Embed Size (px)
Citation preview

ORIGINAL PAPER
Coumarin interacts with auxin polar transport to modify rootsystem architecture in Arabidopsis thaliana
Antonio Lupini • Fabrizio Araniti • Francesco Sunseri •
Maria Rosa Abenavoli
Received: 24 September 2013 / Accepted: 17 January 2014
� Springer Science+Business Media Dordrecht 2014
Abstract Coumarin is a highly active allelopathic com-
pound which plays a key role in plant–plant interactions
and communications. It affects root growth and develop-
ment of many species, but its mode of action has not been
clarified yet. It has been hypothesized that auxin could
mediate coumarin-induced effects on root system. Through
morphological and pharmacological approaches together
with the use of Arabidopsis auxin mutants, a possible
interaction between coumarin and auxin in driving root
system development has been investigated in Arabidopsis
thaliana (Col-0). Coumarin strongly affected primary root
elongation and lateral root development of Arabidopsis
seedlings. In particular, 10-4 M coumarin significantly
inhibited primary root elongation increasing lateral root
number and root hairs length. Further, coumarin addition
was able to restore the negative effects of TIBA and NPA,
two auxin transport inhibitors, which caused a complete
inhibition of lateral root formation. Arabidopsis auxin
mutants differently responded to coumarin compared to
wild type (Col-0). In particular, lax3 mutant showed the
lowest (42 %) inhibition of primary root length, whereas,
eir1-4 mutant had higher inhibition (53 %) compared to
Col-0; conversely, aux1-22 mutant did not show any effect
in response to coumarin. An increase of lateral root number
was observed in pin1 mutant only. Finally, coumarin
increased the root hairs length in eir1-4, lax3, pin1 and
pin3-5 mutants, but not in aux1-22. These results suggested
a functional interaction between coumarin and auxin polar
transport in driving root development in A. thaliana.
Keywords Arabidopsis thaliana � Auxin � Coumarin �Mutants � Root morphology
Introduction
Exploration and exploitation of soil resources such as
water, mineral nutrients by plants is closely associated with
root development (Schlichting 1986) and in particular with
its architecture (root system architecture, RSA), which is
an integrative result of lateral root initiation, morphogen-
esis, emergence and growth (Dubrovsky and Forde 2012)
as well as root hair formation and elongation (Nibau et al.
2008). However, root system is extremely plastic organ,
able to adapt in response to external cues such as nutrients
(Lopez-Bucio et al. 2003), soil moisture and matrix (Hodge
2006) and allelopathic compounds (Whitehead et al. 1982;
Rice 1984). Coumarin is an active allelochemicals widely
distributed in plants kingdom which plays an important
role in plant–plant interaction and communication (Zobel
and Brown 1995). Released into the environment, couma-
rin affected plant growth and development of many spe-
cies, and especially their root system, one of the main
target of allelochemicals (Rice 1984). The negative effects
of coumarin on cell division and root polarity have been
known for a long time (Goodwin and Avers 1950; Avers
and Goodwin 1956), although Neumann (1959) demon-
strated that coumarin markedly stimulated the elongation
of excised segments of Helianthus hypocotyls, comparing
its action to that of the auxin. Coumarin influenced root
morphology and histology (Svensson 1971, 1972; Ku-
pidlowska et al. 1994; Abenavoli et al. 2001), showing a
A. Lupini � F. Araniti � F. Sunseri � M. R. Abenavoli (&)
Dipartimento Agraria, Universita Mediterranea di Reggio
Calabria, Salita Melissari, 89124 Reggio Calabria, RC, Italy
e-mail: [email protected]
123
Plant Growth Regul
DOI 10.1007/s10725-014-9893-0

selective effect on maize and Arabidopsis root types,
inhibiting the primary root elongation and stimulating lat-
eral root formation (Abenavoli et al. 2004, 2008). More
recently, Lupini et al. (2010), by morphological and elec-
trophysiological approaches, indicated the root zone within
20 mm from tip of maize primary root as the most sensitive
to coumarin, suggesting that this effect could be mediated
by auxin. However, the exact mechanism of coumarin on
root growth has not been clarified yet. A complex network
of molecular signaling probably governs coumarin mor-
pho-physiological responses, where auxin transport and/or
biosynthesis could play an important role.
Auxin is considered to be the prime candidate to control
stress-induced morphogenic response ‘‘SIMR’’, character-
ized by inhibition of root elongation and enhanced formation
of lateral roots (Potters et al. 2007). It plays a central role in
organ development and elongation, in shoot/root branching
and plastic growth responses (Zazımalova et al. 2010), in
lateral root initiation (Casimiro et al. 2003; De Smet et al.
2006), primordial (Benkova et al. 2003), and emergence
(Laskowski et al. 2006). Transported in polarized streams
(Polar Auxin Transport, PAT), via auxin transporters, auxin
action depends on its differential distribution (Dubrovsky
et al. 2011; Benkova et al. 2009). Moreover, many evidences
established that PAT is mediated by AUX1/LAX uptake PM
permeases, ATP Binding Cassette subfamily B (ABCB)
transporters, and PIN-FORMED (PIN) carrier proteins
(Petrasek and Friml 2009; Zazımalova et al. 2010). In par-
ticular, in Arabidopsis thaliana, the AUX1/LAX permeases
family encode one AUX1, and three Like AUX1 (LAX1, 2
and 3) (Parry et al. 2001), which control rapid and active
auxin influx in cells. On the other hand, eight members of the
AtPIN family (from 1 to 8) have been identified: PIN5, PIN6
and PIN8, localized to endomembranes, regulate the distri-
bution of cellular auxin homeostasis (Mravec et al. 2009),
and PIN1, PIN2, PIN3, PIN4 and PIN7, localized to plasma
membrane (PM), perform cellular auxin efflux (Mravec et al.
2008).
Recently, Li et al. (2011) demonstrated that 4-methyl-
umbelliferone, a coumarin derivative, increased the
expression of two auxin efflux facilitator genes (PIN2 and
PIN3) in Arabidopsis roots, suggesting that auxin redistri-
bution may directly or indirectly mediate 4-methy-
lumbelliferone-induced root branching.
In this respect, the aim of this work was to investigate,
through morphological and pharmacological approaches
together with the use of Arabidopsis auxin mutants, a
possible interaction between coumarin and auxin in driving
root system development, in A. thaliana. In particular,
auxin transport inhibitors, such as NPA and TIBA, and
several AUX1/LAX and PIN Arabidopsis mutants involved
in active auxin transport/redistribution were employed to
confirm this hypothesis.
Materials and methods
Plant materials and growth conditions
Seeds of A. thaliana (Col-0) and aux1-22, lax3, pin1, eir1-4,
pin3-5 mutants were soaked in distilled water for 30 min,
surface sterilized with 95 % (v/v) ethanol and 5 % (v/v)
commercial bleach for 5 min, rinsed 5 times with sterile
water and then placed to 4 �C for 2 days. Sterilized seeds
were plated (Petri dishes, 120 9 120 mm) on solidified-agar
medium [0.75 % (w/v)] plus sucrose [0.5 % (w/v)], MES
(1 g/L), pH 5.75. They were vertically placed in a growth
chamber (22 �C, 65 % RH, 16/8 h, 300 lmol photon flux
density m-2 s-1) (Lupini et al. 2013). Five uniform seed-
lings, 4-day old, were then transferred to a single plate and
grown with same medium containing different coumarin
(MW 146.15, C99 % purity, Sigma Aldrich) and/or IAA
concentrations ranged from 10-15 to 10-4 M for 7 days.
Furthermore, to determine the effective role of auxin in
driving coumarin-effect on root system, a pharmacological
approach using 1.5 9 10-5 M 2,3,5-triiodobenzoic acid
(TIBA) and 5 9 10-6 M naphthylphthalamic acid (NPA)
(Casimiro et al. 2003) was employed.
Root measurements
Roots image was captured by scanning (STD 1600, Regent
Instruments Inc., Quebec, Canada). Briefly, after 7-day of
treatment the Petri dish containing Arabidopsis seedlings
were positioned on the scanner and a image was captured at
1,200 dots per inch (dpi) of resolution. The root length was
measured using WinRhizo Pro system v. 2002a software
(Instruments Regent Inc., Quebec, Canada) and lateral
roots number was counted manually from the image (Ab-
enavoli et al. 2008). According to Dubrovsky and Forde
(2012), to determine lateral root density, the primary root
was divided in two zones: root branching zone that extends
rootward from the shoot base to the last emerged lateral
root, and lateral root formation zone that spreads rootward
from the end of the root branching zone to last lateral root
primordium. Lateral root density has been expressed as
branching density, defined as the number of lateral roots
per unit length of the root branching zone.
Root hair length (lm) and density (root hair number
mm-2 of primary root) were determined by using a ste-
reoscopic microscopy (Olympus SZX9) and Image Tool v.
3 software (UTHSC, San Antonio, USA).
Mitotic sites determination
Number of mitotic sites was evaluated as described by
Canellas et al. (2002) with some modifications. Briefly,
Arabidopsis seedlings were harvested after 7-day of
Plant Growth Regul
123
treatment and their roots were washed in 50 mM phosphate
buffer (pH 7.4), then transferred in 0.5 % KOH (w/v) and
cooled at 75� C for 20 min. The roots were rinsed once
again in 50 mM phosphate buffer (pH 7.4) and then stained
for 16 h in the dark in haematoxylin solution, containing
0.025 g haematoxylin, 0.0125 g ferric ammonium sulphate
and 1.5 mL of 45 % (v/v) acetic acid. Finally, the roots
were distained in 80 % lactic acid (w/v) at 75 �C for
10–20 s, and the mitotic sites were counted by stereoscopic
microscopy (Olympus SZX9).
Statistical analysis
All experiments were laid out in a completely randomised
design with at least fifteen replications for each. The data
were analyzed by one-way ANOVA comparing within and
among treatments, and means were separated by Tukey’s
Honestly Significant Difference (HSD) test (p B 0.05)
using Systat software (Systat Software Inc, Chicago, USA).
In the mutant experiments, the data were analyzed by
Student’s unpaired t test comparing between treated and
untreated roots.
Results
To assess coumarin responses on Arabidopsis root system,
a coumarin dose response of Arabidopsis seedlings (4 day-
old) of primary root elongation and lateral root number was
performed. Coumarin slightly increased primary root (PR)
length up to 10-6 M, whereas, the highest concentration
(10-4 M) significantly inhibited it (Fig. 1). In addition, at
lowest concentrations, coumarin weakly affected lateral
roots number (LR), which was instead significantly
increased (85.3 % respect to control), at the highest one
(10-4 M) (Fig. 1). Based on these preliminary results,
10-4 M coumarin was adopted in the subsequent experi-
ments as the more effective and interesting dose driving
root modifications.
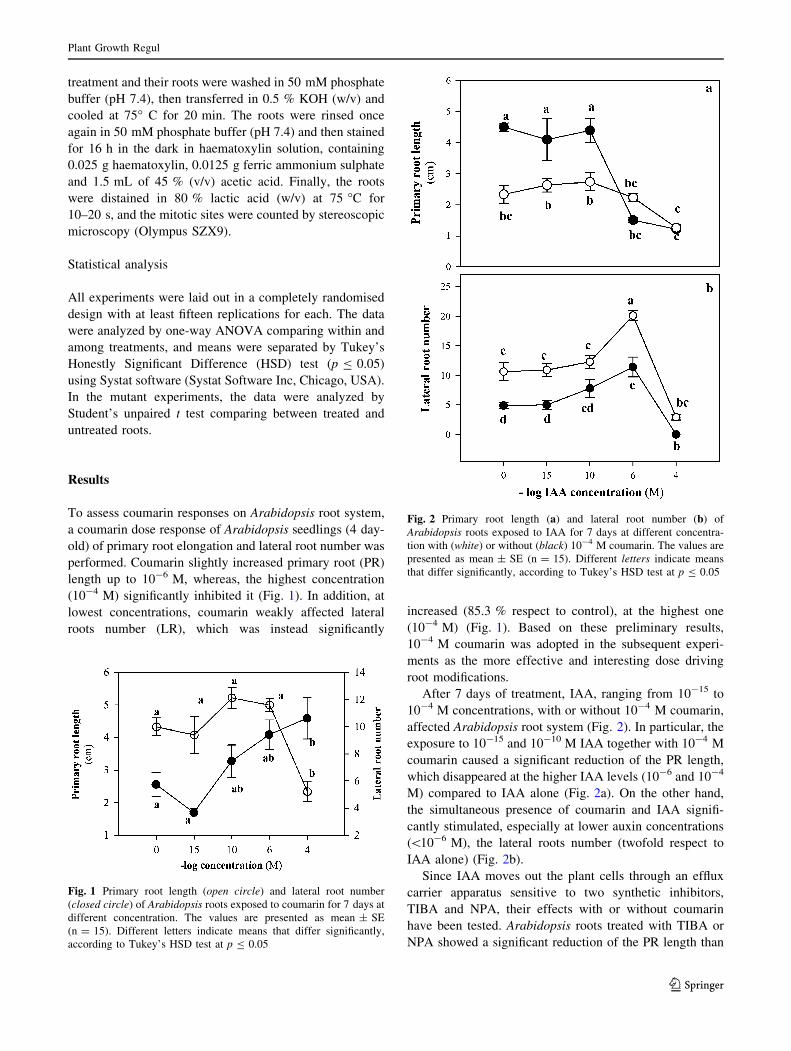
After 7 days of treatment, IAA, ranging from 10-15 to
10-4 M concentrations, with or without 10-4 M coumarin,
affected Arabidopsis root system (Fig. 2). In particular, the
exposure to 10-15 and 10-10 M IAA together with 10-4 M
coumarin caused a significant reduction of the PR length,
which disappeared at the higher IAA levels (10-6 and 10-4
M) compared to IAA alone (Fig. 2a). On the other hand,
the simultaneous presence of coumarin and IAA signifi-
cantly stimulated, especially at lower auxin concentrations
(\10-6 M), the lateral roots number (twofold respect to
IAA alone) (Fig. 2b).
Since IAA moves out the plant cells through an efflux
carrier apparatus sensitive to two synthetic inhibitors,
TIBA and NPA, their effects with or without coumarin
have been tested. Arabidopsis roots treated with TIBA or
NPA showed a significant reduction of the PR length than
Fig. 1 Primary root length (open circle) and lateral root number
(closed circle) of Arabidopsis roots exposed to coumarin for 7 days at
different concentration. The values are presented as mean ± SE
(n = 15). Different letters indicate means that differ significantly,
according to Tukey’s HSD test at p B 0.05
Fig. 2 Primary root length (a) and lateral root number (b) of
Arabidopsis roots exposed to IAA for 7 days at different concentra-
tion with (white) or without (black) 10-4 M coumarin. The values are
presented as mean ± SE (n = 15). Different letters indicate means
that differ significantly, according to Tukey’s HSD test at p B 0.05
Plant Growth Regul
123
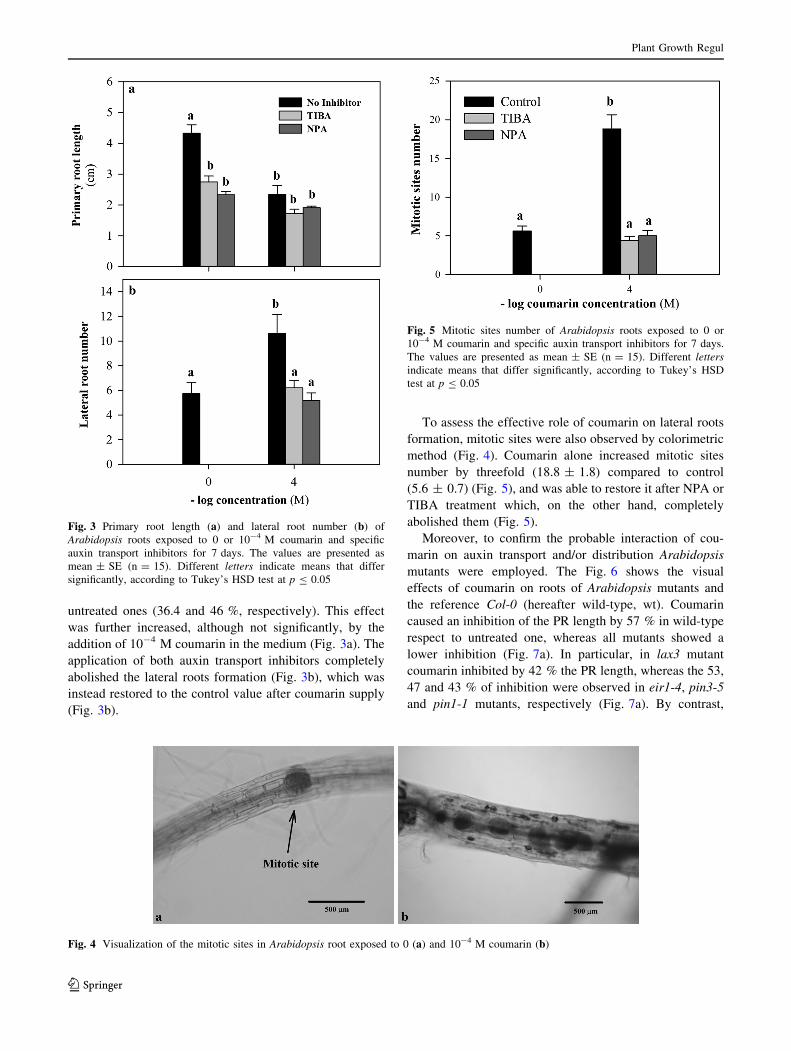
untreated ones (36.4 and 46 %, respectively). This effect
was further increased, although not significantly, by the
addition of 10-4 M coumarin in the medium (Fig. 3a). The
application of both auxin transport inhibitors completely
abolished the lateral roots formation (Fig. 3b), which was
instead restored to the control value after coumarin supply
(Fig. 3b).
To assess the effective role of coumarin on lateral roots
formation, mitotic sites were also observed by colorimetric
method (Fig. 4). Coumarin alone increased mitotic sites
number by threefold (18.8 ± 1.8) compared to control
(5.6 ± 0.7) (Fig. 5), and was able to restore it after NPA or
TIBA treatment which, on the other hand, completely
abolished them (Fig. 5).
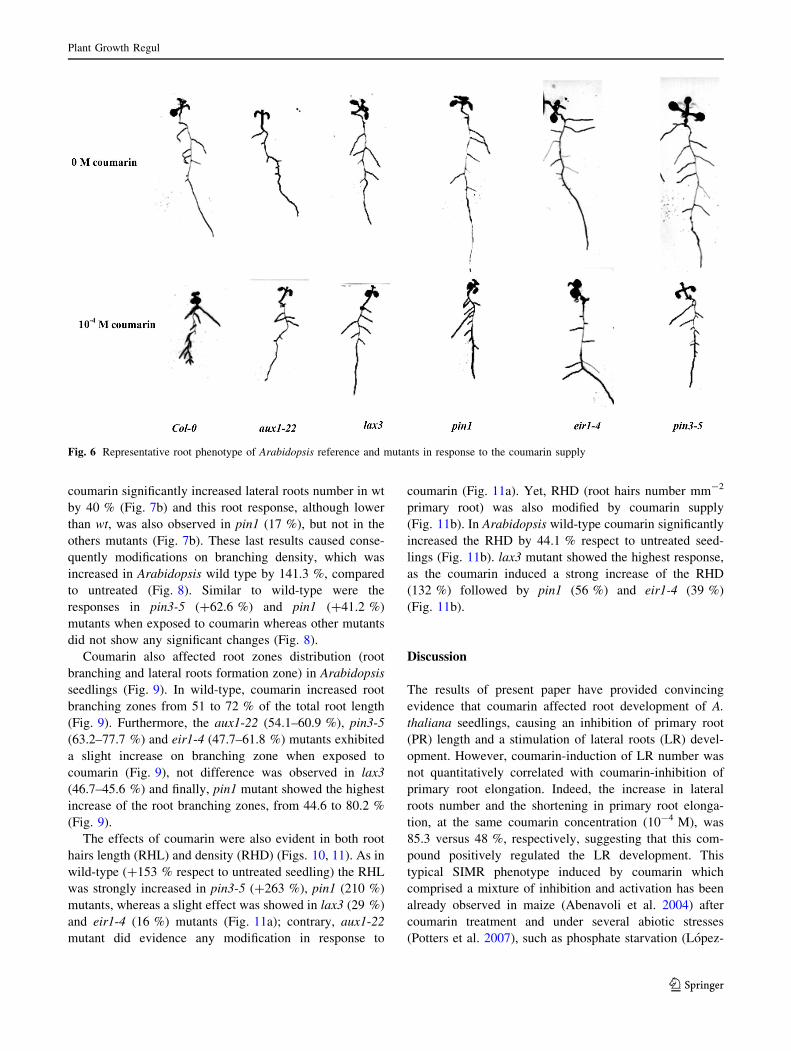
Moreover, to confirm the probable interaction of cou-
marin on auxin transport and/or distribution Arabidopsis
mutants were employed. The Fig. 6 shows the visual
effects of coumarin on roots of Arabidopsis mutants and
the reference Col-0 (hereafter wild-type, wt). Coumarin
caused an inhibition of the PR length by 57 % in wild-type
respect to untreated one, whereas all mutants showed a
lower inhibition (Fig. 7a). In particular, in lax3 mutant
coumarin inhibited by 42 % the PR length, whereas the 53,
47 and 43 % of inhibition were observed in eir1-4, pin3-5
and pin1-1 mutants, respectively (Fig. 7a). By contrast,
Fig. 3 Primary root length (a) and lateral root number (b) of
Arabidopsis roots exposed to 0 or 10-4 M coumarin and specific
auxin transport inhibitors for 7 days. The values are presented as
mean ± SE (n = 15). Different letters indicate means that differ
significantly, according to Tukey’s HSD test at p B 0.05
Fig. 4 Visualization of the mitotic sites in Arabidopsis root exposed to 0 (a) and 10-4 M coumarin (b)
Fig. 5 Mitotic sites number of Arabidopsis roots exposed to 0 or
10-4 M coumarin and specific auxin transport inhibitors for 7 days.
The values are presented as mean ± SE (n = 15). Different letters
indicate means that differ significantly, according to Tukey’s HSD
test at p B 0.05
Plant Growth Regul
123
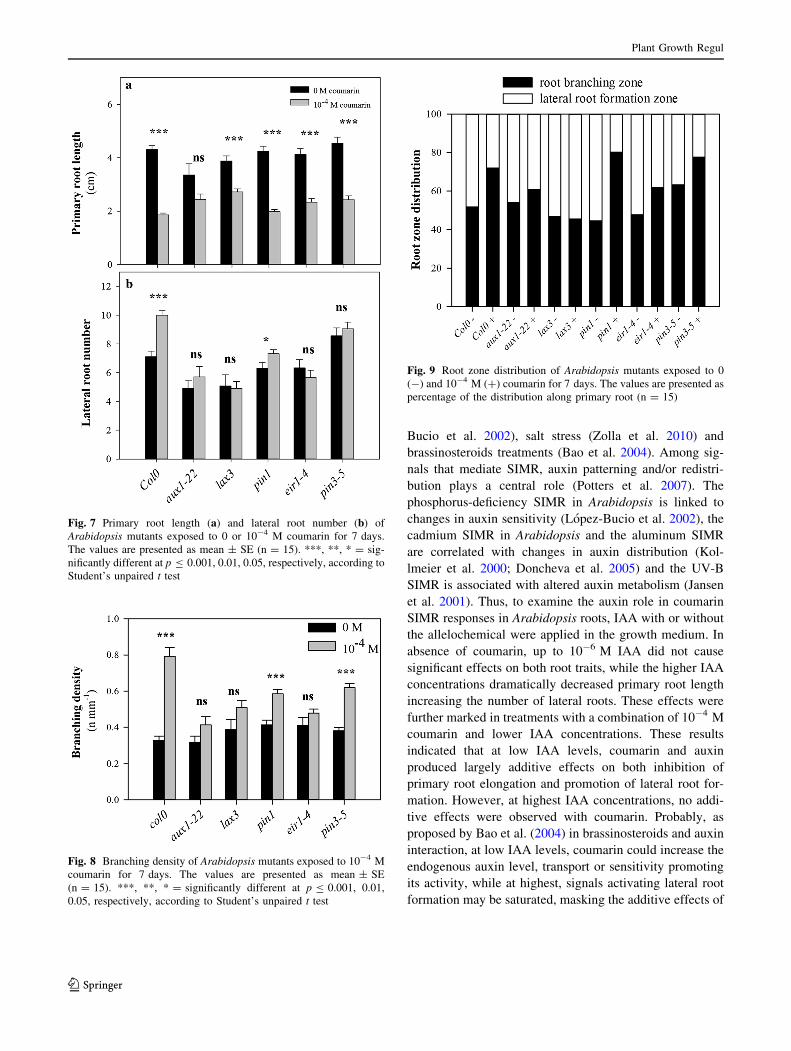
coumarin significantly increased lateral roots number in wt
by 40 % (Fig. 7b) and this root response, although lower
than wt, was also observed in pin1 (17 %), but not in the
others mutants (Fig. 7b). These last results caused conse-
quently modifications on branching density, which was
increased in Arabidopsis wild type by 141.3 %, compared
to untreated (Fig. 8). Similar to wild-type were the
responses in pin3-5 (?62.6 %) and pin1 (?41.2 %)
mutants when exposed to coumarin whereas other mutants
did not show any significant changes (Fig. 8).
Coumarin also affected root zones distribution (root
branching and lateral roots formation zone) in Arabidopsis
seedlings (Fig. 9). In wild-type, coumarin increased root
branching zones from 51 to 72 % of the total root length
(Fig. 9). Furthermore, the aux1-22 (54.1–60.9 %), pin3-5
(63.2–77.7 %) and eir1-4 (47.7–61.8 %) mutants exhibited
a slight increase on branching zone when exposed to
coumarin (Fig. 9), not difference was observed in lax3
(46.7–45.6 %) and finally, pin1 mutant showed the highest
increase of the root branching zones, from 44.6 to 80.2 %
(Fig. 9).
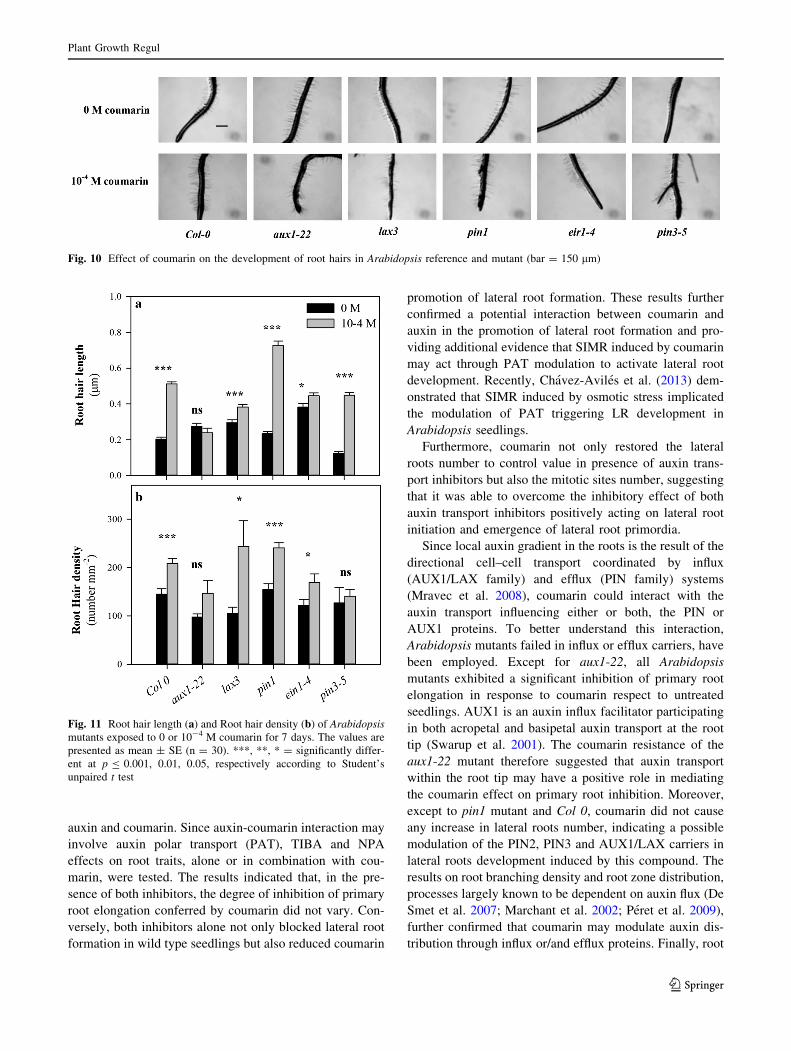
The effects of coumarin were also evident in both root
hairs length (RHL) and density (RHD) (Figs. 10, 11). As in
wild-type (?153 % respect to untreated seedling) the RHL
was strongly increased in pin3-5 (?263 %), pin1 (210 %)
mutants, whereas a slight effect was showed in lax3 (29 %)
and eir1-4 (16 %) mutants (Fig. 11a); contrary, aux1-22
mutant did evidence any modification in response to
coumarin (Fig. 11a). Yet, RHD (root hairs number mm-2
primary root) was also modified by coumarin supply
(Fig. 11b). In Arabidopsis wild-type coumarin significantly
increased the RHD by 44.1 % respect to untreated seed-
lings (Fig. 11b). lax3 mutant showed the highest response,
as the coumarin induced a strong increase of the RHD
(132 %) followed by pin1 (56 %) and eir1-4 (39 %)
(Fig. 11b).
Discussion
The results of present paper have provided convincing
evidence that coumarin affected root development of A.
thaliana seedlings, causing an inhibition of primary root
(PR) length and a stimulation of lateral roots (LR) devel-
opment. However, coumarin-induction of LR number was
not quantitatively correlated with coumarin-inhibition of
primary root elongation. Indeed, the increase in lateral
roots number and the shortening in primary root elonga-
tion, at the same coumarin concentration (10-4 M), was
85.3 versus 48 %, respectively, suggesting that this com-
pound positively regulated the LR development. This
typical SIMR phenotype induced by coumarin which
comprised a mixture of inhibition and activation has been
already observed in maize (Abenavoli et al. 2004) after
coumarin treatment and under several abiotic stresses
(Potters et al. 2007), such as phosphate starvation (Lopez-
Fig. 6 Representative root phenotype of Arabidopsis reference and mutants in response to the coumarin supply
Plant Growth Regul
123
Bucio et al. 2002), salt stress (Zolla et al. 2010) and
brassinosteroids treatments (Bao et al. 2004). Among sig-
nals that mediate SIMR, auxin patterning and/or redistri-
bution plays a central role (Potters et al. 2007). The
phosphorus-deficiency SIMR in Arabidopsis is linked to
changes in auxin sensitivity (Lopez-Bucio et al. 2002), the
cadmium SIMR in Arabidopsis and the aluminum SIMR
are correlated with changes in auxin distribution (Kol-
lmeier et al. 2000; Doncheva et al. 2005) and the UV-B
SIMR is associated with altered auxin metabolism (Jansen
et al. 2001). Thus, to examine the auxin role in coumarin
SIMR responses in Arabidopsis roots, IAA with or without
the allelochemical were applied in the growth medium. In
absence of coumarin, up to 10-6 M IAA did not cause
significant effects on both root traits, while the higher IAA
concentrations dramatically decreased primary root length
increasing the number of lateral roots. These effects were
further marked in treatments with a combination of 10-4 M
coumarin and lower IAA concentrations. These results
indicated that at low IAA levels, coumarin and auxin
produced largely additive effects on both inhibition of
primary root elongation and promotion of lateral root for-
mation. However, at highest IAA concentrations, no addi-
tive effects were observed with coumarin. Probably, as
proposed by Bao et al. (2004) in brassinosteroids and auxin
interaction, at low IAA levels, coumarin could increase the
endogenous auxin level, transport or sensitivity promoting
its activity, while at highest, signals activating lateral root
formation may be saturated, masking the additive effects of
Fig. 7 Primary root length (a) and lateral root number (b) of
Arabidopsis mutants exposed to 0 or 10-4 M coumarin for 7 days.
The values are presented as mean ± SE (n = 15). ***, **, * = sig-
nificantly different at p B 0.001, 0.01, 0.05, respectively, according to
Student’s unpaired t test
Fig. 8 Branching density of Arabidopsis mutants exposed to 10-4 M
coumarin for 7 days. The values are presented as mean ± SE
(n = 15). ***, **, * = significantly different at p B 0.001, 0.01,
0.05, respectively, according to Student’s unpaired t test
Fig. 9 Root zone distribution of Arabidopsis mutants exposed to 0
(-) and 10-4 M (?) coumarin for 7 days. The values are presented as
percentage of the distribution along primary root (n = 15)
Plant Growth Regul
123
auxin and coumarin. Since auxin-coumarin interaction may
involve auxin polar transport (PAT), TIBA and NPA
effects on root traits, alone or in combination with cou-
marin, were tested. The results indicated that, in the pre-
sence of both inhibitors, the degree of inhibition of primary
root elongation conferred by coumarin did not vary. Con-
versely, both inhibitors alone not only blocked lateral root
formation in wild type seedlings but also reduced coumarin
promotion of lateral root formation. These results further
confirmed a potential interaction between coumarin and
auxin in the promotion of lateral root formation and pro-
viding additional evidence that SIMR induced by coumarin
may act through PAT modulation to activate lateral root
development. Recently, Chavez-Aviles et al. (2013) dem-
onstrated that SIMR induced by osmotic stress implicated
the modulation of PAT triggering LR development in
Arabidopsis seedlings.
Furthermore, coumarin not only restored the lateral
roots number to control value in presence of auxin trans-
port inhibitors but also the mitotic sites number, suggesting
that it was able to overcome the inhibitory effect of both
auxin transport inhibitors positively acting on lateral root
initiation and emergence of lateral root primordia.
Since local auxin gradient in the roots is the result of the
directional cell–cell transport coordinated by influx
(AUX1/LAX family) and efflux (PIN family) systems
(Mravec et al. 2008), coumarin could interact with the
auxin transport influencing either or both, the PIN or
AUX1 proteins. To better understand this interaction,
Arabidopsis mutants failed in influx or efflux carriers, have
been employed. Except for aux1-22, all Arabidopsis
mutants exhibited a significant inhibition of primary root
elongation in response to coumarin respect to untreated
seedlings. AUX1 is an auxin influx facilitator participating
in both acropetal and basipetal auxin transport at the root
tip (Swarup et al. 2001). The coumarin resistance of the
aux1-22 mutant therefore suggested that auxin transport
within the root tip may have a positive role in mediating
the coumarin effect on primary root inhibition. Moreover,
except to pin1 mutant and Col 0, coumarin did not cause
any increase in lateral roots number, indicating a possible
modulation of the PIN2, PIN3 and AUX1/LAX carriers in
lateral roots development induced by this compound. The
results on root branching density and root zone distribution,
processes largely known to be dependent on auxin flux (De
Smet et al. 2007; Marchant et al. 2002; Peret et al. 2009),
further confirmed that coumarin may modulate auxin dis-
tribution through influx or/and efflux proteins. Finally, root
Fig. 10 Effect of coumarin on the development of root hairs in Arabidopsis reference and mutant (bar = 150 lm)
Fig. 11 Root hair length (a) and Root hair density (b) of Arabidopsis
mutants exposed to 0 or 10-4 M coumarin for 7 days. The values are
presented as mean ± SE (n = 30). ***, **, * = significantly differ-
ent at p B 0.001, 0.01, 0.05, respectively according to Student’s
unpaired t test
Plant Growth Regul
123

hair formation, process strongly influenced by auxin bal-
ance (Evans et al. 1994; Jones et al. 2009), in wild type and
mutants in response to coumarin have been investigated.
Except to aux1-22 mutant, coumarin treatment led to an
increase in root hair elongation. Considering that this
process is a result of increased auxin concentration by
AUX1-dependent transport in epidermal cells (Evans et al.
1994; Sabatini et al. 1999; Jones et al. 2009), coumarin
might be act via the modulation of auxin influx in root once
again. Furthermore, not differences in root hair density
have been observed in aux1-22 and pin3-5 mutant in
response to coumarin.
In conclusion, the results suggested a functional inter-
action between coumarin and auxin polar transport, and
particularly with AUX1 carrier which determined rootward
auxin transport. Further studies will be needed to better
delineate coumarin mechanisms on regulating auxin
abundance, subcellular trafficking and/or transport activity
of auxin carriers. The dissection of such downstream
organizing events will be a future challenge.
Acknowledgments We would like to thank Eva Benkova (VIB
Department of Plant Systems Biology, UGent), Malcolm Bennett and
Ranjan Swarup (Plant Sciences Division, School of Biosciences,
University of Nottingham) for providing Arabidopsis mutants used in
the present work.
References
Abenavoli MR, De Santis C, Sidari M, Sorgona A, Badiani M, Cacco
G (2001) Influence of coumarin on the net nitrate uptake in
durum wheat. New Phytol 150:619–627
Abenavoli MR, Sorgona A, Albano S, Cacco G (2004) Coumarin
differentially affects the morphology of different root types of
maize seedlings. J Chem Ecol 30:1871–1883
Abenavoli MR, Nicolo A, Lupini A, Oliva S, Sorgona A (2008)
Effects of different allelochemicals on root morphology of
Arabidopsis thaliana. Allelopathy J 22:245–252
Avers CJ, Goodwin RH (1956) Studies on root. IV. Effects of
coumarin and scopoletin on the standard root growth pater of
Phleum pretense. Am J Bot 43:612–620
Bao F, Shen J, Brady SR, Muday GK, Asami T, Yang Z (2004)
Brassinosteroids interact with auxin to promote lateral root
development in Arabidopsis. Plant Physiol 134:1624–1631
Benkova E, Michniewicz M, Sauer M, Teichmann T, Seifertova D,
Jurgens G, Friml J (2003) Local, efflux-dependent auxin gradient
as common module for plant organ formation. Cell 115:591–602
Benkova E, Ivanchenko MG, Friml J, Shishkova S, Dubrovsky JG
(2009) A morphogenetic trigger: is there an emerging concept in
plant developmental biology? Trends Plant Sci 14:189–193
Canellas LP, Olivares FL, Okorokova-Facanha AL, Facanha AR
(2002) Humic acids isolated from earthworm compost enhance
root elongation, lateral root emergence, and plasma membrane
H?-ATPase activity in maize roots. Plant Physiol 130:1951–1957
Casimiro I, Beekman T, Graham N, Bhalerao R, Zhang H, Casero P,
Sandberg G, Bennett MJ (2003) Dissecting Arabidopsis lateral
root development. Trends Plant Sci 8:165–171
Chavez-Aviles MN, Andrade-Perez CL, de la Ryes Cruz H (2013)
PP2A mediates lateral root development under NaCl-induced
osmotic stress throughout auxin redistribution in Arabidopsis
thaliana. Plant Soil 368:591–602
De Smet I, Vanneste S, Inze D, Beeckman T (2006) Lateral root
initiation or the birth of a new meristem. Plant Mol Biol
60:871–887
De Smet I, Tetsumura T, De Rybel B, Frei dit Frey N, Laplaze L,
Casimiro I, Swarup R, Naudts M, Vanneste S, Audenaert D, Inze
D, Bennett MJ, Beeckman T (2007) Auxin-dependent regulation
of lateral root positioning in the basal meristem of Arabidopsis.
Development 134:681–690
Doncheva S, Amenos M, Poschenrieder C, Barcelo J (2005) Root cell
patterning: a primary target for aluminium toxicity in maize.
J Exp Bot 56:1213–1220
Dubrovsky JG, Forde BG (2012) Quantitative analysis of lateral root
development: pitfalls and how avoid then. Plant Cell 24:4–14
Dubrovsky JG, Napsucialy-Mendivil S, Duclercq J, Cheng Y,
Shishkova S, Ivanchenko MG, Friml J, Murphy AS, Benkova
E (2011) Auxin minimum defines a development window for
lateral root initiation. New Phytol 191:970–983
Evans ML, Ishikawa H, Estelle MA (1994) Responses of Arabidopsis
roots to auxin studied with high temporal resolution—compar-
ison of wild-type and auxin-response mutants. Planta 194:
215–222
Goodwin RH, Avers CJ (1950) The effect of coumarin derivatives on
growth of Avena root. Am J Bot 37:224–227
Hodge A (2006) Plastic plants and patchy soils. J Exp Bot 57:
401–411
Jansen MAK, Van den Noort RE, Tan A, Prinsen E, Lagrimini M,
Thorneley RNF (2001) Phenol-oxidizing peroxidases contribute
to the protection of plants from ultraviolet radiation stress. Plant
Physiol 126:1012–1023
Jones AR, Kramer EM, Knox K, Swarup R, Bennett MJ, Lazarus CM,
Leyser HM, Grierson CS (2009) Auxin transport through non-
hair cells sustains root-hair development. Nat Cell Biol 11:78–84
Kollmeier M, Felle HH, Horst WJ (2000) Genotypical differences in
aluminium resistance of maize are expressed in the distal part of
the transition zone. Is reduced basipetal auxin flow involved in
inhibition of root elongation by aluminium? Plant Physiol
122:945–956
Kupidlowska E, Kowalec M, Sulkowski G, Zobel AM (1994) The
effect of coumarins on root elongation and ultrastructure of
meristematic cell protoplast. Ann Bot 73:525–530
Laskowski M, Biller S, Stanley K, Kajstura T, Prusty R (2006)
Expression profiling of auxin-treated Arabidopsis roots: toward a
molecular analysis of lateral root emergence. Plant Cell Physiol
47:788–792
Li X, Gruber MY, Hegedus DD, Lydiate DJ, Gao M-J (2011) Effects
of a coumarin derivative, 4-methylumbelliferone, on seed
germination and seedlings establishment in Arabidopsis.
J Chem Ecol 37:880–890
Lopez-Bucio J, Hernandez-Abreu E, Sanchez-Calderon L, Nieto-
Jacobo MF, Simpson J, Herrera-Estrella L (2002) Phosphate
availability alters architecture and causes changes in hormone
sensitivity in the Arabidopsis root system. Plant Physiol
129:244–256
Lopez-Bucio J, Cruz-Ramırez A, Herrera-Estrella L (2003) The role
of nutrient availability in regulating root architecture. Curr Opin
Plant Biol 6:280–287
Lupini A, Sorgona A, Miller AJ, Abenavoli MR (2010) Short-time
effects of coumarin along the maize primary root axis. Plant
Signal Behav 5:1395–1400
Lupini A, Araniti F, Sunseri F, Abenavoli MR (2013) Gravitropic
response induced by coumarin: evidences of ROS distribution
involvement. Plant Signal Behav 8:e23156
Marchant A, Bhalerao R, Casimiro I, Eklof J, Casero PJ, Bennett M,
Sandberg G (2002) AUX1 promotes lateral root formation by
Plant Growth Regul
123

facilitating indole-3-acetic acid distribution between sink and
source tissues in the Arabidopsis seedling. Plant Cell 14:
589–597
Mravec J, Kubes M, Bielach A, Gaykova V, Petrasek J, Skupa P,
Chand S, Benkova E, Zazımalova E, Friml J (2008) Interaction
of PIN and PGP transport mechanisms in auxin distribution-
dependent development. Development 135:3345–3354
Mravec J, Skupa P, Bailly A, Hoyerova K, Krecek P, Bielach A,
Petrasek J, Zhang J, Gaykova V, Stierhof Y-D, Dobrev PI,
Schwarzerova K, Rolcık J, Seifertova D, Luschnig C, Benkova
E, Zazımalova E, Geisler M, Friml J (2009) Subcellular
homeostasis of phytohormone auxin is mediated by the ER-
localized PIN5 transporter. Nature 459:1136–1140
Neumann J (1959) An Auxin-like Action of coumarin. Science
129:1675–1676
Nibau C, Gibbs DJ, Coates JC (2008) Branching out in new
directions: the control of root architecture by lateral root
formation. New Phytol 179:595–614
Parry G, Delbarre A, Marchant A, Swarup R, Napier R, Perrot-
Rechenmann C, Bennett MJ (2001) Novel auxin transport
inhibitors phenocopy the auxin influx carrier mutation aux1.
Plant J 25:399–406
Peret B, De Rybel B, Casimiro I, Benkova I, Swarup R, Laplaze L,
Beeckman T, Bennett MJ (2009) Arabidopsis lateral root
development: an emerging story. Trends Plant Sci 14:399–408
Petrasek J, Friml J (2009) Auxin transport routes in plant develop-
ment. Development 136:2675–2688
Potters G, Pasternak TP, Guisez Y, Palme KJ, Jansen MAK (2007)
Stress-induced morphogenic responses: growing out of trouble?
Trends Plant Sci 12:98–105
Rice EL (1984) Allelopathy. Academic Press, New York
Sabatini S, Beis D, Wolkenfelt H, Murfett J, Guilfoyle T, Malamy J,
Benfey P, Leyser O, Bechtold N, Weisbeek P, Scheres B (1999)
An auxin-dependent distal organizer of pattern and polarity in
the Arabidopsis root. Cell 99:463–472
Schlichting CD (1986) The evolution of phenotypic plasticity in
plants. Annu Rev Ecol Syst 17:667–693
Svensson SB (1971) The Effect of coumarin on root growth and root
histology. Physiol Plant 24:446–470
Svensson SB (1972) The effect of coumarin on growth, production of
dry matter, protein and nucleic acids in roots of maize and wheat
and the interactions of coumarin with metabolic inhibitors.
Physiol Plant 27:13–24
Swarup R, Friml J, Marchant A, Ljung K, Sanberg G, Palme K,
Bennet M (2001) Localization of the auxin permease AUX1
suggests two functionally distinct hormone transport pathways
operate in the Arabidopsis root apex. Genes Dev 15:2648–2653
Whitehead DC, Dibb H, Hartley RD (1982) Phenolic compounds in
soil as influenced by the growth of different plant species. J ApplEcol 19:579–588
Zazımalova E, Murphy AS, Yang H, Hoyerova K, Hosek P (2010)
Auxin transporters: why so many? Cold Spring Harb Perspect
Biol 2:a001552
Zobel AM, Brown SA (1995) Coumarins in the interaction between
the plant and its environment. Allelopathy J 2:9–22
Zolla G, Heimer YM, Barak S (2010) Mild salinity stimulates a
stress-induced morphogenic response in Arabidopsis thaliana
roots. J Exp Bot 61:211–224
Plant Growth Regul
123





























