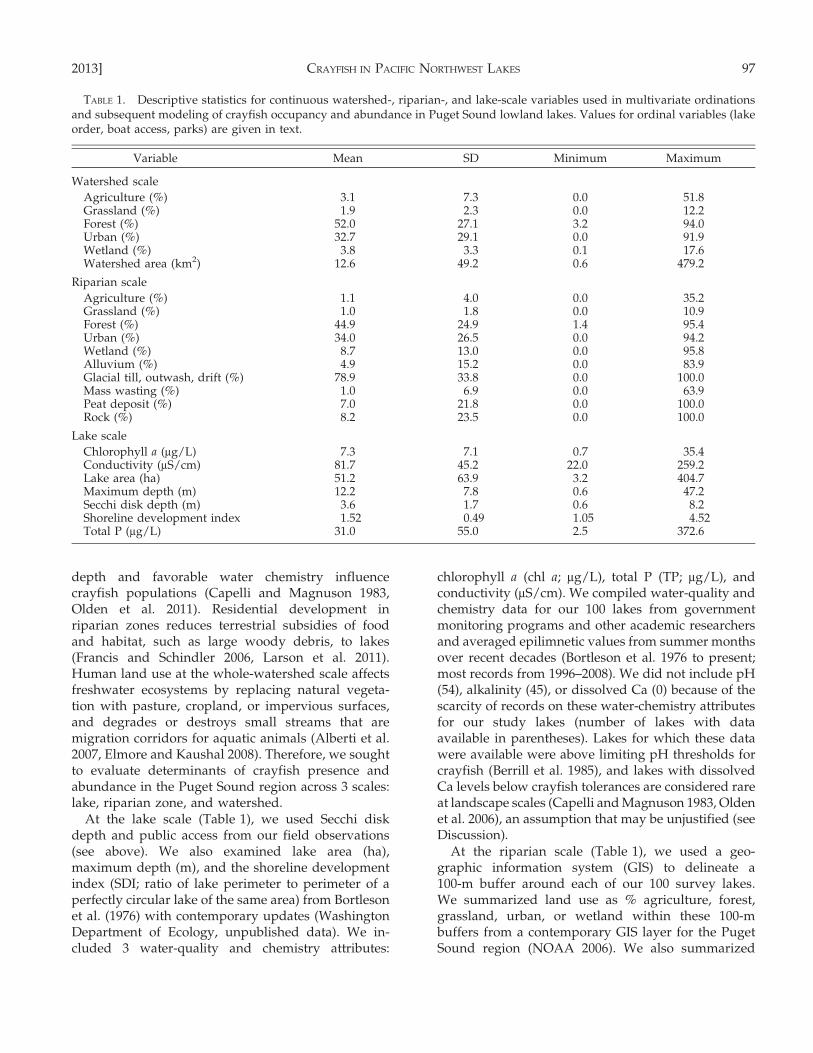
At the lake scale (Table 1), we used Secchi diskdepth and public access from our field observations(see above). We also examined lake area (ha),maximum depth (m), and the shoreline developmentindex (SDI; ratio of lake perimeter to perimeter of aperfectly circular lake of the same area) from Bortlesonet al. (1976) with contemporary updates (WashingtonDepartment of Ecology, unpublished data). We in-cluded 3 water-quality and chemistry attributes:
chlorophyll a (chl a; mg/L), total P (TP; mg/L), andconductivity (mS/cm). We compiled water-quality andchemistry data for our 100 lakes from governmentmonitoring programs and other academic researchersand averaged epilimnetic values from summer monthsover recent decades (Bortleson et al. 1976 to present;most records from 1996–2008). We did not include pH(54), alkalinity (45), or dissolved Ca (0) because of thescarcity of records on these water-chemistry attributesfor our study lakes (number of lakes with dataavailable in parentheses). Lakes for which these datawere available were above limiting pH thresholds forcrayfish (Berrill et al. 1985), and lakes with dissolvedCa levels below crayfish tolerances are considered rareat landscape scales (Capelli and Magnuson 1983, Oldenet al. 2006), an assumption that may be unjustified (seeDiscussion).
At the riparian scale (Table 1), we used a geo-graphic information system (GIS) to delineate a100-m buffer around each of our 100 survey lakes.We summarized land use as % agriculture, forest,grassland, urban, or wetland within these 100-mbuffers from a contemporary GIS layer for the PugetSound region (NOAA 2006). We also summarized
TABLE 1. Descriptive statistics for continuous watershed-, riparian-, and lake-scale variables used in multivariate ordinationsand subsequent modeling of crayfish occupancy and abundance in Puget Sound lowland lakes. Values for ordinal variables (lakeorder, boat access, parks) are given in text.
Variable Mean SD Minimum Maximum
Watershed scale
Agriculture (%) 3.1 7.3 0.0 51.8Grassland (%) 1.9 2.3 0.0 12.2Forest (%) 52.0 27.1 3.2 94.0Urban (%) 32.7 29.1 0.0 91.9Wetland (%) 3.8 3.3 0.1 17.6Watershed area (km2) 12.6 49.2 0.6 479.2
Riparian scale
Agriculture (%) 1.1 4.0 0.0 35.2Grassland (%) 1.0 1.8 0.0 10.9Forest (%) 44.9 24.9 1.4 95.4Urban (%) 34.0 26.5 0.0 94.2Wetland (%) 8.7 13.0 0.0 95.8Alluvium (%) 4.9 15.2 0.0 83.9Glacial till, outwash, drift (%) 78.9 33.8 0.0 100.0Mass wasting (%) 1.0 6.9 0.0 63.9Peat deposit (%) 7.0 21.8 0.0 100.0Rock (%) 8.2 23.5 0.0 100.0
Lake scale
Chlorophyll a (mg/L) 7.3 7.1 0.7 35.4Conductivity (mS/cm) 81.7 45.2 22.0 259.2Lake area (ha) 51.2 63.9 3.2 404.7Maximum depth (m) 12.2 7.8 0.6 47.2Secchi disk depth (m) 3.6 1.7 0.6 8.2Shoreline development index 1.52 0.49 1.05 4.52Total P (mg/L) 31.0 55.0 2.5 372.6
2013] CRAYFISH IN PACIFIC NORTHWEST LAKES 97

surface geology within these riparian buffers becausesome crayfish species prefer firm over soft substratesand use rocky littoral zones of lakes as cover fromaquatic and terrestrial predators (Capelli and Magnu-son 1983, Garvey et al. 2003). The observed preferenceby crayfish for firm and rocky substrates in temperatelakes may be affected by geographic and taxonomicbiases in available studies (see Introduction), andsome crayfish, including many in the genus Procam-barus, may prefer soft or silty substrate for burrowing(Dorn and Volin 2009, Hanshew and Garcia 2012). Wesummarized surface geology within the riparianbuffers as % alluvium, glacial (till, outwash, or drift),mass wasting, peat deposit, or rock using a GIS layerfor Washington State surface geology (WDNR 2010).
At the watershed scale (Table 1), we used GISlayers and tools in the National Hydrography Dataset(NHD; USGS 2010) to delineate contributing water-sheds for each lake by navigating upstream throughflow lines from lake outlets to include all contributingwatershed areas, or in the absence of outlets, byselecting the watershed containing the lake. Wesummarized % land use within these watersheds asabove for riparian buffers and included watershedarea (km2) in our analysis. Last, we used lake order(Martin and Soranno 2006) to represent connectivityand colonization potential for crayfish from otherfreshwater habitats (lakes, streams, or wetlands). Wedefined lake order as lacking inlet or outlet streams(–1; 45% of lakes), having inlet stream(s) only (0; 26%),or having both inlet and outlet streams (1; 29%) basedon NHD flow lines validated by contemporary aerialphotographs (Google EarthH) and field site observa-tions. We included lake order as a watershed attributein anticipation of an association with watershed areaand the potential for land uses, such as urbanization,to affect lake order through burying or diversion ofstreams (Elmore and Kaushal 2008).
Statistical analyses
We used multivariate ordinations to characterizestudy sites at lake, riparian, and watershed scales andretained ordination axes that represented dominantgradients of variation for subsequent models of crayfishoccupancy and abundance. Many of our potentialpredictors, such as % landuse categories or lake depth,Secchi disk depth, and chl a, are not independent ofeach other but can be simplified by ordination into areduced set of orthogonal variables (Beals 2006). Weused Principal Components Analysis (PCA) on corre-lation matrices to generate ordinations for riparian andwatershed scales, and Principal Coordinates Analysis(PCoA) on a distance matrix generated with Gower’s
resemblance coefficient for the lake scale. Conductivity(n = 5) or TP and chl a (n = 18) data were unavailablefor a minority of lakes. These lakes would have had tobe omitted from PCA on correlation matrices or thedata substituted with average values, but Gower’scoefficient accounts for missing data by excludingpredictors from pairwise distance calculations if absentfrom one or both subjects (Pavoine et al. 2009). We ranordinations in the vegan library in R (version 2.11.1; RDevelopment Core Team, Vienna, Austria).
We modeled occupancy as presence (1) or absence(0) in any case where crayfish were detected (CPUE§ 0.05), and we modeled abundance as CPUE(mean number of crayfish/trap at each lake; range= 0–7.70). We included both measures becauseoccurrence can be more tractable than abundance,but identifying conditions that favor high crayfishCPUE might be useful in understanding invasivepopulations, which often are more abundant thannative species (Garvey et al. 2003). Occupancy andabundance are vulnerable to imperfect detectionof organisms (Royle et al. 2005), but CPUE fromcrayfish traps is a standard method that can be usedto monitor temporal trends in distribution andpopulation size (Edwards et al. 2009).
We used classification and regression trees tomodel crayfish occupancy and abundance (dependentvariables) as responses to the first 2 ordination axes(independent variables) from each scale of analysis.We included only the first 2 axes because subsequentordination axes explained small proportions ofvariation in the original variables and were notinfluential in preliminary tree analyses. We usednonnative crayfish (all species) CPUE as a predictorfor P. leniusculus because biotic interactions with othercrayfish could affect distribution and populationpatterns for this species (Garvey et al. 2003, Olden etal. 2011). Classification and regression trees use arecursive partitioning algorithm to split data into endnodes that maximize group homogeneity, whethermeasured by distribution for categorical responsesor mean values for continuous responses (De’athand Fabricius 2000). Single trees are easy to interpretvisually and are robust to a variety of limitations ofother modeling methods. Trees are unaffected bydiffering scales, distributions, or types of predictorvariables; are insensitive to outliers; and account forinteractions with their hierarchical structure (De’athand Fabricius 2000). Single trees can produce poorlyperforming models in cases of small sample sizesand over certain response functions, but ensembleapproaches that develop and average a large numberof trees can overcome these problems (Elith et al.2008).
98 E. R. LARSON AND J. D. OLDEN [Volume 32

We used the boosted ensemble tree method ofElith et al. (2008) to model patterns of crayfishoccupancy and abundance. Relative importance ofpredictor variables from boosted trees, calculated asthe number of times a variable was used weightedby model improvement and averaged across alltrees, was retained and interpreted as the influenceof spatial scales on crayfish occupancy and abun-dance. We developed and reported a single classi-fication tree for occupancy and a single regressiontree for abundance following the methods of De’athand Fabricius (2000) to visualize and interpret ourresults. We report correct classification rates, sensi-tivity (true positive rate), specificity (true negativerate), and Cohen’s k coefficient of agreement asmeasures of individual tree performance. We usedrpart and gbm libraries in R with additional code byElith et al. (2008) to run classification and regressiontree analyses. Trees were supplemented withadditional descriptive statistics and results whenpertinent.
Results
We detected crayfish in 63 of 100 surveyed lakes(Fig. 1, Appendix). Pacifastacus leniusculus was foundin 53% of lakes, whereas P. clarkii was detected in 11%
of lakes. We encountered an unexpectedly diverseassemblage of other nonnative crayfish in the PugetSound region. We found virile crayfish Orconectesvirilis in 2 lakes (Ballinger, Spanaway), Sanborn’scrayfish Orconectes sanbornii in 1 lake (Big), and thewhite river crawfish Procambarus acutus in 1 lake(Echo). Nonnative crayfish identifications were con-firmed by Chris A. Taylor and voucher specimenswere deposited at the Illinois Natural History Survey(Champaign, Illinois). The low prevalence of mostnonnative crayfish species prevented us from model-ing their occupancy and abundance. We attribute thehigh degree of spatial aggregation observed for P.clarkii (Fig. 1) to local releases from schools that usethis crayfish in the classroom (Larson and Olden2008). These local events reduced our ability togeneralize P. clarkii habitat associations across thebroader landscape, so we adjusted our analysis tofocus on P. leniusculus with occasional discussion ofnonnative crayfish.
At the watershed scale, the 1st principal component(axis) represented lakes on a gradient from moreurbanized (positive) to more natural land cover,larger watershed areas, and higher lake orders(negative) (Fig. 2A). The 2nd watershed axis repre-sented lakes with relatively more (positive) or less(negative) agricultural or wetland land cover. At the
riparian scale, the 1st axis represented a gradient oflakes with more natural land cover (positive) to lakeswith more urbanized land cover and glacial geologies(negative), whereas the 2nd axis represented a gradientof lakes with agricultural or wetland land covers andsoft geologies (positive) to lakes with forested riparianzones and rocky substrates (negative) (Fig. 2B). At thelake scale, the 1st axis separated highly productive lakeswith high chl a and TP values (positive) from deeperlakes with greater Secchi disk depths (negative),whereas lakes with larger surface areas and higherSDI values loaded positively on the 2nd axis (Fig. 2C).The first 2 axes of all ordinations were significant(Monte Carlo randomization, 999 permutations; p ,
0.001), but no subsequent axes were significant (all p .
0.500). See Table 2 for correlation coefficients amongordination axes for the 3 scales of analysis.
Our final boosted classification tree for P. leniuscu-lus presence was developed at a learning rate of 0.001,with a tree size of 4, and used just over 1000 trees perthe recommendations of Elith et al. (2008). The 1st lakeaxis was the most important predictor variable,riparian and watershed axes were intermediate, andnonnative crayfish CPUE was the least important(Fig. 3A). The individual classification tree used 2splits and 3 end nodes to model P. leniusculusoccupancy with a 75% correct classification rate,79% sensitivity (42 true presences to 11 false absen-ces), 70% specificity (33 true absences to 14 falsepresences), and a moderate Cohen’s k of 0.496 (p ,
0.001) (Fig. 3B). The 1st split separated sites on the 1st
lake axis of greater lake depth and clarity (P.leniusculus present) to greater lake productivity (P.leniusculus absent), whereas the 2nd split separatedsites on the 2nd riparian axis of forested riparian zoneswith rock substrates (P. leniusculus present) to soft-substrate riparian zones with wetland or agriculturalland covers (P. leniusculus absent).
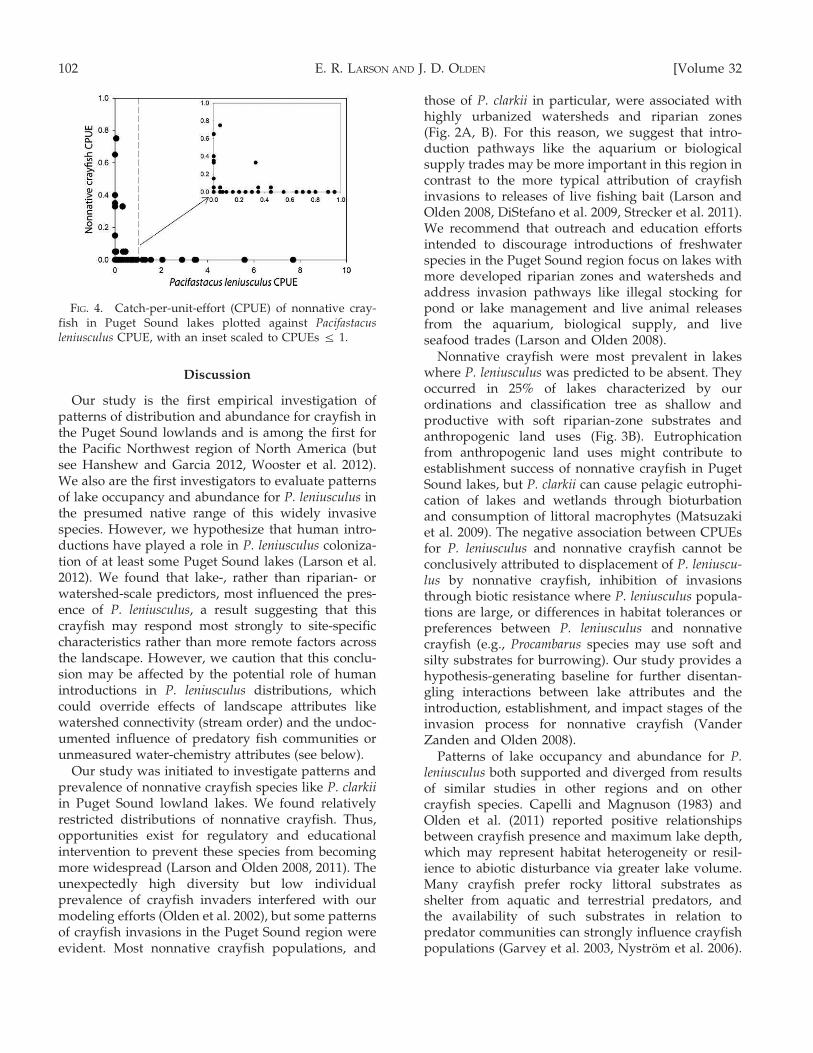
A range of learning rates, tree sizes, and number oftrees did not produce improvements in predictivedeviance for boosted trees of P. leniusculus abundance.Similar difficulties were encountered in developmentof an individual regression tree for abundancebecause no improvement with increases in modelcomplexity (tree size) was detected by cross-validatedrelative error (De’ath and Fabricius 2000). Removingall sites where P. leniusculus was not detected (0CPUE) did not improve model performance. There-fore, we report P. leniusculus abundance as box plotsof CPUE in our classification tree for presence(Fig. 3B). We provide pie charts of nonnative crayfishpresence in terminal nodes of this classification tree(Fig. 3B) and plots of P. leniusculus CPUE vs nonna-tive crayfish CPUE (Fig. 4).
2013] CRAYFISH IN PACIFIC NORTHWEST LAKES 99
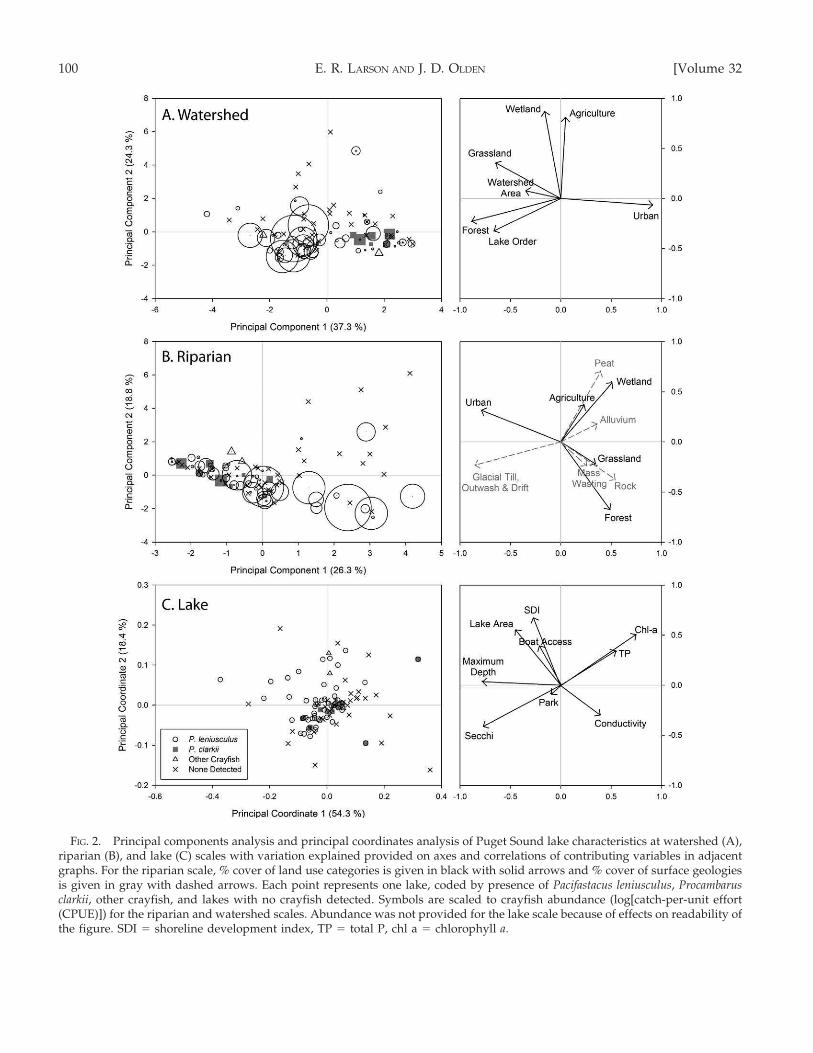
FIG. 2. Principal components analysis and principal coordinates analysis of Puget Sound lake characteristics at watershed (A),riparian (B), and lake (C) scales with variation explained provided on axes and correlations of contributing variables in adjacentgraphs. For the riparian scale, % cover of land use categories is given in black with solid arrows and % cover of surface geologiesis given in gray with dashed arrows. Each point represents one lake, coded by presence of Pacifastacus leniusculus, Procambarusclarkii, other crayfish, and lakes with no crayfish detected. Symbols are scaled to crayfish abundance (log[catch-per-unit effort(CPUE)]) for the riparian and watershed scales. Abundance was not provided for the lake scale because of effects on readability ofthe figure. SDI = shoreline development index, TP = total P, chl a = chlorophyll a.
100 E. R. LARSON AND J. D. OLDEN [Volume 32
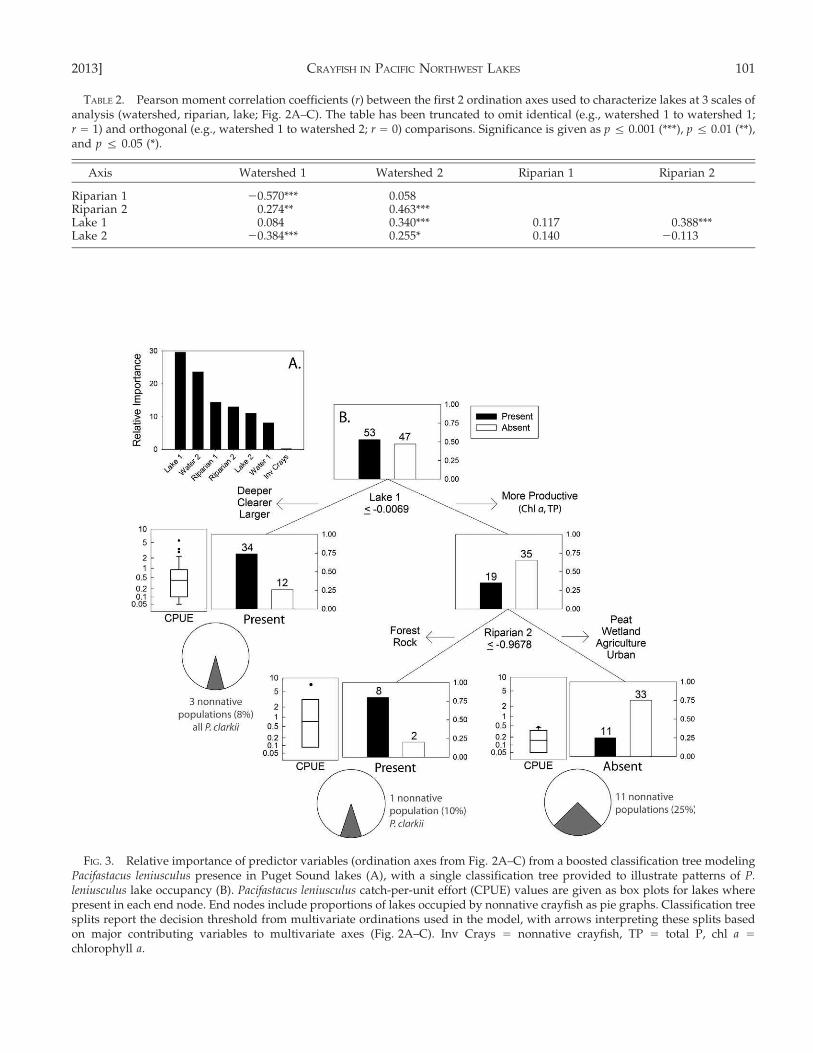
TABLE 2. Pearson moment correlation coefficients (r) between the first 2 ordination axes used to characterize lakes at 3 scales ofanalysis (watershed, riparian, lake; Fig. 2A–C). The table has been truncated to omit identical (e.g., watershed 1 to watershed 1;r = 1) and orthogonal (e.g., watershed 1 to watershed 2; r = 0) comparisons. Significance is given as p ƒ 0.001 (***), p ƒ 0.01 (**),and p ƒ 0.05 (*).
Axis Watershed 1 Watershed 2 Riparian 1 Riparian 2
Riparian 1 20.570*** 0.058Riparian 2 0.274** 0.463***Lake 1 0.084 0.340*** 0.117 0.388***Lake 2 20.384*** 0.255* 0.140 20.113
FIG. 3. Relative importance of predictor variables (ordination axes from Fig. 2A–C) from a boosted classification tree modelingPacifastacus leniusculus presence in Puget Sound lakes (A), with a single classification tree provided to illustrate patterns of P.leniusculus lake occupancy (B). Pacifastacus leniusculus catch-per-unit effort (CPUE) values are given as box plots for lakes wherepresent in each end node. End nodes include proportions of lakes occupied by nonnative crayfish as pie graphs. Classification treesplits report the decision threshold from multivariate ordinations used in the model, with arrows interpreting these splits basedon major contributing variables to multivariate axes (Fig. 2A–C). Inv Crays = nonnative crayfish, TP = total P, chl a =
chlorophyll a.
2013] CRAYFISH IN PACIFIC NORTHWEST LAKES 101
Discussion
Our study is the first empirical investigation ofpatterns of distribution and abundance for crayfish inthe Puget Sound lowlands and is among the first forthe Pacific Northwest region of North America (butsee Hanshew and Garcia 2012, Wooster et al. 2012).We also are the first investigators to evaluate patternsof lake occupancy and abundance for P. leniusculus inthe presumed native range of this widely invasivespecies. However, we hypothesize that human intro-ductions have played a role in P. leniusculus coloniza-tion of at least some Puget Sound lakes (Larson et al.2012). We found that lake-, rather than riparian- orwatershed-scale predictors, most influenced the pres-ence of P. leniusculus, a result suggesting that thiscrayfish may respond most strongly to site-specificcharacteristics rather than more remote factors acrossthe landscape. However, we caution that this conclu-sion may be affected by the potential role of humanintroductions in P. leniusculus distributions, whichcould override effects of landscape attributes likewatershed connectivity (stream order) and the undoc-umented influence of predatory fish communities orunmeasured water-chemistry attributes (see below).
Our study was initiated to investigate patterns andprevalence of nonnative crayfish species like P. clarkiiin Puget Sound lowland lakes. We found relativelyrestricted distributions of nonnative crayfish. Thus,opportunities exist for regulatory and educationalintervention to prevent these species from becomingmore widespread (Larson and Olden 2008, 2011). Theunexpectedly high diversity but low individualprevalence of crayfish invaders interfered with ourmodeling efforts (Olden et al. 2002), but some patternsof crayfish invasions in the Puget Sound region wereevident. Most nonnative crayfish populations, and
those of P. clarkii in particular, were associated withhighly urbanized watersheds and riparian zones(Fig. 2A, B). For this reason, we suggest that intro-duction pathways like the aquarium or biologicalsupply trades may be more important in this region incontrast to the more typical attribution of crayfishinvasions to releases of live fishing bait (Larson andOlden 2008, DiStefano et al. 2009, Strecker et al. 2011).We recommend that outreach and education effortsintended to discourage introductions of freshwaterspecies in the Puget Sound region focus on lakes withmore developed riparian zones and watersheds andaddress invasion pathways like illegal stocking forpond or lake management and live animal releasesfrom the aquarium, biological supply, and liveseafood trades (Larson and Olden 2008).
Nonnative crayfish were most prevalent in lakeswhere P. leniusculus was predicted to be absent. Theyoccurred in 25% of lakes characterized by ourordinations and classification tree as shallow andproductive with soft riparian-zone substrates andanthropogenic land uses (Fig. 3B). Eutrophicationfrom anthropogenic land uses might contribute toestablishment success of nonnative crayfish in PugetSound lakes, but P. clarkii can cause pelagic eutrophi-cation of lakes and wetlands through bioturbationand consumption of littoral macrophytes (Matsuzakiet al. 2009). The negative association between CPUEsfor P. leniusculus and nonnative crayfish cannot beconclusively attributed to displacement of P. leniuscu-lus by nonnative crayfish, inhibition of invasionsthrough biotic resistance where P. leniusculus popula-tions are large, or differences in habitat tolerances orpreferences between P. leniusculus and nonnativecrayfish (e.g., Procambarus species may use soft andsilty substrates for burrowing). Our study provides ahypothesis-generating baseline for further disentan-gling interactions between lake attributes and theintroduction, establishment, and impact stages of theinvasion process for nonnative crayfish (VanderZanden and Olden 2008).
Patterns of lake occupancy and abundance for P.leniusculus both supported and diverged from resultsof similar studies in other regions and on othercrayfish species. Capelli and Magnuson (1983) andOlden et al. (2011) reported positive relationshipsbetween crayfish presence and maximum lake depth,which may represent habitat heterogeneity or resil-ience to abiotic disturbance via greater lake volume.Many crayfish prefer rocky littoral substrates asshelter from aquatic and terrestrial predators, andthe availability of such substrates in relation topredator communities can strongly influence crayfishpopulations (Garvey et al. 2003, Nystrom et al. 2006).
FIG. 4. Catch-per-unit-effort (CPUE) of nonnative cray-fish in Puget Sound lakes plotted against Pacifastacusleniusculus CPUE, with an inset scaled to CPUEs ƒ 1.
102 E. R. LARSON AND J. D. OLDEN [Volume 32

Pacifastacus leniusculus presence was associated withfirmer or rockier riparian surface geologies andabsences were associated with riparian zones charac-terized by peat deposits, alluvium, or wetlands.However, Usio et al. (2006) provided an example ofdisparate habitat use by invasive P. leniusculus inwetlands without rocky substrate. Pacifastacus lenius-culus presence also was associated with forestedriparian zones, which we suggest may benefit crayfishvia subsidies of large woody debris as habitat andterrestrial detritus as an energy source (Francis andSchindler 2006, Larson et al. 2011). For example, thehighest P. leniusculus abundances were observed inlakes with riparian zones and watersheds character-ized by forested rather than agricultural or urban landcover (Fig. 2A).
Despite this apparent pattern, regression treemodels failed to predict P. leniusculus abundancebased on our lake-, riparian-, and watershed-scalepredictors. Poor performance of P. leniusculus abun-dance models might be attributable to lack of data onfish communities in our study lakes. Predatory fishcan strongly influence the presence, abundance, andtrapability of crayfish in lakes (Collins et al. 1983,Garvey et al. 2003, Roth et al. 2007). Predatory fishfound in the Puget Sound region, such as the nativeNorthern Pikeminnow Ptychocheilus oregonensis andnonnative black bass of the genus Micropterus, can behighly efficient predators on crayfish (Poe et al. 1991).We used published (WDFW 2005) and unpublishedfish records from the Washington Department of Fishand Wildlife to document the presence of Micropterusdolomieu or salmoides in 74% of our lakes, the YellowPerch Perca flavescens in 58%, and sunfishes of thegenus Lepomis in 49%. However, many of our lakeshad no contemporary fish records, and fish abun-dance may be more meaningful than presence inexplaining crayfish population processes (Garvey etal. 2003, Roth et al. 2007). Investigation of the effects offish predators on Pacific Northwest crayfish popula-tions is needed, particularly because cambarid cray-fish introduced to this region from eastern NorthAmerica share evolutionary history with the nonna-tive warmwater fish mentioned above, and differen-tial susceptibility to predatory fish is an importantmechanism in crayfish species displacements (Garveyet al. 1994, Mueller and Bodensteiner 2009).
One major difference in our results relative to paststudies of crayfish–lake associations was our lowobserved prevalence and abundance for P. leniusculusin particular and all crayfish in general. Capelli andMagnuson (1983) reported crayfish present in 75% of67 sampled lakes in Wisconsin, Puth and Allen (2004)reported crayfish present in 88% of 35 lakes in this
same lake district, and Edwards et al. (2009) reporteddeclines in crayfish presence from 75% of 100 Ontariolakes from 1989 to 1995 to 67% of the same lakes from2005 to 2007. By contrast, we found crayfish present inonly 63% and the presumed native P. leniusculus inonly 53% of 100 Puget Sound lakes. Abundances, asmeasured by CPUE, also were conspicuously lowerfor P. leniusculus in Puget Sound lakes relative to otherregions and species. Capelli and Magnuson (1983)reported maximum CPUEs of 29.7 crayfish/trap, andGarvey et al. (2003) reported maximum CPUEs .40crayfish/trap for the invasive crayfish O. rusticus.
Our mean CPUE of 0.8 6 1.4 (SD), excluding lakeswhere this crayfish was not detected (0 CPUE), isalso low relative to records from regions whereP. leniusculus is invasive. Usio et al. (2007) reporteda mean CPUE of 7.4 6 5.3 for P. leniusculus from 8lakes in Japan, and Nystrom et al. (2006) reported amean CPUE of 5.6 6 3.1 for P. leniusculus from 10lakes in Sweden. We previously used stable isotopesto show that the broadly omnivorous trophic niche ofP. leniusculus was conserved between its WashingtonState native range and lakes of northern Japan (Larsonet al. 2010), but results here suggest that invasiveimpacts of P. leniusculus might be attributable insteadto higher invasive population densities rather thanany change in ecological function of individual P.leniusculus (Pintor et al. 2009).
Our study may differ from those in other regionsbecause of sampling method or geologic or geomor-phic differences among lake districts. We used asimilar number of traps over equivalent durationsand lake depths as in most of the studies cited above,and our design differed mostly in bait choice. LikeSomers and Stechey (1986), we did not find significantdifferences in CPUE between traps baited with drydog food and traps baited with the canned cat food(Edwards et al. 2009) or beef liver (Capelli andMagnuson 1983, Puth and Allen 2004) used in otherstudies. We do not think this difference in baits canaccount for the low observed prevalence of P.leniusculus in lakes throughout the Puget Soundregion. The ability of CPUE from traps to representcrayfish relative abundance has been evaluatedagainst other measures like mark–recapture popula-tion estimates (Zimmerman and Palo 2011) and visualsurveys by divers (Collins et al. 1983, Olsen et al.1991). These investigators found moderate-to-excellentconcordance between CPUE and other measures ofcrayfish relative abundance. Zimmerman and Palo(2011) observed that CPUE better matched mark–recapture population estimates at higher trappingeffort (R2
= 0.23 at 15 traps, R2= 0.46 at 120 traps),
whereas Capelli and Magnuson (1983) found a strong
2013] CRAYFISH IN PACIFIC NORTHWEST LAKES 103

relationship between intensive lake-wide estimates ofcrayfish biomass from dive surveys and CPUE fromonly 15–18 traps/lake (R2
= 0.94). We acknowledgethat our effort of 20 traps/lake might fail to detectcrayfish presence at low densities, but we note thatCPUE from low trapping efforts and at low crayfishdensities often overestimates relative abundance (e.g.,Olsen et al. 1991, Zimmerman and Palo 2011) in aphenomenon known as hyperstability (Harley et al.2001). Therefore, we think our observation of low P.leniusculus presence and abundance in Puget Soundlowland lakes relative to other regions and speciesprobably is robust.
We could not include pH or dissolved Ca in ourmodels because these data are scarce in monitoringrecords for Puget Sound lowland lakes, probablybecause the minimal impact of acid deposition in ourstudy region relative to some lake districts (i.e., Rodhe1989) has led to a low priority for monitoring thesewater-chemistry attributes. No dissolved Ca valueswere available from our study lakes, but 54 of ourlakes had pH records (mean 7.2, range 6.4–8.6), allabove proposed minimum thresholds for crayfish(Berrill et al. 1985, Olden et al. 2006), and 45 of our lakeshad alkalinity records (mean = 29.7 mg CaCO3/L,range 5.0–84.6 mg CaCO3/L). Greaver et al. (2012)reported that freshwaters of western Washingtonhave moderate to low acid-neutralizing capacity,suggesting potential for impairment from ongoing Nand S deposition. Ca declines in lakes and theirwatershed also have been attributed to land usepractices, such as forest harvest (Jeziorski et al. 2008),and is an issue that may warrant greater scrutiny inthe Puget Sound lowlands. Last, the availability ofrocky littoral substrates as cover from predation andcannibalism is often an important predictor ofcrayfish abundance (Capelli and Magnuson 1983,Garvey et al. 2003, Nystrom et al. 2006), and perhapslakes of the Puget Sound region have less rockysubstrates than other regions with greater crayfishprevalence and abundance. Focused surveys onsubstrate composition in a subset of our study lakesfollowing methods like those of Garvey et al. (2003)could address this question.
In conclusion, lakes of the Puget Sound region wereinfrequently occupied by an unexpectedly diversecommunity of nonnative crayfish dominated by P.clarkii. We also found the presumed native crayfish P.leniusculus to be less prevalent and at lower abun-dances than what has been reported for other nativecrayfish species in other lake districts, or for P.leniusculus elsewhere (i.e., its invasive range in Asiaand Europe). Lake-scale predictors were more influ-ential than riparian and watershed-scale predictors in
model development for P. leniusculus presence, al-though the failure of our models to explain patterns ofP. leniusculus abundance could be caused by the lackof data on predatory fish communities or by theresolution of our data on lake substrate composition(i.e., riparian surface geology). Our study establishes abaseline for monitoring regional change in crayfishcommunities in Puget Sound lakes and demonstratesthat opportunities remain for preventing additionalnonnative crayfish invasions in this lake district. Manyimportant questions remain with respect to definingthe native range of P. leniusculus in the PacificNorthwest, as well as identifying factors that contrib-ute to its invasive success elsewhere in the world.
Acknowledgements
This research was supported by funding fromAchievement Rewards for College Scientists (ARCSFoundation), Washington Sea Grant, the Oregon ZooFuture for Wildlife Grants Program, the WashingtonLake Protection Association Nancy Weller Fellow-ship, and an Anchor QEA Scholarship. Sampling wasconducted under research permits from the Wash-ington Department of Fish and Wildlife (07 to 09-9317) and Washington State Parks and RecreationCommission (090604). J. Benca, F. Lin, M. C. Mims,K. W. Mueller, T. K. Pool, K. Ung, C. A. Wisniewski,and others contributed to field sampling. M. Tamayoassisted with compiling the database of lake charac-teristics, and R. Matthews (Western WashingtonUniversity) generously provided additional lake data.J. W. Adams, D. A. Beauchamp, M. T. Brett, S. S.Chan, B. Cullen, J. J. Lawler, E. Lowery, K. W.Mueller, D. Rose, D. E. Schindler, C. J. Sergeant, andmany others provided suggestions or logisticalsupport in conducting this research. Comments fromA. L. Strecker, M. A. Chadwick, P. Silver, and 3anonymous referees improved this manuscript.
Literature Cited
ABRAHAMSSON, S. A. A., AND C. R. GOLDMAN. 1970. Distribu-tion, density and production of the crayfish Pacifastacusleniusculus Dana in Lake Tahoe, California-Nevada.Oikos 21:83–91.
ACOSTA, C. A., AND S. A. PERRY. 2000. Effective sampling area:a quantitative method for sampling crayfish popula-tions in freshwater marshes. Crustaceana 73:425–431.
ALBERTI, M., D. BOOTH, K. HILL, B. COBURN, C. AVOLIO, S. COE,AND D. SPIRANDELLI. 2007. The impact of urban patterns onaquatic ecosystems: an empirical analysis in Puget lowlandsub-basins. Landscape and Urban Planning 80:345–361.
BEALS, M. L. 2006. Understanding community structure: adata-driven multivariate approach. Oecologia (Berlin)150:484–495.
104 E. R. LARSON AND J. D. OLDEN [Volume 32

BERRILL, M., L. HOLLETT, A. MARGOSIAN, AND J. HUDSON. 1985.Variation in tolerance to low environmental pH by thecrayfish Orconectes rusticus, O. propinquus, and Cambarusrobustus. Canadian Journal of Zoology 63:2586–2589.
BORTLESON, G. C., N. P. DION, J. B. MCCONNELL, AND L. M.NELSON. 1976. Reconnaissance data on lakes in Wash-ington. Water-Supply Bulletin 43. Washington Depart-ment of Ecology, Tacoma, Washington.
BROWNE, R. A. 1981. Lakes as islands: biogeographicdistribution, turnover rates, and species compositionin the lakes of central New York. Journal of Biogeogra-phy 8:75–83.
CAPELLI, G. M., AND J. T. MAGNUSON. 1983. Morphoedaphicand biogeographic analysis of crayfish distribution innorthern Wisconsin. Journal of Crustacean Biology 3:548–564.
CARLTON, J. T. 1996. Biological invasions and cryptogenicspecies. Ecology 77:1653–1655.
COLLINS, N. C., H. H. HARVEY, A. TIERNEY, AND D. W. DUNHAM.1983. Influence of predator density on trapability ofcrayfish in Ontario lakes. Canadian Journal of Fisheriesand Aquatic Sciences 40:1820–1828.
DE’ATH, G., AND K. E. FABRICIUS. 2000. Classification andregression trees: a powerful yet simple technique forecological data analysis. Ecology 81:3178–3192.
DISTEFANO, R. J., M. E. LITVAN, AND P. T. HORNER. 2009. Thebait industry as a potential vector for alien crayfishintroductions: problem recognition by fisheries agenciesand a Missouri evaluation. Fisheries 34(12):586–597.
DORN, N. J., AND J. C. VOLIN. 2009. Resistance of crayfish(Procambarus spp.) populations to wetland dryingdepends on species and substrate. Journal of the NorthAmerican Benthological Society 28:766–777.
EDWARDS, B. A., D. A. JACKSON, AND K. M. SOMERS. 2009.Multispecies crayfish declines in lakes: implications forspecies distributions and richness. Journal of the NorthAmerican Benthological Society 28:719–732.
ELITH, J., J. R. LEATHWICK, AND T. HASTIE. 2008. A workingguide to boosted regression trees. Journal of AnimalEcology 77:802–813.
ELMORE, A. J., AND S. S. KAUSHAL. 2008. Disappearingheadwaters: patterns of stream burial due to urbaniza-tion. Frontiers in Ecology and the Environment 6:308–312.
FRANCIS, T. B., AND D. E. SCHINDLER. 2006. Degradation oflittoral habitats by residential development: woodydebris in lakes of the Pacific Northwest and Midwest,United States. Ambio 35:274–280.
GARVEY, J. E., J. E. RETTIG, R. A. STEIN, D. M. LODGE, AND S. P.KLOSIEWSKI. 2003. Scale-dependent associations amongfish predation, littoral habitat, and distributions ofcrayfish species. Ecology 84:3339–3348.
GARVEY, J. E., R. A. STEIN, AND H. M. THOMAS. 1994. Assessinghow fish predation and interspecific prey competitioninfluence a crayfish assemblage. Ecology 75:532–547.
GREAVER, T. L., T. J. SULLIVAN, J. D. HERRICK, M. C. BARBER, J. S.BARON, B. J. COSBY, M. E. DEERHAKE, R. L. DENNIS, J.-J. B.DUBOIS, C. L. GOODALE, A. T. HERLIHY, G. B. LAWRENCE, L.LIU, J. A. LYNCH, AND K. J. NOVAK. 2012. Ecological effects
of nitrogen and sulfur air pollution in the US: what dowe know? Frontiers in Ecology and Environment 10:365–372.
HANSHEW, B. A., AND T. S. GARCIA. 2012. Invasion of theshelter snatchers: behavioural plasticity in invasive redswamp crayfish, Procambarus clarkii. Freshwater Biology57:2285–2296.
HARLEY, S. J., R. A. MYERS, AND A. DUNN. 2001. Is catch-per-unit-effort proportional to abundance? Canadian Jour-nal of Fisheries and Aquatic Sciences 58:1760–1772.
JACKSON, D. A., P. R. PERES-NETO, AND J. D. OLDEN. 2001. Whatcontrols who is where in freshwater fish communities—the roles of biotic, abiotic, and spatial factors. CanadianJournal of Fisheries and Aquatic Sciences 58:157–170.
JEZIORSKI, A., N. D. YAN, A. M. PATERSON, A. M. DESELLAS,M. A. TURNER, D. S. JEFFRIES, B. KELLER, R. C. WEEBER, D. K.MCNICOL, M. E. PALMER, K. MCIVER, K. ARSENEAU, B. K.GINN, B. F. CUMMING, AND J. P. SMOL. 2008. Thewidespread threat of calcium decline in fresh waters.Science 322:1374–1377.
KEDDY, P. A. 1976. Lakes as islands: the distributionalecology of two aquatic plants, Lemna minor L. and L.trisulca L. Ecology 57:353–359.
LARSON, E. R., C. L. ABBOTT, N. USIO, N. AZUMA, K. A. WOOD,L.-M. HERBORG, AND J. D. OLDEN. 2012. The signal crayfishis not a single species: cryptic diversity and invasions inthe Pacific Northwest range of Pacifastacus leniusculus.Freshwater Biology 57:1823–1838.
LARSON, E. R., AND J. D. OLDEN. 2008. Do schools and golfcourses represent emerging pathways for crayfishinvasions? Aquatic Invasions 3:465–468.
LARSON, E. R., AND J. D. OLDEN. 2011. The state of crayfish inthe Pacific Northwest. Fisheries 36:60–73.
LARSON, E. R., J. D. OLDEN, AND N. USIO. 2010. Decoupledconservatism of Grinnellian and Eltonian niches in aninvasive arthropod. Ecosphere 1(6):art16. doi:10.1890/ES10-00053.1
LARSON, E. R., J. D. OLDEN, AND N. USIO. 2011. Shorelineurbanization interrupts allochthonous subsidies to abenthic consumer over a gradient of lake size. BiologyLetters 7:551–554.
LODGE, D. M., C. A. TAYLOR, D. M. HOLDICH, AND J. SKURDAL.2000. Nonindigenous crayfishes threaten North Amer-ican freshwater biodiversity: lessons from Europe.Fisheries 25(8):7–20.
MARTIN, S. L., AND P. A. SORANNO. 2006. Lake landscapeposition: relationships to hydrologic connectivity andlandscape features. Limnology and Oceanography 51:801–814.
MATSUZAKI, S. S., N. USIO, N. TAKAMURA, AND I. WASHITANI.2009. Contrasting impacts of invasive engineers onfreshwater ecosystems: an experiment and meta-analy-sis. Oecologia (Berlin) 158:673–685.
MCPEEK, M. A. 1990. Determination of species compositionin the Enallagma damselfly assemblages of permanentlakes. Ecology 71:83–98.
MCPHAIL, J. D., C. C. LINDSEY. 1986. Zoogeography of thefreshwater fishes of Cascadia (The Columbia Systemand Rivers North to the Stikine). Pages 615–638 in C. H.
2013] CRAYFISH IN PACIFIC NORTHWEST LAKES 105

Hocutt, and E. O. Wiley. 1986. The zoogeography ofNorth American freshwater fishes. John Wiley and Sons,New York.
MILLER, G. C. 1960. The taxonomy and certain biologicalaspects of the crayfish of Oregon and Washington. MSThesis, Oregon State College, Corvallis, Oregon.
MILLER, G. C., AND J. M. VAN HYNING. 1970. The commercialfishery for fresh-water crawfish, Pacifastacus leniusculus(Astacidae), in Oregon, 1893–1956. Research Reports ofthe Fish Commission of Oregon 2:77–89.
MUELLER, K. W. 2001. First record of the red swamp crayfish,Procambarus clarkii (Girard 1852) (Decapoda, Cambar-idae), from Washington State, U.S.A. Crustaceana 74:1003–1007.
MUELLER, K. W., AND L. R. BODENSTEINER. 2009. Shelteroccupancy by mixed-species pairs of native signalcrayfish and non-native red swamp crayfish held inenclosures. Journal of Freshwater Ecology 24:67–76.
NOAA (NATIONAL OCEANIC AND ATMOSPHERIC ADMINISTRATION).2006. Coastal Change Analysis Program (C-CAP)database. National Oceanic and Atmospheric Adminis-tration, Washington, DC. (Available from: http://www.csc.noaa.gov/crs/lca/ccap.html)
NYSTROM, P., P. STENROTH, N. HOLMQVIST, O. BERGLUND, P.LARSSON, AND W. GRANELI. 2006. Crayfish in lakes andstreams: individual and population responses to preda-tion, productivity and substratum availability. Fresh-water Biology 51:2096–2113.
OLDEN, J. D., D. A. JACKSON, AND P. R. PERES-NETO. 2001.Spatial isolation and fish communities in drainage lakes.Oecologia (Berlin) 127:572–585.
OLDEN, J. D., D. A. JACKSON, AND P. R. PERES-NETO. 2002.Predictive models of fish species distributions: acomment on proper validation and chance predictions.Transactions of the American Fisheries Society 131:329–336.
OLDEN, J. D., J. M. MCCARTHY, J. T. MAXTED, W. W. FETZER, AND
M. J. VANDER ZANDEN. 2006. The rapid spread of rustycrayfish (Orconectes rusticus) with observations on nativecrayfish declines in Wisconsin (U.S.A.) over the past130 years. Biological Invasions 8:1621–1628.
OLDEN, J. D., M. J. VANDER ZANDEN, AND P. T. J. JOHNSON. 2011.Assessing ecosystem vulnerability to invasive rustycrayfish (Orconectes rusticus). Ecological Applications21:2587–2599.
OLSEN, T. M., D. M. LODGE, G. M. CAPELLI, AND R. J. HOULIHAN.1991. Mechanisms of impact of an introduced crayfish(Orconectes rusticus) on littoral congeners, snails, andmacrophytes. Canadian Journal of Fisheries and Aquat-ic Sciences 48:1853–1861.
PAVOINE, J., J. VALLET, A. DUFOUR, S. GACHET, AND H. DANIEL.2009. On the challenge of treating various types ofvariables: application for improving the measurementof functional diversity. Oikos 118:391–402.
PINTOR, L. M., A. SIH, AND J. L. KERBY. 2009. Behavioralcorrelations provide a mechanism for explaining highinvader densities and increased impacts on native prey.Ecology 90:581–587.
POE, T. P., H. C. HANSEL, S. VIGG, D. E. PALMER, AND L. A.PRENDERGAST. 1991. Feeding of predaceous fishes on out-migrating juvenile salmonids in the John Day Reservoir,Columbia River. Transactions of the American FisheriesSociety 120:405–420.
PUTH, L. M., AND T. F. H. ALLEN. 2004. Potential corridors forthe rusty crayfish, Orconectes rusticus, in northernWisconsin (USA) lakes: lessons for exotic invasions.Landscape Ecology 20:567–577.
RODHE, H. 1989. Acidification in a global perspective. Ambio18:155–160.
ROTH, B. M., J. C. TETZLAFF, M. L. ALEXANDER, AND J. F.KITCHELL. 2007. Reciprocal relationships between exoticrusty crayfish, macrophytes, and Lepomis species innorthern Wisconsin lakes. Ecosystems 10:74–85.
ROYLE, J. A., J. D. NICHOLS, AND M. KERY. 2005. Modellingoccurrence and abundance of species when detection isimperfect. Oikos 110:353–359.
SHURIN, J. B., J. E. HAVEL, M. A. LEIBOLD, AND B. PINEL-ALLOUL.2000. Local and regional zooplankton species richness:a scale-independent test for saturation. Ecology 81:3062–3073.
SOMERS, J. M., AND D. P. M. STECHEY. 1986. Variable trapabilityof crayfish associated with bait type, water temperatureand lunar phase. American Midland Naturalist 116:36–44.
STRECKER, A. L., P. M. CAMPBELL, AND J. D. OLDEN. 2011. Theaquarium trade as an invasion pathway in the PacificNorthwest. Fisheries 36:74–85.
TAYLOR, C. A., G. A. SCHUSTER, J. E. COOPER, R. J. DISTEFANO,A. G. EVERSOLE, P. HAMR, H. H. HOBBS, H. W. ROBISON, C.E. SKELTON, AND R. E. THOMA. 2007. A reassessment of theconservation status of crayfishes of the United Statesand Canada after 10+ years of increased awareness.Fisheries 32(8):372–389.
THORSON, R. M. 1980. Ice-sheet glaciation of the Pugetlowland, Washington, during the Vashon Stade (LatePleistocene). Quaternary Research 13:303–321.
USGS (UNITED STATES GEOLOGICAL SURVEY). 2010. The NationalHydrography Dataset v2.0. United States GeologicalSurvey, Reston, Virginia. (Available from: nhd.usgs.gov)
USIO, N., H. NAKAJIMA, R. KAMIYAMA, I. WAKANA, S. HIRUTA,AND N. TAKAMURA. 2006. Predicting the distribution ofinvasive crayfish (Pacifastacus leniusculus) in a KusiroMoor marsh (Japan) using classification and regressiontrees. Ecological Research 21:271–277.
USIO, N., K. NAKATA, T. KAWAI, AND S. KITANO. 2007.Distribution and control status of the invasive signalcrayfish (Pacifastacus leniusculus) in Japan. JapaneseJournal of Limnology 68:471–482.
VANDER ZANDEN, M. J., AND J. D. OLDEN. 2008. A managementframework for preventing the secondary spread ofaquatic invasive species. Canadian Journal of Fisheriesand Aquatic Sciences 65:1512–1522.
WDFW (WASHINGTON DEPARTMENT OF FISH AND WILDLIFE). 2005.Warmwater fishes of Washington. Report FM93-9.Washington Department of Fish and Wildlife, Olympia,Washington.
106 E. R. LARSON AND J. D. OLDEN [Volume 32

WDNR (WASHINGTON DEPARTMENT OF NATURAL RESOURCES). 2010.Washington surface geology GIS Data 3.0. WashingtonDepartment of Natural Resources, Olympia, Washington.(Available from: www.dnr.wa.gov/ResearchScience/Pages/PubData.aspx)
WHITTAKER, R. H., AND C. W. FAIRBANKS. 1958. A study ofplankton copepod communities in the Columbia Basin,southeastern Washington. Ecology 39:46–65.
WOOSTER, D., J. L. SNYDER, AND A. MADSEN. 2012. Environ-mental correlates of signal crayfish, Pacifastacus lenius-culus (Dana, 1852), density and size at two spatial scales
in its native range. Journal of Crustacean Biology 32:741–752.
ZIMMERMAN, J. K. M., AND R. T. PALO. 2011. Reliability of catchper unit effort (CPUE) for evaluation of reintroductionprograms – a comparison of the mark-recapture methodwith standardized trapping. Knowledge and Manage-ment of Aquatic Ecosystems 401:07. doi: http://dx.doi.org/10.1051/kmae/2011016
Received: 2 April 2012Accepted: 2 November 2012
2013] CRAYFISH IN PACIFIC NORTHWEST LAKES 107





























