Embed Size (px)
Citation preview

REVIEW
CURRENTOPINION A systems biology approach for diagnostic and
vaccine antigen discovery in tropical infectiousdiseases
Copyright
www.co-infectiousdiseases.com
Li Liang and Philip L. Felgner
Purpose of review
There is a need for improved diagnosis and for more rapidly assessing the presence, prevalence, andspread of newly emerging or reemerging infectious diseases. An approach to the pathogen-detectionstrategy is based on analyzing host immune response to the infection. This review focuses on a proteinmicroarray approach for this purpose.
Recent findings
Here we take a protein microarray approach to profile the humoral immune response to numerousinfectious agents, and to identify the complete antibody repertoire associated with each disease. The resultsof these studies lead to the identification of diagnostic markers and potential subunit vaccine candidates.These results from over 30 different organisms can also provide information about common trends in thehumoral immune response.
Summary
This review describes the implications of the findings for clinical practice or research. A systems biologyapproach to identify the antibody repertoire associated with infectious diseases challenge using proteinmicroarray has become a powerful method in identifying diagnostic markers and potential subunit vaccinecandidates, and moreover, in providing information on proteomic feature (functional and physicallyproperties) of seroreactive and serodiagnostic antigens. Combining the detection of the pathogen with acomprehensive assessment of the host immune response will provide a new understanding of thecorrelations between specific causative agents, the host response, and the clinical manifestations of thedisease.
Keywords
antigen, immune response, protein microarray, serodiagnostic
Division of Infectious Diseases, Department of Medicine, University ofCalifornia, Irvine, California, USA
Correspondence to Philip L. Felgner, Division of Infectious Diseases,Department of Medicine, University of California, Irvine, CA 92697, USA.Tel: +1 949 824 1407; fax: +1 949 824 9675; e-mail: [email protected]
Curr Opin Infect Dis 2015, 28:438–445
DOI:10.1097/QCO.0000000000000193
INTRODUCTION
A major component of the adaptive immuneresponse to infection is the generation of protectiveand long-lasting humoral immunity. Analyses ofantibody responses against different infectiousagents are critical for diagnosing infectious diseases,understanding pathogenic mechanisms, and thedevelopment and evaluation of vaccines. Proteinmicroarrays are well suited to identify, quantify,and compare individual antigenic responses follow-ing exposure to infectious agents. It can now evalu-ate antibody responses to hundreds, or eventhousands, of recombinant antigens at one time.These large-scale studies have uncovered new anti-genic targets, provided new insights into vaccineresearch and yielded an overview of immunoreac-tivity against almost the entire proteome of certainpathogens. This technology can be applied to the
© 2015 Wolters Kluwer
development of improved serodiagnostic tests,discovery of subunit vaccine antigen candidates,epidemiologic research, and vaccine development,as well as providing novel insights into infectiousdisease and the immune system. In this review, wewill discuss the use of protein microarrays as apowerful tool to define the humoral immuneresponse to bacteria and viruses.
Factors governing selection of the particularantigens recognized are unclear [1,2]. It is not
Health, Inc. All rights reserved.
Volume 28 � Number 5 � October 2015

KEY POINTS
� Protein microarray is a powerful tool in identifyingdiagnostic markers and potential subunit vaccinecandidates.
� Full proteome microarrays have provided valuableinformation on antigen predictions.
� Certain proteomic features (functional and physicallyproperties) have been identified to be associated withseroreactive and serodiagnostic antigens.
� Naive Bayes classification further improves thesensitivity and specificity of this in-silico predictivealgorithm.
Antibody prediction for infectious diseases Liang and Felgner
uncommon for viruses encoding a small numberof proteins to generate antibodies against eachencoded protein. But for infectious agents contain-ing hundreds or thousands of proteins only a subsetof the proteome is recognized and little is knownabout the extent or the characteristics of this subsetof antigens. Methods for making a complete empiri-cal accounting of the immunoproteome havelimitations, particularly when the genome of theorganism is large. The Protein Microarray Labora-tory at University of California Irvine has developeda highly efficient method to determine the humoralimmune response to microbial antigens. We haveapplied this approach to more than 30 medicallyimportant infectious microorganisms [3–33]including Mycobacterium tuberculosis, Plasmodiumfalciparum, Plasmodium vivax, Brucella melitensis,Chlamydia trachomatis, Francisella tularensis, Burkhol-deria pseudomallei, Coxiella burnetii, Borrelia burgdor-feri, Salmonella enterica Typhi, Rickettsia prowazekii,Rickettsia rickettsii, Orientia tsutsugamushi, Bartonellahenselae, Leptospira interrogans, Toxoplasma gondii,Candida albicans, Schistosoma mansoni, and virusesincluding vaccinia herpes simplex viruses 1&2,varicella zoster virus, Epstein–Barr virus, humanpapillomaviruses, HIV, dengue, influenza, WestNile virus, yellow fever, Saint Louis encephalitis,Japanese encephalitis, and chikungunya viruses.After launching this project 10 years ago, we havemade more than 40 000 plasmids, printed theencoded proteins on 25 000 microarrays and probedthe arrays with 15 000 serum specimens in order todetermine disease-associated antibody profiles inpeople infected with each agent. These chips canbe probed with sera from infected patients to deter-mine the immunodominant antigens for each agentand the methodology is amenable to the screeningof sera from very large cohorts numbering in thethousands. When seroreactive and serodiagnostic
Copyright © 2015 Wolters Kluwe
0951-7375 Copyright � 2015 Wolters Kluwer Health, Inc. All rights rese
antigen subsets from different infectious agentsare printed onto the same array, the chip can dis-criminate between patients infected with differentagents and also identify individuals with coinfec-tions or multiple infections. We have shown thatthe individual proteins printed on these arrayscapture antibodies present in serum from infectedindividuals and the amount of captured antibodycan be quantified using fluorescent secondary anti-body. In this way, a comprehensive profile of anti-bodies that result after infection or exposure can bedetermined that is characteristic of the type ofinfection and the stage of disease [9,10,31].
Here we summarize the approximate seroreac-tive and serodiagnostic antigens that were identifiedand published in 30 different organisms, and discussthe antibody response predictions from classifi-cation of reactive antigens based on functionaland physical properties.
PROTEIN MICROARRAY PRODUCTION,PROBING, AND ANALYSIS
Genes were amplified and cloned using a high-throughput PCR and recombination method [29].Open reading frames from genomic DNA or cDNAwere identified and amplified using gene-specificprimers containing about 20 bp nucleotide exten-sion complementary to ends of linearized pXT7vector, which allows homologous recombinationbetween the PCR product and pXT7 vector in com-petent Escherichia coli cells. The resulting fusionproteins also harbored a hemagglutinin epitope atthe 30 end and polyhistidine at the 50 end. Plasmidswere expressed at 24 8C in a 16 h in-vitro transcrip-tion/translation E. coli system (expressway kits fromInvitrogen). For no DNA controls, no plasmid DNAwas added to the same amount of reagent fromin-vitro transcription/translation E. coli system totest E. coli background reactivity. For microarrays,10 ml of reaction was mixed with 3.3 ml 0.2% Tween20 to give a final concentration of 0.05% Tween 20,and printed onto nitrocellulose coated glass FASTslides (Whatman) using an Omni Grid 100 micro-array printer (Genomic Solutions). Sera sampleswere diluted in E. coli lysate (Mclab). Slides wereincubated in biotin-conjugated secondary antibody(Jackson ImmunoResearch) and detected by incu-bation with streptavidin-conjugated SureLight P-3(Columbia Biosciences). Microarray slides werescanned and analyzed using a Perkin Elmer Scan-Array Express HT or Genepix microarray scanner.Intensities were quantified. All signal intensitieswere corrected for spot-specific background. Allforeground values were transformed and normal-ized using a robust linear model or nonlinear
r Health, Inc. All rights reserved.
rved. www.co-infectiousdiseases.com 439

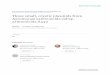
HT PCRCloning
OutputProteinInput
Plasmid
4 hrs
Cell free in vitroExpression
GenomicDNA
2, 000+genes perweek
KN(R)Linear vector
T7
ORF
Laser confocal microarray scanner
HAHISATG
Printing
Probing
Analysis
FIGURE 1. Microarray production, printing, and analysis. Thousands of genes of interest were cloned using a high-throughputmethod, in-vitro expressed in an Escherichia coli cell free system. Protein microarrays were then produced, probed, and datawere analyzed as described in the text. ORF, open reading frame.
Tropical and travel-associated diseases
variance stabilizing normalization to remove sys-tematic effects [24,34,35] (Fig. 1).
PERCENTAGE OF SEROREACTIVEANTIGENS
Discovery of novel antigens associated with infec-tious diseases is fundamental to the development ofserodiagnostic tests and protein subunit vaccinesagainst existing and emerging pathogens. Throughover 10 years of effort, we have identified over 1000antigens associated with infections or vaccinations in30 different organisms (Table 1) [3–6,9–17,23,25
&&
,31–33,36
&
,37–39,40&&
,41,42,43&
,44,45&
,46&
,47&&
],accounting for around 2–5% of bacterial genome;20–57% of viral genome; and 10–45% of parasitegenomes. Antigens differentially reactive amonginfected and healthy controls comprise even smallerpercentage of the genome size: from 0.3 to 3% forbacteria; 16 to 40% for viruses, and 2 to 18% forparasites. Borrelia burgdorferi, however, generatehigher antibody responses against approximately15% of polypeptides during natural infection, ofwhich half are differentially reactive between natu-rally infected and uninfected individuals [10].
Antigens were classified as ‘seroreactive’ withmean reactivity greater than 2–3 standard devi-ations above the mean of the negative controls inmost organisms; and differentially reactive antigensare classified by Benjamini-Hochberg adjusted P
Copyright © 2015 Wolters Kluwer
440 www.co-infectiousdiseases.com
value smaller than 0.05 by comparing the negativegroup with infected or vaccinated individuals.
Full proteome microarrays were constructed foronly a limited number of bacterial species; however,other data were published using partial arrays con-taining only partial proteome, and may over-represent the percentages of seroreactive and sero-diagnostic antigens in the full proteome because thesubset of proteins on the array was selected based onantigenic features seen previously.
ENRICHMENT ANALYSIS REVEALSPHYSICAL PROPERTIES AND CELLULARFUNCTIONS ASSOCIATED WITHIMMUNOGENICITY
Another application for these empirical data is totrain an algorithm to predict reactive antigens insilico, and several studies from our group applyenrichment analyses to identify proteomic featuresthat tend to be seen more frequently in the seror-eactive and serodiagnostic antigen sets [12,17,23].
Efforts to predict antigenicity have relied on afew computational algorithms predicting signalpeptide sequences (signalP), transmembranedomains (TMHMM), or subcellular localization(Psort). The current database from this proteinmicroarray approach contains quantitative antibodyreactivity data against 40 000 proteins derivedfrom 30 infectious microorganisms and more than
Health, Inc. All rights reserved.
Volume 28 � Number 5 � October 2015

Copyright © 2015 Wolters Kluwer Health, Inc. All rights reserved.
Tab
le1
.A
nov
ervi
ewof
sero
reac
tive
antig
ens
and
diffe
rent
ially
reac
tive
antig
ens
iden
tifie
din
diffe
rent
orga
nism
s
Ca
teg
ory
Tota
l#
pro
tein
s#
com
ple
ted
a%
com
ple
ted
%re
act
ive
ant
igen
s%
dif
fere
ntia
llyre
act
ive
ant
igen
sR
efer
ence
s
Virus
esRe
trovi
ruse
s:H
IV1&
2(fi
vesu
btyp
es,
four
clad
es)
83
74
89%
28%
[36
&
]
HPV
viru
ses
(13
type
s)104
104
100%
57%
16%
[32]
Orth
opox
viru
ses:
thre
ety
pes
260
260
100%
20
–22%
–[9
,31]
Her
pes
viru
ses:
HSV
1&
2,
VZV
,EB
V300
270
90%
28.5
–42%
25.9
–41.7
%[3
7]
Flav
ivirus
es:
WN
V,
deng
ue,
YF,
SLE,
JEA
50
50
100%
NP
NP
Alp
havi
ruse
s:C
hiku
ngun
yaC
10
10
100%
NP
NP
Bact
eria
Bruc
ella
mel
itens
isB
3198
3046
95%
4%
1.1
0%
[12]
Chl
amyd
iatra
chom
atis
911
894
98%
2.2
3%
0.5
6%
[3,2
5&&
]
Chl
amyd
iam
urid
arum
911
900
99%
NP
NP
Myc
obac
terium
tube
rcul
osis
C3
990
3899
98%
9.1
3%
0.2
6%
[33]
Fran
cise
llatu
lare
nsis
A1
933
1741
90%
2.7
6%
[11,2
3]
Cox
iella
burn
etiib
B2,0
65
2000
97%
1–2.5
%(Ig
G);
13.1
%(Ig
M)
0.6
–3.3
%(G
);5.1
%(M
)[1
5,3
8]
Borr
elia
burg
dorfer
i1
640
1293
79%
15.5
0%
7.9
7%
[10]
Burk
hold
eria
pseu
dom
alle
iB
5728
1205
21%
8.9
6%
4.0
6%
[6]
Lept
ospi
rain
terr
ogan
sb3
667
3359
92%
5.6
9%
1.0
7%
[39,4
0&&
]
Salm
onel
laen
terica
Typh
ibB
4318
4000
93%
7.3
1%
(GM
)3.4
%(G
M)
[41]
Orien
tiats
utsu
gam
ushi
C1
400
1400
100%
NP
NP
Rick
etts
iarick
etsi
iB
900
900
100%
NP
NP
Barto
nella
hens
elae
B1
493
1433
96%
7.3
3%
3.6
3%
[17]
Ente
ric
toxo
gen
icEs
cher
ichi
aco
liN
PN
P
Para
site
sC
lost
ridi
umdi
ffici
le4000
3297
82%
NP
NP
Plas
mod
ium
falc
ipar
um5
268
4320
82%
13
–21%
2.1
–3.9
%[5
,42,4
3&
]
Plas
mod
ium
viva
x5
300
2200
42%
8.0
4%
–[4
4,4
5&
]
Schi
stos
oma
man
soni
/jap
onic
umc
9000
300
3%
9.8
–44.7
%–
[4,4
6&
]
Toxo
plas
ma
gon
dii
8155
1015
12%
18%
[13]
Nec
ator
amer
ican
us12
000
564
5%
3.9
0%
[47
&&
]
Tryp
anos
oma
cruz
i15
099
240
2%
NP
NP
Hum
anTr
ypan
osom
abr
ucei
8529
214
3%
NP
NP
Hum
anA
utoi
mm
une
arra
yd21
000
1800
9%
2%
[16]
Tota
l1
21
312
>40
000
33%
EBV
,Ep
stei
n–Ba
rrvi
rus;
HIV
,hu
man
imm
unod
efic
ienc
yvi
rus;
HPV
,hu
man
papi
llom
avirus
;H
SV,
herp
essi
mpl
exvi
rus;
JE,
Japa
nese
ence
phal
itis;
SLE,
Sain
tLou
isen
ceph
aliti
s;V
ZV
,va
rice
llazo
ster
viru
s;W
NV
,W
estN
ilevi
rus;
YF,
yello
wfe
ver.
aN
onpu
blis
hed
data
incl
uded
.bBo
thIg
Gan
dIg
Mw
ere
incl
uded
inco
mpu
tatio
n.c IgG
and
IgE
subt
ype
resp
onse
sw
ere
mea
sure
d.dPe
mph
igus
vulg
aris
antib
odie
sw
ere
mea
sure
d.
Antibody prediction for infectious diseases Liang and Felgner
0951-7375 Copyright � 2015 Wolters Kluwer Health, Inc. All rights reserved. www.co-infectiousdiseases.com 441

Table 2. Physical properties and cellular functions associated with immunogenity in bacteria, virus, and parasites
Enriching features Bacteria Viruses Parasites
Predicted functions COG U Intracellular traffickingand secretion
Virion membrane GO function protein binding, catalytic,transporter, transferase activities
COG M-Cell envelope biogenesis,outer memberane
Membrane proteins,core proteins
GO function enzymatic, regulator
COG N-Cell motility andsecretion
GO function structural molecule and ionchannel activity
COG O Posttranslational modifi-cation, protein turnover, chaper-ones
GO ATP biosynthetic process, proteol-ysis; metabolic
COG S GO transport process
Predicted computationally TMHMM¼1 TMHMM TMHMM¼1 to 10
Signal P> ¼0.7 Signal peptide
pSortb Outer Membrane pSort Outer Membrane
pSortb Periplasmic
pI 0–5 pI 7–9
Expression Expression evidence by massspectrometry
Abundance of expression Abundance of expression
COG, Clusters of Orthologous Groups; GO, gene ontology; pI, isoelectric point; TMHMM, transmembrane domains.
Tropical and travel-associated diseases
30 million data points derived from 15 000 patientsera. Interrogation of these data sets has revealedmore than 10 proteomic features that are associatedwith antigenicity allowing an in-silico proteinsequence and functional annotation-based approachto triage the least likely antigenic proteins from thosethat are more likely to be antigenic.
These proteomic enrichment features (Table 2)are: functionally annotated Clusters of OrthologousGroups of proteins (U, M, N, and O) or gene ontol-ogy function and process; computationally pre-dicted features (TMHMM, Signal peptide, pSortOutermembrane, pSort Periplasmic, and isoelectricpoint (pI) less than 5 for bacteria, and pI 7–9 forparasites); and abundance of expression. Thisapproach applied to B. melitensis predicts 37% ofthe bacterial proteome containing 91% of the anti-gens empirically identified by probing proteomemicroarrays [12].
Parasite toxoplasma gondii proteins were assignedby gene ontology functions. Proteins involved inprotein binding, catalytic activity, transporteractivity, and transferase activity were significantlyenriched [13]. Proteins with enzymatic activityother than kinase activity were enriched at 2.0 fold,and proteins with enzyme regulator activity, struc-tural molecule activity, and ion channel activitywere also highly enriched. Proteins with gene ontol-ogy null functions, or involved in nucleotide andnucleic acid binding were underrepresented [13].
Proteins were also assigned by gene ontologyprocess classification. Proteins involved in ATP
Copyright © 2015 Wolters Kluwer
442 www.co-infectiousdiseases.com
biosynthetic process were enriched. Several proteinsinvolved in transport were also significantlyenriched, including ion transport, protein trans-port, vesicle mediated transport, and other transportfunctions. Proteins involved in metabolic process,proteolysis, and signal peptide processing were alsoenriched. Conversely, proteins not assigned withgene ontology process categories were significantlyunderrepresented (0.5 fold; P value 3.301�10�21)[13].
An examination with the Pf proteins on themicroarray based on gene ontological analysisrevealed that approximately 40% of the immuno-genic proteins are expressed in the membrane of theparasite or host erythrocyte and that they are over-represented in the biological process categories of‘pathogenesis,’ ‘cytoadherence to microvascula-ture,’ ‘antigenic variation,’ and ‘rosetting’ [5].
The data set of Vaccinia viral proteins alsoallowed us to identify properties of viral proteinsthat were associated with immunogenicity. Wefound that membrane and core proteins, proteinswith late or early/late temporal expression, andproteins with transmembrane domains were over-represented in the immunoreactive antigen setrelative to the whole proteome. These predictorsare strongest in MVA profiles, as the antibody profileto MVA is more heavily skewed toward structuralproteins. In contrast, early proteins were under-represented relative to the whole proteome, andthere was negligible influence of molecularweight, pI, or the presence of a signal sequence
Health, Inc. All rights reserved.
Volume 28 � Number 5 � October 2015

Table 3. Naive Bayses classification of Brucella melitensis and Salmonella enterica
Top ranked antigens
Brucella melitensis Salmonella enterica
% of totalseroreactive
% of totalserodiagnostic
% of totalseroreactive
% of totalserodiagnostic
1% 6% 15% 4% 5%
2% 11% 21% 4% 9%
5% 27% 45% 9% 25%
10% 44% 82% 19% 39%
20% 63% 91% 38% 64%
25% 72% 94% 45% 72%
50% 88% 100% 72% 86%
75% 94% 100% 100% 96%
100% 100% 100% 100% 100%
Antibody prediction for infectious diseases Liang and Felgner
on immunogenicity. Vaccinia antigens are eitherabundant components of MV particles, such asA10 and L4 [48], or are expressed at high levels ininfected cells, such as I1 and WR148 [49,50]. Theirabundance may contribute to immunogenicityonce released from infected cells, particularly if, likeD13 [51], such proteins have a propensity for self-assembly into macromolecular structures.
Analysis was also done for the herpes simplexvirus-1 antibody profile based on gene ontologycomponent classifiers according to the database atwww.uniprot.org. The percentage of the total num-ber of genes assigned to each gene ontology com-ponent present in the proteome and in theseroreactive antigens was determined, and the ratiowas used to determine the fold enrichment. Theanalysis revealed 12 proteins on the array that wereassigned the gene ontology component virion mem-brane, of which nine were seroreactive. Tegumentproteins were not enriched in the seroreactive anti-gen set [37].
Overall, our data show that the antibody profileis not a random assortment of specificities, butstrongly biased toward the recognition of certainproteomic features. Why we do not observe anti-bodies to all intracellular proteins expressed frominfected cells remains unclear. It is also interesting,to note, that the rules that determine immunoge-nicity might be different from those that defineprotection.
NAIVE BAYES CLASSIFICATION
Individual proteomic features provide some infor-mation about the likelihood of a protein beingseroreactive; however, using all of these featurestogether leads to a better segregation of the hitsfrom the rest of the proteome. To analyze the
Copyright © 2015 Wolters Kluwe
0951-7375 Copyright � 2015 Wolters Kluwer Health, Inc. All rights rese
relationship between all of these features and theseroreactivity of the proteins in a rigorous manner,we used a naive Bayes formulation [52].
We applied a naive Bayes classification approachto assign a relative numerical score to each antigenin the B. melitensis (Bm) proteome. This score reflectsthe relative likelihood that a protein will be reactivebased on its functionally annotated or computation-ally predicted features. Our analyses indicates that91% of serodiagnostic antigens are predictable fromthe top 20% of the genome ranked by this naiveBayes classification approach, and the antigens withenriched features in the top 20% of the genomeaccount for 100% of serodiagnostic antigens withthese features. Without this naive Bayes classifi-cation approach, we would have to clone 37% ofthe genome with enriching features to obtain 91%of serodiagnostic antigens. This analysis greatlyenhances the predictive efficiency compared withprevious studies, will provide a basis for targetedscreens of entire proteomes based on likelihoodof seroreactivity, and help determine trends in thehumoral immune response to gram-negativebacteria. The same approach has been applied toS. enterica and revealed that we would need toscreen only 25% of the genome to be able to identify72% of serodiagnostic antigens (Table 3).
CONCLUSION
The development of protein arrays for profilingthe antibody response generated upon exposureto an infectious agent has allowed for new insightinto the humoral immune response and the identi-fication of potential subunit vaccine candidatesand new diagnostics. No other existing approachcan provide such a thorough perspective of thehumoral immune response to infection. Moreover,
r Health, Inc. All rights reserved.
rved. www.co-infectiousdiseases.com 443

Tropical and travel-associated diseases
it provides a systematic foundation formation onproteomic features (functional and physically pro-perties) of seroreactive and serodiagnostic antigens.The information presented here will allow futureprotein microarray screening to focus efforts onportions of the proteome that most likely containseroreactive proteins, and may also be useful forunderstanding the antibody responses to bacteria,viruses, and parasites.
Acknowledgements
None.
Financial support and sponsorship
This work was supported by NIH grants U01AI078213,AI089686 and AI095916.
Conflicts of interest
There are no conflicts of interest.
REFERENCES AND RECOMMENDEDREADINGPapers of particular interest, published within the annual period of review, havebeen highlighted as:
& of special interest&& of outstanding interest1. Mayers C, Duffield M, Rowe S, et al. Analysis of known bacterial proteinvaccine antigens reveals biased physical properties and amino acid composi-tion. Comp Funct Genomics 2003; 4:468–478.
2. Rappuoli R. Reverse vaccinology, a genome-based approach to vaccinedevelopment. Vaccine 2001; 19:2688–2691.
3. Molina DM, Pal S, Kayala MA, et al. Identification of immunodominant antigensof Chlamydia trachomatis using proteome microarrays. Vaccine 2010;28:3014–3024.
4. Driguez P, Doolan DL, Loukas A, et al. Schistosomiasis vaccine discoveryusing immunomics. Parasit Vectors 2010; 3:4.
5. Crompton PD, Kayala MA, Traore B, et al. A prospective analysis of the Abresponse to Plasmodium falciparum before and after a malaria season byprotein microarray. Proc Natl Acad Sci U S A 2010; 107:6958–6963.
6. Felgner PL, Kayala MA, Vigil A, et al. A Burkholderia pseudomallei proteinmicroarray reveals serodiagnostic and cross-reactive antigens. Proc NatlAcad Sci U S A 2009; 106:13499–13504.
7. Chen C, Bouman TJ, Beare PA, et al. A systematic approach to evaluatehumoral and cellular immune responses to Coxiella burnetii immunoreactiveantigens. Clin Microbiol Infect 2009; 15 (Suppl 2):156–157.
8. Doolan DL, Mu Y, Unal B, et al. Profiling humoral immune responses to P.falciparum infection with protein microarrays. Proteomics 2008; 8:4680–4694.
9. Davies DH, Wyatt LS, Newman FK, et al. Antibody profiling by proteomemicroarray reveals the immunogenicity of the attenuated smallpox vaccinemodified vaccinia virus ankara is comparable to that of Dryvax. J Virol 2008;82:652–663.
10. Barbour AG, Jasinskas A, Kayala MA, et al. A genome-wide proteome arrayreveals a limited set of immunogens in natural infections of humans and white-footed mice with Borrelia burgdorferi. Infect Immun 2008; 76:3374–3389.
11. Sundaresh S, Randall A, Unal B, et al. From protein microarrays to diagnosticantigen discovery: a study of the pathogen Francisella tularensis. Bioinfor-matics 2007; 23:i508–i518.
12. Liang L, Tan X, Juarez S, et al. Systems biology approach predicts antibodysignature associated with brucella melitensis infection in humans. J ProteomeRes 2011; 10:4813–4824.
13. Liang L, Doskaya M, Juarez S, et al. Identification of potential serodiagnosticand subunit vaccine antigens by antibody profiling of toxoplasmosis cases inTurkey. Mol Cell Proteomics 2011; 10:006916; M110.
14. Liang L, Leng D, Burk C, et al. Large scale immune profiling of infectedhumans and goats reveals differential recognition of Brucella melitensisantigens. PLoS Negl Trop Dis 2010; 4:e673.
15. Vigil A, Chen C, Jain A, et al. Profiling the humoral immune response of acuteand chronic Q fever by protein microarray. Mol Cell Proteomics 2011; 10(M110):006304.
Copyright © 2015 Wolters Kluwer
444 www.co-infectiousdiseases.com
16. Kalantari-Dehaghi M, Molina DM, Farhadieh M, et al. New targets of pem-phigus vulgaris antibodies identified by protein array technology. Exp Derma-tol 2011; 20:154–156.
17. Vigil A, Ortega R, Jain A, et al. Identification of the feline humoral immuneresponse to Bartonella henselae infection by protein microarray. PLoS One2010; 5:e11447.
18. Yang ZR, Lertmemongkolchai G, Tan G, et al. A genetic programmingapproach for Burkholderia pseudomallei diagnostic pattern discovery. Bioin-formatics 2009; 25:2256–2262.
19. Tippayawat P, Saenwongsa W, Mahawantung J, et al. Phenotypic andfunctional characterization of human memory T cell responses to Burkholderiapseudomallei. PLoS Negl Trop Dis 2009; 3:e407.
20. Cannella AP, Lin JC, Liang L, et al. Serial kinetics of the antibody responseagainst the complete brucella melitensis orfeome in focal vertebral brucello-sis. J Clin Microbiol 2012; 50:922–926.
21. Tan X, Traore B, Kayentao K, et al. Hemoglobin S and C heterozygosityenhances neither the magnitude nor breadth of antibody responses to adiverse array of Plasmodium falciparum antigens. J Infect Dis 2011;204:1750–1761.
22. Barry AE, Trieu A, Fowkes FJ, et al. The stability and complexity of antibodyresponses to the major surface antigen of Plasmodium falciparum are asso-ciated with age in a malaria endemic area. Mol Cell Proteomics 2011;10:008326; M111.
23. Eyles JE, Unal B, Hartley MG, et al. Immunodominant Francisella tularensisantigens identified using proteome microarray. Proteomics 2007; 7:2172–2183.
24. Sundaresh S, Doolan DL, Hirst S, et al. Identification of humoral immuneresponses in protein microarrays using DNA microarray data analysis tech-niques. Bioinformatics 2006; 22:1760–1766.
25.&&
Patton DL, Teng A, Randall A, et al. Whole genome identification of C.trachomatis immunodominant antigens after genital tract infections and effectof antibiotic treatment of pigtailed macaques. J Proteomics 2014; 108:99–109.
This is the first time that Chlamydia trachomatis immunodominant andpotential vaccine antigens have been identified in nonhuman primates followinginfection.26. Beare PA, Chen C, Bouman T, et al. Candidate antigens for Q fever
serodiagnosis revealed by immunoscreening of a Coxiella burnetii proteinmicroarray. Clin Vaccine Immunol 2008; 15:1771–1779.
27. Doskaya M, Kalantari-Dehaghi M, Walsh CM, et al. GRA1 protein vaccineconfers better immune response compared to codon-optimized GRA1 DNAvaccine. Vaccine 2007; 25:1824–1837.
28. Mochon AB, Jin Y, Kayala MA, et al. Serological profiling of a Candidaalbicans protein microarray reveals permanent host-pathogen interplay andstage-specific responses during candidemia. PLoS Pathog 2010;6:e1000827.
29. Davies DH, Liang X, Hernandez JE, et al. Profiling the humoral immuneresponse to infection by using proteome microarrays: high-throughput vac-cine and diagnostic antigen discovery. Proc Natl Acad Sci U S A 2005;102:547–552.
30. Davies DH, McCausland MM, Valdez C, et al. Vaccinia virus H3Lenvelope protein is a major target of neutralizing antibodies in humansand elicits protection against lethal challenge in mice. J Virol 2005;79:11724–11733.
31. Davies DH, Molina DM, Wrammert J, et al. Proteome-wide analysis of theserological response to vaccinia and smallpox. Proteomics 2007; 7:1678–1686.
32. Luevano M, Bernard HU, Barrera-Saldana HA, et al. High-throughput profilingof the humoral immune responses against thirteen human papillomavirustypes by proteome microarrays. Virology 2010; 405:31–40.
33. Kunnath-Velayudhan S, Salamon H, Wang HY, et al. Dynamic antibodyresponses to the Mycobacterium tuberculosis proteome. Proc Natl AcadSci U S A 2010; 107:14703–14708.
34. Sboner A, Karpikov A, Chen G, et al. Robust-linear-model normalization toreduce technical variability in functional protein microarrays. J Proteome Res2009; 8:5451–5464.
35. Huber W, von Heydebreck A, Sultmann H, et al. Variance stabilization appliedto microarray data calibration and to the quantification of differential expres-sion. Bioinformatics 2002; 18 (Suppl 1):S96–S104.
36.&
Gerns Storey HL, Richardson BA, Singa B, et al. Use of principal componentsanalysis and protein microarray to explore the association of HIV-1-specificIgG responses with disease progression. AIDS Res Hum Retroviruses 2014;30:37–44.
A collection of HIV-specific antibody responses that together were associated withreduced disease progression, by PCA and microarray analyses.37. Kalantari-Dehaghi M, Chun S, Chentoufi AA, et al. Discovery of potential
diagnostic and vaccine antigens in herpes simplex virus 1 and 2 by proteome-wide antibody profiling. J Virol 2012; 86:4328–4339.
38. Vigil A, Ortega R, Nakajima-Sasaki R, et al. Genome-wide profiling of humoralimmune response to Coxiella burnetii infection by protein microarray. Pro-teomics 2010; 10:2259–2269.
39. Lessa-Aquino C, Borges Rodrigues C, Pablo J, et al. Identification of seror-eactive proteins of Leptospira interrogans serovar copenhageni using a high-density protein microarray approach. PLoS Negl Trop Dis 2013; 7:e2499.
Health, Inc. All rights reserved.
Volume 28 � Number 5 � October 2015

Antibody prediction for infectious diseases Liang and Felgner
40.&&
Lessa-Aquino C, Wunder EA Jr, Lindow JC, et al. Proteomic featurespredict seroreactivity against leptospiral antigens in leptospirosis patients.J Proteome Res 2015; 14:549–556.
This is the first full proteome Leptospira interrogans microarray that was con-structed for identifying serodiagnostic antigens, and it also provides an empiricalbasis for predicting antigenicity from Gram-negative bacteria.41. Liang L, Juarez S, Nga TV, et al. Immune profiling with a Salmonella Typhi
antigen microarray identifies new diagnostic biomarkers of human typhoid. SciRep 2013; 3:1043.
42. Baum E, Badu K, Molina DM, et al. Protein microarray analysis of antibodyresponses to Plasmodium falciparum in western Kenyan highland sites withdiffering transmission levels. PLoS One 2013; 8:e82246.
43.&
Campo JJ, Aponte JJ, Skinner J, et al. RTS,S vaccination is associated withserologic evidence of decreased exposure to Plasmodium falciparum liver-and blood-stage parasites. Mol Cell Proteomics 2015; 14:519–531.
These microarray data provide insight into the mechanism by which RTS,S vaccineprotects from Malaria.44. Molina DM, Finney OC, Arevalo-Herrera M, et al. Plasmodium vivax preery-
throcytic-stage antigen discovery: exploiting naturally acquired humoral re-sponses. Am J Trop Med Hyg 2012; 87:460–469.
45.&
Baum E, Sattabongkot J, Sirichaisinthop J, et al. Submicroscopic andasymptomatic Plasmodium falciparum and Plasmodium vivax infections arecommon in western Thailand: molecular and serological evidence. Malar J2015; 14:95.
These microarray data are empirical data for understanding prevalance of Plas-modium in western Thailand.
Copyright © 2015 Wolters Kluwe
0951-7375 Copyright � 2015 Wolters Kluwer Health, Inc. All rights rese
46.&
Gaze S, Driguez P, Pearson MS, et al. An immunomics approach to schisto-some antigen discovery: antibody signatures of naturally resistant and chroni-cally infected individuals from endemic areas. PLoS Pathog 2014;10:e1004033.
Several potentially protective and well tolerated schistosomiasis vaccine antigenswere identified.47.&&
Tang YT, Gao X, Rosa BA, et al. Genome of the human hookworm Necatoramericanus. Nat Genet 2014; 46:261–269.
The authors report sequencing and assembly of the N. americanus genome, andprovide an invaluable resource to boost ongoing efforts toward fundamental andapplied postgenomic research.48. Chung CS, Chen CH, Ho MY, et al. Vaccinia virus proteome: identification of
proteins in vaccinia virus intracellular mature virion particles. J Virol 2006;80:2127–2140.
49. Patel DD, Pickup DJ, Joklik WK. Isolation of cowpox virus A-type inclusionsand characterization of their major protein component. Virology 1986;149:174–189.
50. Liu X, Kremer M, Broyles SS. A natural vaccinia virus promoter with excep-tional capacity to direct protein synthesis. J Virol Methods 2004; 122:141–145.
51. Szajner P, Weisberg AS, Lebowitz J, et al. External scaffold of sphericalimmature poxvirus particles is made of protein trimers, forming a honeycomblattice. J Cell Biol 2005; 170:971–981.
52. Witten IH, Frank E. Data mining: practical machine learning toolsand techniques. 2nd ed. Amsterdam: San Francisco Morgan Kaufman;2005.
r Health, Inc. All rights reserved.
rved. www.co-infectiousdiseases.com 445