Embed Size (px)
Citation preview

J. Insect Physiol. Vol. 43, No. 11, pp. 1015–1021, 1997 1997 Elsevier Science LtdPergamon All rights reserved. Printed in Great Britain0022-1910/97 $17.00 + 0.00PII: S0022-1910(97)00073-5
Contributions to Physiology of the Antenna–Heart in Periplaneta americana (L.) (Blattodea:Blattidae)*MICHAEL RICHTER,† WIELAND HERTEL†‡
Received 17 March 1997; revised 19 May 1997
An accessory pulsatile organ of an open circulatory system in insects supplying the antennaewith haemolymph was investigated. The rhythm of this so-called antenna–heart is generatedby a myogenic automatism and can be neuronally influenced via the nervus cardioantennalis.
The action potentials of the muscle fibres show typical pre-depolarization and mostly noovershoot. A specific membrane resistance (Rm) of about 660 V cm−2 was calculated for thefibres. Some electrical coupling between the muscle fibres is presumed for synchronization ofany myogenically triggered heart beat which could actually be proved experimentally by cur-rent injection in the antenna–heart. However, intercalated disks or gap junctions could notbe found. Nevertheless, a good coupling factor (U2/U1) between all fibres was demonstratedby parallel recordings and can be well described by a conductance model according to fibretopology. 1997 Elsevier Science Ltd. All rights reserved
Periplaneta americana Antenna–heart Myogenic automatism Coupling coefficient Conductance model
INTRODUCTION
Due to the inefficiency of an open circulatory system,long body appendages could not be supplied adequatelywith haemolymph in insects. Therefore, it was necessaryto develop independently working accessory pulsatileorgans such as the antenna–hearts in the head of mostectognatha (Pass, 1985, 1991; Hertel et al., 1985, inpress).
The antenna–heart of Periplaneta americana consistsof two ampullae connected by a thin transversal muscle,the dilator muscle. The non-contractile ampullae arelocated close to the cuticle in front of each antenna andhaemolymph taken in via ostia during contraction of thedilator muscle, is forced through the antennal vessels intothe antennae during muscle dilation (Pass, 1985). Theaction of the transversal dilator is supported by theaccessory dilator muscle which holds it in a slightly tenseposition. The fibres of the accessory dilator, which differmorphologically but not physiologically from transverse
*Dedicated to Prof. Dr. Heinz Penzlin on the occasion of his 65th birth-day.
†Institut fur Allgemeine Zoologie und Tierphysiologie, Friedrich-Schiller-Universitat Jena, Erbertstraße 1, D-07743 Jena, Germany.
‡To whom all correspondence should be addressed.
1015
muscle fibres, are bound into the transverse dilator(Hertel et al., in press). Earlier investigations showed thatthe electrophysiological parameters of the antenna–heartmuscle fibres are comparable with that of the dorsal ves-sel. There is clear evidence for a myogenic automatismof both organs (McCann, 1965; Miller, 1969; Hertel,1971; Hertel et al., 1985). In contrast to the dorsal vessel,the pacemaker activity of the antenna–heart is powerfullyinfluenced by the neuropeptide proctolin (Hertel et al.,1997a).
Electrical coupling between the muscle fibres is anessential requirement for synchronization of the myogen-ically triggered heart beat. Interlocking muscle fibres andthe presence of intercalated disks as are known from ver-tebrate muscles could not be found in the antenna–heart(Hertel et al., in press). Thus, transmission of electricalpotential changes has to occur along the cross directionof the dilator muscle, provided that sufficient current flowis possible. This requires low ohmic contact sitesbetween the fibres whose resistances clearly differ fromthose of membrane resistance. The relationship betweenthe two conductances is characterized by the couplingcoefficient (Socolar, 1977).

1016 MICHAEL RICHTER AND WIELAND HERTEL
MATERIALS AND METHODS
Preparation of experimental animals
For all experiments, male adult Periplaneta americanawere used. The cockroaches were reared under a photop-eriod ratio of 12:12 daylight/darkness ratio and a tem-perature of about 28°C. The antenna–hearts were isolatedfrom the head as were parts of the frontal cuticle of thehead capsule and the antennae. In order to transilluminateand expose the muscle, making it accessible to electro-physiological investigations, an opening was cut into thecuticle between the bases of the antennae. All experi-ments were carried out at room temperature in a modifiedHEPES-buffered ringer solution of mmol l−1: NaCl140.0, KCl 5.0, CaCl2 5.0, MgCl2 1.0, NaHCO3 4.0, glu-cose 5.0, HEPES 5.0, pH adjusted to 7.2 (Hertel et al.,1985).
Experimental equipment
Intracellular electrophysiological measurements werecarried out using conventional microelectrode tech-niques. Floating glass microelectrodes filled with3 mol l−1 KCl or Lucifer yellow were used for intracellu-lar recordings of the beating antenna–heart. In addition,hyperpolarizing current pulses were injected into themuscle cells via the intracellular electrodes by means ofa bridge circuit in order to measure membrane resistanceRinput. In most cases, parallel recordings were obtainedwith two or three floating electrodes to determine thecoupling coefficient. The signals were simultaneouslyrecorded by a storage oscilloscope and a digital taperecorder.
The values represent mean values ± standard devi-ations.
Transmission electron microscopy
For electron microscopy, the hearts were fixed in 2%glutaraldehyde and 2% formaldehyde in 0.1 mol l−1 phos-phate buffer, pH 7.3 for 1 h followed by washing for 1 hand post-fixing with 1% OsO4 in phosphate buffer. Afterthat the antenna-hearts were dehydrated in acetone andembedded in Durcupan ACM. Furthermore, the ultrathinsections were double-stained in uranyl acetate and leadcitrate.
RESULTS
In vitro, the antenna–heart displayed a beat rate of 20–50 min−1 without being synchronized with the frequencyof the dorsal vessel. It produced a power of 136 ± 26 mN(maximum initial stressing 50 mN) at every rhythmicalcontraction.
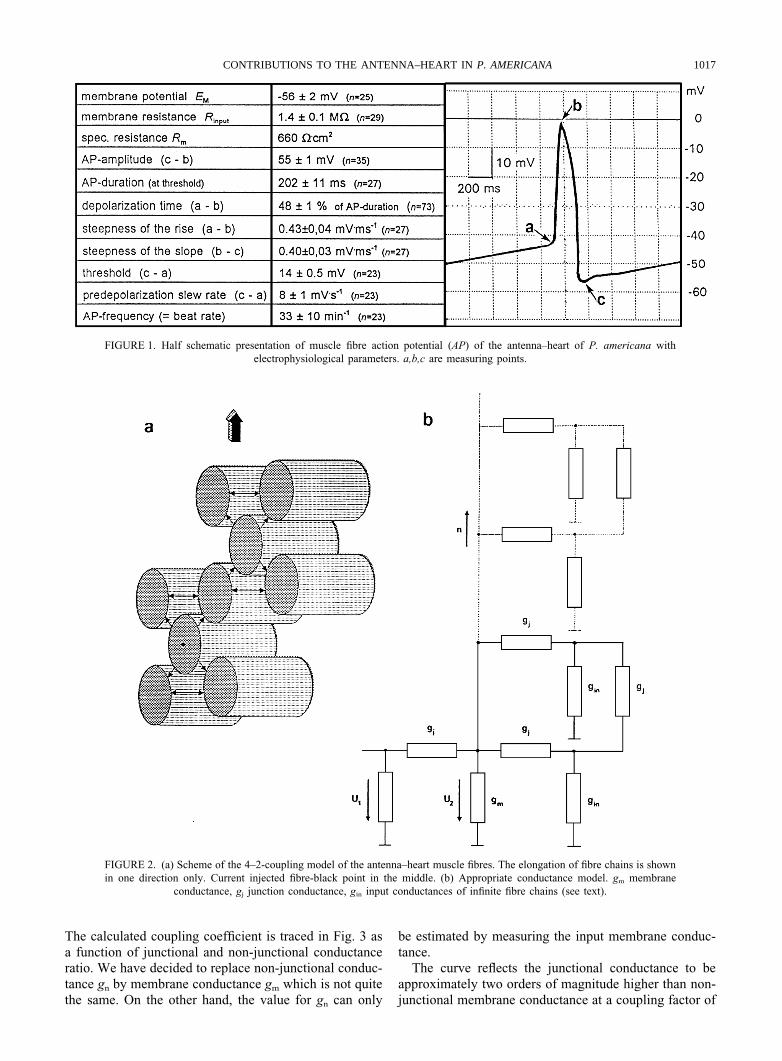
The resting potential (Fig. 1) was defined as themaximum negative membrane potential after repolariz-ation since a new pre-depolarization, the so-called pace-maker activity, restarts at once. All fibres investigatedshowed the same potential course so that no pacemakercentre could be detected. The almost symmetrical and
non-graded action potential was long in duration andmostly without overshoot. The typical values are shownin Fig. 1.
With a cylindrical muscle fibre of about 10 mm indiameter and 1500 mm in length, a specific membraneresistance of Rm = 660 V cm−2 could be calculated.
Electrical coupling in the antenna–heart
Theoretical view. According to the way the current isdivided via electrical contacts between the muscle fibres,an injected current produces a lower voltage drop in anadjacent fibre (Fig. 4). The coupling-quality is estimatedby the coupling-coefficient. It represents the relationshipbetween U2 (voltage drop in the coupled cell) and U1
(input voltage of the current injected cell). According tomorphological findings (Hertel et al., in press), we startout with the assumption that each muscle fibre is electri-cally coupled to 4 other fibres on average in the antenna–heart. Two out of these 4 should normally couple againwith each other. Providing that this 4–2-interconnectionensemble applies for the whole dilator muscle (Fig. 2a),the simple conductance model in Fig. 2b enables the cal-culation of the coupling coefficient U2/U1. Cell 1 withinput conductance gin and non-junctional membrane con-ductance gm couples to n (in case of antenna–heart = 4)cells via junctional conductance gj. Two (n/2 = k) out ofthese 4 cells are linked again via gj with each other. Theprolongation of electrically coupled cell chains is esti-mated by input conductances gin. Under the theoreticalpresumption that all cells have identical values for gj andgm, the perijunctional conductances within the membraneare very minor in comparison with gj and gm and thatintracellular and extracellular medium can be regarded asisopotential, the input conductance of cell 1 is defined as:
gin = gm + 2gjgin
gj + gin
(1)
Equation (1) presumes the input conductance of cell 1 todiffer not from the input conductances of adjacent cellsdue to a supposed infinite chain length. If current isinjected into the first cell generating the membrane volt-age U1 = I1
.gin−1, a part of I1 will cause the voltage drop
U2 in a coupled cell. With the inclusion of Equation (1),the coupling coefficient is given by
U2
U1=
gj
gj + gm + k(gin − gm)k = n/2 (2)
and after solving of Equation (1) for gin and insertioninto Equation (2):
U2
U1=
gj
gm
1 +gj
gm
+ k112
gj
gm
−12
+ !14Sgj
gmD2
+32
gj
gm
+142
1017CONTRIBUTIONS TO THE ANTENNA–HEART INP. AMERICANA
FIGURE 1. Half schematic presentation of muscle fibre action potential (AP) of the antenna–heart of P. americana withelectrophysiological parameters. a,b,c are measuring points.
FIGURE 2. (a) Scheme of the 4–2-coupling model of the antenna–heart muscle fibres. The elongation of fibre chains is shownin one direction only. Current injected fibre-black point in the middle. (b) Appropriate conductance model. gm membrane
conductance, gj junction conductance, gin input conductances of infinite fibre chains (see text).
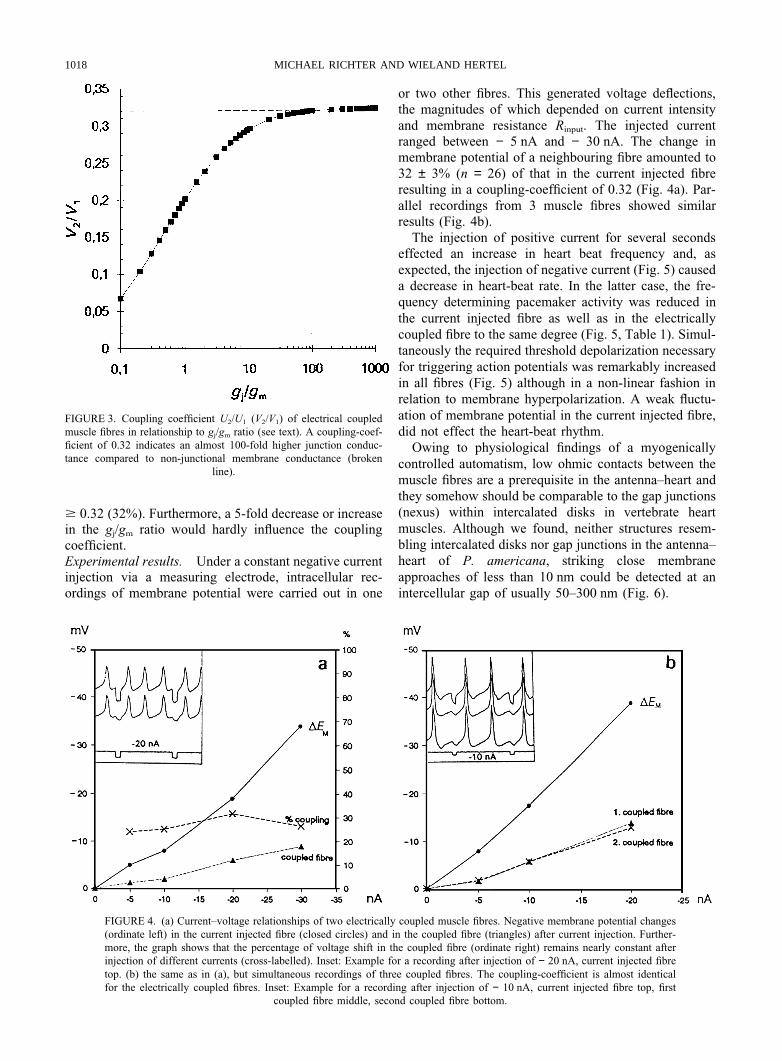
The calculated coupling coefficient is traced in Fig. 3 asa function of junctional and non-junctional conductanceratio. We have decided to replace non-junctional conduc-tance gn by membrane conductance gm which is not quitethe same. On the other hand, the value for gn can only
be estimated by measuring the input membrane conduc-tance.
The curve reflects the junctional conductance to beapproximately two orders of magnitude higher than non-junctional membrane conductance at a coupling factor of
1018 MICHAEL RICHTER AND WIELAND HERTEL
FIGURE 3. Coupling coefficient U2/U1 (V2/V1) of electrical coupledmuscle fibres in relationship to gj/gm ratio (see text). A coupling-coef-ficient of 0.32 indicates an almost 100-fold higher junction conduc-tance compared to non-junctional membrane conductance (broken
line).
$ 0.32 (32%). Furthermore, a 5-fold decrease or increasein the gj/gm ratio would hardly influence the couplingcoefficient.Experimental results. Under a constant negative currentinjection via a measuring electrode, intracellular rec-ordings of membrane potential were carried out in one
FIGURE 4. (a) Current–voltage relationships of two electrically coupled muscle fibres. Negative membrane potential changes(ordinate left) in the current injected fibre (closed circles) and in the coupled fibre (triangles) after current injection. Further-more, the graph shows that the percentage of voltage shift in the coupled fibre (ordinate right) remains nearly constant afterinjection of different currents (cross-labelled). Inset: Example for a recording after injection of − 20 nA, current injected fibretop. (b) the same as in (a), but simultaneous recordings of three coupled fibres. The coupling-coefficient is almost identicalfor the electrically coupled fibres. Inset: Example for a recording after injection of − 10 nA, current injected fibre top, first
coupled fibre middle, second coupled fibre bottom.
or two other fibres. This generated voltage deflections,the magnitudes of which depended on current intensityand membrane resistance Rinput. The injected currentranged between − 5 nA and − 30 nA. The change inmembrane potential of a neighbouring fibre amounted to32 ± 3% (n = 26) of that in the current injected fibreresulting in a coupling-coefficient of 0.32 (Fig. 4a). Par-allel recordings from 3 muscle fibres showed similarresults (Fig. 4b).
The injection of positive current for several secondseffected an increase in heart beat frequency and, asexpected, the injection of negative current (Fig. 5) causeda decrease in heart-beat rate. In the latter case, the fre-quency determining pacemaker activity was reduced inthe current injected fibre as well as in the electricallycoupled fibre to the same degree (Fig. 5, Table 1). Simul-taneously the required threshold depolarization necessaryfor triggering action potentials was remarkably increasedin all fibres (Fig. 5) although in a non-linear fashion inrelation to membrane hyperpolarization. A weak fluctu-ation of membrane potential in the current injected fibre,did not effect the heart-beat rhythm.
Owing to physiological findings of a myogenicallycontrolled automatism, low ohmic contacts between themuscle fibres are a prerequisite in the antenna–heart andthey somehow should be comparable to the gap junctions(nexus) within intercalated disks in vertebrate heartmuscles. Although we found, neither structures resem-bling intercalated disks nor gap junctions in the antenna–heart of P. americana, striking close membraneapproaches of less than 10 nm could be detected at anintercellular gap of usually 50–300 nm (Fig. 6).

1019CONTRIBUTIONS TO THE ANTENNA–HEART INP. AMERICANA
FIGURE 5. Longer lasting injection of sufficient negative current into a fibre of the antenna–heart, drastically reduces heart-beat frequency. Corresponding to coupling-coefficient, the voltage shift in the coupled fibres is smaller than in the currentinjected fibre but note the negative voltage deflections of the same size in both fibres. The steepness of pre-depolarization is
reduced, threshold depolarization rises.
TABLE 1. Effect of negative current injections into muscle fibres of the antenna–heart on the steepness of pacemaker activity in the currentinjected fibre as well as in the electrically coupled fibres
Steepness of pre-depolarization
Current injected fibre 1. Electrically coupled fibre 2. Electrically coupled fibre
Control (n = 8) 19 ± 1 mV s−1 = 100% 19 ± 1 mV s−1 = 100% 18 ± 1 mV s−1 = 100%− 10 nA (n = 7) 77 ± 11% 73 ± 6% 77 ± 13%− 20 nA (n = 5) 34 ± 7% 35 ± 6% 38 ± 8%
The higher steepness of the rise of the control values in these experiments in comparison to normal value (Fig. 1) is attributed to mechanicalmanipulations by the three floating microelectrodes.
FIGURE 6. (a) Extreme membrane approaches (arrow heads) between two muscle fibres, (b) at higher magnification.

1020 MICHAEL RICHTER AND WIELAND HERTEL
The staining of two adjacent fibres with Cascade blueand Lucifer yellow, respectively, indicated that no dye-stuff exchange occurs between the fibres (compare Hertelet al., in press).
DISCUSSION
Though the antenna–heart of P. americana hasdeveloped independently from the dorsal vessel duringphylogenetic evolution (Pass, 1991; Hertel et al., inpress), there exist numerous similarities regarding thefunction of both hearts. How far this can be generalizedfor other insect species has still to be proven since onlythe physiology of P. americana antenna–hearts has beenstudied to date.
The parameters for action potentials and pacemakeractivity of insect myocard (McCann, 1965; McCann andTadkowski, 1980; Miller, 1969, 1974; Richter, 1971) andantenna–heart musculature correspond to each other inmany respects. Muscle potentials are often graded ininsects, hence a multiterminal innervation is required.However, there are also all-or-none action potentials(Pichon and Ashcroft, 1985). Orchard and Lange (1986)described spike-like action potentials in the oviduct ofLocusta migratoria with which the authors explain thebroad range of contractile activity in this organ. Myogen-ically triggered heart muscle fibres, which includesantenna–heart, generate typical all-or-none action poten-tials. Whereas electrical coupling guarantees the synch-ronization of heart beat, innervation of the antenna–heartis necessary for its control (Hertel et al., 1988, 1997a).Distinct pacemaker fibres or regions could not belocalized in the antenna–heart pointing to a myogenicautomatism of every single fibre. Furthermore, the factthat a completely isolated dilator muscle contracts verywell in vitro and that the electrocardiogram does notshow any oscillations (Hertel et al., 1985), render themyogenic automatism a certainty.
In the present paper, a specific membrane resistanceof Rm = 660 V cm−2 was calculated for the antenna–heart.McCann (1966) discussed a specific resistance of1360 V cm−2 in the myocard of Samia cecropia. Gener-ally, the Rm of vertebrate heart muscle fibres rangesbetween 500 and 2000 V cm−2 (Weidmann, 1952; Jener-ick, 1953; Prosser and Brown, 1962; Sperelakis andLehmkuhl, 1964; Irisawa, 1987), so that the specificmembrane resistance of the antenna–heart is quite com-parable with that of vertebrate myocard muscles.
The lack of intercalated disks or similar structures inthe antenna–heart of P. americana is not surprising since,first of all, intercalated disks are responsible for mechan-ical coupling via desmosomes within the zonula adherensand they are strongly loaded in the direction of tension.Since the muscle fibres of the antenna–heart cover thewhole muscle, such a mechanical coupling between themis not necessary and consequently, these structures arenot required for functioning.
The coordination of a myogenically triggered fibre
contraction can only be effected by sufficient electricalcoupling of the fibres. Such coupling could clearly beproven by current injection in the antenna–heart eventhough we have very little knowledge of its morphologi-cal structure. The close membrane approaches seem tobe possible sites of electrical transmission without gapjunctions since we could not find them. The missing gapjunctions explain the non-existing dye coupling betweenthe muscle fibres of the antenna–heart.
It was interesting, that the results of current injectioninto the transverse dilater revealed a coupling-coefficientof about 0.33 for all fibres. This means that a changein potential of 33% in the electrically coupled fibres incomparison to that in the current injected fibre is suf-ficient to drive the frequency of heart beat. The theoreti-cal calculation by analogy with Socolar (1977) resultedin a coupling-coefficient of 0.32 which almost corre-sponds to the experimental value, confirming the theor-etical cell model. Assuming that control of heart-beat rateis brought about via non-junctional conductance, thereduction of potassium conductance gk+ of 25% causesan increase in frequency of about 50% in case of procto-lin (Hertel et al., 1997a). However, an increase in gk+ atthe same percentage after octopamine application, makesthe heart beat disappear (Hertel, unpublished). In bothcases the coupling-coefficient should not be affected sub-stantially, guaranteeing a good synchronous musclefibre contraction.
The calculated relationship of gj/gm = 100 appearsrealistic since Weingart and Maurer (1987) showed arelationship of nexus resistance to membrane resistanceof uncoupled vertebrate heart cells of about 1:300.Deleze (1987) determined a specific nexus resistance of3 V cm−2 in vertebrates. With the above mentioned spe-cific membrane resistances ranging from 500 to2000 V cm−2, this would result in a relationship of about1:100 and 1:700, respectively.
In summary, the antenna–heart of Periplaneta amer-icana represents a myogenically triggered accessoryheart which is neuronally influenced by the nervus car-dioantennalis. The heart-beat rate is controlled by neuro-mediators via changes in appropriate non-junctionalmembrane conductances of all fibres. This is achieved byreleasing the mediators from synaptoid structures into theintercellular space, thus generating an action on multipledistant fibres (Hertel et al., in press). A small post-synap-tic potential in one or in only a few fibres would notalter the heart-beat frequency. Due to cell topology, thecoupling coefficient is rather insensitive to physiologi-cally relevant changes in ion channel conductance ther-eby preserving the quality of synchronous operation ofthe antenna–heart.
REFERENCES
Deleze J. (1987) Cell-to-cell communication in the heart: structure–function correlations. Experientia 43, 1068–1075.
Hertel W. (1971) Untersuchungen zur neurohormonalen Steuerung des

1021CONTRIBUTIONS TO THE ANTENNA–HEART INP. AMERICANA
Herzens der Amerikanischen Schabe Periplaneta americana (L.).Zool. Jb. Physiol. 76, 152–184..
Hertel W., Pass G. and Penzlin H. (1985) Electrophysiological investi-gation of the antennal heart of Periplaneta americana and its reac-tions to proctolin. lin. J. Insect Physiol. 31, 563–572.
Hertel W., Pass G. and Penzlin H. (1988) The effect of the neuropep-tide proctolin and of octopamine on the antennal heart of Per-iplaneta americana. In Symposia Biologica Hungarica, ed. K.Rosza, Vol. 36, pp 351-361. Akademiai Kiado, Budapest.
Hertel W., Rapus J., Richter M., Eckert, M., Vettermann S. and PenzlinH. (1997a) The proctolinergic control of the antenna–heart in Per-iplaneta americana (L.). Zoology 100, 70–79.
Hertel W., Richter M., Pass G., Ude J., Agricola H. and Penzlin H.(1997b) Morphological aspects of the antenna–heart, an accessorypulsatile organ in Periplaneta americana (L.), (Blattodea:Blattidae). Int. J. Insect Morphol. and Embryol. (in press).
Irisawa H. (1987) Membrane currents in cardiac pacemaker tissue.Experientia 43, 1131–1135.
Jenerick H. P. (1953) Muscle membrane potential, resistance, andexternal potassium chloride. J. Cell. Comp. Physiol. 42, 427–448.
McCann F.V. (1965) Unique properties of the moth myocardium. Ann.New York Acad. Sci. 127, 84–99.
McCann F.V. (1966) The effect of intracellular current pulses on mem-brane potentials in the moth heart. Comp. Biochem. Physiol. 17,599–608.
McCann F. V. and Tadkowski T. M. (1980) Ultrastructure and electri-cal activity in embryonic and adult heart cells of the cricket. Comp.Biochem. Physiol. A 66, 447–454.
Miller T. (1969) Initiation of activity in the cockroach heart. Experien-tia Suppl. 15, 206–218.
Miller, T. A. (1974) Electrophysiology of the insect heart. In ThePhysiology of Insecta, ed. M. Rockstein, Vol. 5, pp. 169-200. Aca-demic Press, New York, London.
Orchard I. and Lange A.B. (1986) Neuromuscular transmission in aninsect visceral muscle. J. Neurobiol. 17, 359–372.
Pass G. (1985) Cross and fine structure of the antennal circulatoryorgan in cockroaches (Blattodea, Insecta). J. Morphol. 185, 255–268.
Pass G. (1991) Antennal circulatory organs in onychophora, myriapodaand hexapoda: Functional morphology and evolutionary impli-cations. Zoomorphology 110, 145–164.
Pichon Y. and Ashcroft F. M. (1985) Nerve and Muscle: electricalActivity. In Comprehensive Insect Physiology, Biochemistry andPharmacology, ed G. A. Kerkut and L. I. Gilbert, Vol. 5, pp 85-113. Pergamon Press, Oxford.
Prosser C. L. and Brown F. A. (1962) Comparative Animal Physiology.Saunders Company, Philadelphia, London.
Richter K. (1971) Zur Wirkung von Neurohormon D auf die Membran-potentiale von Muskelzellen des Myocards von Blaberus craniiferBURM. (Insecta: Blattariae), ein Beitrag zur Frage der Regulationdes Insektenherzens. Zool. Jb. Physiol. 76, 51–63.
Socolar S. J. (1977) The coupling coefficient as an index of junctionalconductance. J. Membrane Biol. 34, 29–37.
Sperelakis N. and Lehmkuhl D. (1964) Effect of current on transmem-brane potentials in cultured chick heart cells. J. Gen. Physiol. 47,895–927.
Weidmann S. (1952) The electrical constants of purkinje fibres. J. Phy-siol. 118, 348–360.
Weingart R. and Maurer P. (1987) Cell-to-cell coupling studied in iso-lated ventricular cell pairs. Experientia 43, 1091–1094.
Acknowledgements—We thank Prof. J. Ude, Dr. H.-J. Agricola, Mrs.A. Schmidt and Mrs. B. von Knorre for their kind help in electronmicroscopy. This work has been supported by Deutsche Forschungsge-meinschaft (He 2434/1-1).





























