Embed Size (px)
Citation preview

Meningococcal peptidoglycan and penicillin G susceptibility
1
Correlation between alterations of the penicillin-binding protein 2 and modifications of the
peptidoglycan structure in Neisseria meningitidis with reduced susceptibility to penicillin G
Aude Antignac1, Ivo G. Boneca2, Jean-Claude Rousselle3, Abdelkader Namane3, Jean-
Philippe Carlier4, Julio A. Vázquez5, Andrew Fox6, Jean-Michel Alonso1, and Muhamed-
Kheir Taha1*.
1Unité des Neisseria and Centre National de Référence des Méningocoques, 2Unité de
Pathogénie Bactérienne des Muqueuses, 3Plate-forme de Protéomique, 4Unité des Bactéries
Anaérobies et des Toxines, Institut Pasteur, 28 rue du Dr Roux, 75724 Paris Cedex 15,
France.
5Spanish Reference Laboratory for Meningococci, National Center for Microbiology, Instituto
de Salud Carlos III, Majadahonda, Madrid, 28220, Spain.
6Meningococcal Reference Unit, Manchester Public Health Laboratory, Withington Hospital,
Manchester, M20 2LR, England.
Running title: Meningococcal peptidoglycan and penicillin G susceptibility
* corresponding author
Tel: +33-1-45688438
Fax: +33-1-45688338
Email: [email protected]
Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on June 10, 2003 as Manuscript M304607200 by guest on A
pril 13, 2020http://w
ww
.jbc.org/D
ownloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
2
SUMMARY
Reduced susceptibility to penicillin G in Neisseria meningitidis is directly correlated with
alterations in the penA gene, which encodes the penicillin-binding protein (PBP) 2. Using
purified PBP2s from different backgrounds, we confirmed that the reduced susceptibility to
penicillin G is associated with a decreased affinity of altered PBP2s for penicillin G. Infrared
spectroscopy analysis using isogenic penicillin susceptible strains and strains with reduced
susceptibility to penicillin G suggested that the meningococcal cell wall is also modified in a
penA-dependent manner. Moreover, reverse-phase high pressure liquid chromatography and
mass spectrometry analysis of these meningococcal strains confirmed the modifications of
peptidoglycan components and showed an increase in the peaks corresponding to
pentapeptide-containing muropeptides. These results suggest that the D,D-transpeptidase
and/or D,D-carboxypeptidase activities of PBP2 are modified by the changes in penA gene.
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
3
INTRODUCTION
Neisseria meningitidis is an exclusive human bacterium that usually infects the
nasopharynx. It occasionally provokes invasive infections such as septicemia, meningitis and
arthritis. Meningococcal infections are still a major public health concern due to periodic
epidemics of meningitis in Africa and local outbreaks in developed countries. This concern is
heightened by the increasing prevalence of meningococcal strains with reduced susceptibility
to penicillin G (PenI), one of the major antibiotics used for the treatment of meningococcal
infections.
Penicillin G binds to penicillin-binding proteins (PBPs). SDS-PAGE analysis showed
that N. meningitidis contains three PBPs (1,2): PBP1 encoded by ponA, PBP2 encoded by
penA, and PBP3 encoded by pbp3. Two other putative PBPs have also been identified from
the complete genomic sequences of a serogroup B strain (3) and a serogroup A strain (4) of N.
meningitidis. Meningococcal strains that are highly resistant to penicillin G (MIC > 1 µg/ml)
due to the production of a plasmid-encoded β–lactamase, have been detected in some
countries (5-8). However, meningococcal strains with reduced susceptibility to penicillin G
(PenI) (MICs between 0.125 and 1 µg/ml), have been reported worldwide (9) and this
reduction in susceptibility is at least partly due to the decreased affinity of PBP2 encoded by
an altered penA gene (10,11). N. gonorrhoeae also shows increased resistance to penicillin G
due to the decreased affinity of both PBP1 and PBP2 for penicillin G, combined with
alterations of the gonococcal porin and the overexpression of the MtrCDE efflux pump (12).
N. meningitidis is naturally competent for transformation, which enables the horizontal
transfer of DNA between strains, resulting in mosaic structures at genetic loci and a highly
diverse meningococcal population (13,14). The penA gene is altered as a result of DNA
transfer between meningococcal strains or from commensal Neisseria species (15-18). We
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
4
previously developed a rapid approach to define meningococcal penA alleles based on
restriction fragment length polymorphism (RFLP). Penicillin susceptible (PenS) strains harbor
highly related penA alleles, regardless of their genotype. Conversely, PenI strains harbor
different penA alleles and the PenI phenotype is directly related to alterations in the penA gene
in N. meningitidis (19). The penA gene encodes PBP2, a 60 kDa protein, which is
homologous to high-molecular weight class B PBPs. By analogy with PBP3 from E. coli,
PBP2 probably catalyzes a transpeptidation reaction that is necessary for the cross-linking of
peptidoglycan in the meningococcal cell wall. Alterations of penA gene mostly affect the 3’
transpeptidase-encoding region. The active serine residue SXXK, the SXN and the KTG
motifs, which are all located in this region, are usually conserved, but a high degree of
polymorphism is observed in the surrounding sequences (19).
As penicillin binds covalently to PBP2, the alterations around the active site-encoding
domain probably reduce the affinity of PBP2 for penicillin G, thus accounting for the PenI
phenotype. However, the effects of penA alterations on the enzymatic activity of PBP2
(particularly the transpeptidation reaction involved in peptidoglycan biosynthesis) and the
involvement of such modifications in the PenI phenotype have not been addressed. The lack
of information concerning the structure of the N. meningitidis peptidoglycan has hindered this
analysis. We recently solved this structure using reverse-phase high pressure liquid
chromatography (HPLC) and mass spectrometry (Antignac et al., accompanied submitted
manuscript). The aim of this study was to characterize meningococcal strains with reduced
susceptibility to penicillin G and to study the functional consequences of PBP2 alterations. In
particular, we analyzed the modifications in the peptidoglycan structure in well defined
isogenic strains that only differed by the penA gene.
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
5
EXPERIMENTAL PROCEDURES
Bacterial strains and growth conditions – The meningococcal strains used in this study are
listed in Table I. N. meningitidis was grown for 18 h on GCB medium (Difco) containing
Kellogg supplements (20). E. coli TG1 was used as a host for cloning experiments and E. coli
BL21(DE3) pLysS was used as an expression system (21). E. coli cultures were grown in
Luria-Bertani medium supplemented with 40 µg/ml of kanamycin and 15 µg/ml of
chloramphenicol when necessary. Antibiotic susceptibility testing by the Etest method and
serological typing were performed as previously described (19,22,23). N. meningitidis strain
LNP8013 was transformed with either the entire penA gene or the 3’ end of the penA gene
amplified from PenI strains with oligonucleotides 99-14/99-23 and AA-18/99-23 (Table II),
respectively, as previously described (19). Transformants were selected on GCB medium
containing various concentrations of penicillin G.
Penicillin binding assays - Total meningococcal membrane extracts were prepared as
previously described (24). The crude membranes (30 µg of protein) were incubated for 30
min at 37°C in the presence of increasing concentrations of [3H]benzylpenicillin (20 Ci/mmol,
1 mCi/ml, Amersham Pharmacia Biotech). The reactions were stopped by adding 5 µl of
unlabeled penicillin G (2.5 mg/ml) for 20 min at room temperature. The binding affinity of
penicillin G to purified PBP2s was assayed by competition experiments with
[3H]benzylpenicillin. The purified PBP2s (2 µg of protein) were first incubated with
increasing concentrations of penicillin G, then with saturating concentrations of
[3H]benzylpenicillin and processed as described above. After SDS-PAGE analysis, the gels
were fixed for 30 min in 40% ethanol and 10% acetic acid, washed for 10 min with water and
placed in Amplify (Amersham Pharmacia Biotech) for 30 min. The gels were dried and used
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
6
to expose Hyperfilm-MP (Amersham Pharmacia Biotech) at - 80°C. The signals were
quantified using the ImageQuant program (Molecular Dynamics).
Infrared spectroscopy - Cells that had been grown for 18 hours were harvested by
centrifugation and resuspended in distilled water. The optical densities of the suspensions
were adjusted to yield optimal infrared spectra. A 35 µl aliquot of cells and a 20 µl aliquot of
purified peptidoglycan preparations were transferred to a ZnSe optical plate (sample holder),
vacuum dried, and then used directly for Fourier-transform infrared (FT-IR) spectroscopy.
Each sample was measured in five independent assays. All spectra were recorded between
wave numbers 4000 and 500 cm-1 with an IFS 28/B FT-IR spectrometer (Bruker, Karlsruhe,
Germany). The OPUS software, version 3.02 (Bruker), was used to process the data.
Determination of peptidoglycan structure - Peptidoglycan preparations, reverse-phase HPLC
analysis, desalting of muropeptides and mass spectrometry were performed as previously
described (Antignac et al., accompanied submitted manuscript). Briefly, peptidoglycans were
isolated by an adapted version of the method developed for E. coli (25) with boiling sodium
dodecyl sulfate, digested with muramidase from Streptomyces globisporus and reduced with
sodium borohydride. Muropeptides were separated and desalted by HPLC on a Hypersil ODS
column (ThermoHypersil-Keystone) and submitted to mass spectrometry analysis. All matrix-
assisted laser desorption ionization mass spectrometry (MALDI-MS) and post source decay
(PSD) experiments were carried out on a Voyager DE STR instrument (Applied Biosystems
Inc., Framingham, MA) as previously described (Antignac et al., accompanied submitted
manuscript).
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
7
Molecular typing - Strains were typed by multilocus DNA fingerprinting (MLDF) using five
genes (pilA, pilD, crgA, regF, and igA) as previously described (26-28). Polymorphisms of the
penA and ponA genes were analyzed by RFLP as previously described (19). The ponA gene
was amplified by PCR using oligonucleotides AA-6 and AA-7 (Table II). Amplicons were
then digested with HpaII and analyzed on polyacrylamide gels. The nucleotide sequences of
the penA gene were analyzed as previously described (19).
Purification of recombinant PBP2s - penA’ codes for a water-soluble form of PBP2 lacking
the N-terminal transmembrane domain. penA’ from strains with different penA alleles
(LNP8013, LNP16519, LNP17041, LNP16454, LNP17723, and LNP18425) was PCR-
amplified with oligonucleotides AA-4 (with 5’ BsaI adaptor) and AA-2 (with 5’ XhoI adaptor)
(Table II). These resulting fragments were digested with BsaI and XhoI and ligated into the
NcoI and XhoI sites of pET28b (Novagen) to create genes encoding PBP2s with six C-
terminal histidine residues (His6-tagged PBP2s) expressed under the control of the T7
bacteriophage promoter. These recombinant plasmids were expressed in E. coli BL21(DE3)
pLysS and protein purification was performed using a Ni-NTA agarose column (Qiagen) as
previously described (29). The fractions containing PBP2 were pooled, dialyzed against PBS,
adjusted to 20% glycerol (vol/vol) and stored at – 20°C.
Construction of penA-lacZ transcriptional fusions - The entire penA coding sequence (CDS)
from strain LNP8013 (penA1 allele) was amplified with oligonucleotides 99-1 and 99-2
(Table II) and cloned into pGEM-T Easy (Promega) to obtain the recombinant plasmid pAA3.
A 400-bp fragment located immediately downstream from the penA gene on the
meningococcal chromosome was amplified with oligonucleotides AA-13 (with a 5’ NdeI
adaptor) and AA-14 (with a 5’ SacI adaptor) (Table II). The resulting fragment was cloned
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
8
between the NdeI and SacI sites located downstream from the penA gene in pAA3. A
promotorless lacZ gene (30) was introduced into the SpeI site located downstream from the
penA gene. The aph-3’ gene, encoding kanamycin resistance (31), was amplified with
oligonucleotides KM-6 and KM-7 (Table II), and inserted into the blunt-ended HindIII site at
the end of the lacZ gene. This recombinant plasmid, named pAA8, harbors the penA-lacZ-
aph-3’ operon (Fig. 1A) and was used to transform meningococcal PenS strain LNP8013
(penA1) and PenI strains LNP16969 (penA21) and LNP16635 (penA11) (Table I). Upon
transformation, the recombinant plasmid pAA8 is not maintained and double recombination
occurs. PCR and RFLP analysis were used to select transformants in which lacZ and aph-3’
genes had been integrated correctly, but in which the original penA allele in the chromosome
had been preserved in each strain. β-galactosidase assays were performed as previously
described (32).
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
9
RESULTS
Characterization of meningococcal strains - We first selected a subset of well defined
meningococcal clinical isolates including ten PenI strains and one PenS strain (Table I). We
determined the serogroup, serotype and serosubtype of each strain. The MICs of penicillin G,
amoxicillin, and cefotaxime were determined by the Etest method. The MLDF approach using
five genes (pilA, pilD, crgA, regF, and igA) showed that strains belonged to different genetic
lineages (Table I). RFLP analysis showed that the penA allele of all PenI strains differed from
that of the strain LNP8013 (PenS) (Table I). As alterations of both PBP2 and PBP1 are
associated with an increased level of penicillin resistance in N. gonorrhoeae (12), we also
used RFLP to analyze the ponA polymorphism in sixty-six PenS and PenI meningococal
strains. Several restriction enzymes were tested and only digestion with HpaII gave four
highly related ponA alleles, but no association was observed between ponA alleles and
PenS/PenI phenotypes (Table I and data not shown). It has previously been shown that the
disappearance of PstI site in the ponA gene is correlated with penicillin resistance in
gonococci (12). However, this polymorphism was not found in any of our meningococcal
strains (data not shown).
We next transformed PCR-amplified penA alleles from eight PenI strains into the PenS
strain LNP8013 to generate a set of isogenic meningococcal variants that differed only by
penA alleles. All the transformants (TR) acquired the penA allele from the donor strain and
showed reduced susceptibility to penicillin G, with an MIC greater than 0.125 µg/ml (Table
I).
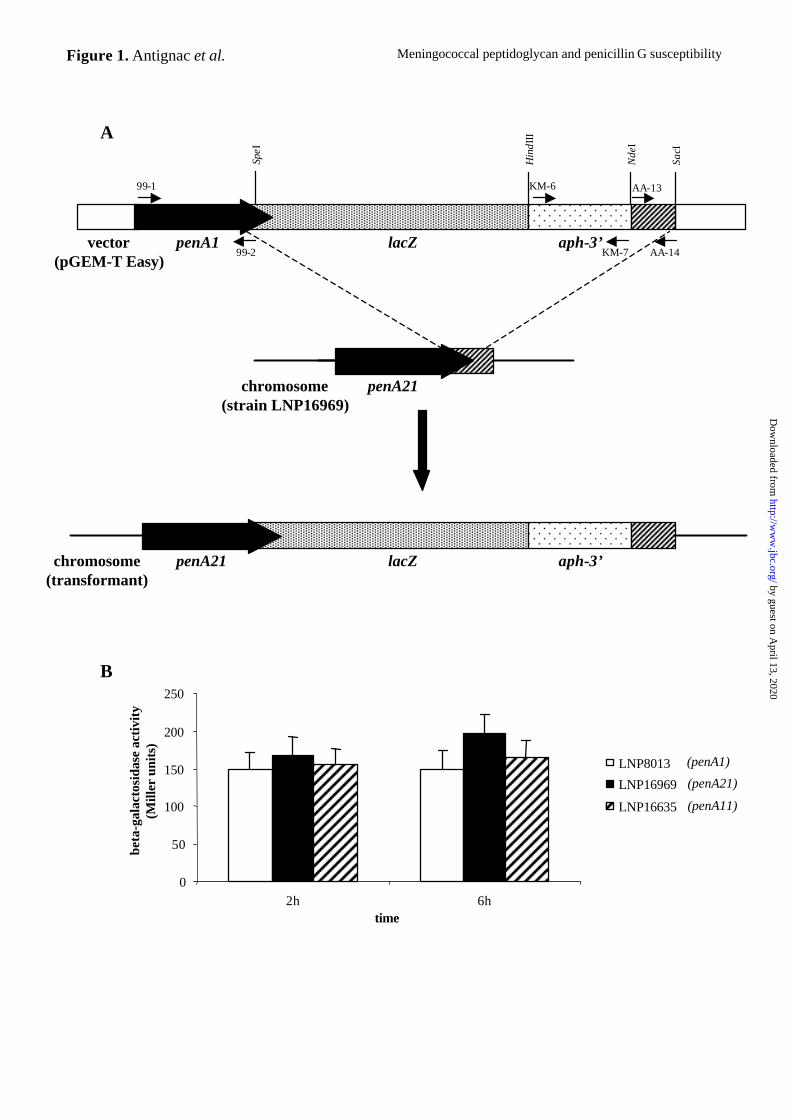
Effects of penA alterations on its own expression - We studied the expression of different
penA alleles by analyzing the expression of penA-lacZ transcriptional fusions. The penA CDS
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
10
from strain LNP8013 was cloned upstream from a promoterless lacZ gene. The resulting
recombinant plasmid, pAA8, also harbored the aph-3’ gene encoding resistance to kanamycin
and the region of the meningococcal chromosome located immediately downstream from the
penA gene (Fig. 1A). Transformation and allelic replacement into the PenS strain, LNP8013,
and two PenI strains, LNP16969 and LNP16635, allowed us to construct strains expressing
lacZ under the control of the promoter region of the chromosomal penA1, penA21 and penA11
genes, respectively (Table I). The β-galactosidase activity reflected the transcription of these
alleles. No significant differences were observed in β-galactosidase activities of the PenS and
PenI strains in our experimental conditions (Fig. 1B). These results show that penA alterations
do not affect the transcription of the penA gene and suggest that the transcriptional level of
penA does not play a role in the PenI phenotype.
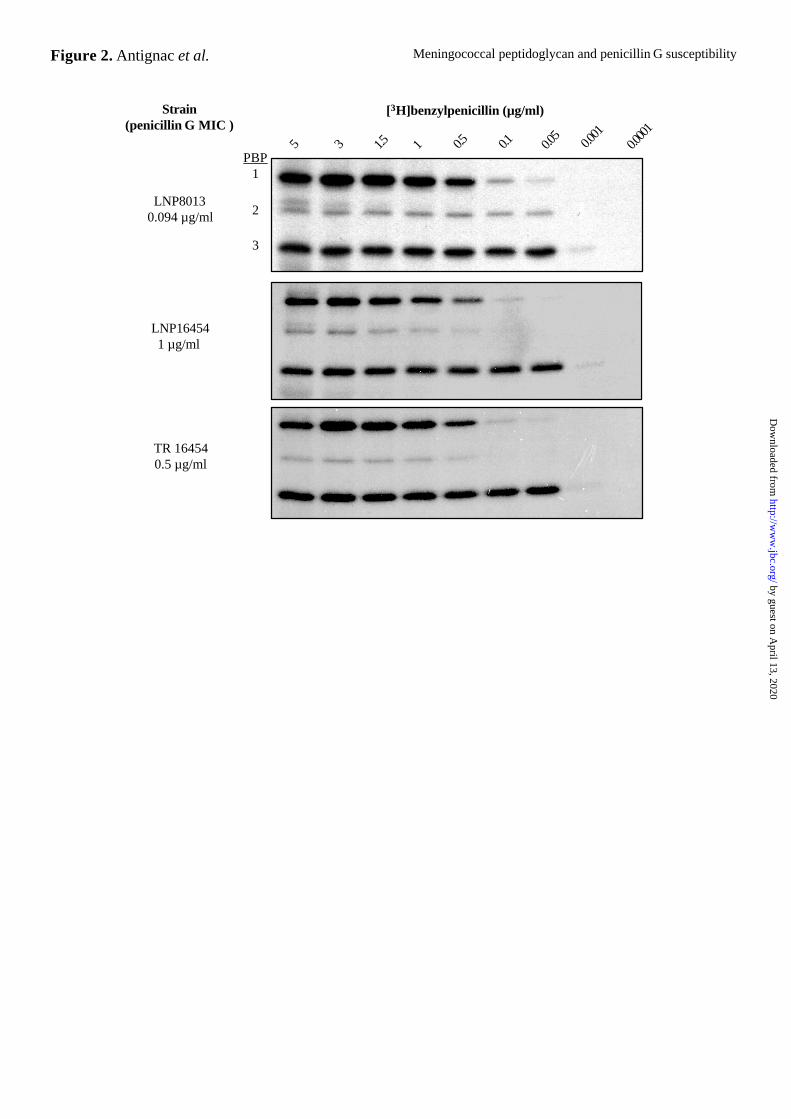
Effects of penA alterations on binding of penicillin to PBP2 – We found that ten times less
[3H]benzylpenicillin bound to PBP2s from all PenI strains than to PBP2 from the PenS strain
in crude membranes (PBP2s from PenI and PenS strains were saturated by 0.5 µg/ml and 0.05
µg/ml [3H]benzylpenicillin, respectively) (Fig. 2). There was no apparent difference in the
amount of [3H]benzylpenicillin bound to the other PBPs (PBP1 and PBP3) between the two
types of strain, even in strains with higher MICs (saturation at 0.5 µg/ml and 0.05 µg/ml
[3H]benzylpenicillin for PBP1 and PBP3, respectively). In transformants, the acquisition of
altered penA genes and reduced susceptibility to penicillin G were accompanied by changes in
the binding of [3H]benzylpenicillin to PBP2s, as in the donor strains, but not to PBP1 and
PBP3 (Fig. 2).
We next studied the binding of penicillin G to purified PBP2s from PenS and PenI
strains. penA alleles lacking the transmembrane-coding region (penA’) were amplified and
cloned into pET28b (Novagen), creating genes coding for water-soluble forms of PBP2s with
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
11
six C-terminal histidine residues. His6-tagged PBP2s were overproduced and purified to
greater than 95% purity (data not shown). We then determined the binding affinity of the
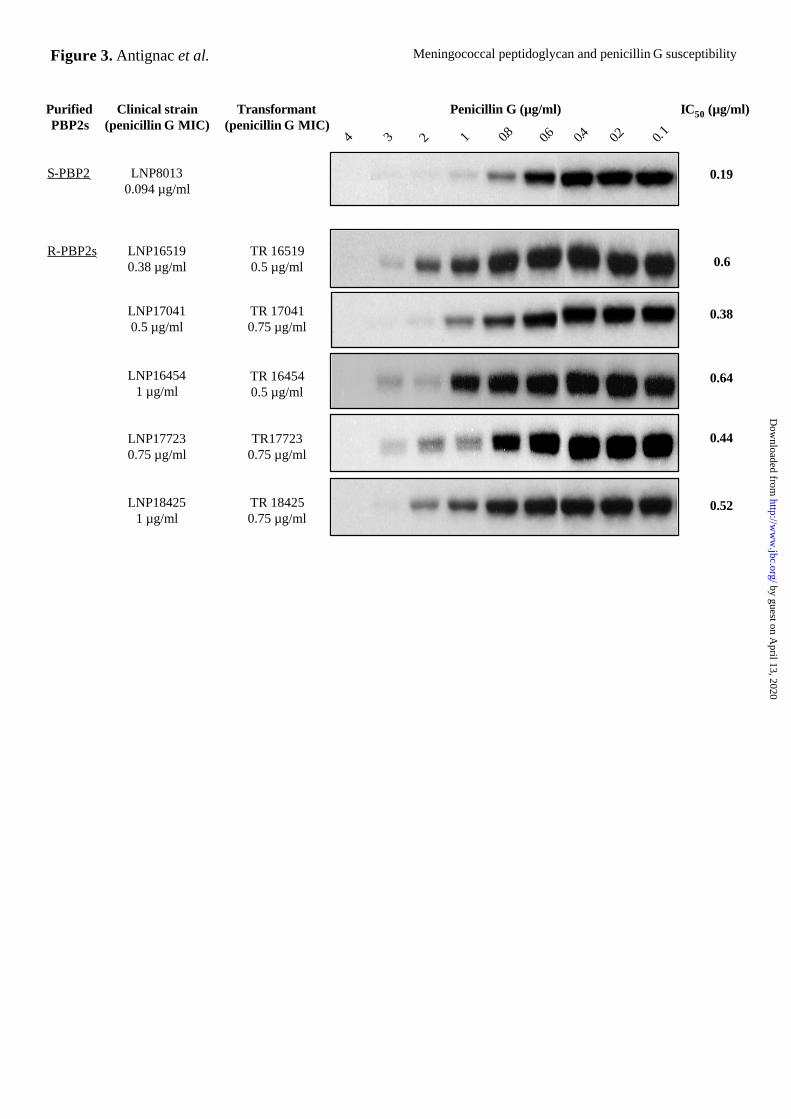
purified PBP2s for penicillin G in competition experiments with [3H]benzylpenicillin. We
tested purified PBP2s from one PenS (S-PBP2) and five PenI (R-PBP2s) strains with different
penicillin G MICs. The binding was scored by determining the concentration of non
radioactive penicillin G that inhibited the binding of saturating concentrations of
[3H]benzypenicillin by 50% (inhibition concentration 50, IC50). R-PBP2s bound less
penicillin G than S-PBP2. The IC50 values and the penicillin G MICs of the corresponding
PenI transformants matched closely. However, no strict correlation was found between the
IC50 values and the penicillin G MICs of the corresponding clinical PenI strains (Fig. 3).
These data strongly suggest that alterations in the penA genes resulted in PBP2s with
decreased affinity for penicillin G. However, the decreased affinity of PBP2s for penicillin G
cannot solely explain the different penicillin G MICs of the clinical PenI isolates.
Effects of penA alterations on the structure of meningococcal PBP2 - We sequenced the
region of the penA gene that encodes the transpeptidase domain of PBP2 of a large collection
of PenI clinical isolates obtained in France, England and Spain as previously described (19).
The deduced amino acid sequences of PBP2 (amino acids 298-581) were aligned and
meningococcal PenI strains showed 8.5 to 14.4% divergence (data not shown). Eight positions
were modified in almost all PenI strains (Fig. 4). These altered positions were located around
the conserved KTG motif. The corresponding residues in PBP2x from S. pneumoniae are part
of the structure that forms the active site (33), particularly the β-sheets 3 and 4 (Fig. 4). This
suggests that these amino acid substitutions are important in determining reduced affinity of
PBP2 for penicillin G. We transformed the PenS strain LNP8013 with a PCR-amplified DNA
fragment corresponding to the 3’ end of the penA gene harboring these eight modifications.
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
12
All the transformants obtained replaced their 3’ end of the penA gene by the transforming
fragment and displayed reduced susceptibility to penicillin G with an MIC ranging from 0.38
to 1 µg/ml. Therefore, this penA fragment was able to confer resistance to a PenS strain,
suggesting that only a small number of amino acid changes are important for decreasing the
affinity of meningococcal PBP2 for penicillin G.
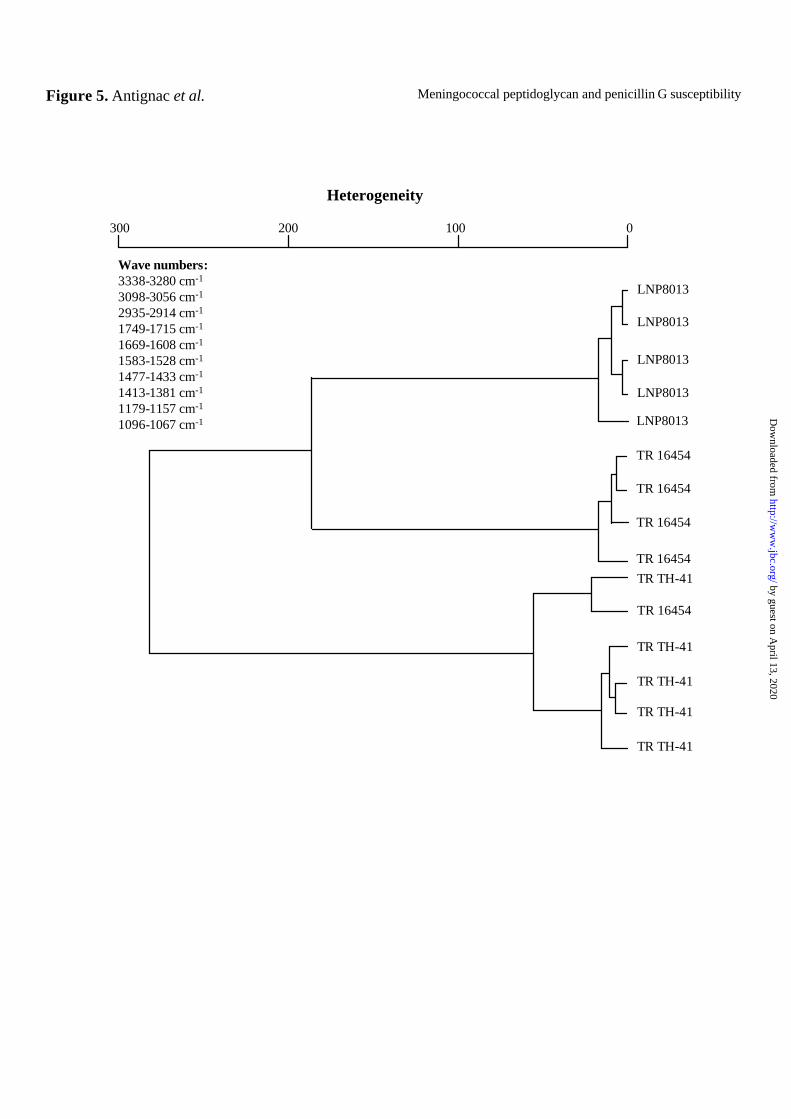
Association between bacterial cell wall alterations and PBP2 modifications - Infrared (FT-
IR) spectroscopy can be used to analyze bacterial cell walls and peptidoglycans. Infrared
spectroscopy has been used to tentatively assign bacterial molecules according to their
vibrational features (34). Spectra for the intact PenS strain (LNP8013) and its isogenic PenI
derivatives (TR 16454, TR TH-41) were recorded and compared between wave numbers 4000
and 500 cm-1. Variations in peptidoglycan structure can be detected by comparing infrared
absorption in the wave numbers regions corresponding to amides and sugar rings bands.
Significant differences were found between amides and carbohydrates absorption bands of the
PenS and isogenic PenI strains (Fig. 5). We calculated the heterogeneity index of absorption at
several wave numbers and used it to construct a dendrogram. This showed that the isogenic
PenI derivatives clustered together and separately from the PenS parental strain (Fig. 5).
Identical results were obtained when we compared purified peptidoglycan preparations from
these strains (data not shown). These data suggest that penA alterations affect the
meningococcal cell wall and possibly the peptidoglycan.
Peptidoglycan structure in meningococcal strains - We recently described the structure of
the peptidoglycan of N. meningitidis, which consists of disaccharide subunits of N-
acetylglucosamine (GlcNAc) and N-acetylmuramic acid (MurNAc) carrying a pentapeptide
(L-Ala-D-Glu-meso-A2pm-D-Ala-D-Ala) (Antignac et al., accompanied submitted
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
13
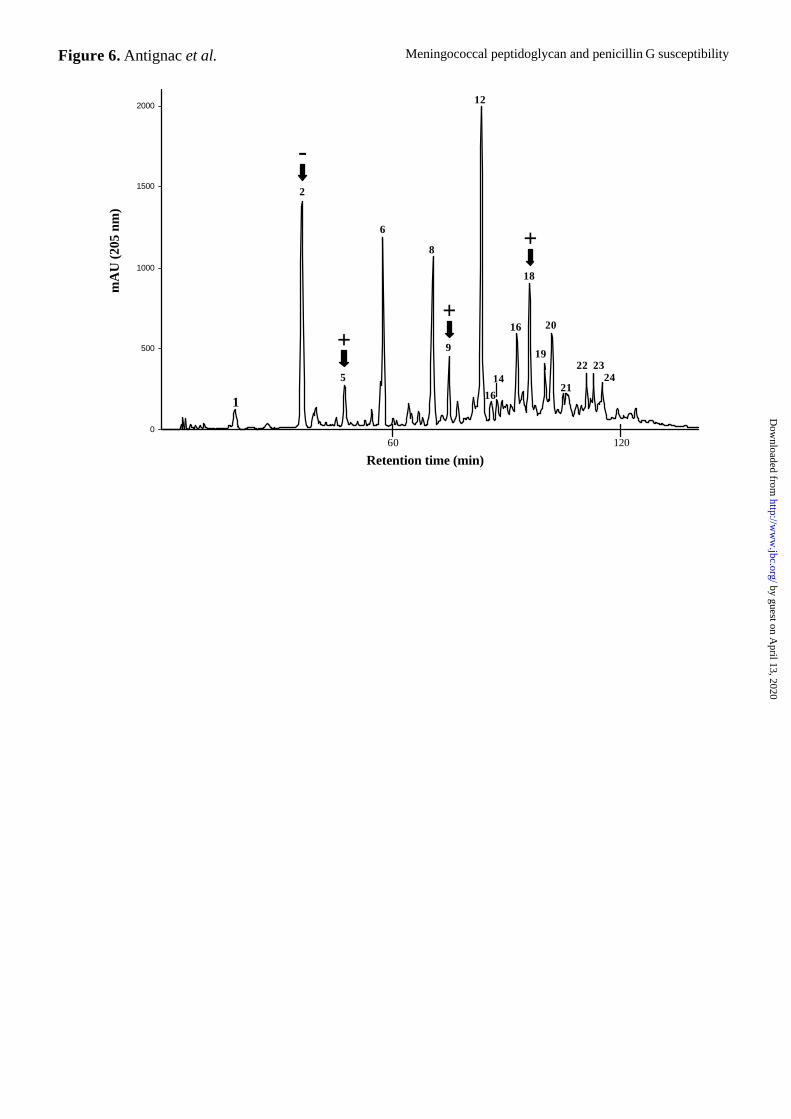
manuscript). To determine the nature of the peptidoglycan structural modifications that could
be induced by penA alterations, we compared the muropeptide composition of the PenS strain
LNP8013 and its isogenic PenI derivatives (Table I). We analyzed purified peptidoglycan
preparations digested with muramidase by HPLC and mass spectrometry. We determined the
structures of 28 different muropeptides species (Antignac et al., accompanied submitted
manuscript) (Fig. 6 and Table III). While no new peaks were detected, differences in the
amplitudes of preexisting peaks were observed between the PenS strain and its seven isogenic
PenI derivatives. These mostly involved an increase in pentapeptide-containing muropeptides
(Fig. 6 and Table III). The amplitude of peak 5, which corresponds to the disaccharide
pentapeptide GlcNAc-MurNAc-L-Ala-D-Glu-meso-A2pm-D-Ala-D-Ala, was significantly
higher in all the isogenic PenI strains than in LNP8013. The amplitudes of peak 9, which
corresponds to the tetra-penta dimer and peak 18, which contains its di-O-acetylated
derivative, were also higher in the isogenic PenI strains. A minor decrease in the amplitude of
peak 2 (tetra monomer) was observed in the isogenic penI strains. The same accumulation of
muropeptides carrying pentapeptide chains in peptidoglycan was also observed when the
parent clinical PenI isolates were compared with strain LNP8013 (Antignac et al.,
accompanied submitted manuscript). Our results suggest that the peptidoglycans from PenS
and the isogenic PenI strains differ. These changes were transformable using penA gene from
a donor PenI strain to a recipient PenS strain and are hence directly linked to penA alterations.
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
14
DISCUSSION
Susceptibility to β-lactams antibiotics can be mediated by three major mechanisms: (i)
decreased binding of antibiotics to PBPs, (ii) antibiotic inactivation by β-lactamase and (iii)
reduced membrane permeability. In N. meningitidis, the first mechanism is usually
incriminated. SDS-PAGE analysis revealed the presence of three PBPs in this bacterium, but
other PBPs may also exist, as suggested from the in silico analysis of the genome of two
completely sequenced meningococcal strains (3,4). We have previously reported that penA
alterations are directly linked to the PenI phenotype (19). Moreover in this study, we showed
that the polymorphism in the ponA gene, encoding PBP1, is not associated with the PenI
phenotype. Our results confirm that alterations of the penA gene, encoding PBP2, decrease the
binding affinity of PBP2 for penicillin G. However, this decreased affinity for penicillin G
may not be the only mechanism accounting for the increased MIC of this antibiotic, as the
binding of penicillin G to purified PBP2s from different clinical PenI strains was not directly
correlated with penicillin G MIC levels (Fig. 3). Penicillin G binds covalently to the active
site of PBPs (35), therefore we expected that the ability of altered PBP2s to bind penicillin
would be affected, as alterations of PBP2s are usually located near to the active site of the
transpeptidase moiety of PBP2. Moreover, the eight residues modified in most PenI isolates
were in the region of the meningococcal PBP2 that is homologous to the active site of PBP2x
from S. pneumoniae (33) and seem to play a major role in the decreased affinity of PBP2 for
penicillin G.
Since PBPs are involved in the late stages of peptidoglycan biosynthesis, their
alterations associated with β-lactam resistance might result in peptidoglycan modifications.
Peptidoglycan is a heteropolymer that is composed of several layers of linear glycan chains of
repeated disaccharide units, N-acetylglucosamine-N-acetylmuramic acid (GlcNAc-MurNAc),
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
15
that are interconnected by peptide cross-links. Peptidoglycan forms a barrier in the cell wall
that preserves cell integrity and shape. In enterococci, staphylococci and pneumococci, altered
PBPs seem to form unusual cross-links, suggesting that the specificity of their transpeptidase
activity is altered (36-40). In S. pneumoniae, resistance to penicillin G occurs in several steps
involving modifications of several PBPs (41,42) and it has been suggested that changes in
peptidoglycan structure are closely related with resistance to β-lactams (43-45). The
peptidoglycan of non-β-lactamase-mediated antibiotic resistant strains of Haemophilus
influenzae has been shown to contain more tripeptides than susceptible strains (46). This may
be due to an increase in the activity of a D,D-carboxypeptidase that would generate more
tripeptides (46). Moreover, the cell wall of H. influenzae resistant strains showed enhanced
inflammatory activity in an animal model of meningitis (47). We recently developed a murine
model that should make it possible to study the effect of modifications in N. meningitidis
peptidoglycan structure on the induction of inflammatory response as well as on
meningococcal virulence (48).
It has been suggested that the degree of cross-linking differs between PenS and PenI
strains of N. gonorrhoeae (49,50), which is closely related to N. meningitidis. However,
isogenic derivatives were not used in this work and HPLC peaks were not characterized (51).
We have shown (Antignac et al., accompanied submitted manuscript) that the reduced
susceptibility to penicillin G of meningococcal PenI clinical isolates was associated with
changes in peptidoglycan structure (accumulation of pentapeptides). Here, we studied the
peptidoglycan structure of several meningococcal isogenic PenI strains obtained by
transformation of the parental susceptible strain with PCR-amplified altered penA genes. We
did not observe any new peak in the HPLC profiles. However, relative changes in the
amplitudes of preexisting peaks were observed. As previously observed in clinical PenI
isolates (Antignac et al., accompanied submitted manuscript), our data indicated an increase
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
16
in muropeptides with pentapeptide chains. As we compared isogenic strains, these
modifications in peptidoglycan structure are directly linked to the alterations in penA gene and
changes in another locus cannot be involved, suggesting that the modified PBP2 in PenI
transformants may have an altered D,D-transpeptidase activity. However, the increase in
muropeptides carrying pentapeptide chains may reflect a decrease in D,D-carboxypeptidase
activity on newly synthesized peptidoglycan chains. This in turn suggests that the
meningococcal PBP2 rather has D,D-carboxypeptidase activity. Most of the modifications
affecting PBP2s from PenI strains were located within a region that is homologous to the
active site of PBP2x from S. pneumoniae (Fig. 4). In fact, these modified sites line the active-
site pocket and therefore are most likely to affect the enzymatic activity of PBP2 (D,D-
transpeptidase and/or D,D-carboxypeptidase). Recently, similar alterations in PBP3 of H.
influenzae have been shown to be associated with β-lactam resistance (52,53). Furthermore,
the same region has been implicated in E. coli PBP5 role in cell morphology and D,D-
carboxypeptidase activity (54).
Genomic analysis of the complete sequence of two strains of N. meningitidis (3,4)
revealed putative genes encoding D,D-carboxypeptidase and endopeptidase. Meningococcal
PBP2 has been shown to interact with the lytic transglycosylase MltA (55). Furthermore, the
E. coli homolog of meningococcal PBP2, PBP3, can form a complex with the lytic
transglycosylase Slt70 and the low molecular weight PBP7/8 (56). Thus, the reduced D,D-
carboxypeptidase activity observed in the isogenic PenI strains could be an indirect effect. The
ability of altered meningococcal PBP2s to engage in protein-protein interactions with the
putative D,D-carboxypeptidase would be affected, leading to the observed phenotype in PenI
strains. However, this hypothesis is less likely as the amino acid modifications are all
predicted to be located in the active-site pocket of meningococcal PBP2, although it is
possible that they change its overall three-dimensional structure. The construction of mutants
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
17
in these genes may help us to understand the effect of penA alterations on the structure of the
N. meningitidis peptidoglycan.
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
18
REFERENCES
1. Barbour, A. G. (1981) Antimicrob Agents Chemother 19, 316-322 2. Dougherty, T. J., Koller, A. E., and Tomasz, A. (1981) Antimicrob Agents Chemother
20, 109-114 3. Tettelin, H., Saunders, N. J., Heidelberg, J., Jeffries, A. C., Nelson, K. E., Eisen, J. A.,
Ketchum, K. A., Hood, D. W., Peden, J. F., Dodson, R. J., Nelson, W. C., Gwinn, M. L., DeBoy, R., Peterson, J. D., Hickey, E. K., Haft, D. H., Salzberg, S. L., White, O., Fleischmann, R. D., Dougherty, B. A., Mason, T., Ciecko, A., Parksey, D. S., Blair, E., Cittone, H., Clark, E. B., Cotton, M. D., Utterback, T. R., Khouri, H., Qin, H., Vamathevan, J., Gill, J., Scarlato, V., Masignani, V., Pizza, M., Grandi, G., Sun, L., Smith, H. O., Fraser, C. M., Moxon, E. R., Rappuoli, R., and Venter, J. C. (2000) Science 287, 1809-1815
4. Parkhill, J., Achtman, M., James, K. D., Bentley, S. D., Churcher, C., Klee, S. R., Morelli, G., Basham, D., Brown, D., Chillingworth, T., Davies, R. M., Davis, P., Devlin, K., Feltwell, T., Hamlin, N., Holroyd, S., Jagels, K., Leather, S., Moule, S., Mungall, K., Quail, M. A., Rajandream, M. A., Rutherford, K. M., Simmonds, M., Skelton, J., Whitehead, S., Spratt, B. G., and Barrell, B. G. (2000) Nature 404, 502-506
5. Fontanals, D., Pineda, V., Pons, I., and Rojo, J. C. (1989) Eur J Clin Microbiol Infect Dis 8, 90-91
6. Dillon, J. R., Pauze, M., and Yeung, K. H. (1983) Lancet 1, 779-781 7. Vazquez, J. A., Enriquez, A. M., De la Fuente, L., Berron, S., and Baquero, M. (1996)
Eur J Clin Microbiol Infect Dis 15, 181-182 8. Botha, P. (1988) Lancet 1, 54 9. Oppenheim, B. A. (1997) Clin Infect Dis 24 Suppl 1, S98-101 10. Spratt, B. G. (1988) Nature 332, 173-176 11. Mendelman, P. M., Campos, J., Chaffin, D. O., Serfass, D. A., Smith, A. L., and Saez-
Nieto, J. A. (1988) Antimicrob Agents Chemother 32, 706-709 12. Ropp, P. A., Hu, M., Olesky, M., and Nicholas, R. A. (2002) Antimicrob Agents
Chemother 46, 769-777 13. Feil, E. J., Maiden, M. C., Achtman, M., and Spratt, B. G. (1999) Mol Biol Evol 16,
1496-1502 14. Holmes, E. C., Urwin, R., and Maiden, M. C. (1999) Mol Biol Evol 16, 741-749 15. Spratt, B. G., Zhang, Q. Y., Jones, D. M., Hutchison, A., Brannigan, J. A., and
Dowson, C. G. (1989) Proc Natl Acad Sci U S A 86, 8988-8992 16. Spratt, B. G., Bowler, L. D., Zhang, Q. Y., Zhou, J., and Smith, J. M. (1992) J Mol
Evol 34, 115-125 17. Saez-Nieto, J. A., Lujan, R., Martinez-Suarez, J. V., Berron, S., Vazquez, J. A., Vinas,
M., and Campos, J. (1990) Antimicrob Agents Chemother 34, 2269-2272 18. Bowler, L. D., Zhang, Q. Y., Riou, J. Y., and Spratt, B. G. (1994) J Bacteriol 176,
333-337 19. Antignac, A., Kriz, P., Tzanakaki, G., Alonso, J. M., and Taha, M. K. (2001) J
Antimicrob Chemother 47, 285-296 20. Kellogg, D. S., Peacock, W. L., Deacon, W. E., Brown, L., and Pirkle, C. I. (1963) J
Bacteriol 85, 1274-1279 21. Studier, F. W., Rosenberg, A. H., Dunn, J. J., and Dubendorff, J. W. (1990) Methods
Enzymol 185, 60-89 22. Abdillahi, H., and Poolman, J. T. (1987) FEMS Microbiol Lett 48, 367-371
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
19
23. Frasch, C. E., Zollinger, W. D., and Poolman, J. T. (1985) Rev Infect Dis 7, 504-510 24. Lujan, R., Zhang, Q. Y., Saez Nieto, J. A., Jones, D. M., and Spratt, B. G. (1991)
Antimicrob Agents Chemother 35, 300-304 25. Glauner, B., Holtje, J. V., and Schwarz, U. (1988) J Biol Chem 263, 10088-10095 26. Guibourdenche, M., Giorgini, D., Gueye, A., Larribe, M., Riou, J. Y., and Taha, M. K.
(1997) J Clin Microbiol 35, 745-750 27. Kriz, P., Giorgini, D., Musilek, M., Larribe, M., and Taha, M. K. (1999) Res
Microbiol 150, 273-280 28. Mayer, L. W., Reeves, M. W., Al-Hamdan, N., Sacchi, C. T., Taha, M. K., Ajello, G.
W., Schmink, S. E., Noble, C. A., Tondella, M. L., Whitney, A. M., Al-Mazrou, Y., Al-Jefri, M., Mishkhis, A., Sabban, S., Caugant, D. A., Lingappa, J., Rosenstein, N. E., and Popovic, T. (2002) J Infect Dis 185, 1596-1605
29. Deghmane, A. E., Petit, S., Topilko, A., Pereira, Y., Giorgini, D., Larribe, M., and Taha, M. K. (2000) Embo J 19, 1068-1078
30. Perkins, J. B., and Youngman, P. J. (1986) Proc Natl Acad Sci U S A 83, 140-144 31. Taha, M. K., Morand, P. C., Pereira, Y., Eugene, E., Giorgini, D., Larribe, M., and
Nassif, X. (1998) Mol Microbiol 28, 1153-1163 32. Miller, J. H. (1972) Experiments in Molecular Genetics. Cold Spring Harbor
Laboratory, Cold Spring Harbor, NY 33. Dessen, A., Mouz, N., Gordon, E., Hopkins, J., and Dideberg, O. (2001) J Biol Chem
276, 45106-45112 34. Naumann, D., Barnickel, G., Bradaczek, H., Labischinski, H., and Giesbrecht, P.
(1982) Eur J Biochem 125, 505-515 35. Hakenbeck, R., and Coyette, J. (1998) Cell Mol Life Sci 54, 332-340 36. van Heijenoort, J., and Gutmann, L. (2000) Proc Natl Acad Sci U S A 97, 5028-5030 37. Garcia-Bustos, J., and Tomasz, A. (1990) Proc Natl Acad Sci U S A 87, 5415-5419 38. Mainardi, J. L., Legrand, R., Arthur, M., Schoot, B., van Heijenoort, J., and Gutmann,
L. (2000) J Biol Chem 275, 16490-16496 39. Mainardi, J. L., Morel, V., Fourgeaud, M., Cremniter, J., Blanot, D., Legrand, R.,
Frehel, C., Arthur, M., Van Heijenoort, J., and Gutmann, L. (2002) J Biol Chem 277, 35801-35807
40. Boneca, I. G., Xu, N., Gage, D. A., de Jonge, B. L., and Tomasz, A. (1997) J Biol Chem 272, 29053-29059
41. Hakenbeck, R. (1998) Electrophoresis 19, 597-601 42. Tomasz, A. (1997) Clin Infect Dis 24 Suppl 1, S85-88 43. Filipe, S. R., and Tomasz, A. (2000) Proc Natl Acad Sci U S A 97, 4891-4896 44. Severin, A., Figueiredo, A. M., and Tomasz, A. (1996) J Bacteriol 178, 1788-1792 45. Severin, A., and Tomasz, A. (1996) J Bacteriol 178, 168-174 46. Burroughs, M. H., Chang, Y. S., Gage, D. A., and Tuomanen, E. I. (1993) J Biol
Chem 268, 11594-11598 47. Burroughs, M., Prasad, S., Cabellos, C., Mendelman, P. M., and Tuomanen, E. (1993)
J Infect Dis 167, 464-468 48. Alonso, J. M., Guiyoule, A., Zarantonelli, L., Ramisse, F., Pirès, R., Antignac, A.,
Deghmane, A. E., Huerre, M., van der Werf, S., and Taha, M. K. (2003) FEMS Microbiol Lett (in press)
49. Garcia-Bustos, J. F., and Dougherty, T. J. (1987) Antimicrob Agents Chemother 31, 178-182
50. Dougherty, T. J. (1985) Antimicrob Agents Chemother 28, 90-95 51. Dougherty, T. J. (1985) J Bacteriol 163, 69-74
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
20
52. Dabernat, H., Delmas, C., Seguy, M., Pelissier, R., Faucon, G., Bennamani, S., and Pasquier, C. (2002) Antimicrob Agents Chemother 46, 2208-2218
53. Ubukata, K., Shibasaki, Y., Yamamoto, K., Chiba, N., Hasegawa, K., Takeuchi, Y., Sunakawa, K., Inoue, M., and Konno, M. (2001) Antimicrob Agents Chemother 45, 1693-1699
54. Ghosh, A. S., and Young, K. D. (2003) J Bacteriol 185, 2178-2186 55. Jennings, G. T., Savino, S., Marchetti, E., Arico, B., Kast, T., Baldi, L., Ursinus, A.,
Holtje, J. V., Nicholas, R. A., Rappuoli, R., and Grandi, G. (2002) Eur J Biochem 269, 3722-3731
56. Romeis, T., and Holtje, J. V. (1994) Eur J Biochem 224, 597-604
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
21
ABBREVIATIONS
Penicillin-susceptible, PenS; reduced susceptibility to penicillin, PenI; penicillin-binding
protein, PBP; high pressure liquid chromatography, HPLC; sodium dodecyl sulfate-
polyacrylamide gel electrophoresis, SDS-PAGE; minimum inhibitory concentration, MIC;
restriction fragment length polymorphism, RFLP; Fourier-transform infrared, FT-IR; matrix-
assisted laser desorption ionization mass spectrometry, MALDI-MS; post source decay, PSD;
multilocus DNA fingerprinting, MLDF; coding sequence, CDS; transformant, TR;
susceptible-PBP2, S-PBP2; resistant-PBP2, R-PBP2; inhibition concentration 50, IC50; N-
acetylglucosamine, GlcNAc; N-acetyl muramic acid, MurNAc; diaminopimelic acid, A2pm.
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
22
ACKNOWLEDGEMENTS
This work was supported by the Pasteur Institute and by a grant from the European
Commission number QLK2-CT-2001-01436. I.G.B. was the recipient of a postdoctoral
fellowship from the Fundação para a Ciência e a Tecnologia, Portugal
(SFRH/BPD/1567/2000).
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
23
FIGURE LEGENDS
Figure 1. Schematic representation of recombinant plasmid pAA8 containing the penA-
lacZ-aph3’ transcriptional fusion and recombination events with the homologous locus
on the meningococcal chromosome (A); β-galactosidase activity assays (B).
Oligonucleotides are represented by small arrows, white boxes correspond to DNA of the
vector pGEM-T easy, black boxes represent meningococcal penA gene and hatched boxes
stand for the meningococcal DNA fragment located downstream from the penA gene. The
recombinant plasmid pAA8 was used to transform N. meningitidis strains LNP8013 (PenS),
LNP16969 (PenI), and LNP16635 (PenI). lacZ was integrated by double crossing over (dotted
lines) downstream from each penA allele creating three transcriptional fusions penA1-lacZ
(strain LNP8013), penA21-lacZ (strain LNP16969) and penA11-lacZ (strain LNP16635). β-
galactosidase activities, which reflect the expression level of the three corresponding penA
alleles, were assayed after 2 h and 6 h of growth.
Figure 2. Binding affinity of PBPs for [3H]benzylpenicillin from one representative PenS
strain (LNP8013), PenI strain (LNP16454), and PenI transformant (TR 16454).
Membranes extracts (30 µg of protein) were incubated with the indicated concentrations of
[3H]benzylpenicillin. After SDS-PAGE analysis, the gels were fixed, treated with Amplify
(Amersham Pharmacia Biotech), and then autoradiographed. The positions of the three PBPs
(PBP1, PBP2 and PBP3) and the penicillin G MICs of each strain are indicated on the left.
Figure 3. Binding affinity of purified S-PBP2 (from PenS strain LNP8013) and R-PBP2s
(from PenI strains LNP16519, LNP17041, LNP16454, LNP17723, and LNP18425) for
penicillin G in competition experiments with [3H]benzylpenicillin. Purified PBP2s (2 µg
of protein) were first incubated with increasing concentrations of penicillin G, then with
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
24
saturating concentrations of [3H]benzylpenicillin. After SDS-PAGE analysis, the gels were
fixed, treated with Amplify (Amersham Pharmacia Biotech), and then autoradiographed.
Bands were quantified using the ImageQuant program (Molecular Dynamics). The
concentrations of penicillin G that inhibited the binding of saturating concentrations of
[3H]benzylpenicillin by 50% (IC50) are indicated on the right. The penicillin G MICs of the
clinical strains and of the corresponding transformants are indicated on the left.
Figure 4. Partial sequences of penicillin-binding protein 2 from meningococcal PenS
strain LNP8013 (Nm PBP2) and of penicillin-binding protein 2x from pneumococcal
PenS strain R6 (Sp PBP2x). The most frequent modified residues (8 residues) in the
sequences from meningococcal PenI strains and the corresponding amino acid changes are
indicated in bold in the sequence of Nm PBP2 (amino acids 443-581). The amino acid
positions associated with resistance in pneumococci are also indicated in bold in the sequence
of Sp PBP2x. The secondary structures (α-helices and β-sheets) of the PBP2x are indicated
according to Dessen et al. (33).
Figure 5. Dendrogram based on the heterogeneity of the infrared absorption spectra in
the wave numbers corresponding to amides (3338-3280 cm-1, 3098-3056 cm-1, 1749-1715
cm-1, 1669-1608 cm-1, 1583-1528 cm-1) and sugar rings bands (2935-2914 cm-1, 1477-1433
cm-1, 1413-1381 cm-1, 1179-1157 cm-1, and 1096-1067cm-1). Each strain (PenS strain
LNP8013 and its PenI isogenic derivatives TR TH-41 and TR 16545) was tested in quintuplet.
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
25
Figure 6. Reverse-phase HPLC analysis of N. meningitidis muramidase-digested
peptidoglycan from the PenS strain LNP8013. Arrows indicate the peaks that differ
between the PenS strain LNP8013 and the seven PenI isogenic strains. The amplitudes of
peaks 5, 9 and 18 were increased in PenI isogenic strains, whereas the amplitude of peak 2
was decreased. The structures of the muropeptides corresponding to these peaks that differ
quantitatively between the strain LNP8013 (PenS) and its isogenic PenI derivatives are
presented in Table III.
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from
Meningococcal peptidoglycan and penicillin G susceptibility
26
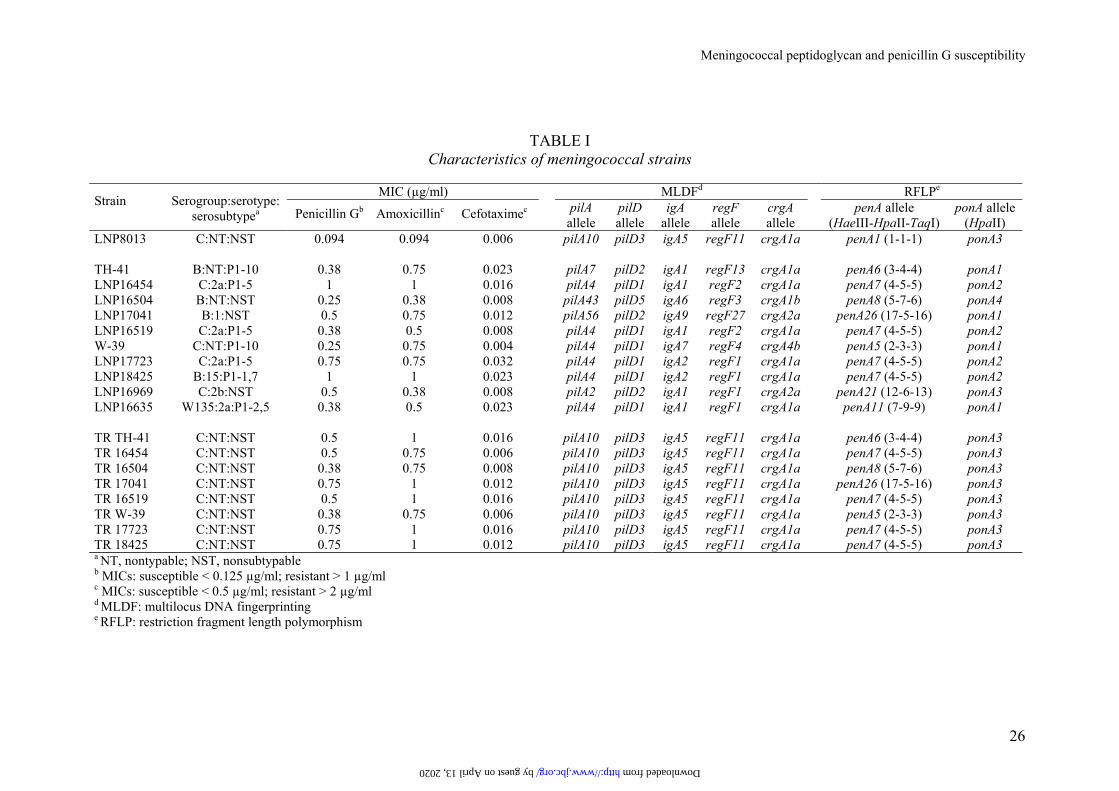
TABLE I Characteristics of meningococcal strains
MIC (µg/ml) MLDFd RFLPe
Strain
Serogroup:serotype: serosubtypea
Penicillin Gb
Amoxicillinc
Cefotaximec pilA allele
pilD allele
igA allele
regF allele
crgA allele
penA allele (HaeIII-HpaII-TaqI)
ponA allele (HpaII)
LNP8013 C:NT:NST 0.094 0.094 0.006 pilA10 pilD3 igA5 regF11 crgA1a penA1 (1-1-1) ponA3 TH-41 B:NT:P1-10 0.38 0.75 0.023 pilA7 pilD2 igA1 regF13 crgA1a penA6 (3-4-4) ponA1 LNP16454 C:2a:P1-5 1 1 0.016 pilA4 pilD1 igA1 regF2 crgA1a penA7 (4-5-5) ponA2 LNP16504 B:NT:NST 0.25 0.38 0.008 pilA43 pilD5 igA6 regF3 crgA1b penA8 (5-7-6) ponA4 LNP17041 B:1:NST 0.5 0.75 0.012 pilA56 pilD2 igA9 regF27 crgA2a penA26 (17-5-16) ponA1 LNP16519 C:2a:P1-5 0.38 0.5 0.008 pilA4 pilD1 igA1 regF2 crgA1a penA7 (4-5-5) ponA2 W-39 C:NT:P1-10 0.25 0.75 0.004 pilA4 pilD1 igA7 regF4 crgA4b penA5 (2-3-3) ponA1 LNP17723 C:2a:P1-5 0.75 0.75 0.032 pilA4 pilD1 igA2 regF1 crgA1a penA7 (4-5-5) ponA2 LNP18425 B:15:P1-1,7 1 1 0.023 pilA4 pilD1 igA2 regF1 crgA1a penA7 (4-5-5) ponA2 LNP16969 C:2b:NST 0.5 0.38 0.008 pilA2 pilD2 igA1 regF1 crgA2a penA21 (12-6-13) ponA3 LNP16635 W135:2a:P1-2,5 0.38 0.5 0.023 pilA4 pilD1 igA1 regF1 crgA1a penA11 (7-9-9) ponA1 TR TH-41 C:NT:NST 0.5 1 0.016 pilA10 pilD3 igA5 regF11 crgA1a penA6 (3-4-4) ponA3 TR 16454 C:NT:NST 0.5 0.75 0.006 pilA10 pilD3 igA5 regF11 crgA1a penA7 (4-5-5) ponA3 TR 16504 C:NT:NST 0.38 0.75 0.008 pilA10 pilD3 igA5 regF11 crgA1a penA8 (5-7-6) ponA3 TR 17041 C:NT:NST 0.75 1 0.012 pilA10 pilD3 igA5 regF11 crgA1a penA26 (17-5-16) ponA3 TR 16519 C:NT:NST 0.5 1 0.016 pilA10 pilD3 igA5 regF11 crgA1a penA7 (4-5-5) ponA3 TR W-39 C:NT:NST 0.38 0.75 0.006 pilA10 pilD3 igA5 regF11 crgA1a penA5 (2-3-3) ponA3 TR 17723 C:NT:NST 0.75 1 0.016 pilA10 pilD3 igA5 regF11 crgA1a penA7 (4-5-5) ponA3 TR 18425 C:NT:NST 0.75 1 0.012 pilA10 pilD3 igA5 regF11 crgA1a penA7 (4-5-5) ponA3 a NT, nontypable; NST, nonsubtypable b MICs: susceptible < 0.125 µg/ml; resistant > 1 µg/ml c MICs: susceptible < 0.5 µg/ml; resistant > 2 µg/ml d MLDF: multilocus DNA fingerprinting e RFLP: restriction fragment length polymorphism
by guest on April 13, 2020 http://www.jbc.org/ Downloaded from

Meningococcal peptidoglycan and penicillin G susceptibility
27
TABLE II Oligonucleotides used in this study
Oligonucleotide Sequence Relevant characteristics
AA-4 5’-GCTGGTCTCCCATGCTGATTGCCCGCGGGCTG-3’ 5’ of the coding strand of penA’ (5’BsaI adaptor) AA-2 5’-CAGCTCGAGAGACGGTGTTTTGACGGCTGC-3’ 5’ of the non-coding strand of penA’ (5’XhoI adaptor) AA-13 5’-AGACATATGTCCCAGTATCAACGAGATTGTTT-3’ 5’ of the coding strand of the region downstream from penA gene (Fig. 1A) AA-14 5’-ATCGAGCTCCCAGCCATTGCGTGATGGA-3’ 5’ of the non-coding strand of the region downstream from penA gene (Fig. 1A) 99-14 5’-GGATAATAATAACGAGAAGTAAAAATGTTG-3’ 5’ of the coding strand of penA AA-18 5’-CACTGACGCACGACGGCGTTTTAC-3’ 5’ of the coding strand of 3’ end of penA 99-23 5’-AGCGCCGTCTGAACCAAAGGGGTTAACTTGCTGAACA-3’ 5’ of the non-coding strand of penA (uptake sequence) 99-1 5’-ATGTTGATTAAGAGCGAATATAAGCC-3’ 5’ of the coding strand of penA (Fig. 1A) 99-2 5’-GATTAAGACGGTGTTTTGACGG-3’ 5’ of the non-coding strand of penA (Fig. 1A) AA-6 5’-ATGATTAAAAAGATTTTAACGACTTGTT-3’ 5’ of the coding strand of ponA AA-7 5’-TTAAAACAGGGAATCCAACTGC-3’ 5’ of the non-coding strand of ponA KM-6 5’-CCCAGCGAACCATTTGAGG-3’ 5’ of the coding strand of aph-3’ (Fig. 1A) KM-7 5’-GCTTTTTAGACATCTAAATCTAGG-3’ 5’ of the non-coding strand of aph-3’ (Fig. 1A)
by guest on April 13, 2020 http://www.jbc.org/ Downloaded from
Meningococcal peptidoglycan and penicillin G susceptibility
28
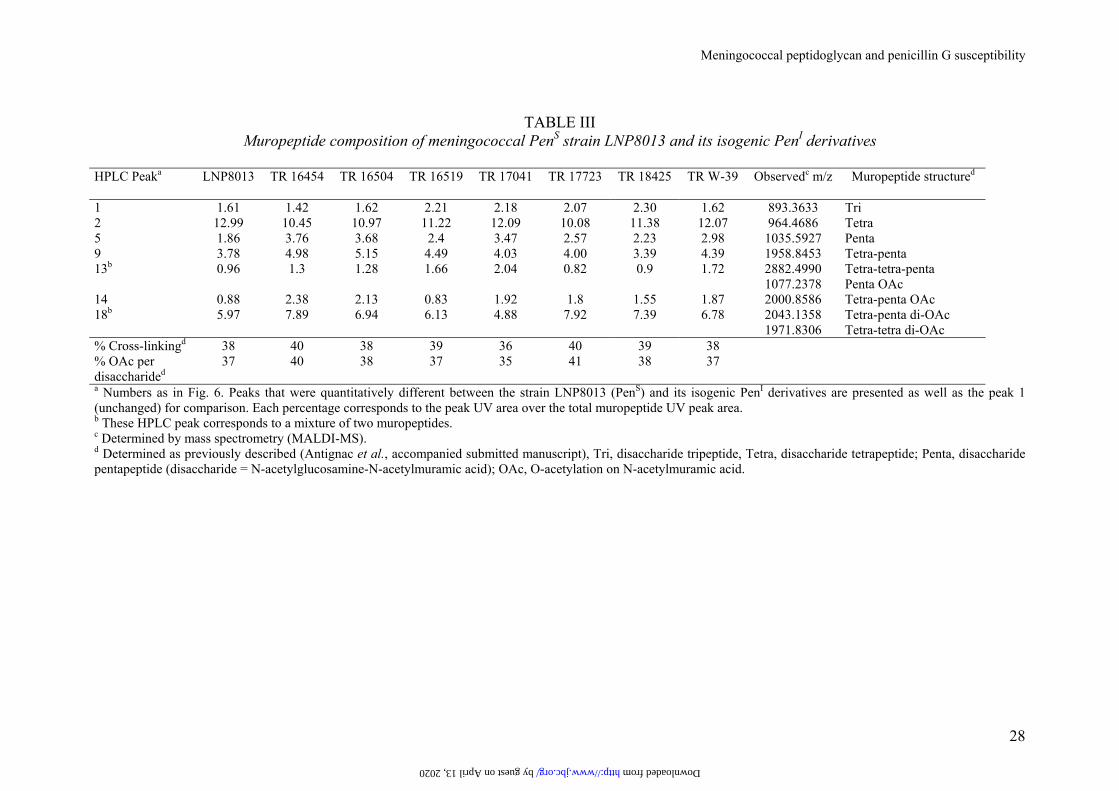
TABLE III
Muropeptide composition of meningococcal PenS strain LNP8013 and its isogenic PenI derivatives
HPLC Peaka
LNP8013 TR 16454 TR 16504 TR 16519 TR 17041 TR 17723 TR 18425 TR W-39 Observedc m/z Muropeptide structured
1 1.61 1.42 1.62 2.21 2.18 2.07 2.30 1.62 893.3633 Tri 2 12.99 10.45 10.97 11.22 12.09 10.08 11.38 12.07 964.4686 Tetra 5 1.86 3.76 3.68 2.4 3.47 2.57 2.23 2.98 1035.5927 Penta 9 3.78 4.98 5.15 4.49 4.03 4.00 3.39 4.39 1958.8453 Tetra-penta 13b 0.96 1.3 1.28 1.66 2.04 0.82 0.9 1.72 2882.4990 Tetra-tetra-penta 1077.2378 Penta OAc 14 0.88 2.38 2.13 0.83 1.92 1.8 1.55 1.87 2000.8586 Tetra-penta OAc 18b 5.97 7.89 6.94 6.13 4.88 7.92 7.39 6.78 2043.1358 Tetra-penta di-OAc 1971.8306 Tetra-tetra di-OAc % Cross-linkingd 38 40 38 39 36 40 39 38 % OAc per disaccharided
37 40 38 37 35 41 38 37
a Numbers as in Fig. 6. Peaks that were quantitatively different between the strain LNP8013 (PenS) and its isogenic PenI derivatives are presented as well as the peak 1 (unchanged) for comparison. Each percentage corresponds to the peak UV area over the total muropeptide UV peak area. b These HPLC peak corresponds to a mixture of two muropeptides. c Determined by mass spectrometry (MALDI-MS). d Determined as previously described (Antignac et al., accompanied submitted manuscript), Tri, disaccharide tripeptide, Tetra, disaccharide tetrapeptide; Penta, disaccharide pentapeptide (disaccharide = N-acetylglucosamine-N-acetylmuramic acid); OAc, O-acetylation on N-acetylmuramic acid.
by guest on April 13, 2020 http://www.jbc.org/ Downloaded from
lacZpenA1 aph-3’
99-1 KM-6 AA-13
99-2 KM-7 AA-14vector
(pGEM-T Easy)Sp
eI
Hin
dIII
Nde
I
SacI
0
50
100
150
200
250
2h 6h
LNP8013
LNP16969
LNP16635
A
B
time
beta
-gal
acto
sida
se a
ctiv
ity(M
iller
uni
ts)
(penA1)
(penA21)
(penA11)
Figure 1. Antignac et al. Meningococcal peptidoglycan and penicillin G susceptibility
penA21chromosome(strain LNP16969)
lacZpenA21 aph-3’chromosome(transformant) by guest on A
pril 13, 2020http://w
ww
.jbc.org/D
ownloaded from
TR 164540.5 µg/ml
LNP164541 µg/ml
LNP80130.094 µg/ml
5 3 1.5 1 0.5 0.1 0.05
0.001
0.0001
1
2
3
PBP
[3H]benzylpenicillin (µg/ml) Strain(penicillin G MIC )
Figure 2. Antignac et al. Meningococcal peptidoglycan and penicillin G susceptibility
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from
LNP80130.094 µg/ml
LNP165190.38 µg/ml
LNP164541 µg/ml
LNP177230.75 µg/ml
LNP184251 µg/ml
LNP170410.5 µg/ml
S-PBP2
R-PBP2s
Clinical strain(penicillin G MIC)
Penicillin G (µg/ml) IC50 (µg/ml)
0.19
0.38
0.64
0.44
0.52
0.6
4 3 2 1 0.8 0.6 0.4 0.2 0. 1
PurifiedPBP2s
TR 165190.5 µg/ml
TR 164540.5 µg/ml
TR177230.75 µg/ml
TR 184250.75 µg/ml
TR 170410.75 µg/ml
Transformant(penicillin G MIC)
Figure 3. Antignac et al. Meningococcal peptidoglycan and penicillin G susceptibility
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from
300 200 100 0
Heterogeneity
LNP8013
LNP8013
LNP8013
LNP8013
LNP8013
TR 16454
TR 16454
TR 16454
TR 16454
TR 16454
TR TH-41
TR TH-41
TR TH-41
TR TH-41
TR TH-41
Wave numbers:3338-3280 cm-1
3098-3056 cm-1
2935-2914 cm-1
1749-1715 cm-1
1669-1608 cm-1
1583-1528 cm-1
1477-1433 cm-1
1413-1381 cm-1
1179-1157 cm-1
1096-1067 cm-1
Figure 5. Antignac et al. Meningococcal peptidoglycan and penicillin G susceptibility
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from
60 120
Retention time (min)
0
500
1000
1500
2000
8
19
1
6
16
12
20
21
22 2324
2
-
5
+ 9
+
18
+
mA
U (2
05 n
m)
Figure 6. Antignac et al. Meningococcal peptidoglycan and penicillin G susceptibility
16
14
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Muhamed-Kheir TahaJean-Philippe Carlier, Julio A Vazquez, Andrew Fox, Jean-Michel Alonso and Aude Antignac, Ivo G Boneca, Jean-Claude Rousselle, Abdelkader Namane,
susceptibility to penicillin Greducedmodifications of the peptidoglycan structure in Neisseria meningitidis with
Correlation between alterations of the penicillin-binding protein 2 and
published online June 10, 2003J. Biol. Chem.
10.1074/jbc.M304607200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on April 13, 2020
http://ww
w.jbc.org/
Dow
nloaded from






























