Embed Size (px)
Citation preview

Cellular Immunology 243 (2006) 75–82
www.elsevier.com/locate/ycimm
CpG-Induced IFN� expands TLR4-speciWc IL-18 responses in vivo
Sameer Gupta a, Meetha P. Gould b, Jennifer DeVecchio c, David H. Canaday d, JeVery J. Auletta e, Frederick P. Heinzel b,c,¤
a Pediatric Pharmacology and Critical Care, Case Western Reserve University School of Medicine, Cleveland, OH 44106, USAb The Center for Global Health and Diseases, Case Western Reserve University School of Medicine, Cleveland, OH 44106, USA
c Medical Research Service, Louis Stokes VA Medical Center, Cleveland, OH 44106, USAd The Department of Medicine, Case Western Reserve University School of Medicine, Cleveland, OH 44106, USA
e Pediatric Hematology/Oncology and Infectious Diseases, Case Western Reserve University School of Medicine, Cleveland, OH 44106, USA
Received 5 June 2006; accepted 27 December 2006Available online 9 February 2007
Abstract
Serum IL-18 responses to LPS increase after pretreatment with CpG-containing DNA. Compared to saline-pretreated controls, micepretreated with CpG for two days produced 20-fold more serum IL-18 when challenged with lipopolysaccharide (LPS). In contrast,IFN�-deWciency or anti-IFN� pretreatment reduced CpG-expanded IL-18 responses to LPS by 67 and 83%, respectively. Mice pretreatedwith either IFN� or CpG comparably increased LPS-inducible serum IL-18 responses. LPS, compared to challenge with other TLR ago-nists, was best able to trigger high serum IL-18 levels in CpG-pretreated mice and this response was TLR4-dependent. CpG, compared topretreatment with other TLR agonists, optimally expanded LPS-induced IL-18 responses that correlated with higher levels of circulatingIFN� levels prior to LPS challenge. High-level serum IL-18 responses were caspase-1-dependent and P2X7 receptor-independent. Weconclude that CpG promotes high-level IL-18 synthesis by an IFN�-dependent and IFN�-suYcient mechanism in vivo that is optimallytriggered by LPS.© 2007 Elsevier Inc. All rights reserved.
Keywords: Interleukin-18; Interferon, type II; Toll-like receptor 9; Receptors; Purinergic P2; Mice
1. Introduction
IL-18. was initially identiWed as an IFN�-inducing fac-tor in serum that was produced in LPS-injected micetreated with heat-killed Propionibacterium acnes [1]. IL-18is now known to mediate critical antimicrobial responsesin animal models of bacterial, fungal, viral and parasiticinfections [2–5]. IL-18 supports both innate and adaptiveimmune production of IFN�, while separately promotingIFN�-independent eVects on polymorphonuclear cellfunction. IL-18 synthesis is triggered by innate immunerecognition of microbial products, such as lipopolysac-charide, which activates IL-18 release from murine mac-
* Corresponding author. Present address: Infectious Diseases, 111-J, JamesA Haley VAMC, 13000 Bruce B Downs Blvd, Tampa, FL 33612, USA.
E-mail address: [email protected] (F.P. Heinzel).
0008-8749/$ - see front matter © 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.cellimm.2006.12.004
rophages or KupVer cells through TLR4-dependent andMyd88-independent signaling pathways [6]. Cytoplasmicstores of pro-IL-18 peptide are proteolytically modiWedby caspase-1 to generate bioactive cytokine exported fromthe cell by a nonclassical, Golgi-independent transportpathway. Signals other than LPS also trigger IL-18 syn-thesis. For example, Chlamydia pneumoniae-induced IL-18 is also produced by Myd88-dependent and TLR4/TLR2-independent pathways [7].
The magnitude of a toll receptor-driven IL-18 responseis modulated by several factors. Triggering of the P2X7receptor by soluble ATP greatly ampliWes the amounts ofIL-18 and IL-1� released by LPS-exposed macrophages[8,9]. IL-18 synthetic capacity also progressively increasesduring repetitive or continuous inXammatory challengeswith LPS [2], after pretreatment with P. acnes [1] or aftertreatment with recombinant IL-12 [10]. Previously, we

76 S. Gupta et al. / Cellular Immunology 243 (2006) 75–82
showed that pretreatment of mice with CpG oligonucleo-tide 1826 for at least two days also dramatically increasedIL-18 serum responses in vivo. Levels of IL-18 elicited byLPS in CpG-pretreated mice were 10- to 20-fold greaterthan in saline-pretreated mice [11]. These expanded serumIL-18 responses subsequently mediated parallel increases inthe LPS-induced IFN� response.
Since CpG stimulates prolonged release of IFN�in vivo prior to LPS challenge [11,12] we hypothesizedthat CpG-induced IFN� contributed to the expansion ofLPS-inducible IL-18 production in vivo. Previously,IFN�-dependent eVects on IL-18 synthesis had beenobserved in P. acnes-pretreated mice [13]. We show thathigh level IL-18 production in CpG-pretreated mice notonly requires prior IFN� activity, but that IFN� alone issuYcient to expand IL-18 production in vivo. We alsoshow that high-level IL-18 responses in CpG-pretreatedmice are optimally triggered by TLR4-dependent stimuli,require caspase-1 expression, but are independent ofP2X7 receptor expression. These Wndings demonstratethat IFN� produced during inXammation dynamicallyregulates the extent of IL-18 production. The restrictiverequirements for CpG and LPS as modulatory and stimu-lating signals, respectively, for high-level IL-18 synthesisfurther suggest that sequential TLR9 and TLR4 activa-tion may uniquely program innate cytokine responses tobacterial challenge.
2. Materials and methods
2.1. Reagents
Salmonella enteriditis lipopolysaccharide was purchasedfrom Sigma Chemical (St Louis, MO). PhosphorothioatedCpG 1826 (TCCATGACGTTCCTGACGTT 5�–3�) waspurchased from Oligos Etc (Wilsonville, OR) and provenfree of LPS contamination by limulus lysate assay (E-Tox-ate, Sigma Chemical Co). Recombinant murine IFN� wasobtained from Peprotech Inc. (Rocky Hill, NJ) and neutral-izing anti-mouse IFN� (XMG1.2; rat IgG1) and irrelevantcontrol rat IgG1 monoclonal antibodies were purchasedfrom Bio Express (West Lebanon, NH). The bacterial lipo-peptide, Pam3CysSerLys4 was purchased from EMCMicrocollections (Tuebingen, Germany). The TLR7 ago-nist, R848, was purchased from PharmaTech (Shanghai,China) and poly(IC) was obtained from Amersham Corp(Piscataway, NJ).
2.2. Mice
Four to six-week old female C3H/HeN and C3H/HeJmice were purchased from Charles River (Lexington, MA).C57BL/6 and C57BL/6 IFN�¡/¡ mice were purchased fromJackson Laboratories (Bar Harbor, ME). C57BL/6 P2X7receptor knockout (P2X7¡/¡) mice [14] were provided byDr. Laurent Audoly (PWzer, Groton, CT), backcrossed atleast 6 generations onto C57BL/6 and maintained at Taconic
until shipment. Caspase-1 deWcient C57BL/6 mice (ICE KO)were obtained from Dr. Richard Flavell (Yale University,New Haven, CT). All mice were housed in the Case WesternReserve University animal facilities under speciWc pathogen-free conditions. For pretreatment with toll agonists, micewere injected i.p. daily for two days with 0.2 ml of PBS aloneor PBS containing 50�g of CpG OD 1826, 50�g ofPam3CysSerLys4 or 20�g of R848. At 72 h after the Wrst con-ditioning dose, mice were challenged by i.p. injection with300�g of LPS (LD90). Because CpG-preconditioned micedeveloped signs of severe toxicity at 6 h after LPS challenge,mice were all euthanized before 6 h post-challenge. Whereindicated, mice were treated with 20�g of recombinantmouse IFN� daily for two days. Selected mice were injectedwith 0.5 mg of XMG1.2 anti-IFN� IgG i.p. 4 h or irrelevantrat IgG1 prior to initial pretreatment with CpG. All proce-dures were approved by the CWRU IACUC.
2.3. ELISA
Cytokine concentrations in serum and conditioned culturemedia were determined for IFN-� using antibody pairs fromBD-Pharmingen, as described [11] Serum IL-18 was mea-sured using an ELISA from Medical and Biological Labora-tories International (Woburn, MA) that detects maturemouse IL-18 with minimal cross-reactivity to precursorforms of IL-18. Mouse IFN� and IL-1� were assayed usingELISA reagents from Pharmingen-BD (San Diego, CA).
2.4. Statistics
Tests for signiWcance of diVerence were made using theStudent parametric t-test.
3. Results
3.1. CpG-pretreatment expands LPS-inducible IL-18 synthesis by an IFN�-dependent mechanism
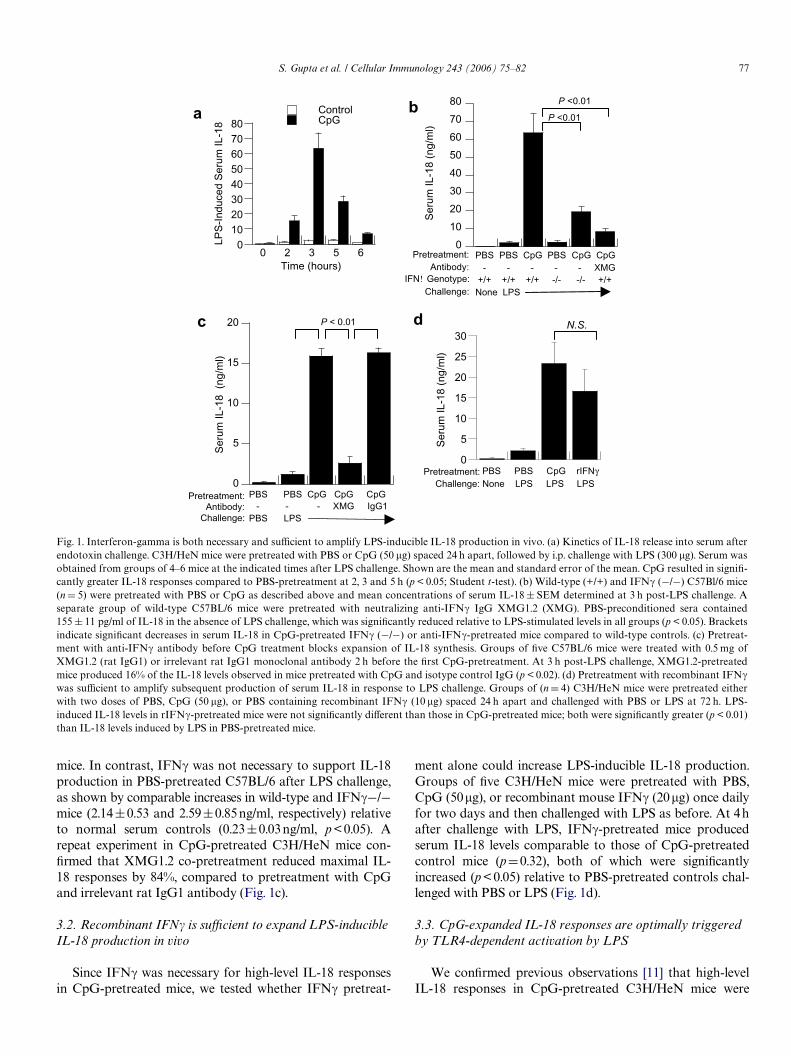
We conWrmed that pretreatment of C3H/HeN mice with50�g of CpG for two days resulted in 10- to 20-fold increasesin LPS-induced serum IL-18 [11], relative to saline-pretreatedmice. These responses peaked between 3 and 5 h after LPSchallenge in both groups (Fig. 1a). Because we previouslyobserved circulating IFN� after CpG-pretreatment, wetested whether IFN� exposure prior to LPS challenge regu-lated IL-18 synthetic capacity (Fig. 1b). Compared to PBS-pretreated controls, CpG-pretreated mice produced 10-foldmore LPS-inducible serum IL-18. However, CpG-pretreatedIFN�¡/¡ mice generated IL-18 concentrations that wereonly one-third of that produced by pretreated IFN�-compe-tent mice (signiWcant at p <0.05). Similarly, combined pre-treatment of wild-type C57Bl/6 mice with anti-IFN�monoclonal antibody 4 h prior to the Wrst CpG- pretreat-ment also reduced the LPS-induced serum IL-18 response to17% of that of CpG-pretreated controls (p < 0.05), althoughthese remained greater than those of PBS-pretreated control
S. Gupta et al. / Cellular Immunology 243 (2006) 75–82 77
mice. In contrast, IFN� was not necessary to support IL-18production in PBS-pretreated C57BL/6 after LPS challenge,as shown by comparable increases in wild-type and IFN�¡/¡mice (2.14§0.53 and 2.59§0.85 ng/ml, respectively) relativeto normal serum controls (0.23§0.03 ng/ml, p <0.05). Arepeat experiment in CpG-pretreated C3H/HeN mice con-Wrmed that XMG1.2 co-pretreatment reduced maximal IL-18 responses by 84%, compared to pretreatment with CpGand irrelevant rat IgG1 antibody (Fig. 1c).
3.2. Recombinant IFN� is suYcient to expand LPS-inducible IL-18 production in vivo
Since IFN� was necessary for high-level IL-18 responsesin CpG-pretreated mice, we tested whether IFN� pretreat-
ment alone could increase LPS-inducible IL-18 production.Groups of Wve C3H/HeN mice were pretreated with PBS,CpG (50�g), or recombinant mouse IFN� (20�g) once dailyfor two days and then challenged with LPS as before. At 4hafter challenge with LPS, IFN�-pretreated mice producedserum IL-18 levels comparable to those of CpG-pretreatedcontrol mice (pD0.32), both of which were signiWcantlyincreased (p < 0.05) relative to PBS-pretreated controls chal-lenged with PBS or LPS (Fig. 1d).
3.3. CpG-expanded IL-18 responses are optimally triggered by TLR4-dependent activation by LPS
We conWrmed previous observations [11] that high-levelIL-18 responses in CpG-pretreated C3H/HeN mice were
Fig. 1. Interferon-gamma is both necessary and suYcient to amplify LPS-inducible IL-18 production in vivo. (a) Kinetics of IL-18 release into serum afterendotoxin challenge. C3H/HeN mice were pretreated with PBS or CpG (50 �g) spaced 24 h apart, followed by i.p. challenge with LPS (300 �g). Serum wasobtained from groups of 4–6 mice at the indicated times after LPS challenge. Shown are the mean and standard error of the mean. CpG resulted in signiW-cantly greater IL-18 responses compared to PBS-pretreatment at 2, 3 and 5 h (p < 0.05; Student t-test). (b) Wild-type (+/+) and IFN� (¡/¡) C57Bl/6 mice(nD 5) were pretreated with PBS or CpG as described above and mean concentrations of serum IL-18§ SEM determined at 3 h post-LPS challenge. Aseparate group of wild-type C57BL/6 mice were pretreated with neutralizing anti-IFN� IgG XMG1.2 (XMG). PBS-preconditioned sera contained155 § 11 pg/ml of IL-18 in the absence of LPS challenge, which was signiWcantly reduced relative to LPS-stimulated levels in all groups (p < 0.05). Bracketsindicate signiWcant decreases in serum IL-18 in CpG-pretreated IFN� (¡/¡) or anti-IFN�-pretreated mice compared to wild-type controls. (c) Pretreat-ment with anti-IFN� antibody before CpG treatment blocks expansion of IL-18 synthesis. Groups of Wve C57BL/6 mice were treated with 0.5 mg ofXMG1.2 (rat IgG1) or irrelevant rat IgG1 monoclonal antibody 2 h before the Wrst CpG-pretreatment. At 3 h post-LPS challenge, XMG1.2-pretreatedmice produced 16% of the IL-18 levels observed in mice pretreated with CpG and isotype control IgG (p < 0.02). (d) Pretreatment with recombinant IFN�was suYcient to amplify subsequent production of serum IL-18 in response to LPS challenge. Groups of (nD 4) C3H/HeN mice were pretreated eitherwith two doses of PBS, CpG (50 �g), or PBS containing recombinant IFN� (10 �g) spaced 24 h apart and challenged with PBS or LPS at 72 h. LPS-induced IL-18 levels in rIFN�-pretreated mice were not signiWcantly diVerent than those in CpG-pretreated mice; both were signiWcantly greater (p < 0.01)than IL-18 levels induced by LPS in PBS-pretreated mice.
01020304050607080
Pretreatment:Antibody:
IFN! Genotype:Challenge:
Seru
mIL
-18
(ng/
ml)
b P <0.01
P <0.01
Pretreatment:Challenge:
PBS PBS CpG rIFNγNone LPS LPS LPS
Seru
mIL
-18
(ng/
ml)
0
5
10
15
20
25
30
0
5
10
15
20
PBS PBS CpG PBS CpG CpG- - - - - XMG
+/+ +/+ +/+ -/- -/- +/+None LPS
N.S.
Time (hours)
LPS-
Indu
ced
Seru
mIL
-18
a
01020304050607080
0 2 3 5 6
ControlCpG
d
Pretreatment:Antibody:
Challenge:
PBS PBS CpG CpG CpG- - - XMG IgG1
PBS LPS
P < 0.01c
Seru
mIL
-18
(ng/
ml)
78 S. Gupta et al. / Cellular Immunology 243 (2006) 75–82
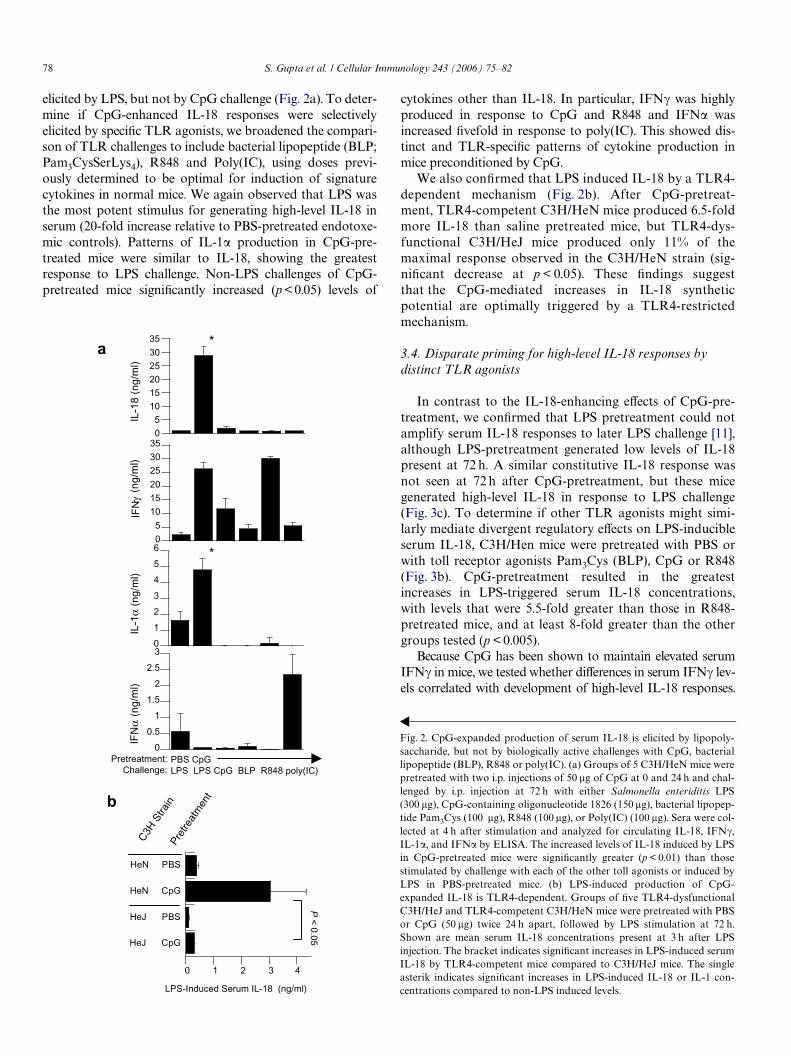
elicited by LPS, but not by CpG challenge (Fig. 2a). To deter-mine if CpG-enhanced IL-18 responses were selectivelyelicited by speciWc TLR agonists, we broadened the compari-son of TLR challenges to include bacterial lipopeptide (BLP;Pam3CysSerLys4), R848 and Poly(IC), using doses previ-ously determined to be optimal for induction of signaturecytokines in normal mice. We again observed that LPS wasthe most potent stimulus for generating high-level IL-18 inserum (20-fold increase relative to PBS-pretreated endotoxe-mic controls). Patterns of IL-1� production in CpG-pre-treated mice were similar to IL-18, showing the greatestresponse to LPS challenge. Non-LPS challenges of CpG-pretreated mice signiWcantly increased (p < 0.05) levels of
0 1 2 3 4
HeN PBS
HeN CpG
HeJ PBS
HeJ CpG
Pret
reat
men
t
P<
0.05
LPS-Induced Serum IL-18 (ng/ml)
05
101520253035
Pretreatment:Challenge:
PBS CpGLPS LPS CpG BLP R848 poly(IC)
IL-1
α(n
g/m
l)IF
NIF
Nα
(ng/
ml)
(ng/
ml)
IL-1
8(n
g/m
l)
*
*
C3H
Stra
in
a
b
05
101520
253035
0
1
2
3
4
5
6
0
0.5
1
1.5
2
2.5
3
γ
cytokines other than IL-18. In particular, IFN� was highlyproduced in response to CpG and R848 and IFN� wasincreased Wvefold in response to poly(IC). This showed dis-tinct and TLR-speciWc patterns of cytokine production inmice preconditioned by CpG.
We also conWrmed that LPS induced IL-18 by a TLR4-dependent mechanism (Fig. 2b). After CpG-pretreat-ment, TLR4-competent C3H/HeN mice produced 6.5-foldmore IL-18 than saline pretreated mice, but TLR4-dys-functional C3H/HeJ mice produced only 11% of themaximal response observed in the C3H/HeN strain (sig-niWcant decrease at p < 0.05). These Wndings suggestthat the CpG-mediated increases in IL-18 syntheticpotential are optimally triggered by a TLR4-restrictedmechanism.
3.4. Disparate priming for high-level IL-18 responses by distinct TLR agonists
In contrast to the IL-18-enhancing eVects of CpG-pre-treatment, we conWrmed that LPS pretreatment could notamplify serum IL-18 responses to later LPS challenge [11],although LPS-pretreatment generated low levels of IL-18present at 72 h. A similar constitutive IL-18 response wasnot seen at 72 h after CpG-pretreatment, but these micegenerated high-level IL-18 in response to LPS challenge(Fig. 3c). To determine if other TLR agonists might simi-larly mediate divergent regulatory eVects on LPS-inducibleserum IL-18, C3H/Hen mice were pretreated with PBS orwith toll receptor agonists Pam3Cys (BLP), CpG or R848(Fig. 3b). CpG-pretreatment resulted in the greatestincreases in LPS-triggered serum IL-18 concentrations,with levels that were 5.5-fold greater than those in R848-pretreated mice, and at least 8-fold greater than the othergroups tested (p < 0.005).
Because CpG has been shown to maintain elevated serumIFN� in mice, we tested whether diVerences in serum IFN� lev-els correlated with development of high-level IL-18 responses.
Fig. 2. CpG-expanded production of serum IL-18 is elicited by lipopoly-saccharide, but not by biologically active challenges with CpG, bacteriallipopeptide (BLP), R848 or poly(IC). (a) Groups of 5 C3H/HeN mice werepretreated with two i.p. injections of 50 �g of CpG at 0 and 24 h and chal-lenged by i.p. injection at 72 h with either Salmonella enteriditis LPS(300 �g), CpG-containing oligonucleotide 1826 (150 �g), bacterial lipopep-tide Pam3Cys (100 �g), R848 (100 �g), or Poly(IC) (100 �g). Sera were col-lected at 4 h after stimulation and analyzed for circulating IL-18, IFN�,IL-1�, and IFN� by ELISA. The increased levels of IL-18 induced by LPSin CpG-pretreated mice were signiWcantly greater (p < 0.01) than thosestimulated by challenge with each of the other toll agonists or induced byLPS in PBS-pretreated mice. (b) LPS-induced production of CpG-expanded IL-18 is TLR4-dependent. Groups of Wve TLR4-dysfunctionalC3H/HeJ and TLR4-competent C3H/HeN mice were pretreated with PBSor CpG (50 �g) twice 24 h apart, followed by LPS stimulation at 72 h.Shown are mean serum IL-18 concentrations present at 3 h after LPSinjection. The bracket indicates signiWcant increases in LPS-induced serumIL-18 by TLR4-competent mice compared to C3H/HeJ mice. The singleasterik indicates signiWcant increases in LPS-induced IL-18 or IL-1 con-centrations compared to non-LPS induced levels.
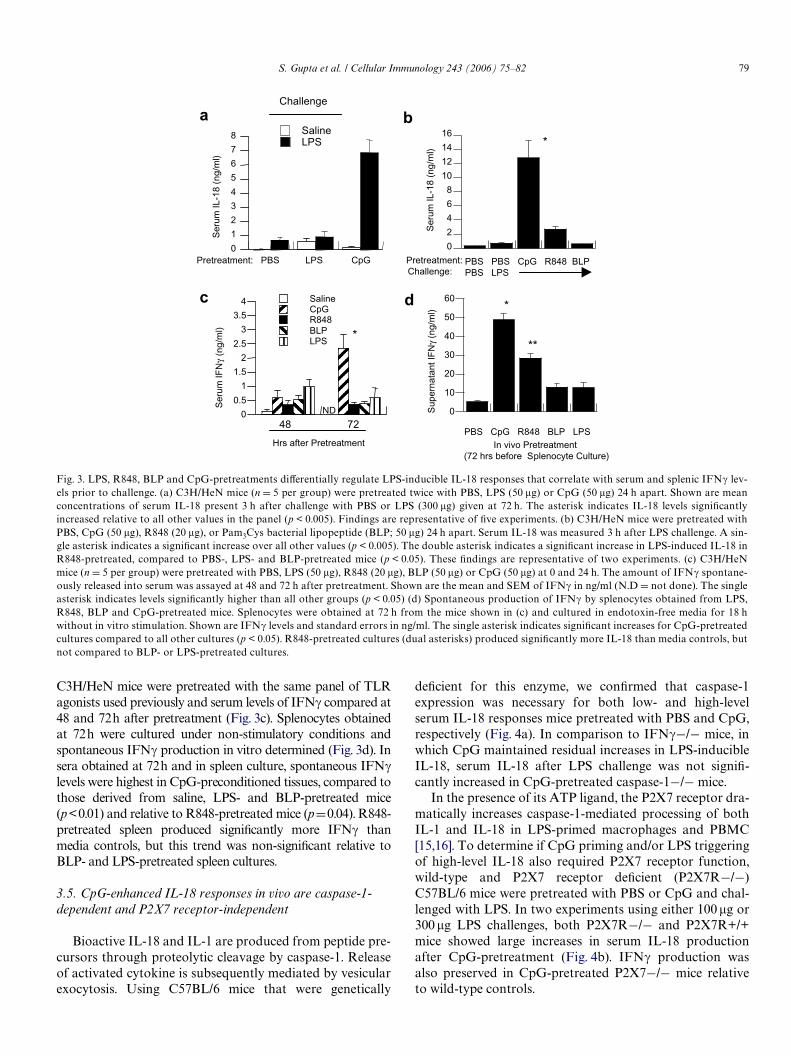
S. Gupta et al. / Cellular Immunology 243 (2006) 75–82 79
C3H/HeN mice were pretreated with the same panel of TLRagonists used previously and serum levels of IFN� compared at48 and 72h after pretreatment (Fig. 3c). Splenocytes obtainedat 72h were cultured under non-stimulatory conditions andspontaneous IFN� production in vitro determined (Fig. 3d). Insera obtained at 72h and in spleen culture, spontaneous IFN�levels were highest in CpG-preconditioned tissues, compared tothose derived from saline, LPS- and BLP-pretreated mice(p<0.01) and relative to R848-pretreated mice (pD0.04). R848-pretreated spleen produced signiWcantly more IFN� thanmedia controls, but this trend was non-signiWcant relative toBLP- and LPS-pretreated spleen cultures.
3.5. CpG-enhanced IL-18 responses in vivo are caspase-1-dependent and P2X7 receptor-independent
Bioactive IL-18 and IL-1 are produced from peptide pre-cursors through proteolytic cleavage by caspase-1. Releaseof activated cytokine is subsequently mediated by vesicularexocytosis. Using C57BL/6 mice that were genetically
deWcient for this enzyme, we conWrmed that caspase-1expression was necessary for both low- and high-levelserum IL-18 responses mice pretreated with PBS and CpG,respectively (Fig. 4a). In comparison to IFN�¡/¡ mice, inwhich CpG maintained residual increases in LPS-inducibleIL-18, serum IL-18 after LPS challenge was not signiW-cantly increased in CpG-pretreated caspase-1¡/¡ mice.
In the presence of its ATP ligand, the P2X7 receptor dra-matically increases caspase-1-mediated processing of bothIL-1 and IL-18 in LPS-primed macrophages and PBMC[15,16]. To determine if CpG priming and/or LPS triggeringof high-level IL-18 also required P2X7 receptor function,wild-type and P2X7 receptor deWcient (P2X7R¡/¡)C57BL/6 mice were pretreated with PBS or CpG and chal-lenged with LPS. In two experiments using either 100 �g or300 �g LPS challenges, both P2X7R¡/¡ and P2X7R+/+mice showed large increases in serum IL-18 productionafter CpG-pretreatment (Fig. 4b). IFN� production wasalso preserved in CpG-pretreated P2X7¡/¡ mice relativeto wild-type controls.
Fig. 3. LPS, R848, BLP and CpG-pretreatments diVerentially regulate LPS-inducible IL-18 responses that correlate with serum and splenic IFN� lev-els prior to challenge. (a) C3H/HeN mice (n D 5 per group) were pretreated twice with PBS, LPS (50 �g) or CpG (50 �g) 24 h apart. Shown are meanconcentrations of serum IL-18 present 3 h after challenge with PBS or LPS (300 �g) given at 72 h. The asterisk indicates IL-18 levels signiWcantlyincreased relative to all other values in the panel (p < 0.005). Findings are representative of Wve experiments. (b) C3H/HeN mice were pretreated withPBS, CpG (50 �g), R848 (20 �g), or Pam3Cys bacterial lipopeptide (BLP; 50 �g) 24 h apart. Serum IL-18 was measured 3 h after LPS challenge. A sin-gle asterisk indicates a signiWcant increase over all other values (p < 0.005). The double asterisk indicates a signiWcant increase in LPS-induced IL-18 inR848-pretreated, compared to PBS-, LPS- and BLP-pretreated mice (p < 0.05). These Wndings are representative of two experiments. (c) C3H/HeNmice (n D 5 per group) were pretreated with PBS, LPS (50 �g), R848 (20 �g), BLP (50 �g) or CpG (50 �g) at 0 and 24 h. The amount of IFN� spontane-ously released into serum was assayed at 48 and 72 h after pretreatment. Shown are the mean and SEM of IFN� in ng/ml (N.D D not done). The singleasterisk indicates levels signiWcantly higher than all other groups (p < 0.05) (d) Spontaneous production of IFN� by splenocytes obtained from LPS,R848, BLP and CpG-pretreated mice. Splenocytes were obtained at 72 h from the mice shown in (c) and cultured in endotoxin-free media for 18 hwithout in vitro stimulation. Shown are IFN� levels and standard errors in ng/ml. The single asterisk indicates signiWcant increases for CpG-pretreatedcultures compared to all other cultures (p < 0.05). R848-pretreated cultures (dual asterisks) produced signiWcantly more IL-18 than media controls, butnot compared to BLP- or LPS-pretreated cultures.
Ser
umIF
Nγ
(ng/
ml)
Hrs after PretreatmentPBS CpG R848 BLP LPS
Sup
erna
tant
IFN
γ(n
g/m
l)In vivo Pretreatment
(72 hrs before Splenocyte Culture)
a b
c d *
**
*
0
2
4
6
8
10
12
14
16
0
10
20
30
40
50
60
Ser
umIL
-18
(ng/
ml)
48 72S
erum
IL-1
8(n
g/m
l)
Challenge
PBS PBS CpG R848 BLPPBS LPS
Pretreatment:Challenge:
Pretreatment: PBS LPS CpG
ND
0
1
2
3
4
5
6
7
8 SalineLPS
0
0.5
1
1.5
2
2.5
3
3.5
4 SalineCpGR848BLPLPS
*
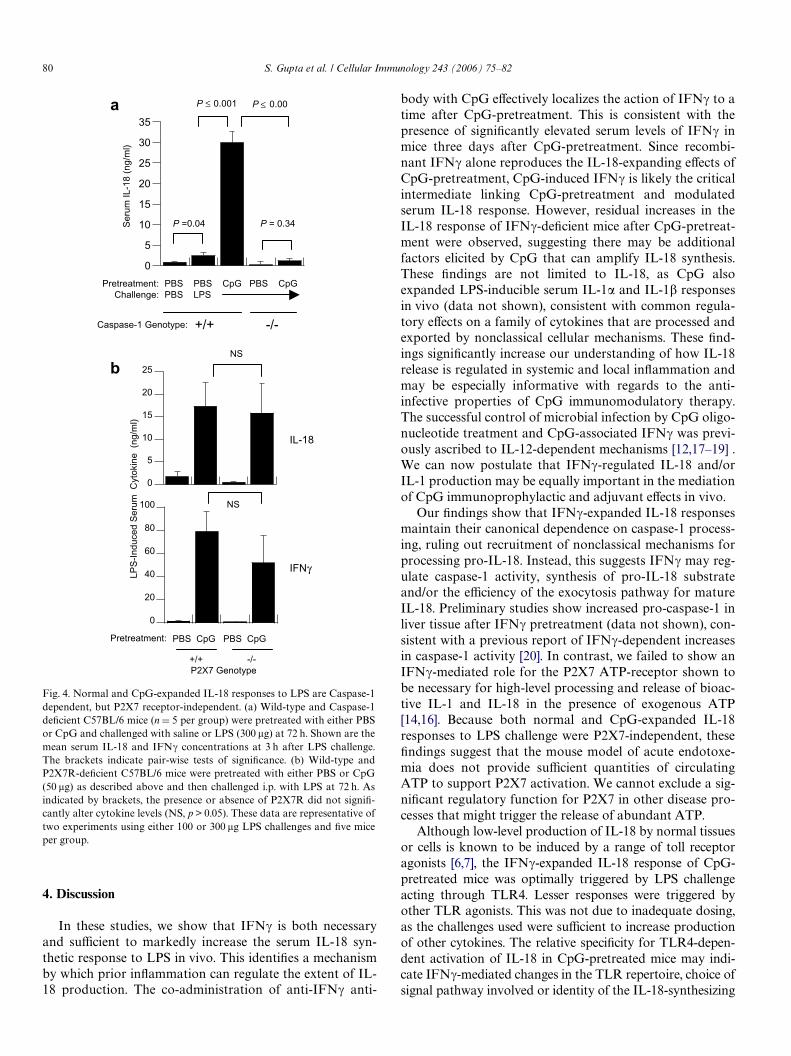
80 S. Gupta et al. / Cellular Immunology 243 (2006) 75–82
4. Discussion
In these studies, we show that IFN� is both necessaryand suYcient to markedly increase the serum IL-18 syn-thetic response to LPS in vivo. This identiWes a mechanismby which prior inXammation can regulate the extent of IL-18 production. The co-administration of anti-IFN� anti-
Fig. 4. Normal and CpG-expanded IL-18 responses to LPS are Caspase-1dependent, but P2X7 receptor-independent. (a) Wild-type and Caspase-1deWcient C57BL/6 mice (n D 5 per group) were pretreated with either PBSor CpG and challenged with saline or LPS (300 �g) at 72 h. Shown are themean serum IL-18 and IFN� concentrations at 3 h after LPS challenge.The brackets indicate pair-wise tests of signiWcance. (b) Wild-type andP2X7R-deWcient C57BL/6 mice were pretreated with either PBS or CpG(50 �g) as described above and then challenged i.p. with LPS at 72 h. Asindicated by brackets, the presence or absence of P2X7R did not signiW-cantly alter cytokine levels (NS, p > 0.05). These data are representative oftwo experiments using either 100 or 300 �g LPS challenges and Wve miceper group.
b
0
20
40
60
80
100
0
5
10
15
20
25
Pretreatment:
LPS-
Indu
ced
Seru
mC
ytok
ine
(ng/
ml)
NS
0
5
10
15
20
25
30
35
Pretreatment:Challenge:
PBS PBS CpG PBS CpGPBS LPS
Seru
mIL
-18
(ng/
ml)
P =0.04
P ≤ ≤0.001 P 0.00
P = 0.34
+/+ -/-Caspase-1 Genotype:
a
PBS CpG PBS CpG
IFNγ
IL-18
+/+ -/-P2X7 Genotype
NS
body with CpG eVectively localizes the action of IFN� to atime after CpG-pretreatment. This is consistent with thepresence of signiWcantly elevated serum levels of IFN� inmice three days after CpG-pretreatment. Since recombi-nant IFN� alone reproduces the IL-18-expanding eVects ofCpG-pretreatment, CpG-induced IFN� is likely the criticalintermediate linking CpG-pretreatment and modulatedserum IL-18 response. However, residual increases in theIL-18 response of IFN�-deWcient mice after CpG-pretreat-ment were observed, suggesting there may be additionalfactors elicited by CpG that can amplify IL-18 synthesis.These Wndings are not limited to IL-18, as CpG alsoexpanded LPS-inducible serum IL-1� and IL-1� responsesin vivo (data not shown), consistent with common regula-tory eVects on a family of cytokines that are processed andexported by nonclassical cellular mechanisms. These Wnd-ings signiWcantly increase our understanding of how IL-18release is regulated in systemic and local inXammation andmay be especially informative with regards to the anti-infective properties of CpG immunomodulatory therapy.The successful control of microbial infection by CpG oligo-nucleotide treatment and CpG-associated IFN� was previ-ously ascribed to IL-12-dependent mechanisms [12,17–19] .We can now postulate that IFN�-regulated IL-18 and/orIL-1 production may be equally important in the mediationof CpG immunoprophylactic and adjuvant eVects in vivo.
Our Wndings show that IFN�-expanded IL-18 responsesmaintain their canonical dependence on caspase-1 process-ing, ruling out recruitment of nonclassical mechanisms forprocessing pro-IL-18. Instead, this suggests IFN� may reg-ulate caspase-1 activity, synthesis of pro-IL-18 substrateand/or the eYciency of the exocytosis pathway for matureIL-18. Preliminary studies show increased pro-caspase-1 inliver tissue after IFN� pretreatment (data not shown), con-sistent with a previous report of IFN�-dependent increasesin caspase-1 activity [20]. In contrast, we failed to show anIFN�-mediated role for the P2X7 ATP-receptor shown tobe necessary for high-level processing and release of bioac-tive IL-1 and IL-18 in the presence of exogenous ATP[14,16]. Because both normal and CpG-expanded IL-18responses to LPS challenge were P2X7-independent, theseWndings suggest that the mouse model of acute endotoxe-mia does not provide suYcient quantities of circulatingATP to support P2X7 activation. We cannot exclude a sig-niWcant regulatory function for P2X7 in other disease pro-cesses that might trigger the release of abundant ATP.
Although low-level production of IL-18 by normal tissuesor cells is known to be induced by a range of toll receptoragonists [6,7], the IFN�-expanded IL-18 response of CpG-pretreated mice was optimally triggered by LPS challengeacting through TLR4. Lesser responses were triggered byother TLR agonists. This was not due to inadequate dosing,as the challenges used were suYcient to increase productionof other cytokines. The relative speciWcity for TLR4-depen-dent activation of IL-18 in CpG-pretreated mice may indi-cate IFN�-mediated changes in the TLR repertoire, choice ofsignal pathway involved or identity of the IL-18-synthesizing

S. Gupta et al. / Cellular Immunology 243 (2006) 75–82 81
cell. Future studies will need to distinguish between thesemechanistic possibilities.
We also observed in these initial studies that precondi-tioning with CpG, compared to other TLR agonists, wasoptimal for expanding IL-18 responses in vivo. Usingin vivo doses of toll agonist that acutely stimulated cyto-kine production in normal mice, Pam3Cys, LPS and R848promoted LPS-triggered IL-18 responses that were signiW-cantly less than those generated in CpG-pretreated mice.Further escalation of LPS and Pam3Cys dosing was limitedby toxicity. Consistent with our Wnding that IFN� alone issuYcient to regulate the amplitude of the IL-18 response,CpG promoted signiWcantly greater levels of splenocyte orserum IFN� in mice at 72 h after pretreatment. R848, whichpromoted lesser increases of LPS-inducible IL-18 also gen-erated increased levels of IFN� in splenocyte culture. Theseobservations suggest that other inXammatory responsesproductive of IFN�, such as seen after repeated dosing withIL-12 [10], may similarly expand IL-18 capacity in vivo.
In summary, we conclude that IFN� centrally promoteshigh-level production of IL-18 in CpG-pretreated mice andthat, compared to other toll agonists, LPS best stimulatesCpG-expanded IL-18 release. Because CpG expands IL-18production in T- and B-cell deWcient mice [11], cells of theinnate immune system are the likely targets of these immuno-modulatory eVects. One limitation of this study is that wehave not identiWed a cell source for IFN�-augmented IL-18synthesis, despite examination of IFN�-preincubated macro-phage and dendritic cell cultures or splenocytes and perito-neal macrophages derived from CpG-pretreated mice (datanot shown). Future studies need to determine if IL-18 is pro-duced by a non-traditional cell source –perhaps of endothe-lial or epithelial origin—that is uniquely recruited for IL-18synthesis in response to circulating IFN�. However, we canconclude that the synthetic pathway involved remainscaspase-1 dependent. Recently, synchronous stimulation ofmultiple toll receptors has been shown to mediate synergisticincreases in cytokine production [21]. The model of innateimmune modulation described here diVers in that the out-come is both sequence- and TLR-speciWc and occurs over aperiod of days. We speculate that the innate immune systemmay possess additional diVerentiative pathways distinct tospeciWc temporal patterns of exposure to microbial PAMPduring infection.
Acknowledgments
Fred Heinzel is supported by the VA Medical ResearchService and by NIH Grant AI45602, JeV Auletta by AI57801and Sameer Gupta by T32 HL07889. We thank Drs. EricPearlman, George Dubyak and Michelle Lin for discussion,encouragement and sharing of reagents in these studies.
References
[1] H. Okamura, H. Tsutsi, T. Komatsu, M. Yutsudo, A. Hakura, T. Tan-imoto, K. Torigoe, T. Okura, Y. Nukada, K. Hattoria, et al., Cloning
of a new cytokine that induces IFN-gamma production by T cells,Nature 378 (1995) 88–91.
[2] V.D. Joshi, D.V. Kalvakolanu, J.D. Hasday, R.J. Hebela, A.S. Cross,IL-18 levels and the outcome of innate immune response to lipopoly-saccharide: importance of a positive feedback loop with caspase-1 inIL-18 expression, J. Immunol. 169 (2002) 2536–2544.
[3] M.G. Netea, A.G. Vonk, M. van den Hoven, I. Verschueren, L.A.Joosten, J.H. van Krieken, W.B. van den Berg, J.W. Van der Meera,B.J. Kullberg, DiVerential role of IL-18 and IL-12 in the host defenseagainst disseminated Candida albicans infection, Eur. J. Immunol. 33(2003) 3409–3417.
[4] G.C. Pien, A.R. Satoskar, K. Takeda, S. Akiraa, C.A. Biron, Cuttingedge: selective IL-18 requirements for induction of compartmentalIFN-gamma responses during viral infection, J. Immunol. 165 (2000)4787–4791.
[5] R.J. Stuyt, M.G. Netea, J.H. van Krieken, J.W. van der Meera, B.J. Kull-berg, Recombinant interleukin-18 protects against disseminated Can-dida albicans infection in mice, J. Infect Dis. 189 (2004) 1524–1527.
[6] E. Seki, H. Tsutsui, H. Nakano, N. Tsuji, K. Hoshino, O. Adachi, K.Adachi, S. Futatsugi, K. Kuida, O. Takeuchi, H. Okamura, J. Fujim-oto, S. Akiraa, K. Nakanishi, Lipopolysaccharide-induced IL-18secretion from murine KupVer cells independently of myeloid diVer-entiation factor 88 that is critically involved in induction of produc-tion of IL-12 and IL-1 beta, J. Immunol. 166 (2001) 2651–2657.
[7] M.G. Netea, B.J. Kullberg, L.E. Jacobs, T.J. Verver-Jansen, J. van derVen-Jongekrijg, J.M. Galama, A.F. Stalenhoef, C.A. Dinarelloa, J.W.Van der Meer, Chlamydia pneumoniae stimulates IFN-gamma syn-thesis through MyD88-dependent, TLR2- and TLR4-independentinduction of IL-18 release, J. Immunol. 173 (2004) 1477–1482.
[8] V.B. Mehta, J. Harta, M.D. Wewers, ATP-stimulated release of inter-leukin (IL)-1beta and IL-18 requires priming by lipopolysaccharideand is independent of caspase-1 cleavage, J. Biol. Chem. 276 (2001)3820–3826.
[9] R. Sluyter, J.G. Dalitza, J.S. Wiley, P2X7 receptor polymorphismimpairs extracellular adenosine 5�-triphosphate-induced interleukin-18 release from human monocytes, Genes Immun. 5 (2004) 588–591.
[10] G. Fantuzzi, D.A. Reeda, C.A. Dinarello, IL-12-induced IFN-gammais dependent on caspase-1 processing of the IL-18 precursor, J. Clin.Invest. 104 (1999) 761–767.
[11] M.P. Gould, J.A. Greene, V. Bhoj, J.L. DeVecchioa, F.P. Heinzel, Dis-tinct modulatory eVects of LPS and CpG on IL-18-dependent IFN-gamma synthesis, J. Immunol. 172 (2004) 1754–1762.
[12] J.S. Cowdery, J.H. Chace, A.K. Yia, A.M. Krieg, Bacterial DNAinduces NK cells to produce IFN-gamma in vivo and increases thetoxicity of lipopolysaccharides, J. Immunol. 156 (1996) 4570–4575.
[13] H. Tsuji, N. Mukaida, A. Harada, S. Kaneko, E. Matsushita, Y. Naka-numa, H. Tsutsui, H. Okamura, K. Nakanishi, Y. Tagawa, Y. Iwakura,K. Kobayashia, K. Matsushima, Alleviation of lipopolysaccharide-induced acute liver injury in Propionibacterium acnes-primed IFN-gamma-deWcient mice by a concomitant reduction of TNF-alpha, IL-12, and IL-18 production, J. Immunol. 162 (1999) 1049–1055.
[14] J.M. Labasi, N. Petrushova, C. Donovan, S. McCurdy, P. Lira, M.M.Payette, W. Brissette, J.R. Wicks, L. Audolya, C.A. Gabel, Absence ofthe P2X7 receptor alters leukocyte function and attenuates an inXam-matory response, J. Immunol. 168 (2002) 6436–6445.
[15] D.G. Perregaux, P. McNiV, R. Laliberte, M. Conklyna, C.A. Gabel,ATP acts as an agonist to promote stimulus-induced secretionof IL-1 beta and IL-18 in human blood, J. Immunol. 165 (2000)4615–4623.
[16] M. Solle, J. Labasi, D.G. Perregaux, E. Stam, N. Petrushova, B.H.Koller, R.J. GriYthsa, C.A. Gabel, Altered cytokine production inmice lacking P2X(7) receptors, J. Biol. Chem. 276 (2001) 125–132.
[17] J.C. Deng, T.A. Moore, M.W. Newstead, X. Zeng, A.M. Kriega, T.J.Standiford, CpG oligodeoxynucleotides stimulate protective innateimmunity against pulmonary Klebsiella infection, J. Immunol. 173(2004) 5148–5155.
[18] A.M. Krieg, L. Love-Homan, A.K. Yia, J.T. Harty, CpG DNAinduces sustained IL-12 expression in vivo and resistance to

82 S. Gupta et al. / Cellular Immunology 243 (2006) 75–82
Listeria monocytogenes challenge, J. Immunol. 161 (1998) 2428–2434.
[19] S. Zimmermann, O. Egeter, S. Hausmann, G.B. Lipford, M. Rocken,H. Wagnera, K. Heeg, CpG oligodeoxynucleotides trigger protectiveand curative Th1 responses in lethal murine leishmaniasis, J. Immu-nol. 160 (1998) 3627–3630.
[20] A.S. Cross, Endotoxin tolerance-current concepts in historical per-spective, J. Endotoxin Res. 8 (2002) 83–98.
[21] G. Napolitani, A. Rinaldi, F. Bertoni, F. Sallustoa, A. Lanzavecchia,Selected Toll-like receptor agonist combinations synergistically trig-ger a T helper type 1-polarizing program in dendritic cells, Nat.Immunol. 6 (2005) 769–776.