Embed Size (px)
Citation preview

Convergent Evolution within
Current Biology 22, 2362–2368, December 18, 2012 ª2012 Elsevier Ltd All rights reserved http://dx.doi.org/10.1016/j.cub.2012.10.048
Report
an Adaptive Radiation of Cichlid Fishes
Moritz Muschick,1 Adrian Indermaur,1
and Walter Salzburger1,*1Zoological Institute, University of Basel, 4051 Basel,Switzerland
Summary
The recurrent evolution of convergent forms is a widespread
phenomenon in adaptive radiations (e.g., [1–9]). For ex-ample, similar ecotypes of anoles lizards have evolved on
different islands of the Caribbean [2, 6], benthic-limneticspecies pairs of stickleback fish emerged repeatedly in post-
glacial lakes [1, 3], equivalent sets of spider ecomorphs havearisen on Hawaiian islands [7, 8], and a whole set of conver-
gent species pairs of cichlid fishes evolved in East AfricanLakes Malawi and Tanganyika [10, 11]. In all these cases,
convergent phenotypes originated in geographic isolationfrom each other. Recent theoretical models, however,
predict that convergence should be common withinspecies-rich communities [12, 13], such as species assem-
blages resulting from adaptive radiations. Here, we presentthe most extensive quantitative analysis to date of an
adaptive radiation of cichlid fishes, discovering multipleinstances of convergence in body and trophic morphology.
Moreover, we show that convergent morphologies are asso-ciated with adaptations to specific habitats and resources
and that Lake Tanganyika’s cichlid communities are charac-
terized by the sympatric occurrence of convergent forms.This prevalent coexistence of distantly related yet ecomor-
phologically similar species offers an explanation for thegreatly elevated species numbers in cichlid species flocks.
Results and Discussion
Adaptive radiation, the rapid evolution of a multitude ofspecies from a common ancestor as a consequence of theiradaptation to various ecological niches, is thought to beresponsible for much of the morphological and ecologicaldiversity on earth [4, 9]. Interestingly, parallel adaptive radia-tions of the same group of organisms frequently produceconvergent forms [1–9], which is commonly understood asthe result of independent adaptations to similar ecologicalconditions [3, 4, 14, 15]. Convergence in morphology andbehavior is typically observed between species that evolvedin geographic isolation [2, 3, 7, 10]. Theoretical models, onthe other hand, predict that convergence should also becommon within species-rich communities [12, 13], thus chal-lenging the standard ecological premises that closely relatedspecies should be ecologically similar [16, 17] and that twospecies cannot coexist in the same niche [18]. Such modelssuggest that there is an alternative strategy for enabling stablecoexistence than to be sufficiently distinct: to be sufficientlysimilar. According to these models, convergent evolutionactually appears to be characteristic in ‘‘species-saturated
*Correspondence: [email protected]
communities’’ [12] and to occur when the number of speciesexceeds the number of available niches [13], as is probablythe case in the exceptionally diverse species flocks of cichlidfishes in the East African Great Lakes Victoria, Malawi, andTanganyika.Against this background we explore the cichlid fish assem-
blage of Lake Tanganyika (LT) (Figure 1A) and provide what isto date the most thorough examination of a cichlid adaptiveradiation. Our integrative study combines molecular phyloge-netic, geometric morphometric, and diet analyses in a data setof more than a thousand specimens from 71 species (seeTable S1 available online and Experimental Procedures). Ourmorphological comparisons focus on two ecologically highlyrelevant characters, overall body shape and the shape of thelower pharyngeal jaw bone (LPJ). The LPJ is the central unitof the pharyngeal jaw apparatus, which is a second setof tooth-bearing jaws in the pharynx used to process food[11, 22] (Movie S1). Finally, we use carbon and nitrogen stableisotope ratios as proxy for trophic ecology—in combinationwith stomach and gut content analyses.We first present a robust phylogenetic framework for the
species flock (Figure 1B), which largely agrees with previousstudies [19, 20]. When clustering the species according tobody and LPJ shape, the phylogenetic structure vanishes(Figures 2A and 2C), indicating that the shape of these traitsis largely uncoupled from the phylogenetic background ofa species. All larger cichlid tribes are broken up into two ormore body and LPJ shape clusters, and the different tribesoverlap in morphospace (Figures S1A and S1B). A largefraction of the sister taxa are not each other’s closest ally inthe morphological cluster analyses, and the cluster treesbased on shape data are incongruent with the molecularphylogeny (body shape: D -lnL = 2885.87; D tree length =1059; PSH < 0.001; PKH < 0.001; LPJ shape: D -lnL = 3709.20;D tree length = 1484; PSH < 0.001; PKH < 0.001). Instead ofcorrelating with phylogeny, species that are morphologicallyalike are, in general, more similar in trophic ecology (Figures2 and S1). This integrated analysis leads to two main observa-tions. First, species from distinct clades are grouped into thesame morphoclusters, whereas sister-species are often quitedistinct morphologically (Figure S2); this suggests prevalentconvergence in body and LPJ shape within the cichlid speciesflock of LT. Second, there appears to be a strong link between(trophic) morphology and ecology in LT cichlids; this suggeststhat, just like in other cases of convergent evolution, naturalselection is the driving force in the evolution of convergentforms [1, 5, 15, 23]. In the following, we provide examples forconvergent species and quantify convergence in sympatry inthe cichlid species flock of LT.Perhaps the most striking case of convergent evolution
within LT’s cichlid assemblage involves Neolamprologusprochilus and the enigmatic ‘‘Ctenochromis’’ benthicola (Fig-ure 3A and indicated in bold in Figures 1 and 2). Both speciesoccur sympatrically and are similar to a degree that even localfishermen, who otherwise ably distinguish species, considerthem as one. In line with this, geometric morphometric anal-yses cluster them together, they have similar stable isotopesignatures (Figures 2 and S1), and they show the same

Figure 1. The Cichlid Species Flock of Lake Tanganyika
(A) Map of East Africa showing the three Great Lakes. Lake Tanganyika (LT) is the oldest lake in East Africa and, consequently, accommodates the genet-
ically, morphologically, and ecologically most diverse cichlid species flock [11, 19].
(B) Maximum-likelihood phylogeny of the 71 Tanganyikan cichlid species in our core data set, based on two nuclear (ednrb1, phpt1) and one mitochondrial
(ND2) marker (2,013 bp in total) and the GTR+G model of molecular evolution. Numbers above the branches depict Bayesian posterior probabilities >0.97.
Full species names are given in Table S1; different colors denote the main cichlid lineages (‘‘tribes’’), some of which are likely to have undergone secondary
subradiations [19–21]. Note that the cichlid adaptive radiations of Lakes Malawi and Victoria consist of one of these tribes only, the Haplochromini (the Tan-
ganyikan representatives of which are often referred to as Tropheini) [21]. Our phylogeny confirms the monophyly of the tribes; at least seven genera
are, however, paraphyletic, which already indicates convergence in traits used to classify them initially. For example, the putative haplochromine
‘‘Ctenochromis’’ benthicola (Cteben) emerges as a member of the Cyphotilapiini, whereas its congener, C. horei (Ctehor) remains within the Tropheini/
Haplochromini. The other paraphyletic genera are Gnathochromis (Gna), Lamprologus (Lam), Limnochromis (Lch), Neolamprologus (Neo), Perissodus
(Per), and Petrochromis (Pet). Images of the fishes were taken directly in the field.
Convergence within a Cichlid Adaptive Radiation2363
stomach contents, namely remnants of the endemic shrimpLimnocaridina sp. (Figure 3A). Yet, whereas N. prochilusbelongs to the Lamprologini, ‘‘C.’’ benthicola—formerly con-sidered a Haplochromini and congener of C. horei—nowemerges as a member of the Cyphotilapiini (Figure 1B). Pair-wise genetic distances of 10.6%and 1.4% in themitochondrialand nuclear DNA, respectively, suggest that the two speciesare separated by several million years of independent evolu-tion, which lies in the range of the eye-catching convergentspecies pairs observed between Lakes Tanganyika andMalawi [10]. But cichlids do not only resemble other endemiccichlids. The rare Baileychromis centropomoides, for
example, is very similar in overall body shape to an endemicLates sp. (Figures 3B and S3).To quantify convergence in the LT cichlid species flock, we
plotted relative morphological distance against phylogeneticdistance for each pair of species and compared it to simula-tions of trait evolution (Figure 4A). Applying a conservativethreshold (see Experimental Procedures), we identify 122and 132 species pairs that are convergent in body and LPJshape, respectively, which is about five times more than pre-dicted by the models. Importantly, more than three quartersof these convergent species pairs overlap in habitat and depthdistribution (Table S2), and they show a significantly greater

Figure 2. Ecomorphological Diversity in Cichlids from Lake Tanganyika
(A) Cluster analysis on the basis of 17 homologous landmarks on body shape.
(B) d13C stable isotope signatures.
(C) Cluster analysis on the basis of eight homologous and six sliding landmarks on the lower pharyngeal jaw bone.
(D) d15N stable isotope signatures.
(E) Results from the stomach and gut content analyses (in volume %).
Outlines in (A) are based on real photographs; images in (C) are taken fromdissected LPJs (see Table S1 for details). Themainmorphoclusters are separated
by gray lines, and the tribes are colored as in Figure 1. Colored dots in (B) and (D) represent average values; gray bars indicate 95% confidence limits of a t
distribution. *marks specieswith too small a sample size, so that 95%confidence intervals were not calculated. The ratio between the rare isotope 13C to 12C
(the d13C value) indicates theprimary carbon source,whichmay vary betweenmacrohabitats (e.g., benthic versus pelagic), whereas the d15N value (15N to 14N)
serves as proxy for the relative trophic level of an organism. Accordingly, in LT cichlids, d13C values correlate with body shape clusters (F = 2.66, p < 0.005),
whereas d15N values correlate with LPJ shape (F = 4.03, p < 0.005). Note that each trophic level is separated by approximately 3.4‰ in d15N from the one
below. To facilitate comparisons, we also included average stable isotope values for some plant and animal species from LT (see box at the bottom).
Current Biology Vol 22 No 242364
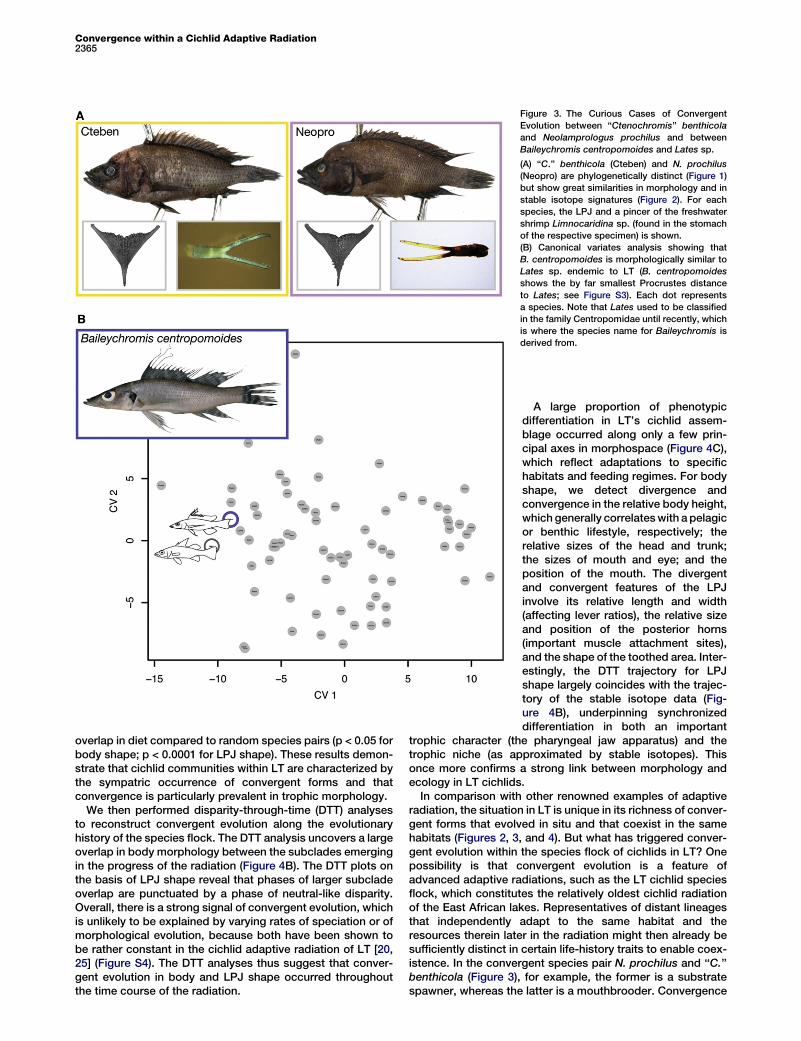
Figure 3. The Curious Cases of Convergent
Evolution between ‘‘Ctenochromis’’ benthicola
and Neolamprologus prochilus and between
Baileychromis centropomoides and Lates sp.
(A) ‘‘C.’’ benthicola (Cteben) and N. prochilus
(Neopro) are phylogenetically distinct (Figure 1)
but show great similarities in morphology and in
stable isotope signatures (Figure 2). For each
species, the LPJ and a pincer of the freshwater
shrimp Limnocaridina sp. (found in the stomach
of the respective specimen) is shown.
(B) Canonical variates analysis showing that
B. centropomoides is morphologically similar to
Lates sp. endemic to LT (B. centropomoides
shows the by far smallest Procrustes distance
to Lates; see Figure S3). Each dot represents
a species. Note that Lates used to be classified
in the family Centropomidae until recently, which
is where the species name for Baileychromis is
derived from.
Convergence within a Cichlid Adaptive Radiation2365
overlap in diet compared to random species pairs (p < 0.05 forbody shape; p < 0.0001 for LPJ shape). These results demon-strate that cichlid communities within LT are characterized bythe sympatric occurrence of convergent forms and thatconvergence is particularly prevalent in trophic morphology.
We then performed disparity-through-time (DTT) analysesto reconstruct convergent evolution along the evolutionaryhistory of the species flock. The DTT analysis uncovers a largeoverlap in body morphology between the subclades emergingin the progress of the radiation (Figure 4B). The DTT plots onthe basis of LPJ shape reveal that phases of larger subcladeoverlap are punctuated by a phase of neutral-like disparity.Overall, there is a strong signal of convergent evolution, whichis unlikely to be explained by varying rates of speciation or ofmorphological evolution, because both have been shown tobe rather constant in the cichlid adaptive radiation of LT [20,25] (Figure S4). The DTT analyses thus suggest that conver-gent evolution in body and LPJ shape occurred throughoutthe time course of the radiation.
A large proportion of phenotypicdifferentiation in LT’s cichlid assem-blage occurred along only a few prin-cipal axes in morphospace (Figure 4C),which reflect adaptations to specifichabitats and feeding regimes. For bodyshape, we detect divergence andconvergence in the relative body height,which generally correlateswith a pelagicor benthic lifestyle, respectively; therelative sizes of the head and trunk;the sizes of mouth and eye; and theposition of the mouth. The divergentand convergent features of the LPJinvolve its relative length and width(affecting lever ratios), the relative sizeand position of the posterior horns(important muscle attachment sites),and the shape of the toothed area. Inter-estingly, the DTT trajectory for LPJshape largely coincides with the trajec-tory of the stable isotope data (Fig-ure 4B), underpinning synchronizeddifferentiation in both an important
trophic character (the pharyngeal jaw apparatus) and thetrophic niche (as approximated by stable isotopes). Thisonce more confirms a strong link between morphology andecology in LT cichlids.In comparison with other renowned examples of adaptive
radiation, the situation in LT is unique in its richness of conver-gent forms that evolved in situ and that coexist in the samehabitats (Figures 2, 3, and 4). But what has triggered conver-gent evolution within the species flock of cichlids in LT? Onepossibility is that convergent evolution is a feature ofadvanced adaptive radiations, such as the LT cichlid speciesflock, which constitutes the relatively oldest cichlid radiationof the East African lakes. Representatives of distant lineagesthat independently adapt to the same habitat and theresources therein later in the radiation might then already besufficiently distinct in certain life-history traits to enable coex-istence. In the convergent species pair N. prochilus and ‘‘C.’’benthicola (Figure 3), for example, the former is a substratespawner, whereas the latter is a mouthbrooder. Convergence
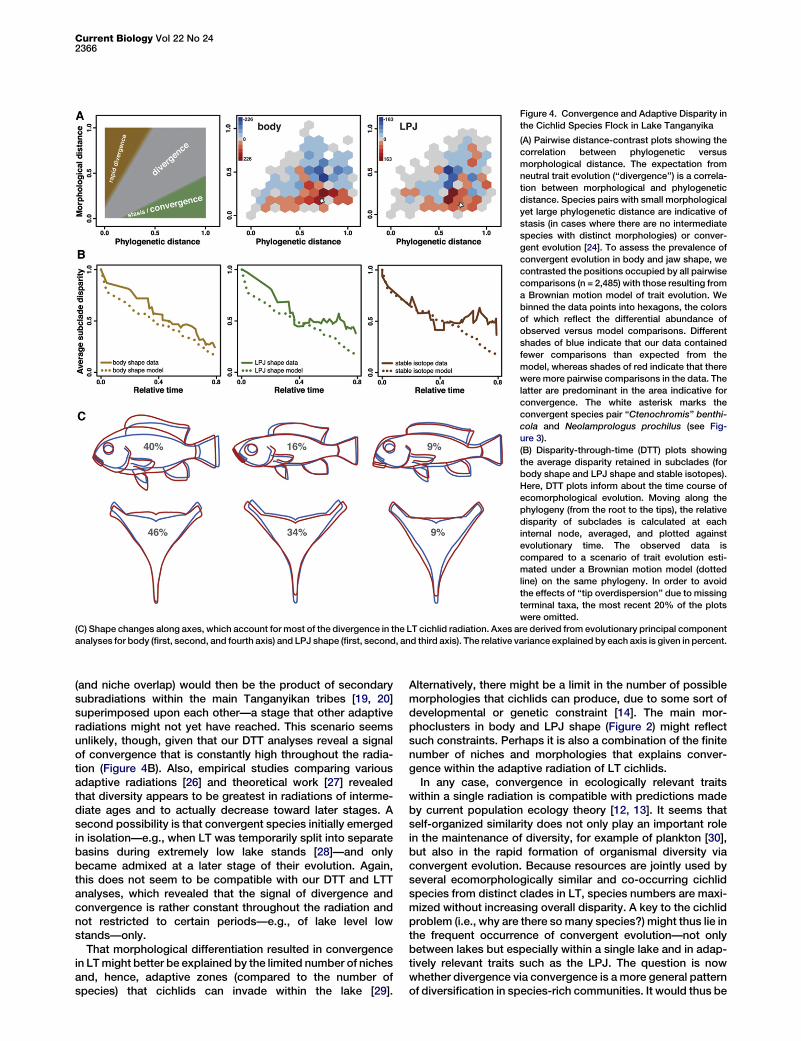
Figure 4. Convergence and Adaptive Disparity in
the Cichlid Species Flock in Lake Tanganyika
(A) Pairwise distance-contrast plots showing the
correlation between phylogenetic versus
morphological distance. The expectation from
neutral trait evolution (‘‘divergence’’) is a correla-
tion between morphological and phylogenetic
distance. Species pairs with small morphological
yet large phylogenetic distance are indicative of
stasis (in cases where there are no intermediate
species with distinct morphologies) or conver-
gent evolution [24]. To assess the prevalence of
convergent evolution in body and jaw shape, we
contrasted the positions occupied by all pairwise
comparisons (n = 2,485) with those resulting from
a Brownian motion model of trait evolution. We
binned the data points into hexagons, the colors
of which reflect the differential abundance of
observed versus model comparisons. Different
shades of blue indicate that our data contained
fewer comparisons than expected from the
model, whereas shades of red indicate that there
were more pairwise comparisons in the data. The
latter are predominant in the area indicative for
convergence. The white asterisk marks the
convergent species pair ‘‘Ctenochromis’’ benthi-
cola and Neolamprologus prochilus (see Fig-
ure 3).
(B) Disparity-through-time (DTT) plots showing
the average disparity retained in subclades (for
body shape and LPJ shape and stable isotopes).
Here, DTT plots inform about the time course of
ecomorphological evolution. Moving along the
phylogeny (from the root to the tips), the relative
disparity of subclades is calculated at each
internal node, averaged, and plotted against
evolutionary time. The observed data is
compared to a scenario of trait evolution esti-
mated under a Brownian motion model (dotted
line) on the same phylogeny. In order to avoid
the effects of ‘‘tip overdispersion’’ due to missing
terminal taxa, the most recent 20% of the plots
were omitted.
(C) Shape changes along axes, which account for most of the divergence in the LT cichlid radiation. Axes are derived from evolutionary principal component
analyses for body (first, second, and fourth axis) and LPJ shape (first, second, and third axis). The relative variance explained by each axis is given in percent.
Current Biology Vol 22 No 242366
(and niche overlap) would then be the product of secondarysubradiations within the main Tanganyikan tribes [19, 20]superimposed upon each other—a stage that other adaptiveradiations might not yet have reached. This scenario seemsunlikely, though, given that our DTT analyses reveal a signalof convergence that is constantly high throughout the radia-tion (Figure 4B). Also, empirical studies comparing variousadaptive radiations [26] and theoretical work [27] revealedthat diversity appears to be greatest in radiations of interme-diate ages and to actually decrease toward later stages. Asecond possibility is that convergent species initially emergedin isolation—e.g., when LT was temporarily split into separatebasins during extremely low lake stands [28]—and onlybecame admixed at a later stage of their evolution. Again,this does not seem to be compatible with our DTT and LTTanalyses, which revealed that the signal of divergence andconvergence is rather constant throughout the radiation andnot restricted to certain periods—e.g., of lake level lowstands—only.
That morphological differentiation resulted in convergencein LTmight better be explained by the limited number of nichesand, hence, adaptive zones (compared to the number ofspecies) that cichlids can invade within the lake [29].
Alternatively, there might be a limit in the number of possiblemorphologies that cichlids can produce, due to some sort ofdevelopmental or genetic constraint [14]. The main mor-phoclusters in body and LPJ shape (Figure 2) might reflectsuch constraints. Perhaps it is also a combination of the finitenumber of niches and morphologies that explains conver-gence within the adaptive radiation of LT cichlids.In any case, convergence in ecologically relevant traits
within a single radiation is compatible with predictions madeby current population ecology theory [12, 13]. It seems thatself-organized similarity does not only play an important rolein the maintenance of diversity, for example of plankton [30],but also in the rapid formation of organismal diversity viaconvergent evolution. Because resources are jointly used byseveral ecomorphologically similar and co-occurring cichlidspecies from distinct clades in LT, species numbers are maxi-mized without increasing overall disparity. A key to the cichlidproblem (i.e., why are there somany species?) might thus lie inthe frequent occurrence of convergent evolution—not onlybetween lakes but especially within a single lake and in adap-tively relevant traits such as the LPJ. The question is nowwhether divergence via convergence is a more general patternof diversification in species-rich communities. It would thus be

Convergence within a Cichlid Adaptive Radiation2367
of great interest to extend the kind of integrative analysisimplemented in this study to other adaptive radiations and,especially, to the cichlid adaptive radiations in Lakes Malawiand Victoria. Even more so, because a recent comparisonacross 46 cichlid adaptive radiations [31] suggests that theLT radiation is an outlier from an otherwise more general trendin cichlid radiations, which appear to be triggered by bothecological opportunity and sexual selection.
Experimental Procedures
Sampling
Sampling was performed under permission from the Department of Fish-
eries, Lake Tanganyika Research Unit, Mpulungu, Zambia. In total, we
sampled more than 1,000 specimens for this study (see Supplemental
Experimental Procedures and Table S1 for further details).
Phylogenetic Analyses
We analyzed one mitochondrial (ND2) and two nuclear (ednrb1, phpt)
markers (see Supplemental Experimental Procedures and Table S1 for
GenBank accession numbers used in this study). We relied on maximum
likelihood and Bayesian methods for phylogenetic analysis using PAUP*,
MRBAYES, and the BEAST package. The appropriate model of molecular
evolution for the heuristic tree searches in PAUP* was determined with
JMODELTEST; MRBAYES was run for ten million generations with a burn-
in of 10%; data were partitioned in BEAST. We first analyzed our core
data set combining the mitochondrial and nuclear DNA sequences in 71
taxa, then the core data set including Baileychromis centropomoides,
and, finally, a mitochondrial data set including the ND2 sequences of 180
taxa (i.e., ca. 90% of all Tanganyika species). Trees derived from the latter
analysis were used for lineage-through-time plots. For incongruence
testing, we applied the Kishino-Hasegawa (KH) and the Shimodaira-
Hasegawa (SH) test implemented in PAUP*.
Geometric Morphometric and Morphological Analyses
We assessed the body shape of 1,049 individuals using landmark-based
geometric morphometrics. xy coordinates of 17 landmarks, distributed
across the whole fish body (see Figure S5A), and the scale of each picture
were recorded using TPSDIG [32]. Aligned Procrustes coordinates were
used for a pooled-within-species regression of shape against centroid
size in MORPHOJ 1.02d [33]. Species averages were then used for principal
component analysis (PCA), for disparity-through-time analyses, and for the
calculation of pairwise distances between species. For LPJ assessment we
recorded coordinates of eight true landmarks and 20 semilandmarks
describing the outline of the bone (Figure S5B). We then clustered the
species according to similarity in body and LPJ shape, using agglomerative
hierarchical clustering in R.
Stomach and Gut Content Analyses
Contents were removed from the intestinal tracts of 506 specimens and
separated up into one or more of the following categories: sand, aufwuchs
(algae), plantmaterial, mollusks, insects (imagines and larvae), crustaceans,
fish (remains), fish eggs, and fish scales. We determined volume (in %) and
weight (in mg) of each category.
Stable Isotope Analysis
White muscle tissue from 727 specimens (see Table S1) was dried,
pulverized, and analyzed on an elemental analyzer (Thermo Finnigan)
coupled to a Finnigan Delta V Advantage Isotope Ratio Mass Spectrometer
(IRMS).
Pairwise Distance-Contrast Plots
To estimate the extent of convergence, we compared the phylogenetic
distance to the morphological distance of each species pair [24]. The
morphological distance was calculated as Euclidean distance from the
pooled-within-species regressions of shape against centroid size using
R’s dist() function. In total, we had 2,485 species comparisons; therefore,
we used hexagonal binning (x = 10 bins) to overcome overplotting. We
also simulated neutral trait evolution on the phylogeny, using Brownian
motion and Ornstein-Uhlenbeck models. Species comparisons that we
derived from these simulations were then compared to our actual data by
subtracting the binning counts of the simulations from those of the data.
We tested for statistical significance of the difference of pointwise means
between simulations and data (each 1/10 of the x axis) by bootstrapping
(1,000 replications).
Disparity-through-Time Analysis
DTT analyses were performed according to Harmon et al. [34], compar-
ing the observed data to a scenario of trait evolution estimated under a
Brownianmotionmodel. Positive deviations of the data from the simulations
indicate a higher overlap in morphospace among subclades than would be
expected under neutral evolution.
Evolutionary PCA
We estimated the ancestral character states for body and LPJ shape at
each node in the phylogeny and calculated the extent and the direction of
shape change along each branch. These branchwise estimates were then
subjected to PCA to find the axes of greatest evolutionary divergence. All
evolutionary PCAs were performed in MORPHOJ.
Supplemental Information
Supplemental Information includes five figures, two tables, Supplemental
Experimental Procedures, and one movie and can be found with this article
online at http://dx.doi.org/10.1016/j.cub.2012.10.048.
Acknowledgments
We thank H. Buscher and various past and present members of the Salzbur-
gerLab for help during fieldwork campaigns; S. Koblmuller for discussions
and help with species identifications; B. Aeschbach, N. Boileau, and
F. Munzel for assistance in the lab; M. Lehmann for help with stable isotope
analyses; H. Bichsel, T. Bosia, M. Gschwind, I. Keller, M. Maurer, N. Rose,
and F. Ronco for help with the stomach content analyses; F. Meury, D.
Moser, W. Moser, and C. Mullis for assistance with transect surveys; J.P.
Montoya and S. Chraiti for help with the CT scans; the Department of Fish-
eries, Republic of Zambia, for research permits and technical support; and
H. Buscher, L. Harmon, L. Keller, C.P. Klingenberg, J.B. Losos, P. Nosil, S.
Ramm, L. Scharer, M. Scheffer, D. Schluter, J.T. Streelman, M. Taylor, the
SalzburgerLab, and two anonymous referees for discussion and valuable
comments. This work was supported by the European Research Council
(ERC; Starting Grant ‘‘INTERGENADAPT’’), the Swiss National Science
Foundation, the National Geographic Society, and the University of Basel.
Received: September 12, 2012
Revised: October 25, 2012
Accepted: October 29, 2012
Published: November 15, 2012
References
1. Schluter, D., and Nagel, L.M. (1995). Parallel speciation by natural
selection. Am. Nat. 146, 292–301.
2. Losos, J.B., Jackman, T.R., Larson, A., Queiroz, K., and Rodriguez-
Schettino, L. (1998). Contingency and determinism in replicated
adaptive radiations of island lizards. Science 279, 2115–2118.
3. Rundle, H.D., Nagel, L., Wenrick Boughman, J., and Schluter, D. (2000).
Natural selection and parallel speciation in sympatric sticklebacks.
Science 287, 306–308.
4. Schluter, D. (2000). The Ecology of Adaptive Radiation (New York:
Oxford University Press).
5. Nosil, P., Crespi, B.J., and Sandoval, C.P. (2002). Host-plant adaptation
drives the parallel evolution of reproductive isolation. Nature 417,
440–443.
6. Losos, J.B., Leal, M., Glor, R.E., De Queiroz, K., Hertz, P.E., Rodrıguez
Schettino, L., Lara, A.C., Jackman, T.R., and Larson, A. (2003). Niche
lability in the evolution of a Caribbean lizard community. Nature 424,
542–545.
7. Blackledge, T.A., and Gillespie, R.G. (2004). Convergent evolution of
behavior in an adaptive radiation of Hawaiian web-building spiders.
Proc. Natl. Acad. Sci. USA 101, 16228–16233.
8. Gillespie, R. (2004). Community assembly through adaptive radiation in
Hawaiian spiders. Science 303, 356–359.
9. Gavrilets, S., and Losos, J.B. (2009). Adaptive radiation: contrasting
theory with data. Science 323, 732–737.

Current Biology Vol 22 No 242368
10. Kocher, T.D., Conroy, J.A., McKaye, K.R., and Stauffer, J.R. (1993).
Similar morphologies of cichlid fish in Lakes Tanganyika and Malawi
are due to convergence. Mol. Phylogenet. Evol. 2, 158–165.
11. Salzburger, W. (2009). The interaction of sexually and naturally selected
traits in the adaptive radiations of cichlid fishes. Mol. Ecol. 18, 169–185.
12. Scheffer, M., and van Nes, E.H. (2006). Self-organized similarity, the
evolutionary emergence of groups of similar species. Proc. Natl.
Acad. Sci. USA 103, 6230–6235.
13. terHorst, C.P., Miller, T.E., and Powell, E. (2010). When can competition
for resources lead to ecological equivalence? Evol. Ecol. Res. 12,
843–854.
14. Brakefield, P.M. (2006). Evo-devo and constraints on selection. Trends
Ecol. Evol. 21, 362–368.
15. Losos, J.B. (2011). Convergence, adaptation, and constraint. Evolution
65, 1827–1840.
16. Peterson, A.T., Soberon, J., and Sanchez-Cordero, V. (1999).
Conservatism of ecological niches in evolutionary time. Science 285,
1265–1267.
17. Losos, J.B. (2008). Phylogenetic niche conservatism, phylogenetic
signal and the relationship between phylogenetic relatedness and
ecological similarity among species. Ecol. Lett. 11, 995–1003.
18. Gause, G.F. (1934). The Struggle for Existence (Baltimore: The Williams
and Wilkins Company).
19. Salzburger, W., Meyer, A., Baric, S., Verheyen, E., and Sturmbauer, C.
(2002). Phylogeny of the Lake Tanganyika cichlid species flock and its
relationship to the Central and East African haplochromine cichlid fish
faunas. Syst. Biol. 51, 113–135.
20. Day, J.J., Cotton, J.A., and Barraclough, T.G. (2008). Tempo and mode
of diversification of lake Tanganyika cichlid fishes. PLoS One 3, e1730.
21. Salzburger, W., Mack, T., Verheyen, E., and Meyer, A. (2005). Out of
Tanganyika: genesis, explosive speciation, key-innovations and
phylogeography of the haplochromine cichlid fishes. BMC Evol. Biol.
5, 17.
22. Fryer, G., and Iles, T.D. (1972). The Cichlid Fishes of the Great Lakes of
Africa: Their Biology and Evolution (Edinburgh: Oliver & Boyd).
23. Harmon, L.J., Kolbe, J.J., Cheverud, J.M., and Losos, J.B. (2005).
Convergence and the multidimensional niche. Evolution 59, 409–421.
24. Winemiller, K.O. (1991). Ecomorphological diversification in lowland
freshwater fish assemblages from five biotic regions. Ecol. Monogr.
61, 343–365.
25. Hoerner, M.E. (2011). Testing for differences in rates of speciation,
extinction, and morphological evolution in four tribes of cichlids
endemic to Lake Tanganyika, East Africa. Evolution 65, 3398–3412.
26. Gillespie, R.G., andBaldwin, B.G. (2009). Island biogeography of remote
archipelagoes. In The Theory of Island Biogeography Revisited, J.B.
Losos and R.E. Ricklefs, eds. (Princeton, NJ: Princeton University
Press), pp. 358–387.
27. Gavrilets, S., and Vose, A. (2005). Dynamic patterns of adaptive radia-
tion. Proc. Natl. Acad. Sci. USA 102, 18040–18045.
28. Cohen, A.S., Lezzar, K.E., Tiercelin, J.J., and Soreghan, M. (1997). New
paleogeographic and lake-level reconstructions of Lake Tanganyika:
implications for tectonic, climatic and biological evolution in a rift lake.
Basin Research 9, 107–132.
29. Ruber, L., and Adams, D.C. (2001). Evolutionary convergence of body
shape and trophic morphology in cichlids from Lake Tanganyika.
J. Evol. Biol. 14, 325–332.
30. Beninca, E., Huisman, J., Heerkloss, R., Johnk, K.D., Branco, P., Van
Nes, E.H., Scheffer, M., and Ellner, S.P. (2008). Chaos in a long-term
experiment with a plankton community. Nature 451, 822–825.
31. Wagner, C.E., Harmon, L.J., and Seehausen, O. (2012). Ecological
opportunity and sexual selection together predict adaptive radiation.
Nature 487, 366–369.
32. Rohlf, F.J. (2006). tpsDIG, Version 2.10. Department of Ecology and
Evolution (Stony Brook, NY: State University of New York).
33. Klingenberg, C.P. (2011). MorphoJ: an integrated software package for
geometric morphometrics. Mol. Ecol. Resour. 11, 353–357.
34. Harmon, L.J., Schulte, J.A., 2nd, Larson, A., and Losos, J.B. (2003).
Tempo and mode of evolutionary radiation in iguanian lizards. Science
301, 961–964.