Embed Size (px)
Citation preview

Food Chemistry 221 (2017) 39–46
Contents lists available at ScienceDirect
Food Chemistry
journal homepage: www.elsevier .com/locate / foodchem
Correlation of the immunostimulatory activities of honeys with theircontents of identified bioactives
http://dx.doi.org/10.1016/j.foodchem.2016.10.0380308-8146/� 2016 Elsevier Ltd. All rights reserved.
⇑ Corresponding author.E-mail addresses: [email protected] (S. Gannabathula), gw.krissansen@
auckland.ac.nz (G.W. Krissansen), [email protected] (L. Bisson-Rowe),[email protected] (M. Skinner), [email protected] (G. Steinhorn),[email protected] (R. Schlothauer).
Swapna Gannabathula a,⇑, Geoffrey W. Krissansen a, Liana Bisson-Rowe a, Margot Skinner b,Gregor Steinhorn c, Ralf Schlothauer c
aDepartment of Molecular Medicine & Pathology, Faculty of Medical and Health Sciences, The University of Auckland, Auckland, New Zealandb Food Science Programme, School of Chemical Sciences, The University of Auckland, Auckland, New ZealandcComvita New Zealand Ltd, Te Puke, New Zealand
a r t i c l e i n f o
Article history:Received 13 May 2016Received in revised form 19 September2016Accepted 10 October 2016Available online 11 October 2016
Keywords:New Zealand honeysNon-New Zealand honeysMonocytesTNF-a
a b s t r a c t
Honeys with unique compositions and properties are worth studying for their health-promoting effects.In order to correlate bioactive content with immunostimulatory activity we compared the abilities of sev-enty eight New Zealand and non-New Zealand honeys to stimulate blood monocytes to release tumournecrosis factor (TNF)-a, and examined the compositions of selected honeys that had widely varyingactivities. All the honeys, except for a Malaysian ‘‘Amber honey” stimulated the release of TNF-a frommonocytes. However, the honeys differed greatly in their immunostimulatory activity, even within thesame honey type. They differed in their contents of immunostimulatory components, including apalbu-mins, arabinogalactan proteins, and apisimin, whose levels did not correlate exactly with immunostim-ulatory activities. We suggest that the immunostimulatory properties of honey may be influenced byother factors, including unidentified immunostimulatory bioactives and immunosuppressive compo-nents; the bioavailability of some bioactives may depend on unidentified factors.
� 2016 Elsevier Ltd. All rights reserved.
1. Introduction
There are several hundred unique types of honey producedworldwide, with each type originating from a different floralsource that is often specific to a particular country (Kaškoniene &Venskutonis, 2010). Honey is classified according to its floralsource. The physical properties of honey vary, depending on watercontent, the floral source, and the proportion of the specific sugars,organic acids, amino acids, enzymes, vitamins, proteins and phyto-chemicals it contains (Ball, 2007; White, 1975). An additional vari-ety of honey, namely honeydew honey, is produced by bees whichharvest honeydew, the sweet secretions of aphids or other plantsap-sucking insects, rather than harvesting nectar (Leroy et al.,2011). The water content of honey may change because of environ-mental factors, such as the weather and humidity, inside the hives(Olaitan, Adeleke, & Iyabo, 2007). Honey composition can changedue to temperature and storage conditions (Stephens et al.,
2015), and the continuous action of the enzymes it contains(White, 1962).
Since antiquity, bee products have been used in traditionalmedicine, with honey being used in the prevention of ulcers, bedsores, and skin infections resulting from burns and wounds(Mandal & Mandal, 2011; Zumla & Lulat, 1989). The use of honeyin wound-healing has been historically anecdotal, but more recentreports claim that honey reduces inflammation and oedema, deb-rides the wound, and promotes granulation, angiogenesis andepithelialization (Molan, 2001a,b). The anti-microbial activities ofhoney have been well studied (Willix, Molan, & Harfoot, 1992),and include both bactericidal and bacteriostatic effects (Cooper,Molan, & Harding, 2002; Wahdan, 1998).
Honey also appears to function as an immunomodulator duringwound-healing. It has been reported to stimulate cells of theimmune system that infiltrate the wound site (Al-Waili, Salom, &Al-Ghamdi, 2011). Unfractionated New Zealand kanuka andmanuka honeys are able to stimulate the release of TNF-a frommonocytes/macrophages (Gannabathula et al., 2012; Raynaudet al., 2013; Tonks, Cooper, Price, Molan, & Jones, 2001; Tonkset al., 2003). We recently reported that arabinogalactan proteins(AGPs) and apisimin, which are commonly present in New Zealandhoneys, are at least partly responsible for stimulating the release of

40 S. Gannabathula et al. / Food Chemistry 221 (2017) 39–46
TNF-a (Gannabathula, Krissansen, Skinner, Steinhorn, &Schlothauer, 2015). The major glycoproteins in honey, namelyapalbumin1 (Apa1; 56 kDa), apalbumin2 (Apa2; 49 kDa), and apal-bumin3 (Apa3; 70 kDa) also stimulate macrophages to releaseTNF-a (Majtan, Kovacova, Bilikova, & Simuth, 2006; Okamotoet al., 2003; Šimúth, Bíliková, Kovácová, Kuzmová, & Schroder,2004).
The present study sought to compare the compositions andimmunostimulatory activities of commercial New Zealand honeysand honeys collected internationally. Through understanding theimmunostimulatory activity and bioactives of different honeyvarieties, it may be possible to identify those that have elevatedactivity that may be suitable for improving human health andwell-being.
2. Materials and methods
2.1. Materials
Commercial samples of New Zealand manuka (Leptospermumscoparium) and clover (Trifolium spp.) honeys were purchased(Countdown, Auckland, NZ). New Zealand and non-New Zealandhoneys (Table 1) were selected and supplied by Comvita New Zeal-and Ltd, Te Puke. Artificial honey was prepared as described(Gannabathula et al., 2015). Lipopolysaccharide (LPS) isolated froma culture of Escherichia coli 0127:B8 was purchased from SigmaAldrich, New Zealand. The Yariv reagent was obtained fromCallaghan Innovation, New Zealand. Mouse anti-human TNF-aantibodies were purchased from BD Biosciences, Auckland, and apolyclonal antiserum raised in mice against synthetic apisimin(synthesized by Peptide 2.0 Inc., Chantilly, VA, USA) was custom-produced by Promab Biotechnologies, Inc., CA. Fresh human bloodmonocytes were obtained from the buffy coats of venous bloodfractionated by centrifugation on Ficoll gradients. Blood wasobtained from healthy adult subjects by informed consent accord-ing to approval 2010/462 from the University of Auckland HumanParticipants Ethics Committee.
2.2. Rocket gel electrophoresis assay
AGPs in honey were quantified by rocket gel electrophoresis,using Gum arabic as a standard, according to a previouslydescribed method (Gannabathula et al., 2015; Komalavilas, Zhu,& Nothnagel, 1991; Wisniewska & Majewska-Sawka, 2007). Theagarose (1%) contained 15 lM Yariv reagent. Electrophoresis wascarried out in a Tris-glycine buffer (25 mM Tris, 200 mM glycine,pH 8.4) for 4–8 h at room temperature (RT) until the rockets werewell developed. The concentrations of AGPs in the samples wereestimated in relation to the area of the precipitin peak formed withknown amounts of Gum arabic.
2.3. Polyacrylamide gel electrophoresis
Honeys were analyzed on a 12% polyacrylamide Tris-glycinesodium dodecyl sulfate (SDS)-gel, and on a 15% polyacrylamideTris-tricine SDS-gel, and the resolved proteins were visualized bystaining with Coomassie blue G-250.
2.4. Western blot analysis
Proteins in honey samples, separated by SDS polyacrylamide gelelectrophoresis (SDS-PAGE) on Tris-tricine gels, were electroblot-ted onto a nitrocellulose membrane. The membranes were blockedwith TBS-Tween buffer containing 5% non-fat milk for 1 h at RT,and incubated overnight with a polyclonal mouse anti-apisimin
antibody diluted 1:5000 in TBS-Tween buffer containing 5% non-fat milk. They were washed and incubated for 2 h in the same buf-fer containing horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG. Immunoreactivity was developed with ECL substratesolution, and the result recorded using a Fujifilm LAS-3000scanner.
2.5. TNF-a ELISA
TNF-a concentrations were measured by ELISA as describedpreviously (Farr, Hurst, & Skinner, 2008; Gannabathula et al.,2012). Briefly, a mouse anti-human TNF-a mAb, used as thecapture antibody, was coated onto ELISA plates at 2 lg/ml, and100 ll of culture supernatant were dispensed into the wells. Afterwashing, a biotinylated mouse anti-human TNF-a antibody at2 lg/ml was added to detect the captured TNF-a. Immunoreactiv-ity was detected with streptavidin-HRP, and developed withtetramethylbenzidine (TMB) Substrate Reagent (BD Biosciences).LPS (100 ng/ml) was included as a standard, against which theresults obtained with 1% solutions of the New Zealand and non-New Zealand honey samples were compared.
2.6. Statistical analysis
Graphs were plotted using Graph pad (Prism). The data wereanalyzed using Statistix 8.1 and significant difference was indi-cated by a p value less than 0.05 (p < 0.05).
3. Results and discussion
3.1. Immunostimulatory activities of eight commercial New Zealandhoneys
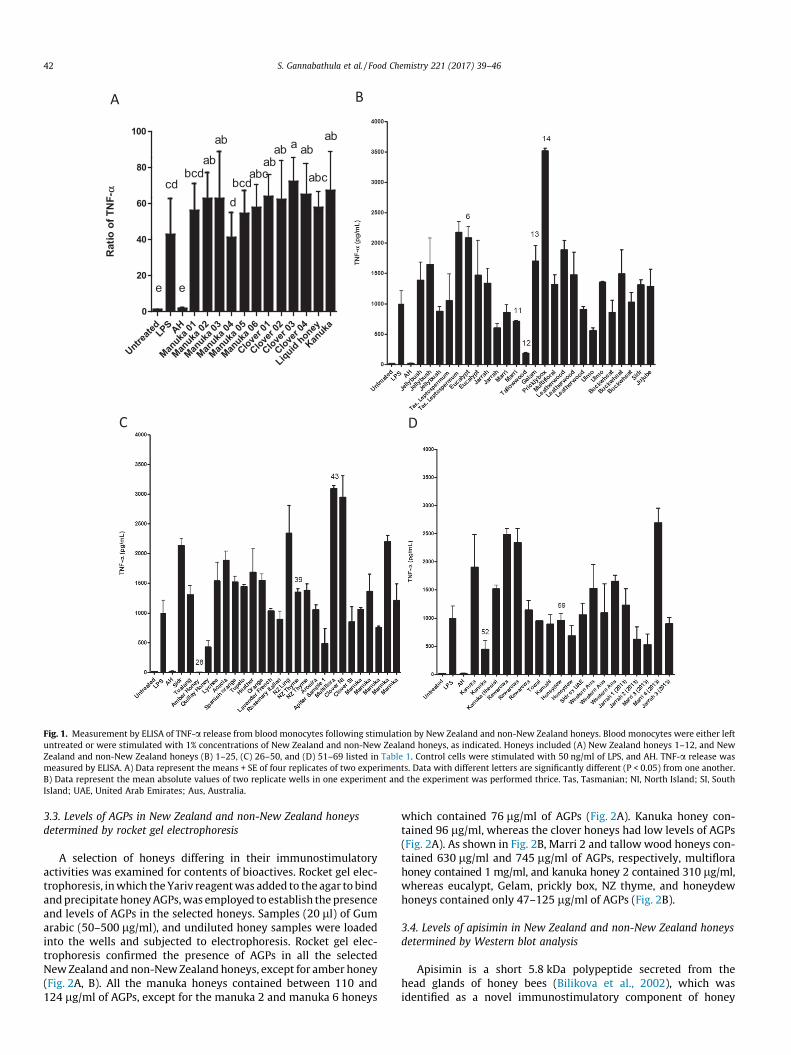
As shown in Fig. 1A, all commercial New Zealand honeysinduced TNF-a release from blood monocytes. Whilst the honeyshad roughly similar immunostimulatory activities, clover 3 honeyhad the strongest activity, followed by kanuka honey, cloverhoneys 1 and 4, and manuka honeys 2 and 3.
3.2. Immunostimulatory activities of forty-eight non-New Zealandhoneys and twenty-one New Zealand honeys
Forty-eight global honeys and twenty-one New Zealand honeyswere screened for immunostimulatory activity, and those of inter-est were selected for further analysis. As shown in Fig. 1B–D, allnon-New Zealand and New Zealand honeys, except for Amberhoney, induced TNF-a release from blood monocytes. The differenthoneys differed greatly in their immunostimulatory activities.Tallowwood honey had the lowest immunostimulatory activityafter amber honey, followed by Quillay and Apiter honeys(Fig. 1B, C). Honeys that displayed strong immunostimulatoryactivity included prickly box, multiflora, and clover NI, which eachinduced the release of P3000 pg/ml of TNF-a (Fig. 1B, C). Otherhoneys with moderate activity that induced the release of 2000–3000 pg/ml of TNF-a included Tasmanian Leptospermum, euca-lypt, Sidr, NZ ling, one of the NZ manuka honeys, two of the threerewarewa honeys, and Marri 4 honey (one of four Marri honeys).
The results suggested a difference in the activities of multiplesamples of a particular honey type. Thus, the activities of 3jellybush honeys differed almost 2-fold, 2 jarrah honeys differedby 2-fold, 4 Marri honeys differed by 4-fold, 2 Ulmo honeys by2-fold, New Zealand kanuka honeys by 3-fold, and New Zealandmanuka honeys by 2-fold. Differences in activity may be due tothe purity of the floral source, aging, or adulteration of the honeys,which would be expected to alter the levels or activities of honey

Table 1New Zealand and non-New Zealand honey samples analyzed in this study.
Serial No. Honey type Origin Plant species
1 Jelly bush 1 Australia Leptospermum polgalifolium2 Jelly bush 2 Australia Leptospermum polgalifolium3 Jelly bush 3 Australia Leptospermum polgalifolium4 Tasmania Leptospermum Tasmania Australia Leptospermum scoparium var.exinium5 Tasmania Leptospermum Tasmania Australia Leptospermum scoparium var.exinium6 Eucalypt 1 Australia Eucalyptus globulus7 Eucalypt 2 Australia Eucalyptus macrorhyncha8 Jarrah 1 Western Australia Eucalyptus marginata9 Jarrah 2 Western Australia Eucalyptus marginata10 Marri 1 Western Australia Corymbia calophylla11 Marri 2 Western Australia Corymbia calophylla12 Tallow wood Tasmania Australia Eucalyptus microcorys13 Gelam Australia Melaleuca cajaputi14 Prickly box Tasmania Australia Bursaria spinosa15 Multifloral Australia Unspecified16 Leatherwood 1 Tasmania Australia Eucryphia lucida17 Leatherwood 2 Tasmania Australia Eucryphia lucida18 Leatherwood 3 Tasmania Australia Eucryphia lucida19 Ulmo 1 Chile Eucryphia cordifolia20 Ulmo 2 Chile Eucryphia cordifolia21 Buckwheat Canada Fagopyrum esculentum22 Buckwheat Probably Japan or China Fagopyrum esculentum23 Buckwheat China via Parodi Fagopyrum esculentum24 Sidr 1 Saudi Arabia Ziziphus spina-christi25 Jujube China Ziziphus zizyphus26 Sidr 2 Yemen Ziziphus spina-christi27 Tualang Malaysia Koompassia excelsa28 Amber honey Malaysia Possibly Tualang honey29 Quillay honey Chile Quillaja saponaria30 Lychee Indonesia Litchi chinensis31 Acacia France Acacia sp.32 Spanish orange Italy Citrus sinensis33 Tupelo USA Nyssa sp.34 Heather Scotland Erica sp.35 Orange USA Citrus sinensis36 Lavender French France Lavendula sp.37 Rosemart Italian Italy Rosemarinus sp.38 NZ Ling New Zealand Erica sp.39 NZ Thyme New Zealand Thymus vulgaris40 NZ Thyme New Zealand Thymus vulgaris41 Aroeira Brazil Schinus terebinthifolius42 Apiter sample 1 Uruguay Unspecified43 Multifloral Samoa Unspecified44 Clover NI New Zealand Trifolium spp.45 Clover SI New Zealand Trifolium spp.46 Manuka 1 New Zealand L. Scoparium47 Manuka 2 New Zealand L. Scoparium48 Manuka 3 New Zealand L. Scoparium49 Manuka 4 New Zealand L. Scoparium50 Manuka 5 New Zealand L. Scoparium51 Kanuka 1 New Zealand Kunzea ericoides52 Kanuka 2 New Zealand Kunzea ericoides53 Kanuka 3 New Zealand Kunzea ericoides54 Rewarewa 1 New Zealand Knightia excelsa55 Rewarewa 2 New Zealand Knightia excelsa56 Rewarewa 3 New Zealand Knightia excelsa57 Towai New Zealand Weinmannia silvicola58 Kamahi New Zealand Weinmannia racemosa59 Honeydew New Zealand Honeydew60 Honeydew New Zealand Honeydew61 Sidr ex UAE Ex Paengaroa, New Zealand Ziziphus spina-christi62 Western Australia Leptospermum 1 Western Australia Leptospermum subtenue63 Western Australia Leptospermum 2 Western Australia Leptospermum subtenue64 Western Australia Leptospermum 3 Western Australia Leptospermum subtenue65 Jarrah 3 Western Australia Eucalyptus marginata66 Jarrah 4 Western Australia Eucalyptus marginata67 Marri 3 Western Australia Corymbia calophylla68 Marri 4 Western Australia Corymbia calophylla69 Jarrah 5 Western Australia Eucalyptus marginata
S. Gannabathula et al. / Food Chemistry 221 (2017) 39–46 41
bioactives. Whilst beekeepers attempt to maintain monoflorality,honeys can nonetheless become blended, which would potentiallylead to an alteration in activity.
The levels of AGPs, apisimin, and apalbumins in some of theabove honeys were determined to provide an explanation for thedifferences in the immunostimulatory activities of the honeys.
A B
Untreate
dLPS AH
Man
uka 01
Manuka
02
Man
uka 03
Man
uka 04
Man
uka 05
Man
uka 06
Clover 0
1
Clover 0
2
Clover 0
3
Clover 0
4
Liquid honey
Kanuka
0
20
40
60
80
100
e
bcd
ab
e
ab aab
bcd abc
b
ab abab
abccdd
Rat
io o
f TN
F-α
C D
Fig. 1. Measurement by ELISA of TNF-a release from blood monocytes following stimulation by New Zealand and non-New Zealand honeys. Blood monocytes were either leftuntreated or were stimulated with 1% concentrations of New Zealand and non-New Zealand honeys, as indicated. Honeys included (A) New Zealand honeys 1–12, and NewZealand and non-New Zealand honeys (B) 1–25, (C) 26–50, and (D) 51–69 listed in Table 1. Control cells were stimulated with 50 ng/ml of LPS, and AH. TNF-a release wasmeasured by ELISA. A) Data represent the means + SE of four replicates of two experiments. Data with different letters are significantly different (P < 0.05) from one another.B) Data represent the mean absolute values of two replicate wells in one experiment and the experiment was performed thrice. Tas, Tasmanian; NI, North Island; SI, SouthIsland; UAE, United Arab Emirates; Aus, Australia.
42 S. Gannabathula et al. / Food Chemistry 221 (2017) 39–46
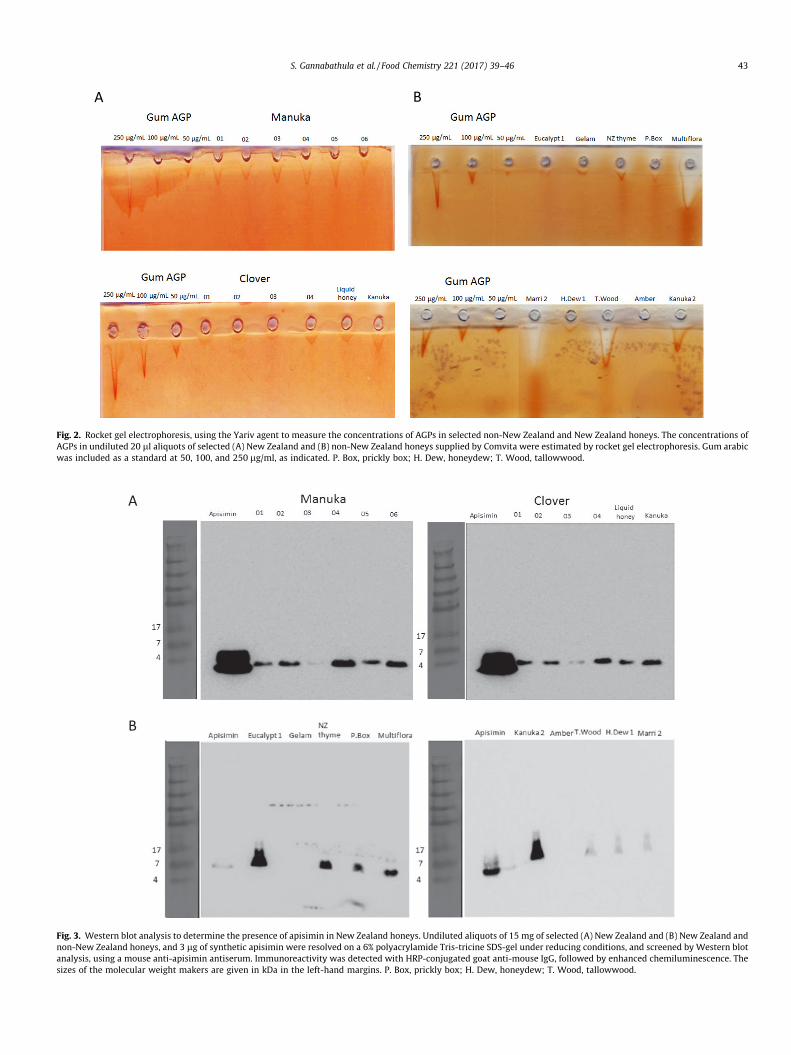
3.3. Levels of AGPs in New Zealand and non-New Zealand honeysdetermined by rocket gel electrophoresis
A selection of honeys differing in their immunostimulatoryactivities was examined for contents of bioactives. Rocket gel elec-trophoresis, inwhich theYariv reagentwas added to the agar to bindandprecipitate honeyAGPs,was employed to establish the presenceand levels of AGPs in the selected honeys. Samples (20 ll) of Gumarabic (50–500 lg/ml), and undiluted honey samples were loadedinto the wells and subjected to electrophoresis. Rocket gel elec-trophoresis confirmed the presence of AGPs in all the selectedNewZealand and non-NewZealand honeys, except for amber honey(Fig. 2A, B). All the manuka honeys contained between 110 and124 lg/ml of AGPs, except for the manuka 2 and manuka 6 honeys
which contained 76 lg/ml of AGPs (Fig. 2A). Kanuka honey con-tained 96 lg/ml, whereas the clover honeys had low levels of AGPs(Fig. 2A). As shown in Fig. 2B, Marri 2 and tallow wood honeys con-tained 630 lg/ml and 745 lg/ml of AGPs, respectively, multiflorahoney contained 1 mg/ml, and kanuka honey 2 contained 310 lg/ml,whereas eucalypt, Gelam, prickly box, NZ thyme, and honeydewhoneys contained only 47–125 lg/ml of AGPs (Fig. 2B).
3.4. Levels of apisimin in New Zealand and non-New Zealand honeysdetermined by Western blot analysis
Apisimin is a short 5.8 kDa polypeptide secreted from thehead glands of honey bees (Bilikova et al., 2002), which wasidentified as a novel immunostimulatory component of honey
A B
Fig. 2. Rocket gel electrophoresis, using the Yariv agent to measure the concentrations of AGPs in selected non-New Zealand and New Zealand honeys. The concentrations ofAGPs in undiluted 20 ll aliquots of selected (A) New Zealand and (B) non-New Zealand honeys supplied by Comvita were estimated by rocket gel electrophoresis. Gum arabicwas included as a standard at 50, 100, and 250 lg/ml, as indicated. P. Box, prickly box; H. Dew, honeydew; T. Wood, tallowwood.
A
B
Fig. 3. Western blot analysis to determine the presence of apisimin in New Zealand honeys. Undiluted aliquots of 15 mg of selected (A) New Zealand and (B) New Zealand andnon-New Zealand honeys, and 3 lg of synthetic apisimin were resolved on a 6% polyacrylamide Tris-tricine SDS-gel under reducing conditions, and screened by Western blotanalysis, using a mouse anti-apisimin antiserum. Immunoreactivity was detected with HRP-conjugated goat anti-mouse IgG, followed by enhanced chemiluminescence. Thesizes of the molecular weight makers are given in kDa in the left-hand margins. P. Box, prickly box; H. Dew, honeydew; T. Wood, tallowwood.
S. Gannabathula et al. / Food Chemistry 221 (2017) 39–46 43
A
B
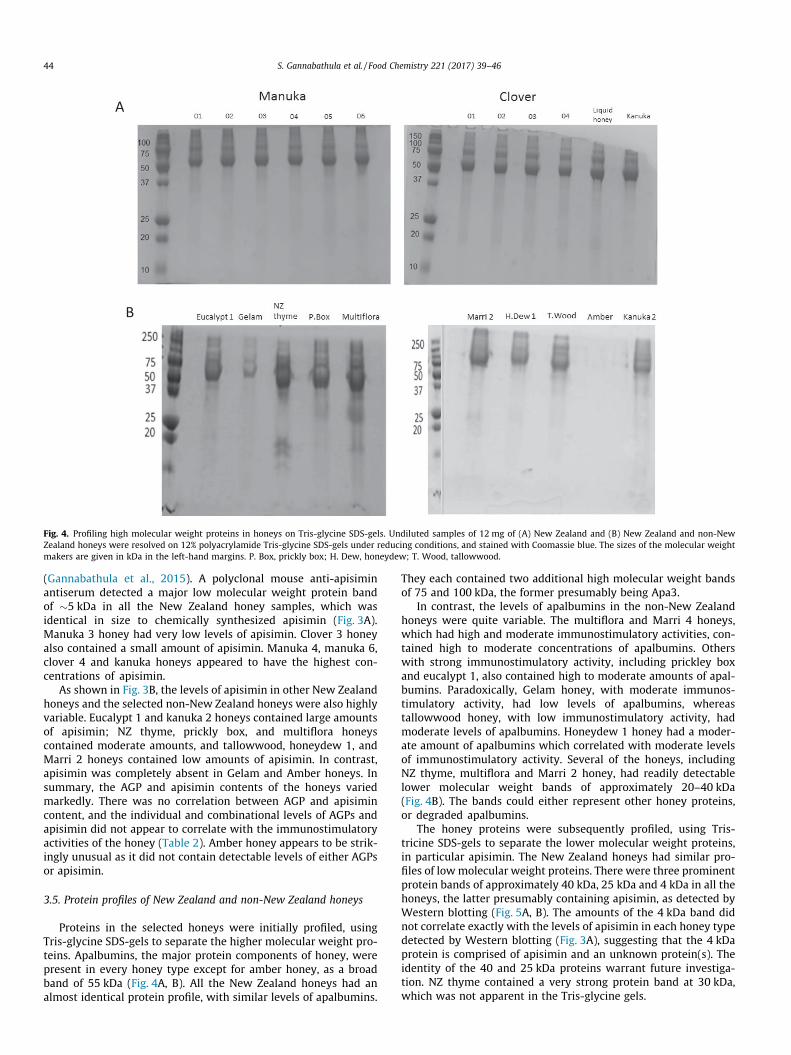
Fig. 4. Profiling high molecular weight proteins in honeys on Tris-glycine SDS-gels. Undiluted samples of 12 mg of (A) New Zealand and (B) New Zealand and non-NewZealand honeys were resolved on 12% polyacrylamide Tris-glycine SDS-gels under reducing conditions, and stained with Coomassie blue. The sizes of the molecular weightmakers are given in kDa in the left-hand margins. P. Box, prickly box; H. Dew, honeydew; T. Wood, tallowwood.
44 S. Gannabathula et al. / Food Chemistry 221 (2017) 39–46
(Gannabathula et al., 2015). A polyclonal mouse anti-apisiminantiserum detected a major low molecular weight protein bandof �5 kDa in all the New Zealand honey samples, which wasidentical in size to chemically synthesized apisimin (Fig. 3A).Manuka 3 honey had very low levels of apisimin. Clover 3 honeyalso contained a small amount of apisimin. Manuka 4, manuka 6,clover 4 and kanuka honeys appeared to have the highest con-centrations of apisimin.
As shown in Fig. 3B, the levels of apisimin in other New Zealandhoneys and the selected non-New Zealand honeys were also highlyvariable. Eucalypt 1 and kanuka 2 honeys contained large amountsof apisimin; NZ thyme, prickly box, and multiflora honeyscontained moderate amounts, and tallowwood, honeydew 1, andMarri 2 honeys contained low amounts of apisimin. In contrast,apisimin was completely absent in Gelam and Amber honeys. Insummary, the AGP and apisimin contents of the honeys variedmarkedly. There was no correlation between AGP and apisimincontent, and the individual and combinational levels of AGPs andapisimin did not appear to correlate with the immunostimulatoryactivities of the honey (Table 2). Amber honey appears to be strik-ingly unusual as it did not contain detectable levels of either AGPsor apisimin.
3.5. Protein profiles of New Zealand and non-New Zealand honeys
Proteins in the selected honeys were initially profiled, usingTris-glycine SDS-gels to separate the higher molecular weight pro-teins. Apalbumins, the major protein components of honey, werepresent in every honey type except for amber honey, as a broadband of 55 kDa (Fig. 4A, B). All the New Zealand honeys had analmost identical protein profile, with similar levels of apalbumins.
They each contained two additional high molecular weight bandsof 75 and 100 kDa, the former presumably being Apa3.
In contrast, the levels of apalbumins in the non-New Zealandhoneys were quite variable. The multiflora and Marri 4 honeys,which had high and moderate immunostimulatory activities, con-tained high to moderate concentrations of apalbumins. Otherswith strong immunostimulatory activity, including prickley boxand eucalypt 1, also contained high to moderate amounts of apal-bumins. Paradoxically, Gelam honey, with moderate immunos-timulatory activity, had low levels of apalbumins, whereastallowwood honey, with low immunostimulatory activity, hadmoderate levels of apalbumins. Honeydew 1 honey had a moder-ate amount of apalbumins which correlated with moderate levelsof immunostimulatory activity. Several of the honeys, includingNZ thyme, multiflora and Marri 2 honey, had readily detectablelower molecular weight bands of approximately 20–40 kDa(Fig. 4B). The bands could either represent other honey proteins,or degraded apalbumins.
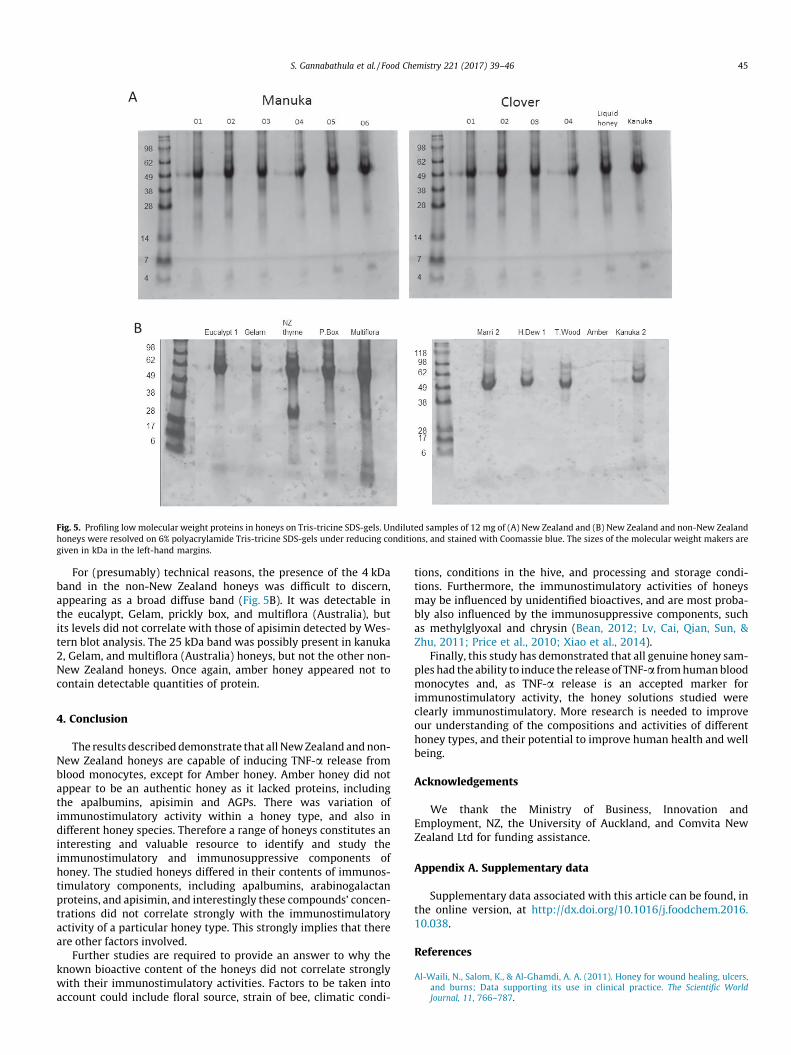
The honey proteins were subsequently profiled, using Tris-tricine SDS-gels to separate the lower molecular weight proteins,in particular apisimin. The New Zealand honeys had similar pro-files of lowmolecular weight proteins. There were three prominentprotein bands of approximately 40 kDa, 25 kDa and 4 kDa in all thehoneys, the latter presumably containing apisimin, as detected byWestern blotting (Fig. 5A, B). The amounts of the 4 kDa band didnot correlate exactly with the levels of apisimin in each honey typedetected by Western blotting (Fig. 3A), suggesting that the 4 kDaprotein is comprised of apisimin and an unknown protein(s). Theidentity of the 40 and 25 kDa proteins warrant future investiga-tion. NZ thyme contained a very strong protein band at 30 kDa,which was not apparent in the Tris-glycine gels.
A
B
Fig. 5. Profiling lowmolecular weight proteins in honeys on Tris-tricine SDS-gels. Undiluted samples of 12 mg of (A) New Zealand and (B) New Zealand and non-New Zealandhoneys were resolved on 6% polyacrylamide Tris-tricine SDS-gels under reducing conditions, and stained with Coomassie blue. The sizes of the molecular weight makers aregiven in kDa in the left-hand margins.
S. Gannabathula et al. / Food Chemistry 221 (2017) 39–46 45
For (presumably) technical reasons, the presence of the 4 kDaband in the non-New Zealand honeys was difficult to discern,appearing as a broad diffuse band (Fig. 5B). It was detectable inthe eucalypt, Gelam, prickly box, and multiflora (Australia), butits levels did not correlate with those of apisimin detected by Wes-tern blot analysis. The 25 kDa band was possibly present in kanuka2, Gelam, and multiflora (Australia) honeys, but not the other non-New Zealand honeys. Once again, amber honey appeared not tocontain detectable quantities of protein.
4. Conclusion
The results describeddemonstrate that all NewZealand andnon-New Zealand honeys are capable of inducing TNF-a release fromblood monocytes, except for Amber honey. Amber honey did notappear to be an authentic honey as it lacked proteins, includingthe apalbumins, apisimin and AGPs. There was variation ofimmunostimulatory activity within a honey type, and also indifferent honey species. Therefore a range of honeys constitutes aninteresting and valuable resource to identify and study theimmunostimulatory and immunosuppressive components ofhoney. The studied honeys differed in their contents of immunos-timulatory components, including apalbumins, arabinogalactanproteins, and apisimin, and interestingly these compounds’ concen-trations did not correlate strongly with the immunostimulatoryactivity of a particular honey type. This strongly implies that thereare other factors involved.
Further studies are required to provide an answer to why theknown bioactive content of the honeys did not correlate stronglywith their immunostimulatory activities. Factors to be taken intoaccount could include floral source, strain of bee, climatic condi-
tions, conditions in the hive, and processing and storage condi-tions. Furthermore, the immunostimulatory activities of honeysmay be influenced by unidentified bioactives, and are most proba-bly also influenced by the immunosuppressive components, suchas methylglyoxal and chrysin (Bean, 2012; Lv, Cai, Qian, Sun, &Zhu, 2011; Price et al., 2010; Xiao et al., 2014).
Finally, this study has demonstrated that all genuine honey sam-ples had the ability to induce the release of TNF-a fromhumanbloodmonocytes and, as TNF-a release is an accepted marker forimmunostimulatory activity, the honey solutions studied wereclearly immunostimulatory. More research is needed to improveour understanding of the compositions and activities of differenthoney types, and their potential to improve human health and wellbeing.
Acknowledgements
We thank the Ministry of Business, Innovation andEmployment, NZ, the University of Auckland, and Comvita NewZealand Ltd for funding assistance.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.foodchem.2016.10.038.
References
Al-Waili, N., Salom, K., & Al-Ghamdi, A. A. (2011). Honey for wound healing, ulcers,and burns; Data supporting its use in clinical practice. The Scientific WorldJournal, 11, 766–787.

46 S. Gannabathula et al. / Food Chemistry 221 (2017) 39–46
Ball, D. W. (2007). The chemical composition of honey. Journal of ChemicalEducation, 84, 1643–1646.
Bean, A. (2012). Investigating the anti-inflammatory activity of honey. University ofWaikato.
Bilikova, K., Hanes, J., Nordhoff, E., Saenger, W., Klaudiny, J., & Simuth, J. (2002).Apisimin, a new serine-valine-rich peptide from honeybee (Apis Mellifera L.)Royal Jelly: Purification and molecular characterization. Federation of EuropeanBiochemical Societies, 528, 125–129.
Cooper, R. A., Molan, P. C., & Harding, K. G. (2002). The sensitivity to honey of Gram-positive cocci of clinical significance isolated from wounds. Journal of AppliedMicrobiology, 93, 857–863.
Farr, J., Hurst, S., & Skinner, M. (2008). Anti-inflammatory effects of kiwifruit. AsiaPacific Journal of Clinical Nutrition, 32, 20–25.
Gannabathula, S., Krissansen, G. W., Skinner, M., Steinhorn, G., & Schlothauer, R.(2015). Honeybee apisimin and plant arabinogalactans in honey costimulatemonocytes. Food Chemistry, 168, 34–40.
Gannabathula, S., Skinner, M. A., Rosendale, D., Greenwood, J. M., Mutukumira, A. N.,Steinhorn, G., & Schlothauer, R. C. (2012). Arabinogalactan proteins contributeto the immunostimulatory properties of New Zealand honeys.Immunopharmacology and Immunotoxicology, 34, 598–607.
Kaškoniene, V., & Venskutonis, P. R. (2010). Floral markers in honey of variousbotanical and geographic origins: A Review. Comprehensive Reviews in FoodScience and Food Safety, 9, 620–634.
Komalavilas, P., Zhu, J.-K., & Nothnagel, E. A. (1991). Arabinogalactan-proteins fromthe suspension culture medium and plasma membrane of rose cells. Journal ofBiological Chemistry, 266, 15956–15965.
Leroy, P. D., Wathelet, B., Sabri, A., Francis, F., Verheggen, F. J., Capella, Q., Thonart, P.,et al. (2011). Aphid-host plant interactions: Does aphid honeydew exactlyreflect the host plant amino acid composition? Arthropod-Plant Interactions, 5,193–199.
Lv, P. C., Cai, T. T., Qian, Y., Sun, J., & Zhu, H. L. (2011). Synthesis, biological evaluationof chrysin derivatives as potential immunosuppressive agents. European Journalof Medicinal Chemistry, 46, 393–398.
Majtan, J., Kovacova, E., Bilikova, K., & Simuth, J. (2006). The immunostimulatoryeffect of the recombinant apalbumin 1-major honeybee Royal Jelly protein-onTNF-a release. International Immunopharmacology, 6, 269–278.
Mandal, M. D., & Mandal, S. (2011). Honey: Its medicinal property and antibacterialactivity. Asian Pacific Journal of Tropical Biomedicine, 1, 154–160.
Molan, P. C. (2001). Potential of honey in the treatment of wounds and burns.American Journal of Clinical Dermatology, 2, 13–19.
Molan, P. C. (2001). Why honey is effective as a medicine. 2. The scientificexplanation of its effects. Bee World, 82, 22–40.
Okamoto, I., Taniguchi, Y., Kunikata, T., Kohno, K., Iwaki, K., Ikeda, M., & Kurimoto,M. (2003). Major Royal Jelly protein 3 modulates immune responses in vitro andin vivo. Life Sciences, 73, 2029–2045.
Olaitan, P. B., Adeleke, O. E., & Iyabo, O. (2007). Honey: A reservoir formicroorganisms and an inhibitory agent for microbes. African Health Sciences,7, 159–165.
Price, C. L., Hassi, H. O., English, N. R., Blakemore, A. I., Stagg, A. J., & Knight, S. C.(2010). Methylglyoxal modulates immune responses: Relevance to diabetes.Journal of Cellular and Molecular Medicine, 14, 1806–1815.
Raynaud, A., Ghezali, L., Gloaguen, V., Liagre, B., Quero, F., & Petit, J.-M. (2013).Honey-induced macrophage stimulation: Ap-1 and Nf-Kb activation andcytokine production are unrelated to LPS content of honey. InternationalImmunopharmacology, 17, 874–879.
Šimúth, J., Bíliková, K., Kovácová, E., Kuzmová, Z., & Schroder, W. (2004).Immunochemical approach to detection of adulteration in honey:Physiologically active royal jelly protein stimulating TNF-a release is aregular component of honey. Journal of Agricultural and Food Chemistry, 52,2154–2158.
Stephens, J.M., Greenwood, D.R., Feamley, L., Bong, J., Schlothauer, R. C., & Loomes, K.M. (2015). Honey production and compositional parameters. Processing andImpact on Active Components in Food, 675–680, ISBN: 978-0-12-404699-3,Chapter 81.
Tonks, A., Cooper, R. A., Price, A. J., Molan, P. C., & Jones, K. P. (2001). Stimulation ofTNF-a release in monocytes by honey. Cytokine, 14, 240–242.
Tonks, A. J., Cooper, R. A., Jones, K. P., Blair, S., Parton, J., & Tonks, A. (2003). Honeystimulates inflammatory cytokine production from monocytes. Cytokine, 21,242–247.
Wahdan, H. A. L. (1998). Causes of the antimicrobial activity of honey. Infection, 26,26–31.
White, J. (1975). Physical characteristics of honey. Honey: A comprehensive survey,207–239.
White, J. W. (1962). Composition of American Honeys. US Dept. of Agriculture, 1–24.Willix, D. J., Molan, P. C., & Harfoot, C. G. (1992). A comparison of the sensitivity of
wound-infecting species of bacteria to the antibacterial activity of manukahoney and other honey. Journal of Applied Bacteriology, 73, 388–394.
Wisniewska, E., & Majewska-Sawka, A. (2007). Arabinogalactan-proteins stimulatethe organogenesis of guard cell protoplasts-derived callus in sugar beet. PlantCell Reports, 26, 1457–1467.
Xiao, J., Zhai, H., Yao, Y., Wang, C., Jiang, W., Zhang, C., Simard, A. R., et al. (2014).Chrysin attenuates experimental autoimmune neuritis by suppressingimmuno-inflammatory responses. Neuroscience, 262, 156–164.
Zumla, A., & Lulat, A. (1989). Honey–a remedy rediscovered. Journal of the RoyalSociety of Medicine, 82, 384–385.





























