Embed Size (px)
Citation preview

CP43-like chlorophyll binding proteins:structural and evolutionaryimplicationsJames W. Murray, James Duncan and James Barber
Division of Molecular Biosciences, Biochemistry Building, South Kensington Campus, Imperial College London, UK SW7 2AZ
CP43, encoded by the psbC gene, is a chlorophyll (Chl)-
binding protein of Photosystem II (PSII), the water-
splitting and oxygen-evolving enzyme of photosyn-
thesis. CP47, encoded by psbB, a Chl-binding protein
of PSII, is closely related to CP43. The Chl-binding six
transmembrane helical unit typified by CP43, is also
structurally related to the N-terminal domains of the
PsaA and PsaB proteins of Photosystem I (PSI) as well as
to the family of light-harvesting proteins encoded by
cyanobacterial isiA genes and prochlorophyte pcb
genes. Here we use recent structural information
derived for PSII and PSI to review similarities and
differences between the various members of the CP43-
like class of light-harvesting proteins, exploring both
functional and evolutionary implications.
CP43-like class of light-harvesting proteins
Plants, algae and cyanobacteria use two photosystems,Photosystem I (PSI) and Photosystem II (PSII), to absorblight energy and to provide the reducing equivalents andchemical energy needed to convert carbon dioxide intocarbohydrates and other organic molecules. PSII catalysesthe light-induced splitting of water leading to thegeneration of dioxygen and reducing equivalents in theform of reduced plastoquinone. PSI works in series withPSII and uses light energy to drive the reduction of NADPusing the reducing equivalents generated by PSII. Theredox coupling of PSII and PSI is provided by thecytochrome b6f complex and plastocyanin [1]. Despitehaving different functional properties, it is now clear thatPSI and PSII are structurally similar and originate from acommon ancestor. In PSII, the two proteins D1 and D2,which each have five transmembrane helices, form thereaction centre and bind most of the cofactors involved inthe water-splitting reaction. The CP43 and CP47 subunitsare closely associated with these two proteins. CP43 andCP47 have six transmembrane helices each and bindchlorophyll a (Chla) molecules, which function as a light-harvesting system for the PSII reaction centre. Bycontrast, the reaction centre proteins of PSI, PsaA andPsaB have eleven transmembrane helices each. The fiveC-terminal transmembrane helices of PsaA and PsaB arearranged in a manner similar to that of the
Corresponding author: Barber, J. ([email protected]).Available online 13 February 2006
www.sciencedirect.com 1360-1385/$ - see front matter Q 2006 Elsevier Ltd. All rights reserved
transmembrane helices of the D1 and D2 proteins.Similarly, the six transmembrane helices that make upthe N-terminal domain of PsaA and PsaB are organized inthe same way as those of CP43 and CP47. Here we discussthese similarities in some detail and extend the analysis toa class of Chl-binding proteins that are related to CP43.These CP43-like proteins are the IsiA proteins induced incyanobacteria in response to depletion of iron, and the Pcbproteins that form the Chla/b-binding proteins ofprochlorophytes*. Unlike PSI and PSII, there are, as yet,no crystal structures of these CP43-like proteins.
Comparison of CP43 and CP47
Hydropathy plots based on primary sequences predictedthat both CP43 and CP47 have six transmembrane heliceswith a large extrinsic domain joining the lumenal ends ofhelices V and VI [2]. Electron crystallographic analyses ofPSII revealed that these six transmembrane helices arearranged in three pairs related by a pseudo-threefold axis[3–5]. These analyses also indicated that the three pairs ofhelices of each protein are positioned on either side of theD1/D2 reaction centre proteins and, in this way, arerelated by the same pseudo-twofold axis that relates thefive transmembrane helices of D1 and D2 proteins and thecofactors that they bind. This organization has beenconfirmed by X-ray crystallography [6–8] (Figure 1a).
The structures of CP43 and CP47 derived from electroncrystallography were restricted to 8–10 A resolution and,therefore, did not provide side-chain information orstructural detail of the loops joining the transmembranehelices. However, they did give the first hints as to theorganization of Chls bound to these two PSII proteins [3].These studies, and later X-ray analyses [6–9], support thenotion that there is structural homology between CP43and CP47 and the N-terminal domains of the PSI reactioncentre proteins PsaA and PsaB [10] (Figure 1), except thatin PsaA and PsaB, the large extrinsic domains joiningtransmembrane helices V and VI of CP43 and CP47 arenot present (Figure 2).
The crystal structure of PSII isolated from thecyanobacterium Thermosynechococcus elongatus [8] hasshown that the large extrinsic domains of CP43 and CP47are different in size (132 and 191 amino acids,
Review TRENDS in Plant Science Vol.11 No.3 March 2006
* pcb genes are also found in some oxyphotobacteria (prochlorophyte-likecyanobacteria) that contain phycobiliproteins and bind only Chla (Fischerellamuscicola) or Chla and Chld (Acaryochloris marina).
. doi:10.1016/j.tplants.2006.01.007

(a) PSII (b) PSI
Figure 1. Top stromal views derived fromX-ray crystallography of (a) PSII [8] and (b)
PSI [9] to highlight the remarkable similarity in the organization of the
transmembrane helices of CP43 and D1 and CP47 and D2 with those of PsaA and
PsaB. The tetrapyrrole head groups of the primary electron donor Chls of PSII
(P680) and PSI (P700) are shown in green and, in this view, are seen from their sides.
Review TRENDS in Plant Science Vol.11 No.3 March 2006 153
respectively) and vary in their secondary structuralfeatures. The large loop of CP43 contains two long andthree short helices, with one being a 310-helix locatedwithin a motif fully conserved in all known sequences ofCP43: GGETMRFWD (residues 352–360†) (highlighted inorange in Figure 2a). The X-ray structure revealed thatthis 310 helix forms part of the water-splitting catalyticcentre providing a ligand, CP43 Glu354, for one of the Mnions of the Mn4-Ca cluster. The nearby CP43 Arg357 alsoseems to be a key side-chain for facilitating deprotonationof substrate water molecules during their oxidation [11].Other conserved regions of the large loop are involved inbinding the PsbO 33-kDa ‘manganese stabilizing’ protein[12]. The corresponding large loop of CP47 is located onthe D2 side of the reaction centre and has two long andfour short helices and three b-sheets. It also containsconserved residues that are involved in binding the PsbOprotein [12]. However, one conserved region is symmet-rically related to the 310-helix of CP43. This regioncontains phenylalanines (FFESFPVIL in T. elongatus,residues 362–370), which together with phenylalanines ofthe D2 protein, occupy the position equivalent to thatbinding the Mn4-Ca cluster on the D1 side.
The transmembrane regions of CP43 and CP47 haveseveral fully conserved histidine residues: ten in CP43 and13 in CP47 (see Supplementary material Figure S1). TheX-ray structure has shown that these His residues ligateChl molecules. Four further Chls were assigned to CP43and one of these is ligated by a fully conserved asparagine(CP43 Asn39) towards its N-terminus (replaced by aHis inCP47) (see Supplementary material Figure S1). Althoughthese ligating residues are not in a transmembrane regionthey are located in a surface helix and positioned withinthe membrane domain. The remaining Chls probably havewater molecules as ligands that are stabilized by side
† Unless stated otherwise, all amino acid numbering is for sequences ofThermosynechococcus elongatus. The Chl numbering for PSII and PSI are as inFerreira et al. [8] (1S5L pdb) and Jordan et al. [9] (1JB0 pdb), respectively.
www.sciencedirect.com
chains: CP43 Ser275 (helix V), CP43Met67 (helix I), CP47Ser241 (helix V) and CP47 Met37 (helix I). The assign-ment of the remaining Chl of CP43 (Chl47) is less certainowing to disorder; the Chl of CP47 (Chl46)† seems to be inpart stabilized by Trp185 (close to the lumenal surface ofhelix IV). Thirteen Chls of CP43 and CP47 are approxi-mately related to each other by the pseudo-twofold axisthat relates the transmembrane helices and cofactors ofthe D1 and D2 proteins. Within CP43, Chls (15,22,19),Chls (20,11,21) and Chls (18,16,17) are related around apseudo-threefold axis that relates the three pairs oftransmembrane helices of this protein. Similarly Chls(25,34,26), Chls (37,35,23), Chls (28,29,30) and (26,27,33)are also related by the pseudo-threefold axis of the CP47transmembrane helices. The majority of the Chls of CP43and CP47 are positioned in two layers towards the stromaland lumenal surfaces, eight in the stromal level for bothCP43 and CP47 and the remainder in the lumenal levelexcept for one Chl in each protein which is midway: CP43Chl13 ligated by CP43 His56, and CP47 Chl24 ligated byCP47 His26 (Figure 2). These two Chls are, therefore,located closely to adjacent Chls in the stromal andlumenal layers forming a stack of similarly orientatedChls: for CP43 Chl13, these are Chl15 (ligated to CP43His53) and Chl18 (ligated to CP43 His430), whereas forCP47 Chl24 the closest Chls in the stromal and lumenallayers are CP47 Chl25 (ligated to CP47 His23) and CP47Chl28 (ligated to CP47 His445). The function of thetransmembrane stack of Chls in CP43 and CP47 isunknown but probably aids energy transfer from onelayer to the other. Exciton interaction between these Chlsmight be the origin of the ‘red’ pigments known to exist inCP43 and CP47 [13]. Several of the histidine residues ofCP43 and CP47 have been mutated (reviewed in [14]),with the resulting phenotypes ranging from havingreduced PSII activity to a complete loss of photoauto-trophic growth because of the lack of assembly of PSII.One such study indicated that the Chl ligated to CP47His114 was the origin of the 695 nm emission at 77 K [15].
Chls can be ligated in an a- or b-configuration [16]. Thea-configuration is where the carbomethoxy group of ring Vof Chl is on the same side as the Mg2C ligand; the oppositeis true for the b-configuration (Figure 3). Usually Chlligation in photosynthetic antenna systems is by thea-configuration; this is true for the majority of the Chlsbound to CP43 and CP47, as can be seen in Figure S1 inthe Supplementary material. However, four Chls of CP43and CP47 have b-ligation and are all located on thestromal side. The possible significance of the b-configur-ation will be discussed later.
Comparison of CP43 and CP47 with the N-terminal
domains of PsaA and PsaB
The six N-terminal transmembrane helices of PsaA andPsaB, the reaction centre proteins of PSI [9], arestructurally homologous to those of CP43 and CP47, asshown in Figures 1 and 2. On superposition of thecrystallographic structural models, the root mean square(RMS) deviation of the Ca-atoms of the transmembranehelices of CP43 and CP47 is 1.4 A. This low value ismaintained when the transmembrane helices of CP43 and
CP43 CP47
(a) (b) (c)
(d) (e) (f)
(g) (h) (i)
(j) (k) (l)
PsaA PsaB
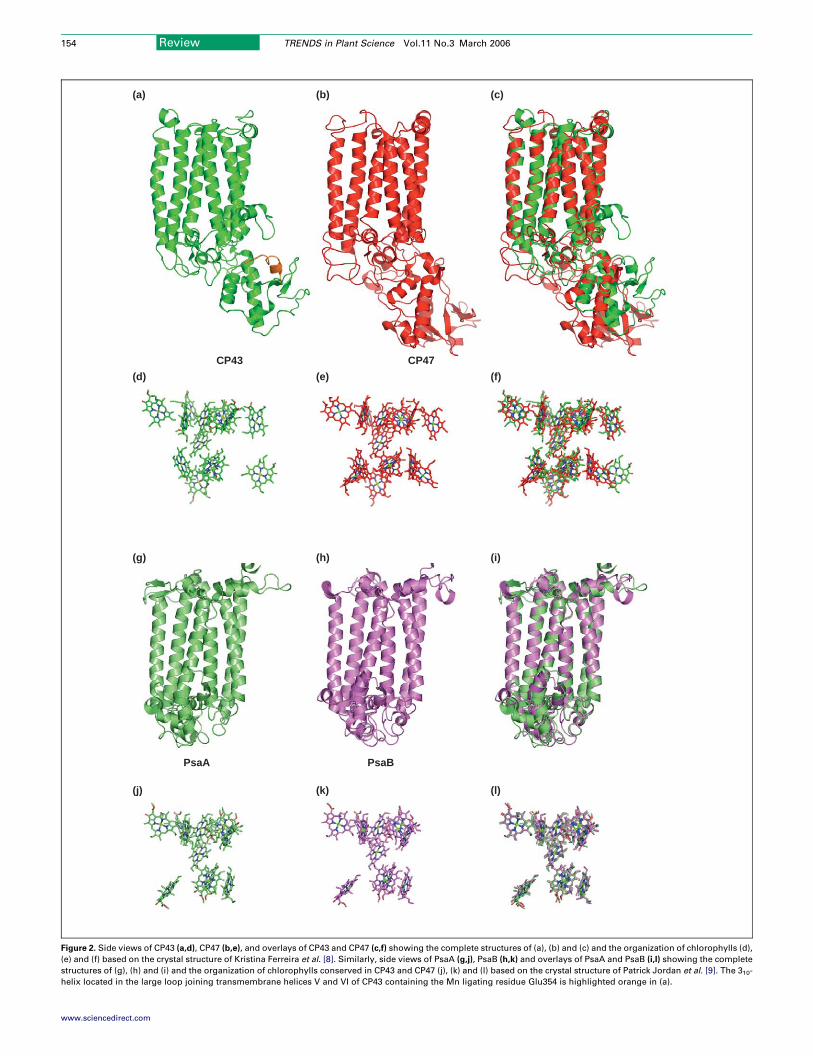
Figure 2. Side views of CP43 (a,d), CP47 (b,e), and overlays of CP43 and CP47 (c,f) showing the complete structures of (a), (b) and (c) and the organization of chlorophylls (d),
(e) and (f) based on the crystal structure of Kristina Ferreira et al. [8]. Similarly, side views of PsaA (g,j), PsaB (h,k) and overlays of PsaA and PsaB (i,l) showing the complete
structures of (g), (h) and (i) and the organization of chlorophylls conserved in CP43 and CP47 (j), (k) and (l) based on the crystal structure of Patrick Jordan et al. [9]. The 310-
helix located in the large loop joining transmembrane helices V and VI of CP43 containing the Mn ligating residue Glu354 is highlighted orange in (a).
Review TRENDS in Plant Science Vol.11 No.3 March 2006154
www.sciencedirect.com

(a) (b)α-configuration β-configuration
Figure 3. Chlorophylls can be ligated in an a- or b-configuration [18]. The
a-configuration is when the carbomethoxy group of ring V of Chl is on the same
side as the Mg2C ligand (a); the opposite is true in the b-configuration (b).
Prochlorothrix CP43
Acar yochloris PcbA
Synechocystis CP43
Spinach CP43Chlamydomonas CP43
Prochl MIT 9313 CP43Synechococcus WH8102 CP43
Prochl MED4 CP43Prochl SS120 CP43
Fischerella muscicola isiA
Prochloron PcbAProchlorothrix PcbAProchlorothrix PcbB
Prochl MIT 9313 CP47Prochl SS120 CP47Prochl MED4 CP47
Synechococcus WH8102 CP47
Prochlorothrix CP47Synechocystis CP47
Synechocystis PsaASynechocystis PsaB
Spinach CP47Chlamydomonas CP47
Prochl SS120 PcbAProchl SS120 PcbF
Prochl MED4 PcbA
Prochl SS120 PcbGProchl MIT 9313 PcbB
Prochl SS120 PcbBProchl SS120 PcbEProchl SS120 PcbCProchl SS120 PcbDProchl SS120 PcbH
Prochl MIT 9313 PcbA
100
100
8396
81
61
54
100
6894
88
51
100
100
70
6664
6968
100
95
92100
99
88
80
84
100
Synechocystis isiAT. elongatus isiA
T. elongatus CP43
T. elongatus CP47
Anabaena PCC 7120 isiA
Fischerella muscicola PcbCProchlorothrix PcbC
TRENDS in Plant Science
Figure 4. Bootstrap consensus phylogenetic tree of CP43, CP47, IsiA, Pcb and the N-
terminal domain of the PSI proteins PsaA and PsaB. The sequences were retrieved
from the Swiss-Prot database and aligned with ClustalX [44]. This phylogenetic tree
was built from the aligned sequences and the MEGA3 (Molecular Evolutionary
Genetic Analysis) software [45] using the neighbour-joining algorism. Boot-
strapping with 500 replicates was used as a test for inferred phylogeny and
bootstrap values are displayed on the nodes. Abbreviation: Prochl, Prochlorococ-
cus marinus.
Review TRENDS in Plant Science Vol.11 No.3 March 2006 155
CP47 are compared with six N-terminal helices of PsaAand PsaB (RMS deviations are w2.2 A). This strikingstructural correspondence between the PSII and PSIproteins is also found when comparing the Chls theybind. Remarkably ten of the 14 Chls bound to CP43 and 12of the 16 Chls of CP47 are conserved within theN-terminal domains of PsaA and PsaB. All theseconserved Chls are ligated by His residues [except PsaAChl1123, which is ligated by an asparagine (Asn359)]. Theconservation of these Chls is also maintained in their a- orb-ligating conformations. It has been argued that thecluster of b-Chls in N-terminal domains of PsaA and PsaBhelp to stabilize the transmembrane helices by providingH-bonding (P. Braun, personal communication) and,therefore, this is also likely to be the case for CP43 andCP47. Moreover, Teodor Silviu Balaban and colleagues[16] argue that in PSI, the b-Chls play an important role inenergy transfer and the same argument can be appliedto PSII.
The conservation of Chls between PSII and PSI meansthat PsaA Chl1104 ligated to His79 and PsaB Chl1203ligated to His52 are equivalent to CP43 Chl13 and CP47Chl24, which bridge the stromal and lumenal Chl layers.In PSI, there are several such ‘bridging’ Chls that wouldfacilitate energy transfer between stromal and lumenalsurfaces. The precise role of PsaA Chl104 and PsaBChl203 has yet to be elucidated.
Sergej Vasil’ev and Doug Bruce [17] investigated theconservation of Chls between CP43 and CP47 and theN-terminal domains of PsaA and PsaB and conducted asimulation of energy transfer between them when theorientation of each Chl was adjusted systematically. Thisanalysis revealed that a Chl in CP43 (CP43 Chl14 ligatedto CP43 His441) and a symmetry related Chl in CP47(CP47 Chl31 ligated to CP47 His466), which are located inhelix VI of both proteins (see Supplementary materialFigure S1) but are not conserved in PSI, had the greatesteffect on modulating energy transfer to the reactioncentre. Similarly, they identified a Chl in PsaA and inPsaB whose orientation greatly influenced the simulatedenergy-transfer calculation. These Chls, PsaA Chl26 andPsaB Chl24, are ligated by PsaA His396 and PsaB His337located in transmembrane helix ‘f ’ (corresponding to
www.sciencedirect.com
helix VI of CP43 and CP47) and are not conserved inPSII. It was concluded that these pairs of Chls, specific toPSII or to PSI act as ‘bridging pigments’ between thelight-harvesting Chls and the corresponding reactioncentre. A diffusion-limited transfer mechanism for PSII isfavoured by some [18]; others argue that energy transfer tothe PSII reaction centre is fast, as it is in PSI, and that therelatively long lifetime of the excited state of PSII is becauseof the shallow excitonic trap of the primary electron donorP680 [19,20]. As far as we are aware, CP43 His441 andCP47 His466, which ligate the ‘regulatory Chls’ identifiedby Vasil’ev and Bruce [17], have not beenmutated to enableinvestigation of their function or to address the controversyabout energy-transfer mechanisms in PSII.
Other CP43-like proteins
In spite of the remarkable structural similarity betweenCP43 and CP47 and the N-terminal domains of PsaA andPsaB, the CP43 protein is phylogenetically more closelyrelated to the IsiA proteins of cyanobacteria and to the Pcbproteins of prochlorophytes andprochlorophyte-like cyano-bacteria as shown in Figure 4. There are no crystal

Review TRENDS in Plant Science Vol.11 No.3 March 2006156
structures available for these CP43-like proteins but it isclear from sequence alignments (see Supplementarymaterial Figure S1) that they contain six transmembranehelices and do not have an extensive extrinsic domainjoining the lumenal ends of helices Vand VI as CP43 does.In the phylogenetic analysis shown in Figure 4, the IsiAproteins are highly conservedwhereas the Pcb proteins aremore divergent.
Comparison of CP43 and IsiA proteins
isiA genes are expressed in response to depletion of ironlevels (iron stress induced) [21], although other stressconditions can turn on this gene, such as high levels oflight [22]. The IsiA protein, like CP43, normally bindsChla although it can be induced to bind some Chlb whenthe gene for the enzyme Chla oxidase (cao gene) isengineered into cyanobacteria [23]. When expressed atrelatively low levels, the IsiA protein forms an 18-mer ringaround the trimeric PSI reaction centre core of cyano-bacteria [24,25]. The ring acts as an efficient light-harvesting system for PSI as shown by steady-state andtransient optical spectroscopy [26,27]. The most-detailed3D structure of this IsiA-PSI supercomplex has beendetermined by electron cryomicroscopy (cryo-EM) of non-stained preparations followed by single particle analysis[28]. Although the electron densitymap had a resolution ofw20 A, it provided a framework for incorporating theX-ray-derived structure of PSI [9] and for IsiA based onthe X-ray structure of CP43 [8].
Figure S1 in the Supplementary material shows theconservation of the membrane-located His residuesbetween CP43 and IsiA using the T. elongatussequences. These His residues are conserved in allknown sequences of CP43 and IsiA, as is theChl-ligating CP43 Asn39. However, there are someinteresting differences. CP43 His56, which ligates theChl linking stromal and lumenal Chl layers, is replacedby a glutamine in IsiA. It is likely that Gln will ligate aChl and that this ‘linker’ pigment is maintained in theIsiA protein. An extra His in helix III adjacent to theequivalent CP43 His164 suggests that an additional Chlis bound in this position, a possibility that will beconsidered in more depth below. The regulatory Chlidentified in CP43 by Vasil’ev and Bruce [17] is ligatedto CP43 His441 and this residue is conserved in IsiA.
Comparison of CP43 with Pcb proteins
Pcb proteins provide the outer light-harvesting systemsof PSII and PSI of prochlorophytes (Prochloron didemni,Prochlorothrix hollandica and Prochlorococcus marinus)[29] and prochlorophyte-like cyanobacteria (Fischerellamuscicola and Acaryochloris marina) [30,31]. The num-ber of pcb genes varies between organisms and betweenstrains [32]. For example, Prochlorococcus MED4, astrain adapted to high light at the surface of the oceans,has one pcb gene whereas the low light, deep-living strainSS120 has eight pcb genes [33,34]. Some pcb genesencode Pcb proteins that form a light-harvesting systemfor PSII, and others are targeted to PSI where they forman 18-mer ring much like that of the IsiA protein [35–39].In Prochloron, Prochlorothrix and Prochlorococcus, the
www.sciencedirect.com
Pcb proteins bind both Chla and Chlb, the ratio varyingbetween different organisms and strains [33]. In the caseof Prochlorococcus MED4, the Chlb level is low comparedwith that of Chla, whereas SS120 has a Chlb:Chla ratiogreater than one. The Pcb proteins of Fischerella bindonly Chla [30] whereas in Acaryochloris the dominantpigment is Chld [31]. In addition, Acaryochloris CP43,CP47, PsaA and PsaB bind high levels of Chld ratherthan Chla [40,41].
Figure S1 in the Supplementary material presents asequence comparison of Pcb proteins aligned withT. elongatus sequences of CP43, CP47, N-terminaldomains of PsaA and PsaB, and IsiA. In many Pcbproteins the residue equivalent to CP43 His 56 of thefirst transmembrane helix remains a His or is a Gln as inIsiA. All Pcb and IsiA proteins share the dissimilarity withCP43 in having an extra His in transmembrane helix III.In other respects, Pcb proteins, like IsiA, have Hisresidues that are conserved with those of CP43, includingthe His of helix VI, which has been proposed to bind the‘linker’ Chl of CP43 [17].
Implications of differences between CP43 and IsiA
and the Pcb proteins
Kristina Ferreira et al. [8] elucidated the structure ofPSII, revealing details of the involvement of the largeextrinsic loop of CP43 in both the ligation of the Mn4-Cacluster of the oxygen-evolving catalytic centre (which hadnot been predicted) and its role as a ‘docking site’ for thePsbO protein. The role of CP43 in binding the PsbO hadbeen suggested from a range of mutational and cross-linking studies (reviewed in [14]). However, these specificproperties of CP43 are related to the water-splittingreaction of PSII and are in addition to its function as alight-harvesting protein. IsiA and Pcb proteins functiononly as Chl-binding proteins involved in light harvestingand, in the case of IsiA, probably also inphotoprotection [42].
Overall it is likely that IsiA and Pcb proteins share thesame Chl binding sites with CP43, at least those that areinvolved directly in ligation with side chains. There is oneinteresting difference in that there is an extra His residuein helix III that is fully conserved in all known IsiA andPcb sequences (see Supplementary material Figure S1).This extra His is likely to ligate a Chl molecule and thequestion is, what is its function?
All structures of Pcb–PSII, Pcb–PSI and IsiA–PSIsupercomplexes derived from electron microscopy seemto exhibit a common feature: the interaction betweenadjacent Pcb or IsiA proteins is tight compared withinteractions with the surfaces of the PSI and PSII reactioncentre cores [28]. Therefore, it is possible that thisproposed extra Chl, not found in CP43, plays a role inaiding energy transfer between adjacent Pcb orIsiA subunits.
Evolutionary significance
The six transmembrane helical chlorophyll-bindingproteins, typified by CP43, is a basic building block ofthe photosynthetic reaction centre and light-harvestingsystems. Given the distinct structural differences between

Review TRENDS in Plant Science Vol.11 No.3 March 2006 157
reaction centre proteins (transmembrane helicesarranged in a ‘hand-shake’ motif) and CP43-like proteins(transmembrane helices arranged in a ‘circular’ motif), itseems likely that the Type-1 reaction centres typified byPSI came about from gene fusion resulting in an eleventransmembrane helical protein. By contrast, ArmenMulkidjanian and Wolfgang Junge [43] hypothesize thatType-1 reaction centres originate from an eleven trans-membrane unit. In our view, no fusion occurred in the caseof PSII. Gene duplication and modification haveultimately led to the modern reaction centres. However,the structure of the CP43-like proteins with theirpseudo-threefold symmetry suggests that the precursorto this family was a single transmembrane helix thatby gene duplication and fusion initially converted toa two-helix bundle that ultimately gave rise to the3!2 pairs of transmembrane helices characteristic ofthe CP43-like family. Whether the IsiA and Pcbproteins were derived from CP43, as argued byMin Chen et al. [32], or from a precursor of CP43 is amatter of debate.
Acknowledgements
We acknowledge financial support from the Biotechnology and BiologicalScience Research Council (BBSRC).
Supplementary data
Supplementary data associated with this article can befound at doi:10.1016/j.tplants.2006.01.007
References
1 Barber, J. and Andersson, B. (1994) Revealing the blue-print ofphotosynthesis. Nature 370, 31–34
2 Barber, J. et al. (2000) Revealing the structure of the photosystem IIchlorophyll binding proteins, CP43 and CP47. Biochim. Biophys. Acta1459, 239–247
3 Rhee, K.H. et al. (1998) Three-dimensional structure of the plantphotosystem II reaction centre at 8 A resolution. Nature 396, 283–286
4 Hankamer, B. et al. (1999) Revealing the structure of the oxygen-evolving core dimer of photosystem II by cryoelectron crystallography.Nat. Struct. Biol. 6, 560–564
5 Hankamer, B. et al. (2001) Three-dimensional structure of thephotosystem II core dimer of higher plants determined by electronmicroscopy. J. Struct. Biol. 135, 262–269
6 Zouni, A. et al. (2001) Crystal structure of photosystem II fromSynechococcus elongatus at 3.8 A resolution. Nature 409, 739–743
7 Kamiya, N. and Shen, J.R. (2003) Crystal structure of oxygen-evolvingphotosystem II from Thermosynechococcus vulcanus at 3.7-A resol-ution. Proc. Natl. Acad. Sci. U. S. A. 100, 98–103
8 Ferreira, K.N. et al. (2004) Architecture of the photosynthetic oxygen-evolving center. Science 303, 1831–1838
9 Jordan, P. et al. (2001) Three-dimensional structure of cyanobacterialphotosystem I at 2.5 A resolution. Nature 411, 909–917
10 Schubert, W.D. et al. (1998) A common ancestor for oxygenic andanoxygenic photosynthetic systems: a comparison based on thestructural model of photosystem I. J. Mol. Biol. 280, 297–341
11 McEnvoy, J.P. and Brudvig, G.W. (2004) Structure-based mechanismof photosynthetic water oxidation. Phys. Chem. Chem. Phys. 6,4754–4763
12 De Las Rivas, J. and Barber, J. (2004) Analysis of the structure of thePsbO protein and its implications. Photosynth. Res. 81, 329–343
13 de Weerd, F.L. et al. (2002) Pathways for energy transfer in the corelight-harvesting complexes CP43 and CP47 of photosystem II.Biophys. J. 82, 1586–1595
14 Eaton-Rye, J.J. and Putnam-Evans, C. (2005) The proximal antennaproteins CP47 and CP43. In Photosystem II: The Water Plastoquinone
www.sciencedirect.com
Oxido-Reductase of Photosynthesis. Advances in Photosynthesis andRespiration Series (Wydrzynski, T.J. et al., eds), pp. 45–70, KluwerAcademic Publishers
15 Shen, G. and Vermaas, W.F.J. (1994) Mutations of chlorophyll ligandsin the chlorophyll-binding CP47 proteins as studied in a Synechocystissp. PCC 6803 photosystem I-less background. Biochemistry 33,7379–7388
16 Balaban, T.S. et al. (2002) Relevance of the diasteriotopic ligation ofmagnesium atoms of chlorophylls in Photosystem I. Biochim. Biophys.Acta 1556, 197–207
17 Vasil’ev, S. and Bruce, D. (2004) Optimization and evolution of lightharvesting in photosynthesis: the role of antenna chlorophyllconserved between photosystem II and photosystem I. Plant Cell 16,3059–3068
18 Gobets, B. et al. (2003) Excitation wavelength dependence of thefluorescence kinetic in Photosystem I. Particles from SynechococcusPCC 6809 and Synechococcus elongatus. Biophys. J. 85, 3883–3898
19 Schatz, G.H. et al. (1988) A kinetic and energetic model for primaryprocesses in Photosystem II. Biophys. J. 54, 397–405
20 Barter, L.M.C. et al. (2001) Relationship between excitation energytransfer, trapping and antenna size in Photosystem II. Biochemistry40, 4026–4034
21 Straus, N. (1994) Iron deprivation: physiology and gene regulation. InThe Molecular Biology of Cyanobacteria (Bryant, D.A., ed.), pp.731–750, Kluwer Academic Publishers
22 Havaux, M. et al. (2005) The chlorophyll-binding protein IsiA isinducible by high light and protects the cyanobacterium SynechocystisPCC 6803 from photooxidative stress. FEBS Lett. 579, 2289–2293
23 Duncan, J. et al. (2003) Exploring the ability of chlorophyll b to bind tothe CP43 0 protein induced under iron deprivation in a mutant ofSynechocystis PCC 6803 containing the cao gene. FEBS Lett. 541,171–175
24 Bibby, T.S. et al. (2001) Iron deficiency induces the formation of anantenna ring around trimeric photosystem I in cyanobacteria. Nature412, 743–745
25 Boekema, E.J. et al. (2001) A giant chlorophyll–protein complexinduced by iron deficiency in cyanobacteria. Nature 412, 745–748
26 Andrizhiyevskaya, E.G. et al. (2002) Spectroscopic properties of PSI-IsiA supercomplexes from the cyanobacterium Synechococcus PCC7942. Biochim. Biophys. Acta 1556, 265–272
27 Melkozernov, A.N. et al. (2003) Time-resolved absorption and emissionshow that the CP43 0 antenna ring of iron-stressed Synechocystis sp.PCC6803 is efficiently coupled to the photosystem I reaction centercore. Biochemistry 42, 3893–3903
28 Nield, J. et al. (2003) Structural analysis of the photosystem Isupercomplex of cyanobacteria induced by iron deficiency. Biochem-istry 42, 3180–3188
29 Matthijs, H.C. et al. (1994) Prochlorophytes: the ‘other’ cyanobacteria.In The Molecular Biology of Cyanobacteria (Bryant, D.A., ed.), pp.49–64, Kluwer Academic Publishers
30 Geiss, U. et al. (2001) The iron-regulated isiA gene of Fischerellamuscicola strain PCC 73103 is linked to a likewise regulated geneencoding a Pcb-like chlorophyll-binding protein. FEMS Microbiol.Lett. 197, 123–129
31 Chen, M. et al. (2002) The major light-harvesting pigment protein ofAcaryochloris marina. FEBS Lett. 514, 149–151
32 Chen, M. et al. (2005) Unique origin and lateral transfer of prokaryoticchlorophyll b and chlorophyll d light harvesting systems. Mol. Biol.Evol. 22, 21–28
33 Partensky, F. et al. (1999) Prochlorococcus, a marine photosyntheticprokaryote of global significance. Microbiol. Mol. Biol. Rev. 63,106–127
34 Hess, W.R. (2004) Genome analysis of marine photosynthetic microbesand their global role. Curr. Opin. Biotechnol. 15, 191–198
35 Bibby, T.S. et al. (2003) Low-light-adapted Prochlorococcus speciespossess specific antennae for each photosystem. Nature 424,1051–1054
36 Bibby, T.S. et al. (2003) Structure of a photosystem II supercomplexisolated from Prochloron didemni retaining its chlorophyll a/b light-harvesting system. Proc. Natl. Acad. Sci. U. S. A. 100, 9050–9054
37 Bumba, L. et al. (2005) Antenna ring around trimeric Photosystem I inchlorophyll b containing cyanobacterium Prochlorothrix hollandica.Biochim. Biophys. Acta 1708, 1–5

Review TRENDS in Plant Science Vol.11 No.3 March 2006158
38 Chen, M. et al. (2005) Iron deficiency induces a chlorophyll d-bindingPcb antenna system around Photosystem I in Acaryochloris marina.Biochim. Biophys. Acta 1708, 367–374
39 Chen, M. et al. (2005) Structure of a large photosystem II super-complex from Acaryochloris marina. FEBS Lett. 579, 1306–1310
40 Miyashita, H. et al. (1996) Chlorophyll d as a major pigment. Nature383, 402
41 Hu, Q. et al. (1998) A photosystem I reaction center driven bychlorophyll d in oxygenic photosynthesis. Proc. Natl. Acad. Sci. U. S.A. 95, 13319–13323
42 Park, Y.I. et al. (1999) Expression of the isiA gene is essential for thesurvival of the cyanobacterium Synechococcus sp. PCC 7942 by
Plant Science meeti
3rd International Symposium Phosphoru
14–19 May
Uberlandia, Minas
http://www.cnpms.em
RNA Biology: Novel Insigh
18–20 May
Penn State Univer
http://plantphysiolo
2nd Pan American Plant Mem
17–20 May
Sheraton Hotel Con
South Padre Isla
http://www.panamericanplantmem
EPSO: Plant Dynamics – From
28 May–1 Ju
Visegrad, H
http://www.epsoweb.org/
www.sciencedirect.com
protecting photosystem II from excess light under iron limitation.Mol.Microbiol. 32, 123–129
43 Mulkidjanian, A.Y. and Junge, W. (1997) On the origin of photosyn-thesis as inferred from sequence analysis – a primordial UV-protectoras common ancestor of reaction centers and antenna proteins.Photosynth. Res. 51, 27–42
44 Ramu, C. et al. (2003) Multiple sequence alignment withCluster series of programs. Nucleic Acids Res. 31, 3497–3500
45 Kumar, S. et al. (2004) MEGA3: integrated software for molecularevolutionary genetics analysis and sequence alignment. Brief. Bioin-form. 5, 150–163
ngs in May 2006
s Dynamics in Soil–Plant Continuum
2006
Gerais, Brazil
brapa.br/simposio/
ts from Plant Systems
2006
sity, PA, USA
gy.cas.psu.edu/
brane Biology Workshop
2006
ference Center
nd, TX, USA
branebiologyworkshop.org/
Molecules to Ecosystems
ne 2006
ungary
catalog/Conf2006.htm