Embed Size (px)
Citation preview

Copepod Diel Migration, Feeding, and the Vertical Flux of PheopigmentsAuthor(s): M. J. Dagg, B. W. Frost and W. E. Walser, Jr.Source: Limnology and Oceanography, Vol. 34, No. 6 (Sep., 1989), pp. 1062-1071Published by: American Society of Limnology and OceanographyStable URL: http://www.jstor.org/stable/2837195 .
Accessed: 14/06/2014 02:48
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
American Society of Limnology and Oceanography is collaborating with JSTOR to digitize, preserve andextend access to Limnology and Oceanography.
http://www.jstor.org
This content downloaded from 195.34.79.20 on Sat, 14 Jun 2014 02:48:56 AMAll use subject to JSTOR Terms and Conditions

Limnol. Oceanogr., 34(6), 1989, 1062-1071 ? 1989, by the American Society of Limnology and Oceanography, Inc.
Copepod diel migration, feeding, and the vertical flux of pheopigments
M. J. Dagg Louisiana Universities Marine Consortium, Chauvin 70344
B. W. Frost School of Oceanography, WB-10, University of Washington, Seattle 98195
W. E. Walser, Jr. Louisiana Universities Marine Consortium
Abstract Feeding and diel vertical migration in the copepods Calanus pacificus Brodsky and Metridia
lucens Boeck were examined during dusk and dawn in Dabob Bay, Washington. Both species migrated from below 75 m during the day into the upper 25 m at night. Feeding on phytoplankton was confined to periods spent in the surface layer.
Metridia lucens arrived in the surface 0.5 h after sunset, fed at high rates throughout the night until 1.0 h before sunrise, and then returned to depth. In contrast, C. pacificus appeared in the surface layer 0.5-1.5 h before sunset but fed at low rates until 1 h after sunset, then increased its feeding rate. Feeding decreased before sunrise, although migratory descent was not completed until after sunrise. These patterns are not in agreement with models of diel migration that predict the onset of feeding 1-2 h before sunset.
During the night in surface waters, these copepods defecated 2.3-2.9 mg pigment m-2. In com- parison, active transport of pigment to deep waters via migratory descent was only 0.01-0.02 mg pigment m-2 d-l.
The feeding behavior of marine copepods can vary over periods as short as several hours, even when food concentrations are essentially constant (Mackas and Burns 1986; Dagg and Grill 1980; Mackas and Bohrer 1976). This observation suggests that the nighttime feeding on phytoplankton by dielly migrating copepods is not a simple function of their presence in a surface, food- rich layer. In addition, it has been proposed that for energetic reasons, dielly migrating copepods ought to begin feeding on phy- toplankton 1-2 h before sunset (Enright 1977). One of the goals of this study was to examine the timing of feeding onset and cessation in copepods that were dielly mi- grating into and out of a phytoplankton-rich surface layer.
Besides being of interest to zooplanktol- ogists, the relationship between feeding and
Acknowledgments This work was supported by NSF grants OCE 85-
14530 (Dagg) and OCE 84-08929 (Frost) and by the Louisiana Universities Marine Consortium (Dagg).
The technical assistance of D. Thoreson and G. Her- on is greatly appreciated.
diel migration has potential oceanographic consequences. Actively feeding copepods produce several fecal pellets each hour, and the feeding pattern of a population influ- ences the flux of material from the euphotic zone. For example, a diel signal in the flux of pheopigments was associated with the diel migration and feeding of large copepods in Dabob Bay, Washington (Lorenzen and Welschmeyer 1983). Also, vertically mi- grating organisms may actively transport significant amounts of material to deeper waters if "the retention time in the guts is long enough for an animal with a full stom- ach leaving shallow depths to reach its max- imum depth before it defecates, yet short enough ... not to transport it back up the water column on its next migration cycle" (Angel 1984, p. 492). A second goal of our study was to determine the significance of this active transport process.
Methods and materials The study was done at a central, 195-m-
deep station in Dabob Bay (47?45.5'N, 122?49.5'W) during 6-8 May and 4-6 Au- gust 1986. Dabob Bay is a moderately pro-
1062
This content downloaded from 195.34.79.20 on Sat, 14 Jun 2014 02:48:56 AMAll use subject to JSTOR Terms and Conditions

Copepodfeeding and migration 1063
ductive fjord, with about 200 g C m-2 yr-1 primary production (Welschmeyer et al. 1984). It is particularly suitable for study of diel migration of zooplankton; due to the bathymetry of the bay and negligible river inflow, tidally induced horizontal motions are small (Kollmeyer 1965; Ebbesmeyer 1973; McGary and Lincoln 1977; Jamart and Winter 1978) and estuarine circulation is negligible (Barnes and Ebbesmeyer 1978). It is reasonable to presume that the same planktonic population is sampled over pe- riods of a few days.
Vertical distribution of copepods was de- termined by vertical hauls with a multinet sampler configured like the Be net (Be 1962), carrying five nets of 1 x 1 -m mouth opening and 333-,um mesh. The water column was sampled over six depth strata: 175-125, 125-75, 75-50, 50-25, 25-10, and 10-0 m. Duplicate vertical hauls were made on two successive days and nights during each cruise. Samples were preserved in a sea- water-Formalin solution. The mean abun- dances of adult female Calanus pacificus and Metridia lucens in replicate subsamples tak- en with a Stempel pipette were used to de- termine the copepod concentrations in each depth stratum. The systematics of the coast- al Pacific population often attributed to Metridia pacifica Brodsky is inconclusive (Thorpe 1980); we follow the taxonomic opinions of Fleminger (1967) and Park (1968) in assigning the name M. lucens Boeck.
Incident solar radiation (400-700-nm wavelength, Einst m-2) was measured con- tinuously throughout both observation pe- riods with a LiCor 195 quantum meter mounted on a post above the ship's super- structure, and average and integrated fluxes over 10-min intervals were calculated and recorded with a LiCor T-550 integrator. From these data we defined sunrise as the start of the first 10-min interval and sunset as the end of the last 10-min interval with positive flux (range of flux during such 10- min intervals was 9-14,uEinst m-2 S-1). On the basis of more extensive data than pre- sented here, times of sunrise and sunset so defined generally agreed to within 30 min with predictions (U.S. Dep. Commerce 1985), but on the mornings of 6 and 7 May
heavy cloud cover caused a 45-55-min de- lay between predicted and observed sunrise.
Phytoplankton standing stock was mea- sured as concentration of chlorophyll a in samples collected at five to seven depths in the upper 30 m (see Frost 1988).
Copepods for gut content analysis were collected in vertical hauls with a 1 -m closing net (227-,um mesh) from two depth strata: a surface stratum of 25-0 m and a deep stratum typically of 135-80 m but of 80- 50 m for part of the August series. During May, gut pigment concentrations were mea- sured in females of C. pacificus; during Au- gust, analyses were made on females of both C. pacificus and M. lucens.
Partial evacuation of gut contents during the time between collection and sorting of copepods causes underestimates of in situ gut contents. In May, this problem was min- imized by restricting the sorting period to < 10 min after the net was brought on deck. When copepods were abundant, six repli- cates of five C. pacificus females each were easily sorted within a few minutes by two of us. When copepods were not abundant, replicate samples could not be sorted within the 10-min period. Our ability to sort rep- licates thus provided a qualitative index of copepod abundance, similar to that of Boyd et al. (1980). Copepods were sorted into vials containing several milliliters of filtered sea- water. The contents of each vial were fil- tered onto a 24-mm GF/F filter. The filter was placed in a cryotube, frozen in liquid nitrogen, and returned to the laboratory for gut pigment analysis.
In August, the potential for partial evac- uation of gut contents during sorting was further reduced. Immediately after each net haul came on board the cod-end contents were poured into a bucket of filtered sea- water. Duplicate or triplicate subsamples were taken by passing small sieves through the water. The collecting portion of each sieve was a removable 47-mm disk of 253- or 333-,gm-mesh Nitex screen. Each disk was immediately placed in a 47-mm plastic Petri dish, wrapped in foil, and frozen in liquid nitrogen. Samples were frozen within 2-3 min after the net came on deck.
In the laboratory, filters containing sorted C. pacificus females from the May cruise
This content downloaded from 195.34.79.20 on Sat, 14 Jun 2014 02:48:56 AMAll use subject to JSTOR Terms and Conditions

1064 Dagg et al.
were removed from the cryotubes and placed directly into grinding tubes with a few milli- liters of 90% aqueous acetone. Each Nitex disk from the August samples was removed from the liquid nitrogen and placed in a dish of filtered seawater. When sufficient cope- pods were present, three replicates of 10 C. pacificus females and 10 M. lucens females were immediately picked by fine forceps, dipped into a second dish of filtered sea- water for a further brief rinse, and placed in grinding tubes with a few milliliters of 90% acetone.
The samples were homogenized in 10 ml of 90% acetone and the fluorescence of the filtrate was measured in a Turner Designs model 10 fluorometer before and after acid- ification with two drops of 10% HCI. Chlo- rophyll and pheopigment (as chlorophyll equivalents) concentrations of each sample were calculated with equations for in vitro fluorometry (Strickland and Parsons 1968) modified slightly to
ng Chl copepod- = k(fo - fa) n
pheopigment (as ng Chl = k(Rfa - fo) equivalent n copepod-1)
where k is the machine calibration constant, fo andfa are the fluorescence readings before and after acidification, R is the acidification ratio, and n is the number of copepods. To- tal pigment content of each copepod was determined by directly summing these two values. All values were then corrected for the small amount of fluorescence contained in starved copepods: 0.74 ng pigment co- pepod-I in C. pacificus and 0.33 in M. lu- cens. All samples were analyzed within 30 d of collection.
During May, we made two shipboard measurements of gut evacuation rate in C. pacificus females. In one, females were fed Thalassiosira weissflogii at 4.0 ,g Chl liter-l before placement in filtered seawater. Levels of gut pigment were measured in subsam- ples at selected intervals for a period of 1 h to determine the rate of gut evacuation (see Dagg and Wyman 1983). The second measurement was made with the approach
of Dagg and Walser (1987); individuals were fed in bottles for 6 h between midnight and dawn, and the ratio of gut pigment to fecal pigment was used to calculate the gut passage time. In addition, several of these experiments were run in the laboratory with C. pacificus females before the August cruise.
Results Timing of migration andfeeding on phy-
toplankton-During May, only a part of the C. pacificus population appeared to be dielly migrating; about half the C. pacificus fe- males were found above 50 m at night and almost the entire population was found be- low 50 m during the day (Fig. 1). During August, almost all of the C. pacificus females were dielly migrating from below 75 m dur- ing the day to above 50 m, and mostly above 10 m, at night (Fig. 2). Metridia lucens fe- males in August were also strongly migrat- ing; almost the entire population was below 75 m during the day and above 50 m, mostly above 25 m, at night (Fig. 2). During both sampling periods, the standing stock of phy- toplankton, measured as Chl a, was strongly concentrated in the upper 15 m (see figure 3 of Frost 1988)
With a few exceptions, C. pacificus col- lected from deep water in May had guts devoid of phytoplankton pigments, and in- dividuals collected from the surface layer had high levels of gut pigments (Fig. 3). This pattern is expected in copepods migrating into and out of a phytoplankton-rich surface layer.
The close temporal spacing of samples during the late afternoon and evening of the second day provides a more detailed picture of the copepods' feeding behavior and per- mits inferences on the timing of migration. During the samplings at noon and 1600 hours, C. pacificus females were absent from the surface stratum (arrows in Fig. 3). In each of the four hauls made from 1730 to 1930 hours, we collected enough C. pacifi- cus for only a single analysis (triangles in Fig. 3), indicating that some of the migrating population was in the surface layer during this time. In three of these samples, indi- viduals contained small but significant amounts of gut pigment, indicating that
This content downloaded from 195.34.79.20 on Sat, 14 Jun 2014 02:48:56 AMAll use subject to JSTOR Terms and Conditions
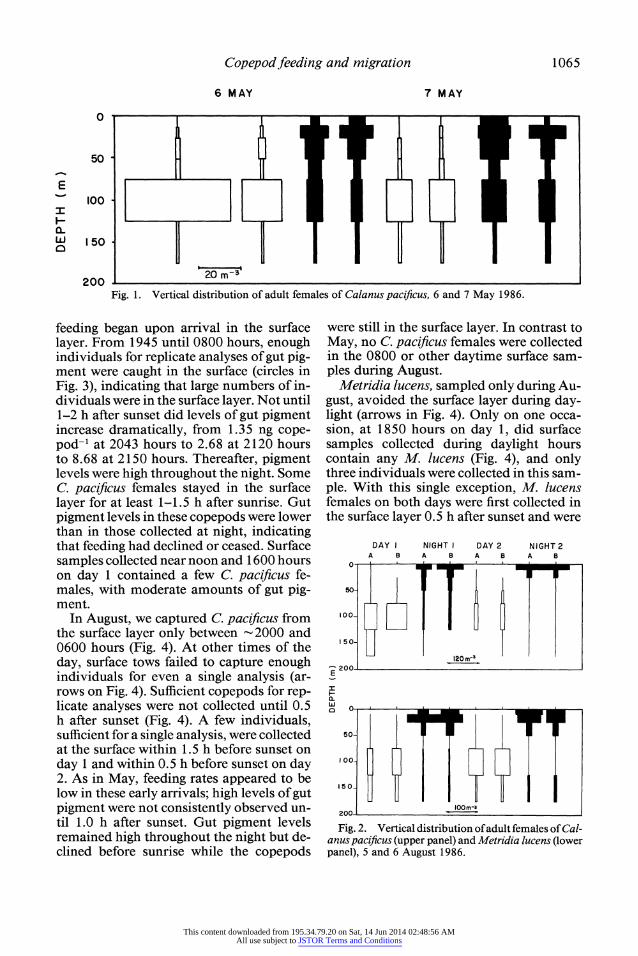
Copepod feeding and migration 1065
6 MAY 7 MAY
0
50
E 100
Fi-. L W 150
200 20m Fig. 1. Vertical distribution of adult females of Calanus pacificus, 6 and 7 May 1986.
feeding began upon arrival in the surface layer. From 1945 until 0800 hours, enough individuals for replicate analyses of gut pig- ment were caught in the surface (circles in Fig. 3), indicating that large numbers of in- dividuals were in the surface layer. Not until 1-2 h after sunset did levels of gut pigment increase dramatically, from 1.35 ng cope- pod-' at 2043 hours to 2.68 at 2120 hours to 8.68 at 2150 hours. Thereafter, pigment levels were high throughout the night. Some C. pacificus females stayed in the surface layer for at least 1-1.5 h after sunrise. Gut pigment levels in these copepods were lower than in those collected at night, indicating that feeding had declined or ceased. Surface samples collected near noon and 1600 hours on day 1 contained a few C. pacificus fe- males, with moderate amounts of gut pig- ment.
In August, we captured C. pacificus from the surface layer only between 2000 and 0600 hours (Fig. 4). At other times of the day, surface tows failed to capture enough individuals for even a single analysis (ar- rows on Fig. 4). Sufficient copepods for rep- licate analyses were not collected until 0.5 h after sunset (Fig. 4). A few individuals, sufficient for a single analysis, were collected at the surface within 1.5 h before sunset on day 1 and within 0.5 h before sunset on day 2. As in May, feeding rates appeared to be low in these early arrivals; high levels of gut pigment were not consistently observed un- til 1.0 h after sunset. Gut pigment levels remained high throughout the night but de- clined before sunrise while the copepods
were still in the surface layer. In contrast to May, no C. pacificus females were collected in the 0800 or other daytime surface sam- ples during August.
Metridia lucens, sampled only during Au- gust, avoided the surface layer during day- light (arrows in Fig. 4). Only on one occa- sion, at 1850 hours on day 1, did surface samples collected during daylight hours contain any M. lucens (Fig. 4), and only three individuals were collected in this sam- ple. With this single exception, M. lucens females on both days were first collected in the surface layer 0.5 h after sunset and were
DAY I NIGHT I DAY 2 NIGHT 2 A B A B A B A B
150 10 n 200
200-
I a-
50-
a 00-
15 0
lOOm-3 200
Fig. 2. Vertical distribution of adult females of Cal- anus pacificus (upper panel) and Metridia lucens (lower panel), 5 and 6 August 1986.
This content downloaded from 195.34.79.20 on Sat, 14 Jun 2014 02:48:56 AMAll use subject to JSTOR Terms and Conditions
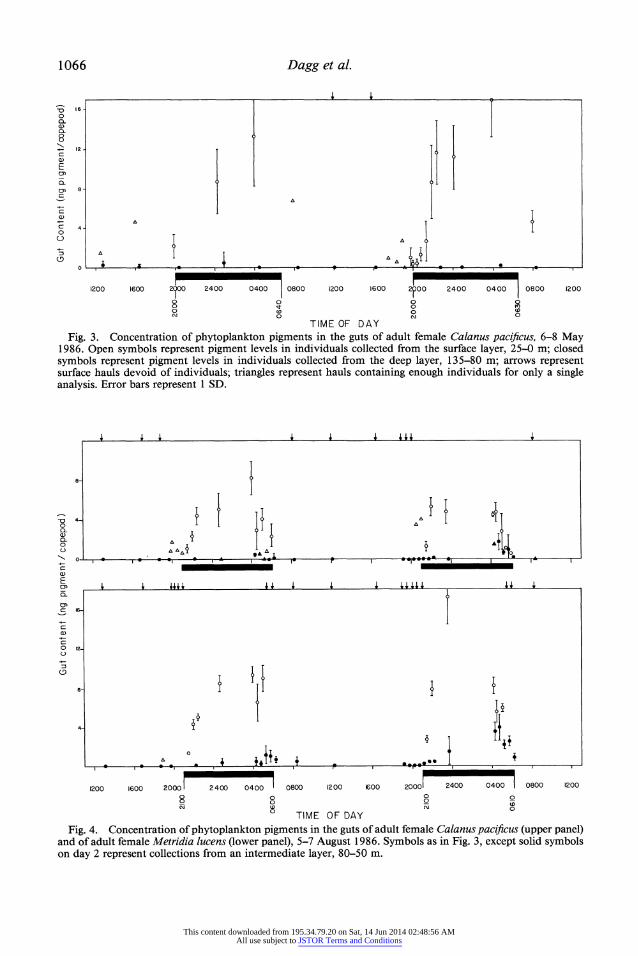
1066 Dagg et al.
0 0.
a)
E
1200 1600 2X0 400400 0800 1200 1600 2pO0240 0400 0800 1200
o 0 0 o o o
oD O ED
0~~~~~~~~~~~~~~~~~~~~~~
TIME OF DAY Fig. 3. Concentration of phytoplankton pigments in the guts of adult female Calanus pacificus, 6-8 May
1986. Open symbols represent pigment levels in individuals collected from the surface layer, 25-0 m; closed symbols represent pigment levels in individuals collected from the deep layer, 135-80 m; arrows represent surface hauls devoid of individuals; triangles represent hauls containing enough individuals for only a single analysis. Error bars represent 1 SD.
? 40 A A^ 4^ X '"
8
4-~~~~~~~~~~~~~~~~~~~ 0~ 0
E
1200 1600 20001 2400 0400 0800 1200 1600 20001 2400 0400 1 .0800 1200
0~~~~~~~~~~~~~~~~~~~~~~~
? TIME OF DAY Fig. 4. Concentration of phytoplankton pigments in the guts of adult female Calanus pacificus (upper panel)
and of adult female Metridia lucens (lower panel), 5-7 August 1986. Symbols as in Fig. 3, except solid symbols on day 2 represent collections from an intermediate layer, 80-50 m.
This content downloaded from 195.34.79.20 on Sat, 14 Jun 2014 02:48:56 AMAll use subject to JSTOR Terms and Conditions
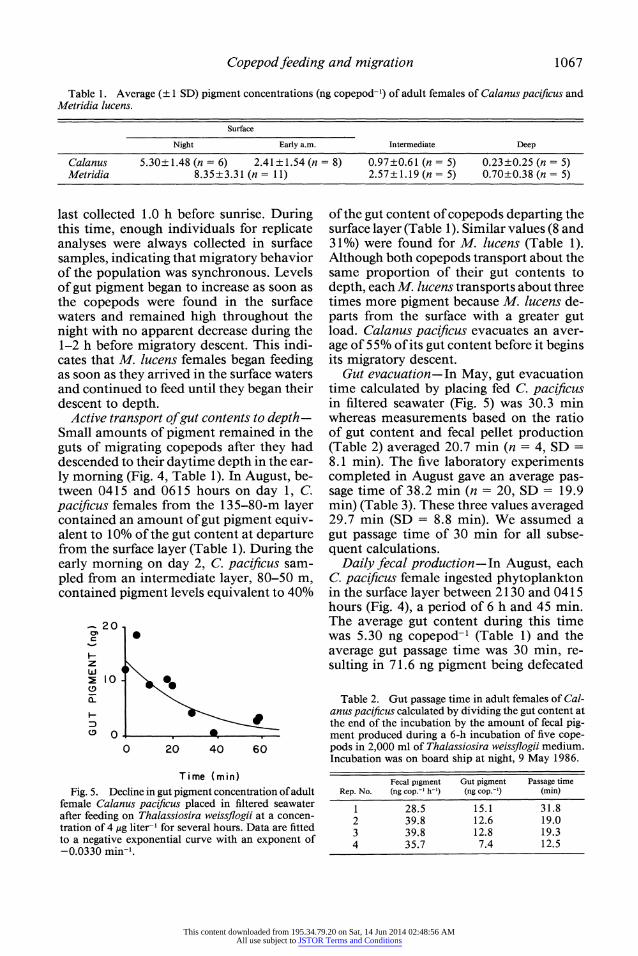
Copepod feeding and migration 1067
Table 1. Average (? 1 SD) pigment concentrations (ng copepod-') of adult females of Calanus pacificus and Metridia lucens.
Surface
Night Early a.m. Intermediate Deep
Calanus 5.30?1.48 (n = 6) 2.41?1.54 (n = 8) 0.97?0.61 (n = 5) 0.23?0.25 (n = 5) Metridia 8.35?3.31 (n = 11) 2.57?1.19(n = 5) 0.70?0.38(n = 5)
last collected 1.0 h before sunrise. During this time, enough individuals for replicate analyses were always collected in surface samples, indicating that migratory behavior of the population was synchronous. Levels of gut pigment began to increase as soon as the copepods were found in the surface waters and remained high throughout the night with no apparent decrease during the 1-2 h before migratory descent. This indi- cates that M. lucens females began feeding as soon as they arrived in the surface waters and continued to feed until they began their descent to depth.
Active transport of gut contents to depth- Small amounts of pigment remained in the guts of migrating copepods after they had descended to their daytime depth in the ear- ly morning (Fig. 4, Table 1). In August, be- tween 0415 and 0615 hours on day 1, C. pacificus females from the 135-80-m layer contained an amount of gut pigment equiv- alent to 10% of the gut content at departure from the surface layer (Table 1). During the early morning on day 2, C. pacificus sam- pled from an intermediate layer, 80-50 m, contained pigment levels equivalent to 40%
20 o 0
z
M 10 *
0 \ 0 20 40 60
Time (m i n) Fig. 5. Decline in gut pigment concentration of adult
female Calanus pacificus placed in filtered seawater after feeding on Thalassiosira weissflogii at a concen- tration of 4 ,ug liter-' for several hours. Data are fitted to a negative exponential curve with an exponent of -0.0330 min-'.
of the gut content of copepods departing the surface layer (Table 1). Similar values (8 and 31%) were found for M. lucens (Table 1). Although both copepods transport about the same proportion of their gut contents to depth, each M. lucens transports about three times more pigment because M. lucens de- parts from the surface with a greater gut load. Calanus pacificus evacuates an aver- age of 5 5% of its gut content before it begins its migratory descent.
Gut evacuation-In May, gut evacuation time calculated by placing fed C. pacificus in filtered seawater (Fig. 5) was 30.3 min whereas measurements based on the ratio of gut content and fecal pellet production (Table 2) averaged 20.7 min (n = 4, SD = 8.1 min). The five laboratory experiments completed in August gave an average pas- sage time of 38.2 min (n = 20, SD = 19.9 min) (Table 3). These three values averaged 29.7 min (SD = 8.8 min). We assumed a gut passage time of 30 min for all subse- quent calculations.
Daily fecal production-In August, each C. pacificus female ingested phytoplankton in the surface layer between 2130 and 0415 hours (Fig. 4), a period of 6 h and 45 min. The average gut content during this time was 5.30 ng copepod-I (Table 1) and the average gut passage time was 30 min, re- sulting in 71.6 ng pigment being defecated
Table 2. Gut passage time in adult females of Cal- anus pacificus calculated by dividing the gut content at the end of the incubation by the amount of fecal pig- ment produced during a 6-h incubation of five cope- pods in 2,000 ml of Thalassiosira weissflogii medium. Incubation was on board ship at night, 9 May 1986.
Fecal pigment Gut pigment Passage time Rep. No. (ng cop.-' h-1) (ng cop.-,) (min)
1 28.5 15.1 31.8 2 39.8 12.6 19.0 3 39.8 12.8 19.3 4 35.7 7.4 12.5
This content downloaded from 195.34.79.20 on Sat, 14 Jun 2014 02:48:56 AMAll use subject to JSTOR Terms and Conditions

1068 Dagg et al.
Table 3. Gut passage time in adult females of Cal- anus pacificus, measured as described in Table 2. Ex- periments were done in the laboratory a few days before the August cruise, at several different phytoplankton concentrations.
Fecal pigment Gut pigment Passage time Exp. (ng cop.-' h-') (ng cop.-,) (min)
1 8.7 4.6 32.1 9.8 4.9 30.2 9.7 4.2 25.8 7.9 1.5 11.3
2 5.0 4.5 53.6 3.3 2.7 48.5 3.7 2.5 40.0 3.3 2.2 40.4
3 5.6 7.9 85.0 6.2 8.0 78.0
19.1 19.3 60.1 5.0 1.0 12.2
4 1.6 0.6 22.1 1.6 0.7 25.6 2.5 1.5 35.7 1.3 0.4 19.1
5 22.1 20.3 54.9 18.0 6.3 34.2 38.0 17.4 27.5 17.2 8.1 28.5
during this time. Assuming that feeding continued at a reduced rate during the pe- riod between 0415 and 0615 hours (Fig. 4), when gut contents averaged 2.41 ng cope- pod-' (Table 1), yields an additional fecal production of 9.6 ng pigment copepod-'. Lastly, during the period between 1930 and 2130 hours, some C. pacificus were in the surface layer feeding at a low rate (Fig. 4, Table 1) resulting in an additional 6.7 ng pigment copepod-I for those individuals that were in the surface. The total fecal produc- tion for those C. pacificus that remain long- est in the surface layer is 87.9 ng pigment copepod-I d-l, 81% of which occurred be- tween 2130 and 0415 h.
Similar calculations for C. pacificus in May yield a nightly defecation of 162.6 ng pig- ment copepod-', higher than the August value. The difference is primarily due to the higher average nighttime feeding level (gut content) during May (10.6 ng copepod-I vs. 5.3 in August) and not to a difference in the feeding duration.
In August, each M. lucens fed for the en- tire period spent in the surface layer, from 2130 to 0500 hours, a period of 7.5 h. Gut
passage time was not measured for M. lu- cens but is assumed to be the same as C. pacificus-30 min. Defecation is therefore equivalent to the volume of pigment in 15.0 guts or 125.3 ng pigment copepod-' night-' .
Population effects-The number of C. pa- cificus females in the upper 25 m at mid- night during the August study ranged from 965 to 1,570 m-2 (Fig. 2). Total fecal pro- duction by the population of C. pacificus females during the period of strongest feed- ing was therefore between 69 and 112 ,ug pigment m-2. During the early morning pe- riod of reduced feeding, an additional 9-15 ,ug pigment m-2 was produced. Lastly, the component of the population (arbitrarily as- sumed to be 10%) that feeds for an addi- tional 2 h at a reduced rate in the early evening produced an additional 0.6-1.1 ,ug pigment m-2. The total population of C. pacificus females produced between 79 and 129 ,ug pigment m-2 d-' by defecation.
The number of M. lucens females in the upper 25 m at midnight during the August study ranged from 17,575 to 21,835 m-2 (Fig. 2). Total fecal production by the pop- ulation of M. lucens during the night was therefore between 2,201 and 2,735 ,ug pig- ment m2.
Total fecal production in the upper 25 m by both copepod populations ranged be- tween 2.3 and 2.9 mg pigment m-2 night-'. Incorporation of the uncertainties in gut passage time into these calculations would increase this range. Effectively all of the fe- cal production of pigments occurred be- tween 2130 and 0500 hours.
The amount of pigment actively trans- ported from the surface layer to the deep layer is small. A total of 0.13-0.16 mg pig- ment m-2 is present in the guts of these copepod populations at the time of migra- tory onset, 0.05-0.06 mg m-2 remains in the guts at intermediate depths, and only 0.01-0.02 mg m-2 remains to be defecated at depth by the time the copepods complete their downward migration. This is < 1% of pigment defecated by these copepods in sur- face waters during the night.
Discussion Migration patterns-When populations
of suspension-feeding copepods dielly mi-
This content downloaded from 195.34.79.20 on Sat, 14 Jun 2014 02:48:56 AMAll use subject to JSTOR Terms and Conditions

Copepodfeeding and migration 1069
grate from deep waters containing little or no phytoplankton into surface waters where phytoplankton is abundant, a diel rhythm in the ingestion of phytoplankton is ex- pected (Gauld 1953), has been observed previously (e.g. Simard et al. 1985; Runge and Ingram 1988), and was observed in mi- gratory individuals of C. pacificus and M. lucens in this study. Our intensive sam- pling, however, revealed a temporal pattern of gut pigments that suggested additional behavior modification of ingestion in C. pa- cificus. Although some C. pacificus entered the surface layer up to 2.5 h before sunset, gut pigments increased substantially only well after sunset, indicating low ingestion of phytoplankton during dusk. Similarly, gut pigments declined in C. pacificus before mi- gratory descent, indicating decreased inges- tion of phytoplankton during dawn periods spent in the surface layer. Evidently in C. pacificus there is an additional constraint on feeding behavior, independent of the ver- tical migration, that restricts the most in- tense foraging on phytoplankton to the hours of darkness.
In most previous studies of variations in gut pigments of migrating copepods, the sampling was not sufficiently frequent to de- tect a pattern such as we observed for C. pacificus. However, of particular relevance are reports of nocturnal increase in gut pig- ments in nonmigratory, surface-dwelling copepods (Nicolajsen et al. 1983; Head et al. 1 985; Steams 1986). We agree with Head et al. (1985) and Stearns (1986) that vertical migration and feeding activity may be con- trolled independently and that a major fac- tor constraining feeding to nocturnal hours could be the risk of detection by visual planktivores. Feeding copepods may attract visual predators by their foraging move- ments and enhanced visibility, which re- sults from their guts being packed with food (Steams 1986). The occurrence of adult fe- males of C. pacificus in the stomachs of planktivorous fish collected at night in Da- bob Bay during both May and August cruis- es (Bollens and Frost 1989) indicates that these copepods were susceptible to preda- tion.
Predation is perceived to be a major driv- ing force for not only the occurrence of diel
vertical migration in planktonic copepods (Zaret and Suffern 1976; Fancett and Kim- merer 1985) but also for seasonal and geo- graphical variability of migration behavior (Ohman et al. 1983; Frost 1988). In this study, changes between May and August in the proportion of migrating C. pacificus fe- males and in the timing of migration indi- cate a more cohesive and temporally more focused vertical migration in August than in May. This behavioral change correlates with increased abundance of planktivorous fish which, based on stomach content anal- ysis, were predators of C. pacificus in Dabob Bay (Bollens and Frost 1989).
Enright (1977) offered a very different hy- pothesis to explain the timing of vertical migration and onset of feeding in planktonic suspension-feeders. To optimize their growth rate "migrant grazers ought to begin feeding about an hour or two before sunset" (Enright 1977, p. 871). Our data on C. pa- cificus do not support this hypothesis since most of the population did not arrive in the surface layer until after sunset and early mi- grants ingested phytoplankton at apparently reduced rates for several hours at dusk. Nor were our observations on vertical migration and feeding of M. lucens supportive of En- right's hypothesis. Indeed, we found, as did Enright and Honegger (1977), that in its ver- tical migration M. lucens (=M. pacifica) en- tered and left the surface layer at times high- ly correlated with sunset and sunrise.
The same predatory constraint that af- fected C. pacificus could have been acting on M. lucens to even more tightly coordi- nate its migration in relation to sunset and sunrise, as well as its ingestion of phyto- plankton in relation to migration. Although adult females of M. lucens in Dabob Bay are similar in appearance (color, opacity) and only slightly smaller than adult C. pa- cificus, they are markedly more active (Mackas and Bums 1986; Wong 1988; our unpubl. observations) and hence potentially more conspicuous and attractive to visual zooplanktivores. Adult females ofM. lucens also were found in the stomachs of plank- tivorous fish collected in August (Bollens and Frost 1989).
Gut passage time-Typically, gut passage times have been determined by placing co-
This content downloaded from 195.34.79.20 on Sat, 14 Jun 2014 02:48:56 AMAll use subject to JSTOR Terms and Conditions

1070 Dagg et al.
pepods into filtered water and monitoring the decline in gut contents over time. Gut evacuation measured in this manner has been variously described as a linear (e.g. Yen 1982; Murtaugh 1984), a power (e.g. Baars and Oosterhuis 1984; Huntley et al. 1987), and, most commonly, an exponen- tial rate (e.g. Dagg and Wyman 1983; Ki0r- boe and Tiselius 1987). The form of equa- tion chosen to describe the evacuation process is important because the calculated ingestion rate is a product of the gut content and the gut residence time. It is now clear that food passes through the gut more slowly as the food level in the gut decreases (e.g. Dagg and Walser 1987), suggesting that, for copepods placed in filtered water, the initial part of the evacuation curve is most rep- resentative of the in situ process. Deter- mining which part of the curve is the "ini- tial" part remains somewhat subjective.
A better method is to determine the gut passage time while the copepods are ac- tively feeding (Dagg and Walser 1987; this study). The wide range in gut passage times for C. pacificus in this study, however, sug- gests that further methodological improve- ments are necessary; perhaps a longer in- cubation period or a method for collecting pellets more gently is needed.
The assumption that gut passage time in M. lucens is the same as in C. pacificus is reasonable. The same proportion (about 10%) of gut contents remains in both species after migration to depth in the early morn- ing, suggesting similar gut evacuation rates. Also, similar values have been reported by others; gut passage times of 24 min for C. pacificus and 36 min for M. lucens were measured by Mackas and Burns (1986), and Batchelder (1986) derived a gut passage time for M. lucens (=paciflca) of 54 min (but dis- cusses the possibility that it is an overesti- mate). Reported values for other copepod species are generally in the range of 20-90 min, depending on feeding activity, tem- perature, and experimental conditions (Ki0rboe et al. 1982; Dagg and Wyman 1983; Baars and Oosterhuis 1984; Dagg and Walser 1987; Ki0rboe and Tiselius 1987).
Active transport-Angel (1984) suggested that active vertical transport of material in the guts of migrators may be significant un-
der certain conditions, but it was not here. Only about 10% of the material present in the guts of these copepods at the beginning ofthe early morning descent remained when they reached their daytime depth. Active transport via gut contents is even less sig- nificant (< 1%) when compared to the amount of fecal material produced in the surface layer during the entire night of graz- ing. Furthermore, since pigments produced by grazing copepods within the euphotic zone of Dabob Bay are conserved during sinking to the bottom (Welschmeyer et al. 1984), we can assume the 2.3-2.9 mg pig- ment m-2 night-' defecated in the surface layer at night sinks to the bottom. There- fore, the active transport of pigments to the bottom in the guts of migrating copepods is an insignificant fraction of the total pigment flux in this system. References ANGEL, M. V. 1984. Detrital organic fluxes through
pelagic ecosystems, p. 475-516. In Flows ofenergy and materials in marine ecosystems. NATO Conf. Ser. 4, Mar. Sci. V. 13. Plenum.
BAARS, M. A., AND S. S. OOSTERHUIS. 1984. Diurnal feeding rhythms in North Sea copepods measured by gut fluorescence, digestive enzyme activity and grazing on labelled food. Neth. J. Sea Res. 18: 97- 119.
BARNES, C. A. AND C. C. EBBESMEYER. 1978. Some aspects of Puget Sound's circulation and water properties, p. 209-228. In B. J. Kjerfve [ed.] Es- tuarine transport processes. Univ. S. Carolina.
BATCHELDER, H. P. 1986. Phytoplankton balance in the oceanic subarctic Pacific: Grazing impact of Metridia pacifica. Mar. Ecol. Prog. Ser. 34: 213- 225.
Bt, A. W. H. 1962. Quantitative multiple opening- and-closing plankton samplers. Deep-Sea Res. 9: 144-151.
BOLLENS, S. M., AND B. W. FROST. 1989. Zooplank- tivorous fish and variable diel vertical migration in the marine planktonic copepod Calanus paci- ficus. Limnol. Oceanogr. 34: 1072-1083.
BOYD, C. M., S. L. SMITH, AND T. J. COWLES. 1980. Grazing patterns of copepods in the upwelling sys- tem off Peru. Limnol. Oceanogr. 25: 583-596.
DAGG, M. J., AND D. W. GRILL. 1980. Natural feeding rates of Centropages typicus females in the New York Bight. Limnol. Oceanogr. 25: 597-609.
, AND W. E. WALSER, JR. 1987. Ingestion, gut passage, and egestion by the copepod Neocalanus plumchrus in the laboratory and in the subarctic Pacific Ocean. Limnol. Oceanogr. 32: 178-188.
, AND K. D. WYMAN. 1983. Natural ingestion rates of the copepods Neocalanus plumchrus and N. cristatus calculated from gut contents. Mar. Ecol. Prog. Ser. 13: 37-46.
This content downloaded from 195.34.79.20 on Sat, 14 Jun 2014 02:48:56 AMAll use subject to JSTOR Terms and Conditions

Copepod feeding and migration 1071
EBBESMEYER, C. C. 1973. Some observations of me- dium scale water parcels in a fjord: Dabob Bay, Washington. Ph.D. thesis, Univ. Washington. 212 p.
ENRIGHT, J. T. 1977. Diurnal vertical migration: Adaptive significance and timing. Part 1. Selective advantage: A metabolic model. Limnol. Ocean- ogr. 22: 856-872.
, AND H.-W. HONEGGER. 1977. Diurnal ver- tical migration: Adaptive significance and timing. Part 2. Test of the model: Details of timing. Lim- nol. Oceanogr. 22: 873-886.
FANCETT, M. S., AND W. J. KIMMERER. 1985. Vertical migration of the demersal copepod Pseudodiap- tomus as a means of predator avoidance. J. Exp. Mar. Biol. Ecol. 88: 31-43.
FLEMINGER, A. 1967. Distributional atlas of calanoid copepods in the California current region, part 2. Calif. Mar. Res. Comm. CalCOFI Atlas No. 7. 213 p.
FROST, B. W. 1988. Variability and possible adaptive significance of diel vertical migration in Calanus pacificus, a planktonic marine copepod. Bull. Mar. Sci. 43: 675-694.
GAULD, D. T. 1953. Diurnal variations in the grazing of planktonic copepods. J. Mar. Biol. Assoc. U.K. 31: 461-473.
HEAD, E. J. H., L. R. HARius, AND C. ABou DEBS. 1985. Effect of daylength and food concentration on in situ diurnal feeding rhythms in Arctic co- pepods. Mar. Ecol. Prog. Ser. 24: 281-288.
HUNTLEY, M., V. MARIN, AND F. ESCRITOR. 1987. Zooplankton grazers as transformers of ocean op- tics: A dynamic model. J. Mar. Res. 45: 911-945.
JAMART, B. M., AND D. F. WINTER. 1978. A new approach to the computation of tidal motions in estuaries, p. 261-281. In J. C. J. Nihoul [ed.], Hy- drodynamics of estuaries and fjords. Elsevier.
KI0RBOE, T., F. MOHLENBERG, AND H. NICOLAJSEN. 1982. Ingestion rate and gut clearance in the planktonic copepod Centropages hamatus (Lillje- borg) in relation to food concentration and tem- perature. Ophelia 21: 181-194.
, AND P. TIsELIUs. 1987. Gut clearance and pigment destruction in a herbivorous copepod, Acartia tonsa, and the determination of in situ grazing rates. J. Plankton Res. 9: 525-534.
KOLLMEYER, R. C. 1965. Water properties and cir- culation in Dabob Bay in autumn, 1962. M.S. the- sis, Univ. Washington. 111 p.
LoRENzEN, C. J., AND N. A. WELSCHMEYER. 1983. The in situ sinking rates of herbivore fecal pellets. J. Plankton Res. 5: 929-933.
McGARY, N., AND J. H. LINCOLN. 1977. Tide prints: Surface tidal currents in Puget Sound. Wash. Sea Grant Publ.
MACKAS, D., AND R. BOHRER. 1976. Fluorescence analysis of zooplankton gut contents and an in- vestigation of diel feeding patterns. J. Exp. Mar. Biol. Ecol. 25: 77-85.
, AND K. E. BuRNs. 1986. Poststarvation feed- ing and swimming activity in Calanus pacificus and Metridia pacifica. Limnol. Oceanogr. 31: 383- 392.
MURTAUGH, P. A. 1984. Variable gut residence time: Problems in inferring feeding rate from stomach fullness of a mysid crustacean. Can. J. Fish. Aquat. Sci. 41: 1287-1293.
NICOLAJSEN, H., F. MOLENBERG, AND T. KI0RBOE. 1983. Algal grazing by the planktonic copepods Centropages hamatus and Pseudocalanus sp. Diurnal and seasonal variation during the spring phytoplankton bloom in the Oresund. Ophelia 22: 15-31.
OHMAN, M. D., B. W. FROST, AND E. B. COHEN. 1983. Reverse diel vertical migration: An escape from invertebrate predators. Science 220: 1404-1407.
PARK, T. S. 1968. Calanoidcopepods from the central North Pacific Ocean. Fish. Bull. 66: 527-572.
RUNGE, J. A., AND R. G. INGRAM. 1988. Underice grazing by planktonic calanoid copepods in rela- tion to a bloom of ice microalgae in southeastern Hudson Bay. Limnol. Oceanogr. 33: 280-286.
SImARD, Y., G. LACROIX, AND L. LEGENDRE. 1985. In situ twilight grazing rhythm during diel vertical migrations of a scattering layer of Calanus fin- marchicus. Limnol. Oceanogr. 30: 598-606.
STEARNS, D. E. 1986. Copepod grazing behavior in simulated natural light and its relation to noctur- nal feeding. Mar. Ecol. Prog. Ser. 30: 65-76.
STRICKLAND, J. D. H., AND T. PARSONS. 1968. A practical handbook of seawater analysis. Bull. Fish. Res. Bd. Can. 167.
THORPE, A. C. 1980. Comparative morphology of sibling species of Metridia (Copepoda: Calanoida). M. lucens, M. pacifica, and a species indeterminata from Indian Arm, British Columbia. M.S. thesis, Univ. British Columbia. 75 p.
U.S. DEPARTMENT OF COMMERCE. 1985. Tide tables 1986. High and low water predictions. West Coast of North and South America.
WELSCHMEYER, N. A., A. E. COPPING, M. VERNET, AND C. J. LORENZEN. 1984. Diel fluctuation in zoo- plankton grazing rate as determined from the downward vertical flux of pheopigments. Mar. Biol. 83: 263-270.
WONG, C. K. 1988. The swimming behavior of the copepod Metridia pacifica. J. Plankton Res. 10: 1285-1290.
YEN, J. 1982. Predatory feeding ecology of Euchaeta elongata Esterly, a carnivorous marine copepod. Ph.D. thesis, Univ. Washington. 130 p.
ZARET, T. M., AND J. S. SummERN. 1976. Vertical mi- gration in zooplankton as a predator avoidance mechanism. Limnol. Oceanogr. 21: 804-813.
Submitted: 19 April 1988 Accepted: 25 March 1989
Revised: 12 June 1989
This content downloaded from 195.34.79.20 on Sat, 14 Jun 2014 02:48:56 AMAll use subject to JSTOR Terms and Conditions





























