Embed Size (px)
Citation preview

Proc. Nati. Acad. Sci. USAVol. 80, pp. 6136-6140, October 1983Physiological Sciences
Amiloride reduces the taste intensity of Na+ and Li+ saltsand sweeteners
(psychophysiology/neurophysiology/transport)
SUSAN S. SCHIFFMAN*, ELAINE LOCKHEAD*, AND FRANS W. MAESt*Department of Psychiatry, Duke University, Durham, NC 27710; and tDepartment of Zoology, Groningen State University, Haren, The Netherlands
Communicated by Knut Schmidt-Nielsen, June 30, 1983
ABSTRACT The diuretic amiloride, a potent inhibitor of so-dium transport in a variety of epithelial systems, was applied tothe human tongue. Application of amiloride reduced the taste in-tensity of sodium and lithium salts and of sweeteners ranging widelyin chemical structure. The sweeteners included saccharides, gly-cosides, dipeptides, proteins, and amino acids. Amiloride did notaffect perception of potassium or calcium salts, bitter and sourtastes, or amino acids without a sweet or salty component. Thesefindings were supported by neurophysiological studies in rat, whichshowed that amiloride diminished the NaCI response relative toKCL. The results are consistent with the position that an amiloride-sensitive transport mechanism is involved in taste perception ofsodium and lithium salts and of sweeteners.
The prevailing theory of transduction in gustation has been thatthe lingual epithelium, including taste buds, is relatively im-permeable to electrolytes (1, 2). Transduction has been pre-sumed to occur by physical adsorption of the stimulus to re-ceptors in the membrane but not to involve penetration of themembrane itself. This theory originated primarily from the workof Beidler (1), who reported that the integrated electrical ac-tivity in rat chorda tympani nerve could be related to salt con-centration by a simple Langmuir adsorption isotherm.
However, numerous lines of evidence indicate that varioussalts do penetrate lingual membranes. The most direct supportfor ion transport across lingual epithelium is the recent findingby DeSimone et al. (3) that epithelium isolated from dog tongueand mounted between symmetrical Krebs-Henseleit buffer so-lutions actively transports sodium ions. Hyperosmotic NaCl so-lutions produced increased potentials and transepithelial cur-rents. The diuretic amiloride (4), a potent inhibitor of sodiumtransport in a wide variety of cellular and epithelial transportsystems (5, 6), substantially reduced the hyperosmotic re-sponse.
Other studies support the finding by DeSimone et al. thatthe lingual epithelium is penetrated by ions. Hayashi (7) foundthat potassium, cesium, and rubidium ions can rapidly pene-trate frog tongue epithelium. Hailback et al. (8) in a study ofvascular architecture of filiform and fungiform papillae ob-tained evidence for active NaCl transport in cat tongue. In ad-dition, reduced pyridine nucleotide fluorescence from smalltaste-bud containing patches of catfish barbel epithelium sug-gests that salts and CN- can cross the epithelial barrier (:).
Although these studies clearly indicate that ions penetratelingual epithelium, they do not demonstrate directly whetherthis process is related to the sense of taste. In the present study,amiloride (N-amidino-3,5-diamino-6-chloropyrazine carbox-amide), which reduces sodium entry across the mucosal or outersurface of the epithelium (5, 6), was found to reduce the per-
ceived taste intensity of Na' and Li' and of sweeteners whenapplied to the human tongue. In addition, amiloride was foundto inhibit the neural NaCl response recorded from single unitsin the nucleus tractus solitarius in the rat. These findings sug-gest that sodium transport is directly involved in the perceptionof taste in both humans and rats.
METHODHuman Psychophysical Studies. The diuretic, amiloride hy-
drochloride, was obtained from Merck Sharp & Dohme and di-luted in deionized water. Three modes of application were usedto deliver amiloride to the tongue in five separate experiments;500 ,uM was used in experiments 1-3 and 50 ,.M in experi-ments 4 and 5.
Experiment 1. (i) Stimuli. The taste stimuli and concentra-tions that served as standards are given at the top of the solidbars in Fig. 2 A-E. Most of the standard concentrations havebeen determined to impart a moderately intense taste in pre-vious studies (9, 10). Dilutions both higher and lower than thestandard (e.g., 0.20 M NaCl) that differed from one another bya factor of 2 were used as test stimuli (e.g., 0.05 M, 0.10 M,0.20 M, and 0.40 M NaCl). Concentrations intermediate be-tween two successive dilutions were included as well (e.g., 0.30M NaCl).The sweeteners tested varied widely in chemical structure.
They included: acetosulfam (methyl derivative of oxathiazinonedioxide: 3,4-dihydro-6-methyl-1,2,3-oxathiazin-4-one-2,2-diox-ide potassium salt), aspartame (dipeptide: L-aspartyl-L-phenyl-alanine methyl ester), calcium cyclamate (calcium cyclohexylsulfamate), fructose (monosaccharide), glucose (monosaccha-ride), neohesperidin dihydrochalcone (dihydrochalcone glyco-side), rebaudioside A (diterpene glycoside), stevioside (diter-pene glycoside), sucrose (disaccharide), and thaumatin (severaldistinct proteins at M, 18,000-21,000).
(ii) Subjects. The subjects ranged in age from 18 to 35 yr andwere selected after a screening process that determined theirability to make fine discriminations in concentrations of NaCl,ranging from 0.05 M to 0.60 M; persons unable to discriminate0.10 M from 0.20 M NaCl did not participate in the experi-ments.
(iii) Mode of application of amiloride. Pieces of chromatog-raphy paper (Whatman no. 1, 0.16-mm thickness) cut in theshape of half tongues were soaked in the amiloride solution ordeionized water (control) for 10 min. Then, two pieces of paper,one impregnated with amiloride and one water control, wereplaced on the tongue (see Fig. 1A) for 2.5 min and removed,whereupon a fresh set was placed on the tongue for another 2.5min and removed for a total application time of 5 min.
t Brand, J. G., Kron, S. J. & Senseman, D. M., Fifth Annual Meetingof the Association for Chemoreception Sciences, Apr. 27-May 1, 1983,Sarasota, FL.
6136
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertise-ment" in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Dow
nloa
ded
by g
uest
on
Apr
il 23
, 202
0 D
ownl
oade
d by
gue
st o
n A
pril
23, 2
020
Dow
nloa
ded
by g
uest
on
Apr
il 23
, 202
0

Proc. Natl. Acad. Sci. USA 80 (1983) 6137
ACHROMATOGRAPHY CHROMATOGRAPHYPAPER SOAKED PAPER SOAKEDIN DEIONIZED IN AMILORIDEWAT ER
B
TEST STIMULI TEST STANDARD
(Concentrations adjusted (Applied to Amilorideto match perceived I0 side of tongue)intensity of standard) A
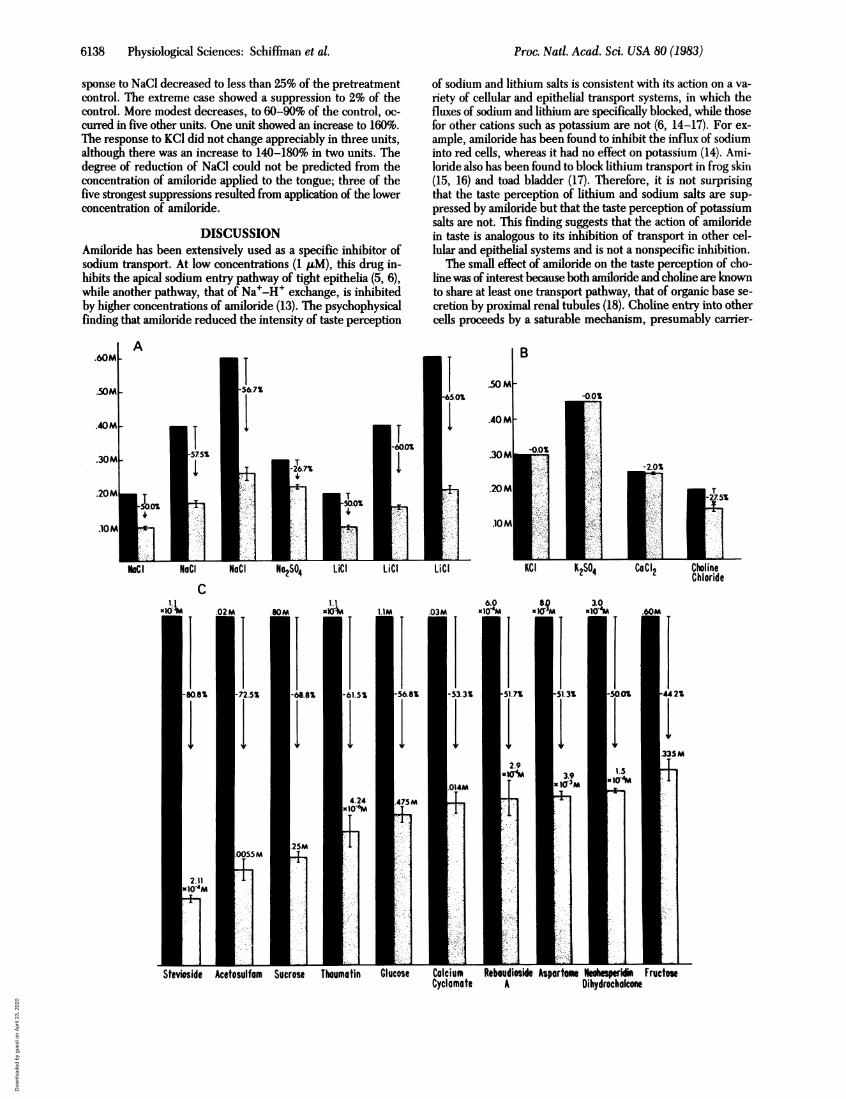
FIG. 1. (A) Amiloride was delivered to one half of the tongue in ex-periments 1, 2, and 4. A filter paper soaked in deionized water was ap-plied to the other side and served as a control. (B) A standard concen-tration of a taste stimulus impregnated in 1.27-cm circles of chro-matography paper was placed on the side of the tongue to which ami-loride had been applied. Test stimuli were applied to the nonamiloride-treated side, and the concentrations were adjusted to match the per-ceived intensity of the standard.
(iv) Testing procedure. The test stimuli were delivered to thetongue impregnated in 1.27-cm circles of chromatography pa-per as shown in Fig. 1B. The concentration to be matched was
placed on the side of the tongue adapted to amiloride. The con-centration on the other side was adjusted to match the per-ceived intensity of the standard placed on the side of the tongueadapted to amiloride. The adjustment process was achieved inthe following manner. At each trial, a pair of 1.27-cm circles(one standard and one test stimulus) was applied simulta-neously and symmetrically to the two sides of the tongue withforceps. If the sensation of the test stimulus was perceived tobe greater than the intensity of the standard applied simulta-neously to the "amiloride side" of the tongue, then a lower di-lution was used on the next trial. Alternatively, a higher con-centration was applied after the perceived intensity on the controlside was judged to be lower. A total of 10 trials (pairs of circles)were used, with the subject's response determining the orderof concentrations applied to the nonamiloride side. The con-centration at which a subjectjudged the test stimulus to be higher50% of the time and lower 50% of the time was considered tomatch the intensity on the side of the tongue adapted to ami-loride. A brief deionized water rinse (15 ml) was used after eachpair.
Experiment 2. This approach was identical to that of exper-iment 1, except that the presence of amiloride (500 MiM) wasreinforced by reapplication for 30 sec between each of the 10pairs during the matching process. NaCl (0.40 M) and stevio-side (0.0011 M) were used as stimuli.
Experiment 3. In this experiment the mode of application ofamiloride differed from that in experiments 1 and 2 in that ami-loride (500 MiM) was dissolved in the same solution as the stan-dard taste stimulus (i.e., 0.40 M NaCl or 0.0011 M stevioside).There was no preadaptation with half tongues. The standardstimuli dissolved in amiloride were impregnated in 1.27-cm cir-cles of chromatography paper and placed on one side of thetongue. Test stimuli dissolved in deionized water and also im-pregnated in 1.27-cm circles were applied simultaneously tothe other half of the tongue. The adjustment process was iden-tical to that described for the first approach.
Experiment 4. This was identical to experiment 2 except thatamiloride was used at 50 puM.
Experiment 5. This was the same as experiment 3 except thatamiloride was used at 50 AMM.One crucial aspect in all five experiments was the proper ap-
plication of the paper circles. When the circles were removedfrom the soaking solution with forceps, they were shaken to re-move excess liquid to prevent any excess from spreading acrossthe tongue. In addition, the circles were symmetrically placedin order to insure stimulation of similar numbers of receptorsand thus reliable responses.
Neurophysiological Studies. Single-unit (extracellular) re-cordings [see Woolston and Erickson (11) for method] were madefrom 11 neurons in the nucleus of the solitary tract in sevenrats. Prior to treatment with amiloride, the mouth was stim-ulated with a concentration series of NaCl to determine a mod-erate (half maximal) response for each neuron. For five neu-rons, 0.25 M KC1 was delivered as a control. Stimuli were appliedto the whole mouth for 5 sec with an interstimulus distilled waterrinse every 30 sec. Initial and maintenance doses of amiloridein distilled water (100 MM for five units, 500 A.M for six units)were applied by perfusing the oral cavity with 5 ml of 100 pMamiloride for 5 min. The stimuli were reapplied with a main-tenance dose of 2.5 ml of amiloride over 1 min given after eachpresentation. The total number of spikes over a 5-sec interval(minus the average background activity prior to each stimula-tion) were determined both prior to amiloride and after ami-loride treatment.
RESULTSHuman Psychophysical Studies. The results from experi-
ment 1 in which the tongue was adapted to 500 p.M amilorideimpregnated in chromatography paper are given in Fig. 2. InFig. 2A, it can be seen that amiloride reduced the taste inten-sity of both sodium and lithium salts. For example, the per-ceived intensity of 0.20 M, 0.40 M, and 0.60 M NaCl decreasedby 50.0%, 57.5%, and 56.7%, respectively. That amiloride hadno effect on taste perception of potassium or calcium salts isshown in Fig. 2B. However, amiloride appeared to have a mod-est effect on perception of choline chloride. Fig. 2C indicatesthat the perceived intensity of all the sweet tastes tested, in-cluding those for compounds ranging widely in chemical struc-ture, such as sweet-tasting saccharides, glycosides, dipeptides,and proteins, was inhibited by amiloride. The sweeteners areplotted in the order of percentage reduction in perceived tasteintensity. In Fig. 2D, it can be seen that amino acid compoundswith a sweet or salty component were also inhibited by ami-loride.Some subjects commented that one of the amino acids, L-
histidine HCl, had a relatively weak taste at 0.027 M. It wasused at this concentration to match the concentration of its D-enantiomer that had a moderate taste at 0.027 M. Amilorideinhibited the perception of 0.20 M L-histidine-HCl by 52.5%.Histidine-HCl has been found to have a salty taste componentin a previous study (12). Amiloride had no effect on the tasteof L-tryptophan, a bitter tasting amino acid. It also had no ef-fect on other bitter tastes or sour tastes (Fig. 2E). When ami-loride had an effect, the inhibition was perceived immediately,at the beginning of the adjustment process.The results of the three modes of application of amiloride at
both 500 and 50 ,uM are shown in Fig. 3A for NaCl and Fig.3B for the sweetener stevioside. All three methods gave similarresults, although there was less inhibition in three of the fourcases when amiloride and the stimulus were dissolved in thesame solution. The higher concentration (500 ,uM) had a greaterinhibitory effect than did 50 ,uM.
Neurophysiological Studies. Amiloride reduced the singleunit activity in 10 of the 11 units tested. In five units the re-
Physiological Sciences: Schiffman et A
Dow
nloa
ded
by g
uest
on
Apr
il 23
, 202
0
6138 Physiological Sciences: Schiffman et al.
sponse to NaCl decreased to less than 25% of the pretreatmentcontrol. The extreme case showed a suppression to 2% of thecontrol. More modest decreases, to 60-90% of the control, oc-curred in five other units. One unit showed an increase to 160%.The response to KCl did not change appreciably in three units,although there was an increase to 140-180% in two units. Thedegree of reduction of NaCl could not be predicted from theconcentration of amiloride applied to the tongue; three of thefive strongest suppressions resulted from application of the lowerconcentration of amiloride.
DISCUSSIONAmiloride has been extensively used as a specific inhibitor ofsodium transport. At low concentrations (1 j.M), this drug in-hibits the apical sodium entry pathway of tight epithelia (5, 6),while another pathway, that of Na+-H+ exchange, is inhibitedby higher concentrations of amiloride (13). The psychophysicalfinding that amiloride reduced the intensity of taste perception
Proc. Natl. Acad. Sci. USA 80 (1983)
of sodium and lithium salts is consistent with its action on a va-riety of cellular and epithelial transport systems, in which thefluxes of sodium and lithium are specifically blocked, while thosefor other cations such as potassium are not (6, 14-17). For ex-ample, amiloride has been found to inhibit the influx of sodiuminto red cells, whereas it had no effect on potassium (14). Ami-loride also has been found to block lithium transport in frog skin(15, 16) and toad bladder (17). Therefore, it is not surprisingthat the taste perception of lithium and sodium salts are sup-pressed by amiloride but that the taste perception of potassiumsalts are not. This finding suggests that the action of amiloridein taste is analogous to its inhibition of transport in other cel-lular and epithelial systems and is not a nonspecific inhibition.The small effect of amiloride on the taste perception of cho-
line was of interest because both amiloride and choline are knownto share at least one transport pathway, that of organic base se-cretion by proximal renal tubules (18). Choline entry into othercells proceeds by a saturable mechanism, presumably carrier-
CholineChloride
C1.1Im 1.1 6.0 BP 3.0
X1FM .02M MxWm A LIM .03M Xle0M x19-M x110M .60MT _T _r T T _T _T _ T _ T _
2.1
2.11iX10-4AT
.;
OOSSM
T::
.: :..0-.
.:-...
'''.-.::'1
.2SMI-.-..
Stevioside Acetosulfam Sucrose
I61.51
1
4.24X10%-M
Thoumatin
6.81
47SM
T.........
-::..'''.::,
.. 7.
::
0-,.-
.-..:
...._ 3.31
.014MA
1~V..
.-.
51.71
I2.9
1.3S
3.9x 1FM
D.0S
1.5
x 04M
Glucose Calcium Rebaudioside Aspartame ebeeid FructoseCyclamate A Dihydrochalcone
-4.21
.335M
.1
I
31
4
Dow
nloa
ded
by g
uest
on
Apr
il 23
, 202
0
Proc. Natl. Acad. Sci. USA 80 (1983) 6139
mediated, but it is not yet known whether this can be inhibitedby amiloride.
It was surprising that amiloride blocked perception of all thesweet-tasting compounds. That this was not simply a nonspe-cific blunting of taste sensation is suggested by the fact thatbitter and sour tastes used in this experiment were not af-fected. The reduction in sweet tastes may indicate that sweet-ness is in part mediated by an amiloride-sensitive sodium chan-nel. Alternatively, amiloride may also occupy the same receptorsite utilized by sweet-tasting compounds. One theory (19) pos-tulates that a necessary condition for sweetness is a pair of hy-drogen bonds 3 A apart. An AH-B configuration in the stim-ulus molecule is presumed to interact with a complementaryAH-B site in the taste-receptor membrane, where A and B areelectronegative atoms and H is a hydrogen atom that is part ofa polarized system A-H. Amiloride, like sweet-tasting com-pounds, also has an AH-B unit capable of intermolecular hy-drogen bonding. Either one or both mechanisms may be in-volved in the inhibition of sweet taste.The results of neurophysiological studies in rats from indi-
vidual medullary taste neurons indicate that amiloride sup-presses the response to NaCl but leaves the response to KC1relatively unaffected. Further experiments with a larger num-ber of neurons and a variety of concentrations will permit quan-titative comparison of neural and psychophysical responses.However, the qualitative agreement between the psychophys-ical and neural data reported here is striking.The psychophysical results are also consistent with the al-
teration in ion-transport properties reported by DeSimone etal. (3, 20, §) in an in vitro preparation of dog tongue epithelium
§ DeSimone, J. A., Schiffman, S. S., Maes, F. W, Heck, G. L.,DeSimone, S. K. & Mierson, S., Fifth Annual Meeting of the Asso-ciation for Chemoreception Sciences, Apr. 27-May 1, 1983, Sarasota,FL.
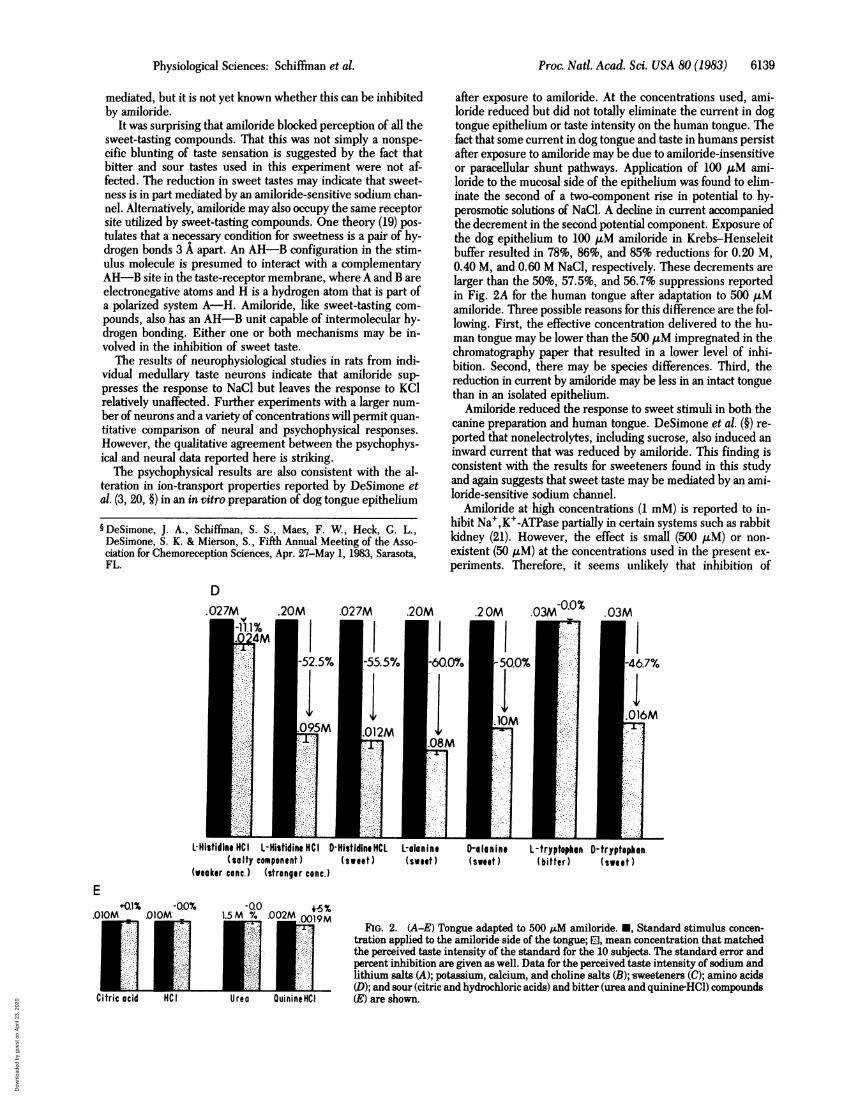
after exposure to amiloride. At the concentrations used, ami-loride reduced but did not totally eliminate the current in dogtongue epithelium or taste intensity on the human tongue. Thefact that some current in dog tongue and taste in humans persistafter exposure to amiloride may be due to amiloride-insensitiveor paracellular shunt pathways. Application of 100 ,M ami-loride to the mucosal side of the epithelium was found to elim-inate the second of a two-component rise in potential to hy-perosmotic solutions of NaCl. A decline in current accompaniedthe decrement in the second potential component. Exposure ofthe dog epithelium to 100 ,M amiloride in Krebs-Henseleitbuffer resulted in 78%, 86%, and 85% reductions for 0.20 M,0.40 M, and 0.60 M NaCl, respectively. These decrements arelarger than the 50%, 57.5%, and 56.7% suppressions reportedin Fig. 2A for the human tongue after adaptation to 500 AuMamiloride. Three possible reasons for this difference are the fol-lowing. First, the effective concentration delivered to the hu-man tongue may be lower than the 500 p.M impregnated in thechromatography paper that resulted in a lower level of inhi-bition. Second, there may be species differences. Third, thereduction in current by amiloride may be less in an intact tonguethan in an isolated epithelium.
Amiloride reduced the response to sweet stimuli in both thecanine preparation and human tongue. DeSimone et al. (§) re-ported that nonelectrolytes, including sucrose, also induced aninward current that was reduced by amiloride. This finding isconsistent with the results for sweeteners found in this studyand again suggests that sweet taste may be mediated by an ami-loride-sensitive sodium channel.
Amiloride at high concentrations (1 mM) is reported to in-hibit Na+,K+-ATPase partially in certain systems such as rabbitkidney (21). However, the effect is small (500 ,uM) or non-existent (50 ,uM) at the concentrations used in the present ex-periments. Therefore, it seems unlikely that inhibition of
D.027M .20M .027M .20M .2 OM .03M00% 03M
.0MI
6.7%
L-Histidine NCI L-Histidine HCI D-HistidineHCL L-olonine D-olonine L-tryptophon D-tryptophon(salty component) (sweet) (sweet) (sweet) (bitter) (sweet)
(weaker conc.) (stronger conc.)
Eo.1% -0.0% -o.o 4- %
.OlOM OlOM 1.5 M % 002 0019M
Citric acid HCI Urea OuinineHCI
FIG. 2. (A-E) Tongue adapted to 500 1,M amiloride. *, Standard stimulus concen-tration applied to the amiloride side of the tongue; El, mean concentration that matchedthe perceived taste intensity of the standard for the 10 subjects. The standard error andpercent inhibition are given as well. Data for the perceived taste intensity of sodium andlithium salts (A); potassium, calcium, and choline salts (B); sweeteners (C); amino acids(D); and sour (citric-and hydrochloric acids) and bitter (urea and quinine HCl) compounds(E) are shown.
Physiological Sciences: Schiffinan et al.
Dow
nloa
ded
by g
uest
on
Apr
il 23
, 202
0

6140 Physiological Sciences: Schiffiman et al..
50y M AMILORIDE
B
5OOM AMILORIDE 5OyuM AMILORiDE-
FIG. 3. (A) Amiloride inhibited the perceived taste intensity of NaClwhen applied at both 500,M and 50 ,uM. The results from experiments1-5 are given in columns a-e, respectively. (B) Amiloride inhibited theperceived taste intensity of the sweetener stevioside when applied at500 ,uM and 50 ,uM. The results from experiments 1-5 are given in col-umns a-e, respectively.
Na+,K+-ATPase plays a significant role in the present study.It should be pointed out that the shape of. the classic re-
sponse-concentration curve- of Beidler (1) is entirely compat-ible with a saturable transport pathway for sodium of the kindknown to be inhibited by amiloride as suggested by the present.results. In addition, the adsorptive mechanism proposed byBeidler may be appropriate for other types of stimuli, such as
certain nonelectrolytes:
In conclusion, the findings reported here suggest that a.so-dium transport pathway is involved in the perception of taste.Multidisciplinary data including psychophysical judgments fromhumans, neurophysiological recordings from rat, along with thetransepithelial potential differences from. isolated lingual epi-thelium of dog and rat reported by DeSimone et al. (3, 20, §)suggest that an amiloride-sensitive transport mechanism plays.a role in the taste mechanism for sodium and lithium salts aswell as for sweeteners. These results are consistent with pre-vious studies in a variety of other cellular and epithelial systemsthat have shown that' amiloride inhibits sodium transport.The authors thank J. A. DeSimone, S. A. Simon, G. L. Heck, R. P.
Erickson, J. M. Gill; and J. Garvin for their helpful comments on thispaper. This research was supported in part by National Institute on Ag-ing Grant AG00443 and National Science Foundation Grant BNS 8021604. The experiments on sweeteners were supportedin part by a grantfrom Searle.
1. Beidler, L. M. (1954)J. Gen. Physiol. 38, 133-139.2. Kamo,. N., Miyake, M., Kurihara, K. & Kobatake, Y. (1974)
Biochim. Biophys. Acta 367; 11-21.3. DeSimone, J. A., Heck, G. L. & DeSimone, S. K. .(1981) Science
214, 1039-1041.4. Cragoe,. E. J., Jr. (1979) in Amiloride and Epithelial Sodium
Transport, eds. Cuthbert, A. W., Fanelli,. E. M., Jr., & Scria-bine, A. (Urban & Schwarzenberg, Baltimore), pp. 1-20.
5. Bentley, P. J. (1968)J. Physiol. (London) 195, 317-330,6. Benos, D. J. (1982) Am. J. Physiol. 242,_C131-_C145.7. Hayashi, H. (1978) Jpn. J. Physiol. 28, 33-45.8. Hallback, D1 A., Jodal, M. &'Lundgren, 0. (1979) Acta.Physial.
Scand. 105, 469-480.9. Schiffman, S. S. & Erickson, R. P. (1971) Physiol. Behav. 7, 617-
633.10. Schiffinan, S. S., Lindley, M. G., Clark, T. B. & Makino, C. (1981)
Neurobiol. Aging 2, 173-185.11. Woolston, D. C. & Erickson, R. P. (1979) J.'Neurophysiol. 42, 1390-
1409.12. Schiffnan, S. S. & Dackis, C. (1975) Percept Psychophys. 17, 140-
146.13. Kinsella, J L. & Aronson, P. S. (1981) Am. J. Physiol. 2411 F374-
F379.14. Aceves, J. & Cerijido, M. (1973)J. Physiol. 229, 709-718.15. Candia, 0. A. & Chiarandini, D. J. (1973) Biochim. Biophys. Acta
307, 578-589.16. Morel, F. & Leblanc, G. (1975) Pflugers Arch.'358, 135-157.17. Herrera, F. C. (1972)'Am. J. Physiol. 222, 499-502.18. Rennick, B. R. (1981) Am. J. Physiol. 240, F83-F89..19. Shallenberger, R. S; & Acree, T. E. (1971)' in Handbook of Sen-
sory Physiology, ed. Beidler, L. M. (Springer, Berlin), pp. 221-227.
20. DeSimone, J. A., Heck, G. L., Mierson, S. & DeSimone, S. K.(1983) J. Gen. Physiol., in press.
21. Soltoff, S. P. & Mandel, L. J. (1983) Science 220, 957-959.
A
500-M AMILORIDE
Proc. Natl. Acad Sci. USA 80 (1983)'
Dow
nloa
ded
by g
uest
on
Apr
il 23
, 202
0

Proc. NatL Acad Sci. USA 81 (1984)
Correction. In the article "Avian carcinoma virus MH2 con-tains a transformation-specific sequence, mht, and sharesthe myc sequence with MC29, CMII, and OK10 viruses" byNancy C. Kan, Christos S. Flordellis, Claude F. Garon, Pe-ter H. Duesberg, and Takis S. Papas, which appeared innumber 21, November 1983, of Proc. Natl. Acad. Sci. USA(80, 6566-6570), the authors wish to clarify the following.The authors referred to the recent finding of a high degree ofrelationship (identity) between transforming sequences ofthe avian retrovirus MH2 (the mht gene) and the murine ret-rovirus 3611-MSV (the raf gene) as "unpublished data." Themht sequence is unpublished data of the authors, and the rafsequence is unpublished data of G. Mark and U. Rapp (Lab-oratory of Viral Carcinogenesis, National Cancer Institute-Frederick Cancer Research Facility, Frederick, MD 21701).The sequence comparison was made by J. Lautenberger.
Correction. In the article "Amiloride reduces the taste in-tensity of Na' and Li' salts and sweeteners" by Susan S.Schiffman, Elaine Lockhead, and Frans W. Maes, which ap-peared in number 19, October 1983, of Proc. Natl. Acad.Sci. USA (80, 6136-6140), the authors request that the fol-lowing acknowledgment be added. "F.W.M. was supportedby grants from the Netherlands Organization for the Ad-vancement of Pure Research ZWO and from the NationalScience Foundation to Dr. R. P. Erickson."
1608 Corrections





























