Embed Size (px)
Citation preview

CONCEPTUAL FRAMEWORKS AND THE PHILOSOPHICAL FOUNDATIONS OF GENERAL LIVING SYSTEMS THEORY’
by Charles E . Warren,’ Marshall Allen,2 and James W. Haefner3
The continued expansion and fragmentation of biological disciplines impede educa- tion, communication, and efficient advance of biological research. Reversing these trends may require a unification of theories and concepts from all levels of biological organization. One form of this unification is the statement of generalizations that apply to all living systems.
We explore the philosophical foundations of general theories of living systems by analyzing conceptual frameworks as they apply to biology. This analysis examines the relation of empirical observation, theories, and conceptual frameworks within the context of an individual scientist’s conceptual continuum. Also presented are a small set of translevel generalizations that articulate our conceptual framework of living systems in terms of organismic system organization, the environmental system-organismic system dyad, system capacity, and system incorporation. A set of procedural rules are stated which suggest minimum criteria for the evaluation of the explanatory adequacy of biological theories. The relation of this work to other similar analyses and syntheses of biological knowledge is discussed.
KEY WORDS: living systems, conceptual framework, translevel, philosophy, generalizations, hierarchy.
M
INTRODUCTION
ILLER (1978) has documented the M rapid proliferation and fragmenta- tion of biological knowledge in recent dec- ades. We share his concern that synthesis and integration are necessary lest the value of current research be lost in a plethora of facts. Miller (1978) has presented one such synthesis in great detail. Working indepen- dently, we have converged on the approach taken by him and other general systems theorists: abstraction across levels of orga- nization to uncover the commonalities of diverse systems.
While Miller (1978) has sought to enu- merate in detail mechanisms and hy- potheses of the structure and function of
’ The authors gratefully acknowledge the Depart- ment of Fisheries and Wildlife, Oregon State Univer- sity, and the Curriculum in Ecology, University of North Carolina, for technical assistance and manu- script preparation. The work of James W. Haefner was supported by National Science Foundation Grants BMS 74-2oooO (to G. S. Innis) and DEB 7805313 during a portion of this research. ’ Department of Fisheries and Wildlife. Oregon
State University, Corvdis. Curriculum in Ecology, University of North Caro-
lina, Chapel Hill.
296
Behavioral Science. Volume 24, 1979
multilevel systems, we attempt here to de- scribe what we feel are the necessary phil- osophical foundations for a general theory of living systems. Thus, our objectives are somewhat different from Miller’s; we desire to delve deeper into the conceptual analysis of many of the concepts fundamental to the analysis of biological systems. We are, therefore, concerned with the concepts of (a) a conceptual framework that describes the general theory of living systems, (b) the environment of a system, (c) the relation of systems and the environments of their sub- systems (incorporation), (d) concordance between systems, subsystems, suprasys- terns, and their respective environments, and (e) system capacity relative to system performance.
We approach these problems in the fol- lowing way. First, we describe the relation between theories, explanations, and con- ceptual frameworks. We advance the thesis that all theory is embedded in a broad and extensive individualized conceptual frame- work which can be articulated and debated. Second, we analyze some fundamental sys- tems concepts by specifying one conceptual framework of general living systems in the form of four translevel generalizations. And

CONCEPTUAL FRAMEWORKS AND LIVING :SYSTEMS 297
third, we suggest that adequate explanation of particular biological systems requires at- tention to a collection of procedural rules that specify the limits of system knowledge and the necessity of translevel syntheses.
UNDERSTANDING AND EXPLANATION
We take the overall goal of the biological sciences to be the understanding of living phenomena, a goal pursued through expla- nations of observational experience. Al- though the nature of explanation continues to be debated in the philosophy of science (e.g., Suppe, 1974), we believe that, most simply and generally, any explanation is a translation of the unfamiliar to the familiar that satisfies those seeking explanation. But familiarity and satisfaction are relative to the conceptual context within which any explanation is proposed and which deter- mines whether or not understanding is achieved. Only recently have some modern philosophers of science emphasized the en- compassing contexts of explanations. To the early logical positivists, most notably Rudolf Carnap, the meaning of an explan- atory proposition was its “method of veri- fication.” This was generally taken to be a deductive argument leading to logical con- sequences that could be related to obser- vational experience, which was thus ex- plained. Internal consistency and predictive power became the dominant criteria of ex- planatory adequacy; little attention was di- rected to conceptual presuppositions and other aspects of “external” adequacy of ex- planations.
Nagel (1961), Popper (1959), and other philosophers recognized that theoretical constructs used in explanation do not in any simple way emerge from observational experience. They are, instead, products of the scientific imagination. But these philos- ophers continued to emphasize internal consistency and verification or falsification by means of observational experience as the principal, if not the only, criterion of explanatory adequacy. Other philosophers, certainly Kant (1781/1958), but more re- cently Hanson (1958), have insisted that the meaning of explanations comes as much from their encompassing contexts of pre- conceptions, beliefs, and background
knowledge of the persons involved as from the internal consistency and observational correspondence criteria emphasized by the logical positivists.
In this view, other important criteria of explanatory adequacy are involved. “Exter- nal adequacy,” which to the positivists had meant primarily predictive power, is seen to entail also the degree of completeness with which an explanation represents total understanding af natural systems. “Heuris- tic power,” the illumination of new prob- lems and approaches to their solution, is another criterion of explanatory adequacy. Finally, “aesthetic appeal,” is a third crite- rion important to modern concepts of ex- planation.
The intellectual and observational activ- ities of an individual biologist can be viewed as a continuum extending from observa- tional experiences (“data”) through gener- alizations of these (“empirical generaliza- tions”) to various attempts at systematic local explanation (“models” and “theories”) and overall conceptualization of a domain (“conceptual framework”), and finally to his richest, most fundamental feelings and beliefs about the living world. Fig. 1 illus- trates this view of an individual’s concep- tual continuum. (Not shown are the rela- tions between a collection of individual con- ceptual frameworks that constitute a sci- entific community.) From observation statements and empirical generalizations to feelings and beliefs, there is a gradient of abstraction, characterized perhaps by a re- ciprocal decrease in verifiability and an in- crease in explanatory breadth. Within the conceptual continuum of an individual sci- entist, we take a conceptual framework to exist at the most encompassing level ame- nable to reasonably complete and adequate verbal articulation. Hence, this is the most inclusive part of the continuum that can be made available to communal scrutiny and use. What we are calling a conceptual framework ought not, then, be construed as a “paradigm” in the senses of Kuhn (1962).
Each individual scientist comes to under- stand his domain of investigation according to his total background, much of which is unique to him, but some of which must be common to all who are trained in a partic-
Behavioral Science, Volume 24, 1979

298 CHARLES E. WARREN, MARSHALL ALLEN, AND JAMES W. HAEFNER
FIG. 1. The conceptual continuum of an individual scientist. Scientific problems are identified by interactions between observations and the theoretical and conceptual context. The conceptual continuum is a hypothetical structure composed of aU aspects of the scientific activity: observation, generalizations, theories, conceptual frameworks, and deeper, more general philosophical beliefs. Falsification and verifiability increase, but explan- atory breadth decreases, as statements pertain more to observations than to conceptual frameworks.
ular science. The conceptual framework of each scientist, then, entails his general view of a natural empirical domain and his judg- ments of what is problematic about this domain and what sorts of explanations are possible and satisfying as to form and con- tent. But science must be communal as well as personal. An effective scientific commu- nity can exist only to the extent that there is overlap of the conceptual continua of scientists investigating a given domain. In such overlap there exists a common view of that domain. This provides a potential for a “communal conceptual framework,” which would entail generally accepted cri- teria of explanatory adequacy, something like those noted above. The work of Miller (1978) is notable in that it includes both empirical generalizations and a statement of Miller’s conceptual framework. We be- lieve further abstraction and generality can be achieved in articulation of conceptual
frameworks for biology other than provided by Miller.
The universality of several of the great theories of physics appears to have pro- vided that science with much explanatory coherency, without there having been artic- ulated what we take to be higher level conceptual frameworks. But in biology there have been no such well-articulated universal theories. Hence, the absence of explicated communal conceptual frame- works has allowed biology to develop with- out much agreement as to the relative im- portance of problems, how they ought to be approached, and what are to be the form and content of adequate explanations. Judgments on these matters have been so personal or limited to such restricted groups of biologists that general coherency of biological explanation is not apparent. Some biologists, in seeking a more adequate biological explanation, have used the theo-
Hehaworal Science, Volume 24. 1979

CONCEPTUAL FRAMEWORKS AND LIVING SYSTEMS 299
retical forms of the physical sciences. In our view these applications of physical expla- nations are often incomplete and do not treat many legitimate biological phenom- ena. This may be due to a misunderstand- ing about the nature and possibilities of explanation and its role in advancing un- derstanding of living phenomena. Conse- quently, there is a need for public debate of the structure of conceptual frameworks for biology, agreement upon appropriate ap- proaches to and criteria for adequate expla- nation, and rigorous tests of the theories and hypotheses entailed by a conceptual framework.
To our knowledge the nature and func- tion of conceptual frameworks (as distinct from paradigms, conceptual continua, and theories) have been considered mainly by Korner (1970), Harre (1964), and Gutting (1973). In his “categorical framework,” Korner describes a framework as the logical scheme by which an individual categorizes the objects of experience. Harre, in his “general conceptual system,” emphasizes framework as a view of the world in which the basic concepts are communally speci- fied by the science of a period. Gutting presents the fullest discussion of the nature and function of communal conceptual frameworks, and our ensuing discussion owes much to his insights concerning the relationships between explanations, indi- vidual conceptual frameworks, and their interrelationships with any existing or po- tential communal conceptual framework.
NATURE OF CONCEPTUAL FRAMEWORKS
The entire domain of biology extends over the various levels of biological organi- zation, each investigated according to the special interests and goals of one or more subdisciplines. Adequate explanation of the integrity of living phenomena would require a coherent and powerful system of thought embodying separate advances of the diverse biological sciences. Because specialized ed- ucation is necessary for these separate ad- vances, the biologist concentrates early on specialized knowledge and receives little preparation for later broadly integrative thinking. Moreover, this training and the existing institutions supporting and con-
ducting research often tend to discourage attempts at synthesis and novel concep- tualization. Were there to be a common conceptual framework adequate for biol- ogy, one that could be taught and used in general form and appropriately interpreted in all domains of biology, these individual and institutional barriers to the develop- ment of more universal biological thought would be reduced.
Such a framework ought to help biolo- gists to perceive and articulate the most fundamental problems in their particular domains of investigation and to see the interrelationships between these problems and their solutions and those in other do- mains. Along with some procedural rules, the framework should make apparent those approaches likely to be most effective in solving problems. Because of its origin from and adequacy for all areas of biology, such a communal conceptual framework not only should strengthen explanatory efforts within each discipline but should also pro- vide common criteria for evaluating the adequacy of proposed research and expla- nations. This could facilitate communica- tion and use of knowledge among diciplines. In short, the roles of conceptual frameworks are to aid (a) in the identification of sci- entific problems, (b) in the origin of theo- ries, (c) in defining relationships between diverse theories, and (d) in evaluating the- ories-which theories themselves, in their greater particularity, generally cannot do.
Individual conceptual frameworks un- doubtedly serve some of these functions, even if they are only vaguely perceived, and a communal conceptual framework should serve most of them. But how are conceptual frameworks to be articulated to serve these functions in the best possible manner? We believe that the basis of a conceptual frame- work should be a general description of living phenomena, interpretable in terms of any particular biological phenomenon or system. As such, it could be articulated as a set of generalizations stating the interre- lationships between the most universal and powerful theoretical concepts of biology.
The fundamental concepts to be em- ployed in the set of law-like generalizations forming the basis of a conceptual frame-
Hrhavaml Science, Volume 24, 1979

300 CHARLES E. WARREN, MARSHALL ALLEN, AND JAMES W. HAEFNER
work need to be of a primitive or theoretical nature, radically different from empirical concepts. Although philosophers may de- bate the meaningfulness of the distinction between “theoretical” and “observational” (Nagel, 1971), our purpose here demands highly abstract concepts that can be differ- ently and yet usefully interpreted for each of the disciplines of biology. And the gen- eralizations articulating these concepts should, at least informally, subsume the theories and empirical generalizations of the different biological disciplines. Theoret- ical concepts, such as “niche” or “selective force,” from population biology, having meaning or applicability to problems in only one or a few areas of biology, cannot serve our purpose. Rather, the theoretical concepts we employ must have an even higher level of abstractness and generality giving them heuristic power within every biological discipline and synthetic power over the knowledge of all disciplines.
In the articulation of a tentative concep- tual framework, we have used natural lan- guage. Because mathematics and other for-
mal languages simplify deductive reasoning and make it clearer and more certain, for- mal theories are desired. But formalization leads to considerable loss of richness of expression and possible meaning, a loss that would reduce the effectiveness of concep- tual frameworks.
BIOLOGICAL CONCEPTUAL FRAMEWORK
We will take a biological system to be composed of an organismic system together with its spacially and temporarily coexten- sive environmental system, which is essen- tial for the persistence of the organismic system (Fig. 2). Such biological systems can be conceived to exist at any level of biolog- ical organization from the molecular level to the level of the biological community. One can think of a biological system within an individual organism as a physiological system, and a biological system including an individual organism as a whole, a species population, or a biological community as an ecological system. In our view, an orga- nismic system at any level of organization
r BIOLOGICAL SYSTEM 3
FIG. 2. The components of a biological (living) system. A dynamic association between environmental systems and organismic systems constitutes biological or living systems. The environmental system has additional structure comprising an effective environment in immediate direct contact with the organismic system, a generative environment that produces effective environmental conditions, and an extraenvironment that has an undescribed relation to the organismic system.
Behavioral Science. Volume 24, 1979

CONCEPTUAL FRAMEWORKS AND LIVING SYSTEMS 30 1
would not be defined simply as a phenom- enon existing in any single, momentary space-time frame, but rather as a contin- uum persisting in time and space from some origin to some future time at which the organismic system and thus its environ- mental system cease to exist (Fig. 2). A coextensive environmental system is best interpreted as being specific to the orga- nismic system under consideration, espe- cially to its level of organization.
In the biological system generalizations that follow, certain other abstract concepts will be identified with theoretical or prim- itive terms, such as “capacity.” As such, these terms will retain their greatest heu- ristic power if left undefined. The meaning of primitive terms is intended to emerge from their context in propositions, or in some uses of scientific deductive systems a posteriori through the translation of theo- rems to empirical generalizations via “rules of correspondence” (Nagel, 1961). That is, these terms can only be defined ostensively by indicating in what particular interpre- tations and deductive systems they are in- volved (see Achinstein, 1968). Thus, we have not defined our primitive terms by stating necessary and sufficient conditions because interpretation is necessary, and this is best left to the reader with his own particular systems of interest. As a result, after presenting the set of generalizations, we shall only interpret or illustrate, not define, these concepts and their interrela- tionships.
I. Organismic system operation, per- formance, and function. Any perform- ance of any organismic system is an out- come of its operation, which consists of the interactive performances of its subsystems, and has functions or plays operational roles in the structure, organization, or rep- lication of the organismic system and a more encompassing system.
11. Organismic system potential ca- pacity, realized capacity, and perform- ance. The potential capacity of any orga- nismic system predetermines all possible sequences of realized capacities, which in turn determine all possible performances, any occurring sequence of realized capac- ities depending on the environmental sys-
tem through time, and any occurringper- formance depending on the immediately effective environment.
111. Organismic system-environmen- tal system relativity. Any performance of any organismic system requires space, time, energy, materials, and information, which are provided in particular forms and limited amounts by its coextensive en- vironmental system; potential and real- ized capacities determine the forms and amounts that will permit performance and thus determine the possible environmental systems within which the organismic sys- tem could persist.
IV. Organismic system incorpora- tion. Any organismic system tends to in- corporate in some degree not only its or- ganismic subsystems but also their partic- ular environmental systems.
In generalization I , we distinguish be- tween the operation, performance, and function of any organismic system or sub- system on any level of organization. Here we take performance of an organismic sys- tem or subsystem to be anything that this system does as a whole as an outcome of its operation. In this view, operation of an organismic system or subsystem as a whole becomes the interactions of all perfor- mances of organismic subsystems on suc- cessively lower levels of organization. Thus, in a sense, performance is what an organ- ismic system or subsystem does as a whole, whereas operation is how this is accom- plished. Function is the operational role or roles a performance of an organismic sys- tem or subsystem plays in the structure, organization, and replication of its subsys- tems or the more encompassing system of which it is a part. Determination of function thus requires appropriate conceptualization of systems on higher and lower levels of organization.
Generalization I1 asserts that from some original potential capacity any organismic system passes through a particular se- quence of realized capacities jointly deter- mined by its potential capacity and the actually prevailing environmental system (Fig. 3). If environmental conditions at any time had been different, a different environ- mental system would have prevailed, and
Behavioral Science, Volume 24, 1979

302 CHARLES E. WARREN, MARSHALL ALLEN, AND JAMES W. HAEFNER
FIG. 3. The capacity of an organismic system. Each organismic system possesses a potential capacity to behave in certain ways. The interaction of system capacity and the state of the environmental system determine the system performance realized at any moment in time.
the sequence of capacities actually realized would also have been different. This is il- lustrated in Fig. 3 by the circles (symboliz- ing organismic systems) inside the environ- mental system. Alternative realized capac- ities occur because the performance of the organismic system at any time is jointly determined by the realized capacity and environmental conditions existing at that time.
We can make this clearer by supposing system organization to be a universal class of performances of organismic systems. Among these classes are system structure and system replication. One can imagine using greater resolution to further specify these performance classes, much as Miller (1978) has done by specifying 19 essential subsystems. In any case, different sets of performances in these classes might be ex- pected to occur at each stage, or even each state, of an organismic system in accord- ance with the realized capacity and envi- ronmental conditions occurring at that time, as we attempt to illustrate in Fig. 3. In some theoretical sense, then, potential
capacity entails all possible sequences of realized capacities and all possible perfor- mances in all possible environmental sys- tems. A simple example of capacity is the human competence to speak and under- stand a large (possibly infinite) number of sentences of a natural language such as English. While this competence or capacity exists a t any one time, the actual sentences spoken or understood (i.e., the realized sen- tences) will depend on the environment of the speaker. We believe this characteristic to be common to all organismic/environ- mental system dyads. Thus, any perform- ance of any organismic system can be ex- plained as a consequence of its capacity and its environmental system.
Generalization I11 further specifies the interrelationships among an organismic system, its potential and realized capacities, its performances, and its environmental system. First, this generalization asserts that any performance of any organismic system requires from a coextensive environ- mental system space, time, energy, materi- als, and information in particular forms and
Behavioral Science, Volume 24, 1979

CONCEPTUAL FRAMEWORKS AND LIVING SYSTEMS 303
amounts. Second, generalization I11 asserts that the potential and realized capacities of the organismic system determine the forms and amounts of environmental require- ments that wdl permit its performance. This determines the possible environmen- tal systems within which the organismic system could possibly persist.
We cannot conceive of any natural or- ganismic system persisting apart from an environmental system, as though in a void. Neither can we conceive of an environmen- tal system except in relation to some orga- nismic system (Fig. 2). This necessary rel- ativity of organismic and environmental systems is extended to the components of an environmental system by potential and realized capacities determining their nec- essary forms and amounts and is main- tained only so long as their provision by the environmental system is adequate for es- sential organismic system performance.
Much inadequacy of physiological, ge- netic, and ecological explanations has fol- lowed from failure to ded effectively with the notion of environment. It may be help- ful to think of the coextensive enuironmen- tal system as having two phases: an effective environment composed of conditions im- pinging immediately and directly on the organismic system at any time during its persistence; and a generative environment composed of sequences of conditions lead- ing to those in the effective environment (Fig. 2). It is not helpful in explanation to allow infinite regress of a defined environ- mental system, and one may choose to in- clude in the generative environment only those parts of sequences of environmental conditions necessary to the explanation being sought. Definition of an organismic system of interest together with its environ- mental system composed of effective and generative phases would define a biological system. But no natural system, biological or other, exists in a void, and perhaps we should think of the biological system of interest as existing in a coextensive ex- traenvironment composed of conditions that for conceptual or methodological rea- sons have been excluded from the environ- mental system but that may ultimately be important to the persistence of the or- ganismic system.
Generalization IV asserts that any or- ganismic system tends to incorporate in some degree not only its organismic subsys- tems but also their particular environmen- tal systems. By incorporate we mean to integrate or combine together so as to form one harmonious or consistent whole. The notion of any system incorporating its sub- systems is self-evident. Not so evident is the notion of an organismic system tending to incorporate the environments of its sub- systems, the implication being that in some important sense the environment of a sub- system is not at all the same as the envi- ronment of the encompassing system. Yet, in our opinion, failure to make and use this distinction has seriously weakened biologi- cal explanation and raised the problem of wholes and parts (Bertalanffy, 1933).
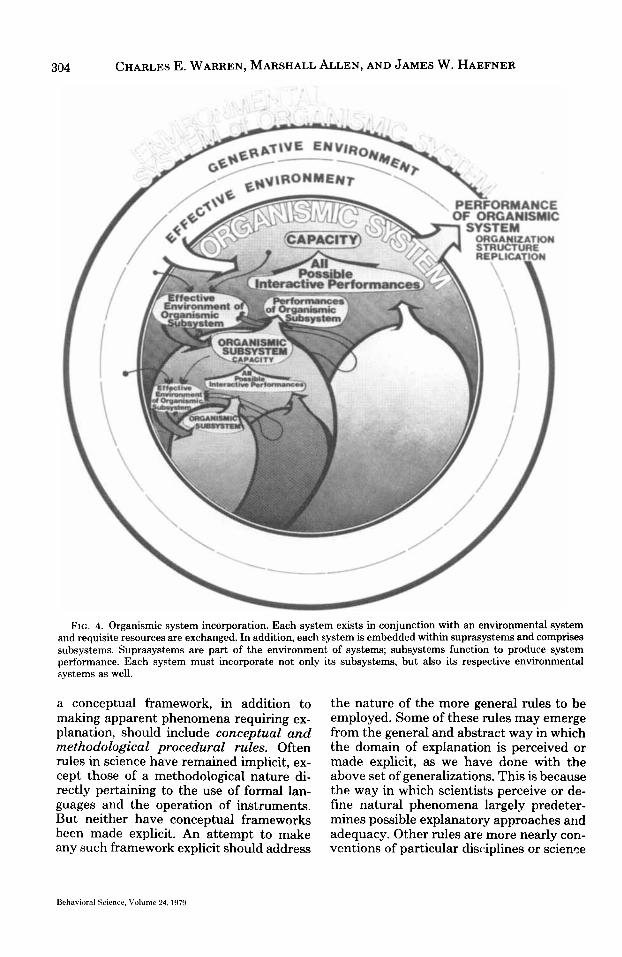
Fig. 4 illustrates, among other matters, the incorporation of subsystem environ- ments. Into the environment of any organ- isimic subsystem comes energy, materials, and information from other subsystems and, in some sense, from the organismic system as a whole, as well as directly from the environment of the organismic system (Fig. 4). The performance of a subsystem is thus determined not only by its potential and realized capacities but by inputs- t,hrough its level-specific environmental system-from other subsystems, the sys- tem as a whole, and perhaps directly from the environment of the encompassing sys- tem. This view is sufficient and perhaps necessary to explain the integration of or- ganismic systems through space and time.
Fig. 4 also provides one view of the pos- sible entailment of potential and realized capacities. The potential and realized ca- pacities of any organismic system entail all possible interactive performances of its sub- systems in all possible environments and thus determine all possible operations and performances of the organismic system as a whole. Organization of organismic sys- tems through time may lead to increasing incorporation of subsystems and their en- vironments.
SOME PROCEDURAL RULES
To suggest approaches to the explanation of natural phenomena and provide more explicitly for the evaluation of explanations,
Behavioral Science, Volume 24, 1979
304 CHARLES E. WARREN, MARSHALL ALLEN, AND JAMES W. HAEFNER
FIG. 4. Organismic system incorporation. Each system exists in conjunction with an environmental system and requisite resources are exchanged. In addition, each system is embedded within suprasystems and comprises subsystems. Suprasystems are part of the environment of systems; subsystems function to produce system performance. Each system must incorporate not only its subsystems, but also its respective environmental systems as well.
a conceptual framework, in addition to making apparent phenomena requiring ex- planation, should include conceptual and methodological procedural rules. Often rules in science have remained implicit, ex- cept those of a methodological nature di- rectly pertaining to the use of formal lan- guages and the operation of instruments. But neither have conceptual frameworks been made explicit. An attempt to make any such framework explicit should address
the nature of the more general rules to be employed. Some of these rules may emerge from the general and abstract way in which the domain of explanation is perceived or made explicit, as we have done with the above set of generalizations. This is because the way in which scientists perceive or de- fine natural phenomena largely predeter- mines possible explanatory approaches and adequacy. Other rules are more nearly con- ventions of particular disciplines or science
Behavioral Science, Volume 24, 1979

CONCEPTUAL FRAMEWORKS AND LIVING SYSTEMS 305
in general, and some appear to have both definitional and conventional origins.
Perhaps many, if not most, biologists will find the generalizations more acceptable than the rules we have stated, even those rules emerging quite directly from the gen- eralizations. This may especially be SO for rules 3 and 4. But disagreement is an inev- itable consequence of making personal con- ceptual frameworks explicit. Moreover, we appreciate that rules can be broken, but recognize that the explanatory utility of breaking rules ought to be, in the balance, greater than any resulting explanatory in- adequacy. The following set of conceptual and methodological rules is not intended to be complete.
Rule 1. Only the performance of an or- ganismic system or subsystem can be mea- sured, its capacity and its operation being representable only indirectly and incom- pletely.
Rule 2. Measurements of the perform- ance of an organismic system or subsystem without relevant measurements on its coextensive, level-specific environmental system are of little explanatory value.
Rule 3. Operational explanation of an organismic system should take into ac- count the performances and operations of subsystems on successively lower levels of organization and cannot be based only on knowledge of subsystems on the lowest levels of organization.
Rule 4 . Explanatory generalizations pertaining directly to any one level of or- ganization of an organismic system should contain at least one concept specific to that level and should subsume conceptual, methodological, or other sorts of indeter- minacy that may exist with respect to lower levels of organization.
Rule 5. Perception and explanation of organismic and environmental systems are always partial relative to the space and time dimensions and the components of the systems.
So long as generalizations I and 11, the distinctions between capacity, operation, and performance there asserted, and the ways in which we illustrate these concepts (Figs. 3 and 4) are accepted, rule 1 seems to be evident. We may be able to measure the
performance, output, or change of any or- ganismic system or subsystem on any level of organization. But if operation is those interactive performances of subsystem on lower levels of organization that lead to a given performance, we cannot reasonably expect to measure all of these lower level performances, much less their interactions. Knowledge of interactions between objects is an interpretation of measurements made on the performances of these objects. In- deed, such complex causal webs are gener- ally explained by defining restricted sys- tems that appear to relate the few lower level performances we do measure to the higher level performance we would explain in operational terms. Capacity, as we em- ploy it, is a strictly theoreticd concept, in the sense that it can never be directly or completely evaluated. At most, we can mea- sure it only as a set of performances, each determined under a different set of environ- mental conditions. And we can never hope to measure all possible performances under all possible sets of environmental condi- tions. (This rule ultimately raises the ques- tion of the nature of causality and we ally ourselves with the Humean-Kantian view that it is a precondition for knowledge.)
Rule 2 in part also follows from general- izations I and 11. If performance is deter- mined not only by capacity but also by environmental conditions and can be ex- pected to change with the latter, we cannot explain performance except in relation to information on environmental conditions. Rule 2 also asserts that the environmental conditions of the organismic system or sub- system whose performance we would ex- plain are specific for that system, especially its level of organization. This follows from generalizations 111 and IV, as we interpret these. (See Bonner, 1969, and Waddington, 1957, for a similar discussion.)
Rule 3 will not be acceptable to many of those working in areas of biology in which the common practice has been to propose operational explanations of high-level sys- tem performance on the basis of informa- tion about subsystems on much lower levels of organization and without knowledge of the performances of intermediate-level subsystems. To the extent that generaliza-
Behavioral Science, Volume 24. IS79

306 CHARLES E. WARREN, MARSHALL ALLEN, AND JAMES W. HAEFNER
tions I, 11, 111, and IV provide an adequate theoretical description of any organismic system whatsoever in its relationships to its environmental system, we see no way of avoiding the assertion of rule 3 that reason- ably adequate operational explanations of the performance of organismic systems can- not be based only on knowledge of subsys- tems at the lowest levels of organization. Thus, disagreement with rule 3 ought to be based on some inadequacy found in the theoretical description provided by these four generalizations.
In no way does rule 3 assert that knowl- edge of organismic subsystems, however low their levels of organization, is without value; it only places vague boundaries on application of such knowledge for explana- tory purposes. (These concerns are raised by Nagel, 1961, and Simon, 1971, in dis- cussing the autonomy of biology as a sci- ence.)
The second part of rule 4, which asserts that explanatory generalizations pertaining to any one level of organization must sub- sume whatever kinds of indeterminacy that may occur at lower levels, is so much a part of present philosophy and practice of sci- ence as to require no justification here. But this is not so with regard to the first part of rule 4, which asserts that generalizations pertaining directly to the explanation of any one level of organization should contain at least one concept specific to that level. This rule implicitly questions the possibil- ity and perhaps the explanatory utility of the complete reduction of one area of sci- ence to another, say, biology to physics, or organismic biology to molecular biology. Of course, the possibility of successful reduc- tion must remain an open question. Nev- ertheless, we are not aware of anything in the philosophy or the practice of science that offers much promise of successful ex- planations that avoid level-specific con- cepts. (Similar views are expressed by Lev- ins, 1966, and Lewontin, 1974.)
Rule 5 should not be controversial. Its statement seems important only because enthusiasm for a very general explanation can on occasion lead to neglect of the limits of its domain of explanatory utility. Percep- tion and explanation of any natural or- ganismic and environmental system must
always be incomplete. (Strongest consider- ation of this in biology has been by von Uexkull, 1909, and in philosophy by Kant, 178 1/ 1958.)
FRAMEWORK ELEMENTS IN BIOLOGY
Although the way in which we have ar- ticulated this conceptual framework may be original, much, if not most, of its content has been recognized by others. Certainly most scientists acknowledge the utility of theoretical concepts and abstract general- izations and some have recognized the need for articulated frameworks such as this one. Indeed, the history of biological science in this century can be viewed to have been as much an attempt to articulate a general theoretical biology as it has been the de- velopment of new empirical subdisciplines (Allen, 1975). Early attempts to erect a general theoretical biology, although not widely acceptable, raised some concepts to theoretical status and stimulated discussion at the conceptual framework level, if not at the level of formal theory (Fig. 1).
The biological system concept of Berta- lanffy (1933) is indicative of relatively early efforts to establish concepts interpretable at all levels of biological organization. Ber- talanffy (1933), Woodger (1930a, 1930b, 1931), Russell (1945) and others strove for frameworks within which theoretical con- cepts could be systematically interpreted. Important in these frameworks was the no- tion of “levels of biological organization” integrated through their hierarchical ar- rangement. This organizational view of the various domains of biological science plus the theory of evolution by natural selection provides much of the integration of biolog- ical thought that exists today (MacMahon, Phillips, Robinson, & Schimpf, 1978). Be- yond the advancing of theoretical concepts and interpretive views, there were efforts to achieve law-like generalizations that could be interpreted in terms of phenomena at aLl levels of biological organization. This effort is clear in the work of Bertalanffy and Woodger, even though their generalizations are widely scattered in their writings. Spe- cific listings or collations of generalizations have appeared more recently (Feibleman, 1954; Kostler, 1967; Weiss, 1969; Miller, 1978). Yet the need for a general conceptual
BrhavliJral Science, Volume 24, 1979

framework for biology, noted in the conclu- sion of a symposium by such eminent biol- ogists as Simpson, Mayr, Wright, Weiss, and Gerard (Gerard, 1958) has not attracted much attention. Nevertheless, we believe that both the intent and the content of the conceptual framework here tentatively pro- posed have firm roots in modern biology.
Our concept of “organismic system” de- scends in both idea and spirit from the discussion of organismal biologists of the early part of this century. This concept was also applied to levels of organization other than the individual organism, as did Clem- ents (1916) in his concept of a plant com- munity as an organism. But, too often, ap- plication of this concept has suffered from general misunderstanding of proper use of analogy and metaphor in biological expla- nation (Wheeler, 1911; Emerson, 1939; Clements, 1916). Our notion that an “envi- ronmental system” is associated with each organismic system is evident in the thinking of some organismal biologists (Haldane, 1913; von Uexkull, 1909). But the concept of environment (one of the most important in biological explanation) is subtle and re- quires careful analysis. This has been at- tempted by Mason and Langenheim (1957) and Fry (1947), but the theoretical use of this concept has generally been disappoint- ing.
The concept of capacity that we use is very close in idea and explanatory intent to Aristotle’s “potentia.” This notion has been extensively employed in attempts to under- stand the generation and development of organisms. A notion resembling our concept of “potential capacity” is apparent in the writings of Waddington (1957,1975), whose “canalized development” represents a col- lection of potential organismal develop- mental trajectories as well as those actually realized in any segment of time. The real- ized developmental trajectories are deter- mined in part by a coextensive environ- ment. Chomsky (1957) emphasized the fun- damental distinction between language competence and language performance.
Finally, our concept of “incorporation” (that an organismic system incorporates not only its subsystems but also their level- specific coextensive environments) appears in the writings of Weiss (1969) and Miller
(1978). In his “canon of determinancy,” Weiss views an organism as a system “en- compassing and operating through the agency of subsystems, each of which in turn, contains and operates through groups of systems of still lower order . . . ” (Weiss, 1969, p. 33). Although the notion of encom- passing and encompassed systems is rec- ognized in general systems theory (Mesa- rovid, Macko, & Takahara, 1970), formula- tions of this theory in our opinion neither entail nor explain the capacity of biological systems and subsystems. Indeed, the ex- planatory power of the theoretical concept of capacity has not been generally recog- nized.
Even though the conceptual and meth- odological rules that we have advanced have historical antecedents and contempo- rary advocates, there has been little recog- nition of the need for rules setting the con- ditions more or less necessary for the expla- nation and understanding of biological phe- nomena. For the most part, rules have been the concern of philosophers of science, in- dividual scientists generally proceeding on the basis of their own intuition and the common practice in their disciplines. There is ample evidence that this has generally been quite effective and that some rules are operative, although usually at a subliminal level. But unified explanations crossing bi- ological domains and levels of organization may not be achieved by inconsistent and only partial use of whatever may be the minimal set of rules essential for adequate biological explanation. But such a set of rules need not be arbitrary. The general nature of all biological phenomena, as un- derstood in any period, should be definable by some set of abstract generalizations. And any such definition should indicate the min- imal conditions or rules for further expli- cation of biological phenomena.
Because of the similarity of this work to that of Miller (1978), some discussion of the relation of the two conceptualizations is needed. It is hoped that an improved view of living systems will emerge as a result of this comparison. One of the most striking differences is that we have not attempted to enumerate the necessary subsystems for any evolutionarily persistent system. Miller (1978) has identified 19 of these subsys-
CONCEPTUAL FRAMEWORKS AND LIVING SYSTEMS 307
Hehaworal SuenLe. Volume 24, 1979

308 CHARLES E. WARREN, MARSHALL ALLEN, AND JAMES W. HAEFNER
tems, and a consequence is that objects that do not possess the subsystems will not be systems. Thus, a single amoeba may be a system, but a population of amoebae is not, because at the population level (in nonsocial, noncolonial species) many of the essential subsystems are not definable. In the present formulation, however, a popu- lation may be considered an organismic system since it is possible to apply the four generalizations at the population level. We believe many ecologists will feel uncomfort- able with a conceptualization that does not permit populations, species, or ecosystems to be systems.
On the other hand, there are many simi- larities between these two approaches. While Miller (1978) presents a much more detailed and empirically testable structure, both studies have common concepts. For example, both studies have identified simi- lar definitions and roles for the concept of system performance or process (in the sense of the word employed by Miller, 1978). In addition, both studies have emphasized the importance of identifying the uses different systems have for requisite resources (time, space, matter, energy, and information) in achieving a particular process or perform- ance. Finally, Miller has alluded to the con- cept of incorporation which we have explic- itly emphasized (generalization IV) when he wrote: “A mutality exists among the components of a system. Each makes its contribution toward the processes of the whole and receives in return a portion of the benefits derived from those activities. This mutual interrelationship extends across the levels” (Miller, 1978, p. 1025).
Both the similarities and dissimilarities between these two approaches to general theoretical biology are encouraging and stimulating. That they have converged from independent sources indicates that substantive issues are being addressed. Cer- tainly it substantiates the fact that a need for analyses of this sort exists.
SOME POSSIBLE DEVELOPMENTS
The foregoing may seem to be little more than the efforts of three biologists dabbling in the philosophy of science. Moreover, some readers may not be comfortable with
our apparent unconcern for the academic separation of biology and philosophy. But isolation of biology from philosophy may prevent attainment of more universal bio- logical explanation and understanding. Such a goal requires not only doing more theoretical and empirical work but also making judgments on how best to proceed in this. And judgments of this sort are of a philosophical nature. Still, for all the con- tributions of philosophy to science and of science to philosophy, the relationship be- tween the philosopher and the scientist has, with a few outstanding exceptions, re- mained an uneasy one. To the extent that there persists mutual lack of appreciation and knowledge of problems and approaches among philosophers and scientists, the ad- vancement of scientific understanding may be impeded.
Biological education prepares biologists in the special knowledge of their disciplines, but a t the expense of adequate education in integrative perspectives and procedures. Philosophers well prepared in the latter are not usually prepared to work with the de- tails of biological knowledge. Although, from Aristotle to the present, some philos- ophers have considered biology, the philos- ophy of science could more precisely be termed the philosophy of physics, to which it more nearly applies and from which it mainly developed. The philosophy of sci- ence is mostly about physics, its patterns of explanation, great theories, and the infel- lectual processes of its most eminent prac- titioners, whose own writings have often addressed important philosophical prob- lems. But how is it that philosophers, not trained in the detailed knowledge of physics any more than in that of biology, have managed to deal more adequately with physics than with biology?
We wonder if it is because the great the- ories of physics, a t least since Galileo, have provided philsophers with well-articulated and high-level summaries of knowledge of the physical world in a way that no extant biological theory has done or can do. These physical theories have made it possible for philosophers to grasp the essence of physi- cal explanation and explore its form and processes, without needing to master all the
Behavioral Science. Volume 24. 1979

CONCEPTUAL FRAMEWORKS AND LIVING SYSTEMS 309
details of physical knowledge. Biological knowledge, on the other hand, is a knowl- edge of details; it offers little having the nature of adequate comprehensive sum- maries, articulated in a way that philoso- phers can appreciate. We do not see that there will soon be any such formal biologi- cal theories: It is yet to be shown that the biological domain is amenable to adequate explanations having the form of physical theory.
Conceivably, until adequate biological theory in one form or another is available, conceptual frameworks articulated as sys- tems of abstract generalizations somewhat as we have done could provide philosophers with the sort of summaries of biological knowledge they would need to discuss bi- ology. Then biology could benefit more from knowledgeable philosophical analysis. But the initial tasks of articulating concep- tual frameworks would need to be accom- plished by biologists. The outcome could well be that biologists and philosophers to- gether would conclude that neither the phi- losophy nor explanatory patterns of biology can be just the same as those of physics. In any case, an integration of biological thought and practice and a recognition of criteria of explanatory adequacy resulting from efforts to develop conceptual frame- works for biology would justify such efforts by biologists.
REFERENCES
Achinstein, P. Concepts of science: A philosophical analysis. Baltimore: The Johns Hopkins Press, 1968.
Allen, M. W. J. H. Woodger and the emergence of supra-empirical orders of discussion in early 20th century biology. Master’s thesis, Oregon State Univ. Corvallis, 1975.
Bertalanffy, L. von. Modern theories of development. Oxford: Oxford Univ. Press, 1933.
Bonner, J . T. Analogies in biology. In J. R. Gregg and F. T. C. Harris (Eds.), Form and strategy in science. Dordrecht: D. Reidel, 1969, pp. 251- 255.
Chomsky, N. Syntactic structures. The Hague: Mou- ton, 1957.
Clements, F. Plant succession. Washington, D. C.: Carnegie Institute of Washington, (Publ. No.
Emerson, A. E. Social coordination and the superor- ganism. American Midland Naturalist, 1939,
242, 1-512) 1916.
21, 182-206.
Feibleman, J. K. Theory of integrative levels. British Journal of Philosophy and Science, 1954, 5, 59-66.
Fry, F. E. J. Effects of the environment on animal activity. University of Toronto Studies, Biol. Ser. 55, Ontario Fisheries Research Labora- tory Publication, 1947, 68, 1-62.
Gerard, R. (Ed.). Concepts of biology. National Re- search Council Publications, 1958, 560, 1-215.
Gutting, G. Conceptual structures and scientific change. Studies in History and the Philosophy of Science, 1973,4,209-230.
Haldane, J. B. S. Organism and environment. New York: MacMillan, 1913.
Hanson, N. R. Patterns of discovery. Cambridge: Cambridge Univ. Press, 1958.
Harre, R. Matter and method. New York: St. Martins Press, 1964.
Kant, I. [Critique of pure reason] (N. K. Smith, trans.). London: MacMillan, 1958. (Originally published, 1781.)
Koestler, A. The ghost in the machine. New York MacMillan, 1967.
Korner, S. Categorial frameworks. New York Barnes and Noble, 1970.
Kuhn, T . The structure of scientific revolutions. Chi- cago: Univ. of Chicago Press, 1962.
Levins, R. Evolution in changing environments. Princeton: Princeton Univ. Press, 1966.
Lewontin, R. The genetic basis of evolutionary change. Syracuse: Syracuse Univ. Press, 1974.
MacMahon, J . A., Phillips, D. L., Robinson, J. V., 8z Schimpf, D. J. Levels of biological organization: An organism-centered approach. Bioscience,
Mason, H., & Langenheim, J . Language analysis and the concept environment. Ecology, 1957, 38, 325-340.
Mesarovid, M. D., Macko, D., Takahara, Y. Theory of hierarchical multilevel systems. New York Ac- ademic Press, 1970.
Miller, J . G. Living systems. New York: McGraw-Hill, 1978.
Nagel, E. The structure of science. New York Har- court, Brace and World, 1961.
Nagel, E. Theory and observation. In E. Nagel, S. Bromberger, & A. Grunbaum, Observation and theory in science. Baltimore: The Johns Hop- kins Press, 1971, pp. 15-45.
Popper, K. The logic of scientific discovery. New York Basic Books, 1959.
Russell, E. S. The directedness of organic activities. Cambridge: Cambridge Univ. Press, 1945.
Simon, M. The matter oflife. New Haven, Conn.: Yale Univ. Press, 1971.
Suppe, F. (Ed.). The structure of scientific theories. Urbana, Ilk Univ. Illinois Press, 1974.
Uexkull, J. J . von. Umwelt und Znnenwelt. Berlin: Springer, 1909.
Waddington, C. H. The strategy of the gene. London: George Allen and Unwin, 1957.
Waddington, C. H. The evolution of an evolutionist. Ithaca, N.Y.: Cornell Univ. Press, 1975.
Weiss, P. The living system: Determinism stratified
1978, 28, 700-704.
Behavioral Science, Volume 24, 1979

310 CHARLES E. WARREN, MARSHALL ALLEN, AND JAMES W. HAEFNER
In A. Koestler & J. R. Smythies (Eds.), Beyond reductionism. Boston: Beacon Press, 1969, pp. 3-55.
Wheeler, W. M. The ant colony as an organism. Jour- nal ofMorphology, 1911, 22, 307-325.
Woodger, J. H. The concept of organism and the relation between embryology and genetics. Part I. Quarterly Review of Biology, 1930, 5, 1-22. (4
Woodger, J. H. The concept of organism and the relation between embryology and genetics. Part 11. QuurterZy Review of Biology, 1930, 5, 438- 463. ( 6 )
Woodger, J. H. The concept of organism and the relation between embryology and genetics. Part 111. Quarterly Review of Biology, 1931, 6, 178- 207.
Behavioral Science. Volume 24. 1979





























