Embed Size (px)
Citation preview

Contact with Existing Adipose Tissue Is Inductive
for Adipogenesis in Matrigel
JOHN L. KELLY, M.B., B.Ch., B.A.O.,1,2* MICHAEL W. FINDLAY, M.B.B.S., B.Sc.,1,2*KENNETHR.KNIGHT, Ph.D.,1,2 ANTHONYPENINGTON,M.B.B.S.,1,2 ERIKW.THOMPSON, Ph.D.,1,2
AURORA MESSINA, Ph.D.,1,2 and WAYNE A. MORRISON, M.B.B.S., M.D.1,2
ABSTRACT
The effect of adipose tissue on inductive adipogenesis within Matrigel (BD Biosciences) was assessed byusing a murine chamber model containing a vascular pedicle. Three-chamber configurations that variedin the access to an adipose tissue source were used, including sealed- and open-chamber groups that hadno access and limited access, respectively, to the surrounding adipose tissue, and a sealed-chamber groupin which adipose tissue was placed as an autograft. All groups showed neovascularization, but varied inthe amount of adipogenesis seen in direct relation to their access to preexisting adipose tissue: openchambers showed strong adipogenesis, whereas the sealed chambers had little or no adipose tissue;adipogenesis was restored in the autograft chamber group that contained 2- to 5-mg fat autografts. Theseshowed significantly more adipogenesis than the sealed chambers with no autograft ( p< 0.01). Autograftswith 1mg of fat were capable of producing adipogenesis but did so less consistently than the largerautografts. These findings have important implications for adipose tissue engineering strategies and forunderstanding de novo production of adipose tissue.
INTRODUCTION
ADIPOSE TISSUE GRAFTS are widely used in reconstructive
and cosmetic surgery. For reconstruction of large de-
fects after cancer or trauma, such as breast reconstruction
after mastectomy, larger volumes of adipose tissue are re-
quired than can be provided by simple fat grafting. There
has, therefore, recently been increasing interest in the pos-
sibility of creating new fat tissue by tissue engineering
techniques.1–11
Kawaguchi et al.1 demonstrated that if Matrigel, a base-
ment membrane–rich extracellular matrix derived from a
mouse sarcoma,12 is injected subcutaneously in mice with
the proangiogenic growth factor basic fibroblast growth
factor (bFGF), the gel will be replaced by mature adipose
tissue over 3–10 weeks. If bFGF is not added, adipose tissue
does not form; as a result, the authors suggested a critical
role for angiogenesis in the process of adipose tissue for-
mation. Subsequent studies have suggested that the me-
chanism for the new tissue formation is inductive, with the
surrounding tissues having an important role in providing
stem cells for the de novo adipogenesis.13 We recently de-
scribed an in vivo murine model of tissue engineering for
vascularized adipose tissue.2 This involves a subcutaneously
implanted 5-mm-long, 0.05-mL-volume silicone chamber
that ensleeves the superficial epigastric vessels. This induces
spontaneous angiogenesis and adipogenesis, and the result-
ing product is based on a vascular pedicle.
The development of a human equivalent to Matrigel for
adipose tissue engineering requires a better understanding of
the mechanisms involved in such inductive adipose tissue
engineering, particularly the influence of neovascularization
and the role of the surrounding tissue. To date, the relative
roles of the surrounding tissue and neovascularization in
1Bernard O’Brien Institute of Microsurgery, Melbourne, Australia.2Department of Surgery, University of Melbourne, St. Vincent’s Hospital, Melbourne, Australia.
*These authors contributed equally to the work.
TISSUE ENGINEERINGVolume 12, Number 7, 2006# Mary Ann Liebert, Inc.
2041

inductive adipogenesis remain unclear. Our murine ‘‘cham-
ber model,’’ comprising a perivascular silicone tubing
chamber and modified from the method first described by
Cronin et al.,2 is ideal for investigating these factors because
it provides an isolated environment for such studies. Silicone
was retained as the chamber material for several reasons,
including its proven use in the existing model, its ready
availability, its pliable handling, its minimal tissue reaction,
and its established use in other animal models for purposes
such as nerve entubulation repair.14
METHODS
Chamber insertion and subcutaneous Matrigel plugs
Tissue engineering chambers were inserted bilaterally in a
method modified from Cronin et al.,2 as shown in Fig. 1.
Eight-week-old male C57/Bl6 mice were anesthetized with
4% intraperitoneal chloral hydrate (Merial, Paramatta, Aus-
tralia) and given 5 mg/kg of Carprofen (Pfizer, West Ryde,
Australia) subcutaneously for analgesia. The groin regions
were depilated. A longitudinal incision was made in the
groin, and the superficial epigastric vessels were isolated,
then stripped meticulously of adipose tissue for 7–8 mm and
wrapped in a 5-mm length of 3.35-mm-internal-diameter
silicone tubing (Dow Corning, Midland, MI), which had
been slit along its length. The tubing was allowed to close
around the vessel and was anchored to adjacent muscle
fascia with a 10/0 nylon suture (Davis & Geck, St. Louis,
MO). One end was sealed with bone wax (Ethicon, Somer-
ville, NJ), taking care not to occlude the emerging vessels,
and the chamber was filled with growth factor reduced
Matrigel (BD Biosciences, Bedford, MA) containing 1 mg/
mL of human recombinant bFGF (Sigma-Aldrich, Sydney,
Australia) and 80 U/mL of unfractionated sodium heparin
(APP, Schaumburg, IL). The steps to complete the chamber
insertion were then varied from this point to give 3 different
chamber configurations (Fig. 2) and a no-chamber control.
There was a minimum of 5 animals in each group. In group 1
the second chamber end was also sealed with wax, except
where the vascular pedicle exited the chamber, forming a
completely sealed chamber. In group 2 the chamber was
partially sealed with a wax plug with a 1-mm defect forming
an ‘‘open’’ chamber. This opening abutted the surrounding
epigastric fat pad, providing some contact with adipose tis-
sue. Group 3 was identical to group 1 except that a piece of
autologous adipose tissue from the wound bed (epididymal
fat pad) was inserted into the chamber as a free graft before
sealing, as in group 1 (Fig. 1B). Two different amounts of fat
were inserted: in group 3A, 2–5 mg was used (approximately
4–10% of the chamber volume), and in group 3B, 1 mg was
used (mean weight, 1.03 mg� 0.05 mg [SE], approximately
2% of the chamber volume). In group 4, 300 mL of the same
Matrigel mixture as used in the chamber groups was injected
subcutaneously in the flank of the animal as a no-chamber
control. The skin was closed with 9-mm Auto clips (Becton
Dickinson, Sparks, MD).
The St. Vincent’s Hospital institutional ethics committee
approved all experiments in advance, and animal manage-
ment complied with the Australian Code of Practice for the
care and use of animals for scientific purposes.
Chamber removal and assessment
All animals were anesthetized without recovery at 6
weeks after chamber insertion, and the chambers were ex-
posed surgically (Fig. 1C). Vessel patency within the cham-
bers was assessed macroscopically by looking for bleeding
from the cut ends of the pedicle as it exited either end of the
FIG. 1. (A) Chamber insertion showing depilated right groin with
silicone chamber (short arrow) around superficial inferior epigastric
pedicle (long arrow) before application of bone wax as chamber
sealant. (B) two- to 5-mg adipose tissue autograft (long arrow)
within chamber with Matrigel and the vascular pedicle (short arrow)
running through it. (C) Chamber depicted end-on at harvest, with
new tissue (long arrow) supplied by the vascular pedicle exiting the
chamber (short arrow). (D) Transillumination of a chamber speci-
men showing neovascularization in specimen (arrow). (Color
images available online at www.liebertpub.com/ten.)
FIG. 2. Diagrams of chamber configurations. (A) Sealed. (B)
Open. (C) Sealed with adipose tissue autograft (arrow). 1, wax
plug; 2, superficial inferior epigastric pedicle; 3, silicone chamber.
2042 KELLY ET AL.

chamber. Contents of the chambers and subcutaneous plugs
were removed and weighed, and the volume was determined
by weighing the tissue while suspending it in a 0.9% sodium
chloride solution (normal saline) at room temperature.15
Tissue was routinely fixed in 10% neutral buffered formalin
(Australian Biostain, Traralgon, Australia) and then em-
bedded in paraffin. Serial histologic sections were stained
routinely with hematoxylin and eosin and Masson’s tri-
chrome. The percentages of adipose tissue, residual Matrigel,
fibrous granulation tissue, and vessels were determined by
point-counting every 20th 5-mm section of each specimen by
using digital video imaging ( JVC, TK C1480E) and an au-
tomated, systematic point-counting system (CAST system,
Olympus, Albertslund, Denmark). Only living adipose tis-
sue was counted; any nonviable fat, including graft remnants
where seen, was not included in the count. Nonviable fat was
distinguished from viable fat morphologically because it
displays decreased intensity of staining, loss of nucleus,
breakdown of cell membrane integrity, interstitial mono-
nuclear cell infiltration, and formation of lipid cysts. Lipid
cysts have a typical appearance of a space surrounded by a
cyst wall containing multiple fibroblastoid cells with or
without an inflammatory infiltrate.
It was not possible to distinguish between engrafted and
newly produced fat in all specimens; therefore, percentage
and volume of total adipose tissue were determined and
compared with the volume of adipose tissue at insertion.
Vessels were identified morphologically by their typical
endothelial lining and by the presence of erythrocytes within
their lumen. Results from such vessel counting have corre-
lated well with point-counting of vessels labeled by using
methods, such as lectin staining or India ink perfusion, ap-
plied in previous studies (results not shown). The proportion
of each tissue type was then multiplied by the total volume of
the chamber specimens to determine the volume of each
tissue. Shrinkage from histologic processing was assumed to
be constant across the 4 tissue types.
Statistical analysis
Mean tissue percentages and volumes were compared for
statistical significance by using 1-way analysis of variance
with post hoc analysis, with Dunnett’s test as appropriate
(GraphPad Prism Software, San Diego, CA). Statistical
significance was set at p< 0.05.
RESULTS
Macroscopic appearance and vessel patency
The vessel patency rate of all chambers was over 90%
and independent of chamber type. Those chambers in
which the vascular pedicle had thrombosed were excluded
from the study; however, no tissue grew in these chambers
regardless of experimental group. Comparison of the spe-
cimens from the different groups, using transillumination
and low magnification (�12.5), showed 2 distinct patterns.
Vascularization of Matrigel occurred from the vascular
pedicle and extended to the periphery of the specimen,
spreading out over the surface of the specimen in all cases
(Fig. 1D). Where fat had been left in contact with Matrigel,
the specimens were opaque. Because Matrigel is transpar-
ent, opacity signified tissue/adipogenic replacement.
Histology
The same patterns of neovascularization as seen macro-
scopically were observed microscopically in all chamber
groups, with new vessels branching from the main pedicle
and extending through Matrigel to the periphery of the
specimen (Fig. 3). Most specimens had a fine fibrous cap-
sule containing additional vessels. Fundamental differences
were evident in the relative proportions of Matrigel and
adipose tissue, and the distribution of these, in the different
chamber configurations.
Sealed-chamber group. There was the same new vessel
growth as seen in the other groups, and fibroblastoid cells,
but few adipocytes were evenly distributed within the
Matrigel (Fig. 3A). Any adipocytes seen were usually close
to the vascular pedicle and may be due to residual adipose
tissue left on the vascular pedicle at chamber insertion.
Autograft chamber group. These showed adipocytes
evenly distributed throughout the Matrigel (Fig. 3C and D).
Occasionally, small clusters of lipid cysts with typical
multicellular, dense connective tissue cyst walls, with or
without a surrounding inflammatory infiltrate, were present
at the site of the original autograft. Adipogenesis within the
1-mg autograft chamber group showed 2 patterns. Ap-
proximately 60% of chambers showed good adipogenesis
with the chamber specimens being indistinguishable from
the 2- to 5-mg autograft chamber specimens. The remainder
showed little or no adipogenesis despite the autograft rem-
nants being visible, similar to the sealed-chamber specimens
without an autograft.
Open-chamber group. There was marked adipogenesis
within Matrigel in proximity to the open end of the cham-
ber, and little or no adipogenesis seen at the opposite end of
the chamber, where the chamber was completely sealed
(Fig. 3B).
Matrigel plug group. Subcutaneous injection of 300 mL
of growth factor reduced Matrigel, together with 1 mg/mL
of bFGF (Fig. 4) and heparin, resulted in angiogenic in-
vasion of the plug and almost complete replacement by fat
by 6 weeks, similar to that reported by Kawaguchi et al.1
These specimens formed a no-chamber positive control of
in vivo adipogenesis.
Findings of the point-counting are summarized in Figs. 5
and 6. The chambers seeded with adipose tissue grafts had
ADIPOGENESIS IN MATRIGEL 2043

significantly more adipose tissue than the sealed- or open-
chamber groups (Fig. 5). The difference in the percentage of
adipose tissue between chamber groups was maximal and
most consistent when a 2- to 5-mg autograft was included in
the chamber ( p< 0.01 compared with sealed chamber). The
1-mg autograft group showed greater variability in percen-
tage of adipose tissue, with most specimens having a per-
centage of adipose tissue similar to that of the 2- to 5-mg
autograft group and others having very little adipogenesis.
Because of this variability in response, no statistically sig-
nificant difference in the mean adipose tissue percentage or
volume was detected when we compared the 1-mg autograft
group to the sealed-chamber specimens. Sealed chambers,
with minimal or no access to adipose tissue, contained the
least adipose tissue (<5%). Open chambers with intermediate
access to preexisting adipose tissue developed intermedi-
ate amounts of adipose tissue, but this was not statis-
tically significant when compared with the sealed-chamber
specimens.
Vessels extended from the vascular pedicle to the per-
iphery of the specimens in all groups. Neovascularization
was similar across all chamber configurations, making up
10% of the chamber volume.
Granulation tissue was also measured. This was pre-
dominantly found at entrance and exit points for the pedicle
and around the periphery of the specimen, where it formed a
fine fibrous capsule. The volume of fibrous tissue produced
did not differ between the groups (approximately 20–25% of
the chamber volume in each case).
A significant residual volume of Matrigel was still evident
at 6 weeks, although this was less in the specimens in which
significant adipose tissue had formed ( p< 0.01 when com-
paring the sealed-chamber and 2- to 5-mg autograft groups).
DISCUSSION
The development of an injectable substance for human
use that induces its own replacement by the de novo growth
of adipose tissue would be of great benefit to many patients
across multiple medical specialties. Such inductive adipo-
genesis has the benefit that it does not require the addi-
tion of stem cells harvested from the patient. Matrigel is a
murine prototype of such a substance. When injected sub-
cutaneously in mice, it induces angiogenesis followed by
adipogenesis until its volume is replaced by adipose tissue.1
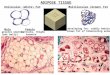
FIG. 3. Histologic characteristics of representative chamber
specimens by chamber group (stained with hematoxylin and eosin).
Section of whole specimen on left, higher-power view of re-
presentative area on right. (A) Sealed-chamber specimen showing
good neovascularization but few adipocytes. (B) Open chamber
with open end to left, where strong adipogenesis is seen. (C) two- to
5-mg autograft chamber specimen showing a strong adipogenic
response throughout the specimen. (D) one-mg autograft chamber
specimen showing a strong adipogenic response throughout.
FIG. 4. Transverse section of subcutaneous Matrigel plug
(�100) showing replacement of Matrigel (light blue) with adipose
tissue (Masson’s trichrome stain). (Color images available online
at www.liebertpub.com/ten.)
2044 KELLY ET AL.

Matrigel is derived from a sarcoma in mice and therefore
cannot be used for tissue engineering in humans.12 It is
hoped that a better understanding of the mechanism by
which adipogenesis occurs within Matrigel will facilitate
the development of an analog suitable for human use.
The mechanism whereby Matrigel induces de novo adi-
pogenesis remains uncertain. Previous research has fo-
cussed on the role of Matrigel when combined with bFGF
in promoting angiogenesis, which, in turn, appears to sup-
port adipogenesis, possibly via recruitment of precursors
from the surrounding tissues.13 The potential role of an-
giogenesis in facilitating adipose tissue production has been
studied in vitro by the co-culture of endothelial cells, adi-
pocytes, and preadipocytes in various combinations,16–18
but to date it has not been possible to isolate each of these
components in vivo.
In a simple mouse model of tissue engineering we have
examined the interaction between existing vessels and tis-
sue in the inductive formation of new adipose tissue within
Matrigel. We have shown that Matrigel with bFGF, when
excluded from an adipose tissue source, will become vas-
cularized but will not develop significant adipose tissue.
When a free graft of adipose tissue is added to the chamber,
or when one end of the chamber is open for access to
Adipose
05
101520253035404550556065
Matrigel Fibrous Vasc/Ped
Tissue Type
Per
cent
age
Tiss
ueby
Poi
nt C
ount
ing
Percentage Tissue by Chamber Type
Sealed
Open
2–5-mg Autograft
1-mg Autograft
FIG. 5. Tissue percentage by chamber type–adipose tissue percentages compared with analysis of variance; the differences were
significant [F(3,22) ¼ 76.9; p¼ 0.003] when compared with the sealed-chamber group. On the post hoc test, only the 2- to 5-mg
autograft group differed significantly from the sealed-chamber group ( p< 0.01).
Adipose Matrigel Fibrous Vasc/Ped
Tissue Type
Vol
ume
Dis
plac
emen
t (µL
)
Tissue Volume by Chamber Type
Sealed
Open
2–5-mg Autograft
1-mg Autograft
0
1
2
3
4
5
6
7
FIG. 6. Tissue volume by chamber type–adipose tissue volumes compared with analysis of variance; the differences were significant
[F(3,22) ¼ 5.9; p¼ 0.0057] when compared with the sealed-chamber group. On the post hoc test, only the 2- to 5-mg autograft group
differed significantly from the sealed-chamber group ( p< 0.01).
ADIPOGENESIS IN MATRIGEL 2045

surrounding fat, new adipose tissue forms within the Ma-
trigel adjacent to this preexisting tissue. The degree of
volume change in the autograft specimens could not be
explained by adipocyte hypertrophy because adipocyte size
in the chamber specimens at harvest was similar to the
graft adipocyte size at insertion.
It is evident in our in vivo model that a suitable matrix, a
patent vascular pedicle, and bFGF are not sufficient by
themselves to generate fat. A tissue source in addition to a
vascular pedicle is essential for the inductive growth of new
adipose tissue. Such definition has not been possible in
subcutaneous models but is afforded by the isolative nature
of our sealed chamber. We found that adipose tissue ap-
peared to grow best in close proximity to an existing or
grafted fat source. Toriyama et al.,13 using the subcutaneous
Matrigel model in which fat abuts the Matrigel plug, ob-
served by electron microscopy that multiple layers of cells at
the periphery differentiate into adipocytes or endothelial
cells, and progressively invade the Matrigel. They observed
that the new fat derives at the point where existing fat con-
tacts the Matrigel, consistent with our observations. The
absence of significant adipogenesis in our sealed-chamber
model supports the assertion that access to surrounding tis-
sues is important for inductive adipogenesis within Matrigel.
No statistically significant differences were detected in
the vascularity of the chamber specimens across the differ-
ent chamber configurations. This finding suggests that an-
giogenesis in Matrigel only requires access to an existing
vasculature or that the influence of existing tissue (e.g., the
autograft) on angiogenesis was negligible. The development
of neovascularization throughout Matrigel within sealed
chambers did not itself induce an adipogenic response. The
few adipocytes seen in the sealed-chamber specimens may
be residual adipocytes left on the vascular pedicle at
chamber insertion. Alternatively, they may represent adi-
pogenesis produced by recruitment of mesenchymal stem
cells via the circulation in the absence of preexisting tissue.
Unlike neovascularization, neoadipogenesis in Matrigel /
bFGF requires more than access to an existing vasculature.
The factor or factors provided by existing fat that promote
neoadipogenesis are not yet determined. Mesenchymal stem
cells have been identified in adipose tissue,19 and thus the
adipose tissue in or near our chambers may be acting as a
source of preadipocytes. Stem cells make up only a very
small percentage of the cells within each tissue,20 and this is
consistent with the variable adipogenic response found with
inclusion of a 1-mg autograft within the chamber when
compared with the more consistent adipogenesis seen with
2- to 5-mg autografts. If stem cells were the major con-
tribution of the preexisting adipose tissue (either host or
grafted) in our system, this would suggest that angiogenesis
in a sealed chamber leads to minimal or no recruitment of
mesenchymal stem cells via the circulation. Also, recent
studies in our laboratory using human adipose tissue xeno-
grafts show that the new adipose tissue is predominantly
murine, suggesting that the graft is inductive of preadipocyte
recruitment rather than providing stem cells directly.21
Other factors, such as the release of extracellular matrix
components,15,16 paracrine factors,17 or chemokines from
the autograft, may also play a role in fostering the neoadi-
pogenesis by stimulating adipogenesis from precursors re-
cruited from the circulation. Experiments designed to isolate
each of these pathways to examine their role in tissue-
induced adipogenesis are in progress.
Increasing evidence suggests close interactions between
angiogenesis and adipogenesis. The precise nature of these
interactions remains unclear. A paracrine-based interaction
between endothelial cells and preadipocytes has been
shown in vivo to involve vascular endothelial growth factor
(VEGF) and VEGF receptor-2 signaling within the en-
dothelial cells.22 Aoki et al.18 showed that co-culture of
endothelial cells and mature adipocytes (in vitro) actively
promotes immature preadipocyte development. These pre-
adipocytes appeared predominantly at the adhesion sites
between the endothelial and mature adipocytes. This effect
was not reproduced by endothelial cell–conditioned media
or by certain endothelial cell–produced cytokines, sug-
gesting that endothelial cells are involved in the enlarge-
ment mechanism of adipose tissue mass through their direct
adhesion to mature adipocytes. Antiangiogenesic agents
have been shown to cause vascular remodeling and reduce
adipose tissue mass in murine models of obesity in a dose-
dependent fashion, confirming a regulation of adipose tis-
sue by its vasculature.23 Hypoxia has been shown to inhibit
adipogenesis via the hypoxia-inducible factor-1–regulated
gene DEC1/Stra13, which causes repression of peroxisome
proliferator-activated receptor-g.24 These findings are con-
sistent with our observation that no fat formed when the
epigastric vessel pedicle was thrombosed, but the absence
of adipogenesis in the sealed chamber, despite considerable
angiogenesis, indicates that angiogenesis is necessary but
not sufficient in the Matrigel/bFGF environment to cause
neoadipogenesis.
The current clinical strategies for the use of adipose
tissue in surgical reconstruction and augmentation rely on
engraftment and revascularization of the adipose tissue
graft, rather than on de novo adipogenesis. Our data in-
dicate an important inductive role of preexisting adipose
tissue, whether vital or isolated, in generating new adipose
tissue with Matrigel with bFGF. Greater understanding of
the mechanism of adipogenesis in response to Matrigel in
mice takes us closer to developing a human equivalent for
human use and has important implications for chamber
design for the production of larger volumes of tissue.
ACKNOWLEDGMENTS
We gratefully acknowledge the surgical assistance from
Sue McKay, Liliana Pepe, Anna Deftereos, and Amanda
Rixon from the Experimental Medical Surgical Unit,
St. Vincent’s Hospital Melbourne. Grant support from the
2046 KELLY ET AL.

National Health and Medical Research Council and Na-
tional Australia Bank is gratefully acknowledged.
REFERENCES
1. Kawaguchi, N., Toriyama, K., Nicodemou-Lena, E., Inou, K.,
Torii, S., and Kitagawa, Y. De novo adipogenesis in mice
at the site of injection of basement membrane and basic fi-
broblast growth factor. Proc Natl Acad Sci U S A 95, 1062,
1998.
2. Cronin, K.J., Messina, A., Knight, K.R., Cooper-White, J.J.,
Stevens, G.W., Penington, A.J., and Morrison, W.A. New
murine model of spontaneous autologous tissue engineering,
combining an arteriovenous pedicle with matrix materials.
Plast Reconstr Surg 113, 260, 2004.
3. Green, H., and Kehinde, O. Formation of normally differ-
entiated subcutaneous fat pads by an established preadipose
cell line. J Cell Physiol 101, 169, 1979.
4. Mandrup, S., Loftus, T.M., MacDougald, O.A., Kuhajda, F.P.,
and Lane, M.D. Obese gene expression at in vivo levels by fat
pads derived from s.c. implanted 3T3-F442A preadipocytes.
Proc Natl Acad Sci U S A 94, 4242, 1997.
5. Patrick, C.W., Chauvin, P.B., Hobley, J., and Reece, G.P.
Preadipocyte seeded PLGA scaffolds for adipose tissue en-
gineering. Tissue Eng 5, 139, 1999.
6. von Heimburg, D., Zachariah, S., Low, A., and Pallua, N.
Influence of different biodegradable carriers on the in vivo
behavior of human adipose precursor cells. Plast Reconstr
Surg 108, 411, 2001.
7. von Heimburg, D., Zachariah, S., Heschel, I., Kuhling, H.,
Schoof, H., Hafemann, B., and Pallua, N. Human pre-
adipocytes seeded on freeze-dried collagen scaffolds in-
vestigated in vitro and in vivo. Biomaterials 22, 429, 2001.
8. Kimura, Y., Ozeki, M., Inamoto, T., and Tabata, Y. Adipose
tissue engineering based on human preadipocytes combined
with gelatin microspheres containing basic fibroblast growth
factor. Biomaterials 24, 2513, 2003.
9. Patrick, C.W., Jr., Zheng, B., Johnston, C., and Reece, G.P.
Long-term implantation of preadipocyte-seeded PLGA scaf-
folds. Tissue Eng 8, 283, 2002.
10. Selye, H., Jean, P., Cantain, M., and Lemire, Y. Induction of
adipose tissue development by mechanical means. Plast Re-
constr Surg 24, 250, 1959.
11. Xaymardan, M., Gibbins, J., and Zoellner, H. Adipogenic
healing in adult mice by implantation of hollow devices in
muscle. Anat Rec 267, 28, 2002.
12. Kleinman, H.K., McGarvey, M.L., Liotta, L.A., Robey, P.G.,
Tryggvason, K., and Martin, G.R. Isolation and character-
ization of type IV procollagen, laminin, and heparan sulfate
proteoglycan from the EHS sarcoma. Biochemistry 21, 6188,
1982.
13. Toriyama, K., Kawaguchi, N., Kitoh, J., Tajima, R., Inou, K.,
Kitagawa, Y., and Torii, S. Endogenous adipocyte precursor
cells for regenerative soft-tissue engineering. Tissue Eng 8,
157, 2002.
14. Odaka, M., Uchiyama, Y., Oka, Y., and Tamaki, T. Muscle
Evaluation of morphological and functional regeneration of
rat nerve-muscle units after temporary and permanent tubu-
lization. Nerve 28, 194, 2003.
15. Weibel, E.R. Stereological Methods, Vol. 1: Practical Meth-
ods for Biological Morphometry. London, Academic Press,
1979.
16. Shillabeer, G., Forden, J.M., and Lau, D.C.W. Induction of
preadipocyte differentiation by mature fat cells in the rat. J Clin
Invest 84, 381, 1989.
17. Varzaneh, F.E., Shillabeer, G., and Wong, K.L., Lau, D.C.W.
Extracellular matrix components secreted by microvascular
endothelial cells stimulate preadipocyte differentiation in vi-
tro. Metabolism 43, 906, 1994.
18. Aoki, S., Toda, S., Sakemi, T., and Sugihara, H. Coculture of
endothelial cells and mature adipocytes actively promotes
immature preadipocyte development in vitro. Cell Struct
Funct 28, 55, 2003.
19. Zuk, P.A., Zhu, M., Ashjian, P., De Ugarte, D.A., Huang, J.I.,
Mizuno, H., Alfonso, Z.C., Fraser, J.K., Benhaim, P., and
Hedrick, M.H. Human adipose tissue is a source of multi-
potent stem cells. Mol Biol Cell 13, 4279, 2002.
20. Asakura, A., and Rudnicki, M.A. Side population cells from
diverse adult tissues are capable of in vitro hematopoietic
differentiation. Exp Hematol 30, 1339, 2002.
21. Stillaert, F., Findlay, M.W., Palmer, J., Cheang, S., Messina,
A., Abberton, K., Morrison, W.A., and Thompson, E.W.
(Manuscript in preparation).
22. Fukumura, D., Ushiyama, A., Duda, D.G., Xu, L., Tam, J.,
Krishna, V., Chatterjee, K., Garkavtsev, I., and Jain, R.K.
Paracrine regulation of angiogenesis and adipocyte differ-
entiation during in vivo adipogenesis. Circ Res 93, e88, 2003.
23. Rupnick, M.A., Panigrahy, D., Zhang, C.Y., Dallabrida, S.M.,
Lowell, B.B., Langer, R., and Folkman, M.J. Adipose tissue
mass can be regulated through the vasculature. Proc Natl
Acad Sci U S A 99, 10730, 2002.
24. Yun, Z., Maecker, H.L., Johnson, R.S., and Giaccia, A.J.
Inhibition of PPAR gamma 2 gene expression by the HIF-1-
regulated gene DEC1/Stra13: a mechanism for regulation of
adipogenesis by hypoxia. Dev Cell 2, 31, 2002.
Address reprint requests to:
Wayne A. Morrison, M.D.
Hugh Devine Professor of Surgery
University of Melbourne Department of Surgery
St. Vincent’s Hospital
Melbourne, Australia 3065
E-mail: [email protected]
ADIPOGENESIS IN MATRIGEL 2047