Embed Size (px)
Citation preview

Journal of Clinical Neuroscience 21 (2014) 893–898
Contents lists available at ScienceDirect
Journal of Clinical Neuroscience
journal homepage: www.elsevier .com/ locate/ jocn
Review
Complexities of lysophospholipid signalling in glioblastoma
http://dx.doi.org/10.1016/j.jocn.2014.02.0130967-5868/� 2014 Elsevier Ltd. All rights reserved.
⇑ Corresponding author. Tel.: +61 3 9035 8586E-mail address: [email protected] (A. Morokoff).
Wayne Ng a,b,c,f, Alice Pébay a,d, Katharine Drummond a,b,c,f, Antony Burgess a,e, Andrew H. Kaye a,b,c,Andrew Morokoff a,b,c,⇑a University of Melbourne, Parkville, VIC, Australiab Department of Surgery, Royal Melbourne Hospital, Grattan Street, Parkville, VIC 3050, Australiac Department of Neurosurgery, Royal Melbourne Hospital, Parkville, VIC, Australiad Centre for Eye Research Australia, Royal Victorian Eye and Ear Hospital, East Melbourne, VIC, Australiae Walter and Eliza Hall Institute of Medical Research, Parkville, VIC, Australiaf Melbourne Brain Centre at Royal Melbourne Hospital, Parkville, VIC, Australia
a r t i c l e i n f o
Article history:Received 12 January 2014Accepted 22 February 2014
Keywords:AutotaxinGlioblastoma multiformeLysophosphatidic acid
a b s t r a c t
Glioblastoma multiforme (GBM) is the most malignant brain tumour and continues to have a very poormedian survival of 12–16 months despite current best therapies. These aggressive tumours always recurafter treatment and are defined by their ability to diffusely infiltrate and invade normal brainparenchyma. Autotaxin is overexpressed in GBM, and is a potent chemotactic enzyme that produceslysophosphatidic acid. Lysophospholipid (LPL) signalling is known to increase invasion of solid tumoursand is also dysregulated in GBM. The LPL pathway has been shown to interact with known cancer-relatedsignalling pathways, including those for epidermal growth factor and yes-associated protein, which arealso dysregulated in GBM. The interactions between these pathways provide insights into the complex-ities of cancer signalling and suggest potential novel targets for GBM.
� 2014 Elsevier Ltd. All rights reserved.
1. Introduction
Glioblastoma multiforme (GBM) is the most malignant (WorldHealth Organization grade IV) glioma and continues to have a verypoor median survival of 12–16 months despite current besttherapies (maximal safe resection with concurrent temozolomidechemotherapy and radiation therapy) [1]. These aggressive tu-mours always recur after treatment and are defined by their abilityto diffusely infiltrate and invade normal brain parenchyma. Thusthe search for targeted agents inhibiting cell proliferation, survivaland invasion has intensified. Research into the epidermal growthfactor receptor (EGFR) pathway has led to clinical trials of EGFRand phosphatidylinositol-3-kinase (PI3K) inhibitors that modulatecell survival and proliferation in pre-clinical models. However,results from early EGFR inhibitor trials have not delivered on theirpromise and PI3K inhibitor trials are ongoing [2–4]. These trialshave made it obvious that strategies combining therapies againstmultiple targets are required to account for the existence ofcomplex pathway interactions and redundancies. For example,matrix metalloproteases (MMP) degrade extracellular matrix com-ponents to produce more favourable conditions for cell migration
and invasion [5]. However, they also cleave and activate growthfactors such as epidermal growth factor (EGF) [6]. The function ofintegrins and their influence on cell morphology and migrationhas also been enlightening [7,8]. More recently, other promisingfactors have been identified, including autocrine motility factorreceptor, heparin-binding epidermal growth factor, ephrin-B3,netrin 4 and autotaxin (ATX) [9]. The latter three are of interestbecause their role in cell migration and motility in neural stemcells suggests a similar role in glioma-derived cancer stem cells,that have a putative role in GBM progression [10,11]. ATX in partic-ular is a powerful chemotactic enzyme involved in lysophospho-lipid (LPL) signalling, and its recent prominence in the literaturehas highlighted the importance of lipid signalling within complexintracellular pathway interactions. This review focuses on the rolethat LPL signalling may play in gliomagenesis and its potential as atarget in the treatment of this highly malignant disease.
2. Lysophosphatidic acid
Lysophosphatidic acid (LPA) and sphingosine-1-phosphate(S1P) are the main membrane-derived lipid signalling molecules.LPA has a 3-carbon glycerol backbone, with an attached single acylor alkyl chain of varying length which imparts some differences inreceptor efficacy [12]. Whilst some LPA production may occur

894 W. Ng et al. / Journal of Clinical Neuroscience 21 (2014) 893–898
intracellularly, much of it is produced extracellularly by secretedenzymes. There are three known pathways: (1) cleavage of LPL(such as lysophosphatidylcholine) by lysophospholipase D, (2)deacylation of phosphatidic acid by phospholipase A1 and A2,and (3) mild oxidation of low-density lipoprotein (non-enzymatic).
Lysophospholipase D is now more commonly known as ATXwhich derives its name from early characterisation of its stimula-tory effect on melanoma cell motility [13–15]. It is also known asectonucleotide pyrophosphatase and phosphodiesterase-2(ENPP-2) and is part of the family of ENPP enzymes which are tra-ditionally known for their involvement in nucleotide metabolism[13]. LPA production is mainly via ATX catalysis. This is confirmedby the uniformly fatal outcome of homozygous ATX (Atx�/�) knock-out mice and the 50% reduction in circulating plasma LPA levels inembryos with heterozygous ATX (Atx+/�) expression [16,17].
The phospholipase A enzymes play an important role indetermining the position of the acylation of the phosphoglycerolbackbone and so, whilst there is currently no evidence of its upreg-ulation, it may still be an important pathway in pathological states,as it may influence the action of liberated LPA species by playing arole in determining the dominant LPA species produced (acyl oralkyl) [18].
Physiologic LPA is present in small amounts in tissues since itregulates its own production via negative feedback inhibition ofits main synthesising enzyme, ATX [19]. Thus, the influence of thissignalling pathway is largely autocrinic/paracrinic [12–14].Further, circulating platelets and erythrocytes secrete LPA whichis then bound to albumin to protect it from rapid enzymatic degra-dation by lysophospholipases, lipid phosphate phosphatases andLPA acyl transferases [12]. Cancer cells can produce increasedamounts of ATX/LPA and it has been previously reported that somemalignant effusions (such as those of ovarian cancer) have elevatedlevels of LPA compared to other malignancies [2,20,21]. In addition,circulating plasma levels of LPA can be elevated in malignancy andthis may be the result of elevated production (induction ofATX) [14].
3. LPA receptors
Only a brief review of the LPA receptors will be provided here asthere are many other detailed reviews available [12,16,22–24]. Atthe time of writing, the International Union of Basic and ClinicalPharmacology had recognised six definitive G-protein coupled-LPA receptors (collectively LPAR) designated LPA1–6 (Table 1).Broadly, the receptors fall into two families: endothelial differenti-ation gene (Edg) and non-Edg (purinergic) receptors [12]. LPA1
(Edg2), LPA2 (Edg4) and LPA3 (Edg7) are members of the Edg family
Table 1LPA receptors and their relevance to cancer research
LPA receptor Coupled G-proteins Relevance to cancer research
Edg LPA1 Gi/o, Gq, G12/13 – Dominant LPA receptor exp– Increased expression in can– Linked to cell proliferation,
LPA2 Gi/o, Gq, G12/13 – Not present in adult CNS– Not markedly elevated in G
LPA3 Gi/o, Gq – Expressed in adult CNS– Not markedly elevated in G
Non-Edg LPA4 Gs, Gq, G12/13 – Activation of LPA4 has beenantagonistic (anti-migration,
LPA5 Gq, G12/13Increase in cAMP notshown to be Gs-mediated
– Activation has been shown
LPA6 G12/13, (possible role for Gs, Gi) – Possible similar function to– May regulate vascular perm
cAMP = cyclic adenosine monophosphate, CNS = central nervous system, Edg = endothelia
and are the best characterised to our knowledge with LPA1 beingthe dominant LPA receptor in the central nervous system (CNS)[13,23,25]. LPA1 is coupled to the G-proteins, Gi/o, Gq and G12/13,which allows it to signal via multiple pathways, including majorcancer-related pathways such as mitogen-activated protein kinase(MAPK), Akt/PKB, and small GTPases such as Rho/ROCK [12,26].LPA1-induced PI3K signalling (which activates Akt/PKB) via thep110b/c subunits (of PI3K) has also been reported [27–30]. WhilstLPA2 is expressed in embryonic brain it has little to no expressionin the adult CNS [31]. LPA3 is expressed in the brain, but unlikeLPA1 and LPA2 it is coupled to the G-proteins, Gi/o and Gq, but notG12/13 and so is less responsive to LPL than LPA1 [27]. As LPA3 doesnot signal via G12/13, it is not involved in changes in cell morphol-ogy, and therefore unlikely to be involved in cell migration [27].
The non-Edg or purinergic family of LPA receptors are geneti-cally distinct from the original Edg family of LPA receptors [23].Importantly, this results in the non-Edg family having an increasedaffinity for alkyl-LPA species, as opposed to the Edg family havingincreased affinity for the acyl variants [22,23]. Current members ofthe non-Edg family include LPA4 (P2Y9), LPA5 (GPR92) and LPA6
(P2Y5). LPA4 signals via Gs, Gq and G12/13-proteins and probablyplays a role in cell motility and migration [23]. Mouse embryonicfibroblasts (MEF) derived from Lpa4 (null) knockout mice havebeen reported to exhibit hypersensitivity to LPA induced motility.These MEF had increased levels of phosphorylated Akt (pAkt) whenstimulated by LPA. The elevated pAkt levels and associated motileresponse were attenuated when Lpa4 was reintroduced into thecells, suggesting that LPA4 has an action which might suppress sig-nalling activity of the LPA1 receptor [32]. LPA5 (GPR92) and LPA6
(P2Y5) were both discovered subsequent to LPA4 and to our knowl-edge there is no reported role for LPA5 in tumourigenesis. LPA6
however, may cause morphological changes in vascular endothelialcells and therefore may play a minor or indirect role in gliomagen-esis via effects on vascular development [22,23,27].
LPL have a wide range of physiological and pathological effectsowing to the myriad G-proteins they are coupled to. As a result,they modulate numerous normal physiologic processes, includingcell proliferation and apoptosis, cell differentiation, cell adhesionand migration, cell morphology (including neurite retraction andsynaptic cleft shape modulation), normal CNS development andautoimmunity [12,13,16,18,25,33–35]. Dysregulation of theseevents is strongly implicated in tumourigenesis.
4. LPA signalling in cancer
Since Stracke et al. discovered the promotile effects of ATX onmelanoma cells in 1992, the LPA pathway has been investigated
Reference
ressed in adult CNS Kishi et al. [2]cers including GBMsurvival, migration/invasion
Kishi et al. [2]BM
Kishi et al. [2], Choi et al. [16]BM
shown to be functionallyanti-proliferation) to LPA1
Kato [59], Yanagida and Ishii [23], Lee et al. [32]
to reduce cell migration Jongsma et al. [54]
LPA4 Yanagida and Ishii [23]eability
l differentiation gene, GBM = glioblastoma multiforme, LPA = lysophosphatidic acid.

W. Ng et al. / Journal of Clinical Neuroscience 21 (2014) 893–898 895
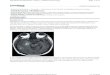
for its role in tumour invasion and metastasis [36]. However, itsoriginal role in nucleotide metabolism could not be directlyreconciled with this biological effect. It was the subsequent discov-ery by Umezu-Goto et al. [14] and Tokumura et al. [15] that ATXhad additional lysophospholipase D activity that triggered furtherinterest in LPA. Recent elucidation of the complex interactions ofLPA with known cancer signalling pathways will be discussedbelow (Fig. 1).
4.1. LPA interacts with cell survival pathways
EGFR signalling is commonly upregulated in GBM cells eithervia over-expression, EGFR gene amplification, or via the EGFR typeIII mutation, which produces a constitutively active form of EGFR[37,38]. EGFR activates downstream PI3K signalling which regu-lates cell survival via abrogation of apoptotic cell signals and thisis dysregulated in glioma. However, clinical trials with EGFR mono-clonal antibodies [3] were less promising than anticipated, possiblybecause of intracellular pathway cross-communication and redun-dancy [28,39].
It has been reported that LPA can intracellularly transactivateEGFR via the Gi-protein and that this is independent of normal li-gand activation of EGFR by EGF [28–30,40]. Further, it is possiblethat transactivation of p110b/c subunits of PI3K via Gi-protein cou-pled LPAR is more important than transactivation of p110a [41].This LPA-Gi-protein coupled signalling link has been reported inmany cancer cell lines including those from prostate, breast, colonand GBM [28,39].
Recently, Schleicher et al. reported that LPA signalling confersradioresistance to the murine glioma cell line GL-261 [42]. Inhibi-tion of LPA signalling with a combination ATX/LPAR inhibitor,a-bromomethylene phosphonate lysophosphatidic acid (BrP-LPA),resulted in impaired survival signals in this cell line when exposedto 3 Gy of irradiation. Knockdown of LPA1 and LPA3 (but not LPA2)signalling using siRNA resulted in reduced pAkt and also correlatedwith reduced survival in these cells. These results point to thehypothesis that LPA1 and LPA3 play an important role in radioresis-tance, via interaction with the PI3K/Akt pathway.
Fig. 1. Overview of intracellular interactions between lysophosphatidic acid (LPA) and eoverexpression or elevated activity of autotaxin, LPA1–3, EGFR and phosphoinositol-3-kigrowth factor, EGFR = epidermal growth factor receptor, ERK = extracellular signaMAPK = mitogen-activated phosphokinase, mTOR = mammalian target of rapamycin,trisphosphate, PI3K = phosphoinositol-3-kinase, PTEN = phosphatase-tensin homologue,
4.2. LPA signalling in cell proliferation
In addition to enhancing cell survival, transactivation of EGFRby LPA also upregulates cell proliferation via MAPK dependentmitogenic signalling. Whilst the transactivation of PI3K signallingcan be EGF independent, the transactivation of MAPK appears tobe dependent on agonistic activation of EGFR. Further, MAPKtransactivation may be linked to activation of PI3K [43]. Its trans-activation is also cell type dependent and has also been confirmedto be G-protein dependent [29,30,39,43]. Overall, transactivation ofMAPK by LPA appears to be more regulated (than PI3K) and in factmay also require (or be augmented by) concurrent MMP activity,which is also upregulated in gliomas [5,6].
4.3. LPL and ATX signalling influences cancer cell migration andinvasion
Alterations in cell morphology are tightly regulated and play animportant role in terms of chemotactic effect [44]. It is likely thatATX and LPA play their roles in glioma migration and invasionvia effects on cell morphology. Overexpression of ATX also corre-lates with increased invasiveness of breast cancer cells comparedto normal breast cells [45]. LPAR overexpression has also been re-ported in ovarian cancers and this has been postulated to create anautocrine/paracrine loop promoting proliferation (MAPK signal-ling) and suppressing apoptosis (PI3K signalling) [6,39,43]. Treat-ment of animals with metastatic ovarian cancer with Ki16425(selective LPA1/3 receptor antagonist) was demonstrated to havean in vivo anti-tumour effect without obvious adverse healtheffects on the treated nu/nu mice [46]. Also, use of a pan-LPARantagonist to treat hepatic metastases in an orthotopic breastcancer model showed significant reduction in tumour deposit size[19]. LPA4 has been shown to promote invadopodia formation inHT1080 fibrosarcoma cells, with Lpa4 shRNA abolishing this effect[47]. Invadopodia are involved in invasion and metastasis and areenriched with MMP, which are known for their key role in invasionand metastasis by providing a favourable milieu for cells to invade
pidermal growth factor receptor (EGFR) signalling pathways. Mutations leading tonase are often found in glioblastoma multiforme. ATX = autotaxin, EGF = epidermall-regulated kinases, LPA = lysophosphatidic acid, LPC = lysophosphatidylcholine,PIP2 = phosphatidylinositol 4,5-bisphosphate, PIP3 = phosphatidylinositol (3,4,5)-ROCK = Rho-associated kinase.

896 W. Ng et al. / Journal of Clinical Neuroscience 21 (2014) 893–898
through and also cleave and activate growth factors such as EGF[5,6].
In glioma, LPA has been shown to induce astrocyte motility bydramatically altering astrocyte shape [24,33,48]. ATX overexpres-sion has also been shown to enhance the invasion of U87 andU251 malignant glioma cells (autocrine effect) through oligoden-drocyte monolayers in vitro, whilst simultaneously reducing theadhesiveness of oligodendrocytes (paracrine effect). Depletion orinactivation of ATX ameliorates these effects. These findings mayprovide clues to the underlying pathogenesis of in vivo whitematter tract invasion of glioma cells [9].
Overexpression of ATX has been shown to be proportionatelyhigher at the leading invasive edges of tumours [9]. There are cur-rently no definitive reports to address whether the overexpressionof ATX and upregulation of LPAR is specific to certain cell typeswithin the tumour. But microglia may be recruited to the invasiveedge of the tumour by various chemokines (including LPA) andsubsequently facilitate invasion by producing ATX, LPA and EGF[49].
The dominant LPA receptors involved in mediating cell migra-tion and motility appear to be LPA1 and LPA4. LPA1 receptors havebeen linked to tumour cell motility in vitro and have been reportedto be overexpressed in GBM cell lines (SNB-78, SNB-75, SF-268,SF539 and SF-295) in combination with overexpression of ATX.This receptor/ligand dual overexpression may contribute to autoc-rinic stimulation of GBM cell motility and this can be abolishedwhen the cells are treated with the LPA1-3 receptor antagonist,Ki16425 [2].
Early work on the cellular mechanisms of LPA induced motilityrevealed a link to Rho signalling through either Gi (pertussis toxinsensitive) or G12/13-protein [13,50,51]. The subsequent Rho-dependent cytoskeletal rearrangement was reported to induce cellrounding in astrocytes and this may contribute to producing disco-hesive cells [51]. However, there have been some contradictoryreports with regards to Rho-related LPA signalling. For example,LPA induced glioma cell migration can be inhibited by blockingRho activation. Contrary to this, stimulation of Rho activity has alsobeen demonstrated to cause immobilisation of glioma cells [52].Recall the different evolutionary pathways of the Edg andnon-Edg families of LPAR and there may exist functional antago-nism between them to provide an explanation for these inconsis-tencies. Lee et al. reported that LPA4 is likely a functionalantagonist of LPA1, leading to a reduction in cell motility in MEFcells as well as DLD1 colorectal cancer cell lines and B103 neuronalcell lines [32]. It is possible that if LPA4 antagonises LPA1, that LPA4
Table 2Evidence for the potential therapeutic effect of lysophosphatidic acid and autotaxin modu
Compound Mechanism Animal Cell line
Ki16425 Selective LPA1/3 antagonist Balb-c nu/nu mice CHOb3wMDA-BO
Cyclic phosphatidicacid analogs
Selective ATX inhibitor NA A2058
NSC 48300 bithionol Small molecule ATXinhibitors
NA A2058
BrP-LPA Dual pan-LPA antagonist/ATX inhibitor
Balb-c nu/nu mice MDA-M
BrP-LPA Dual pan-LPA antagonist/ATX inhibitor
Balb-c nu/nu mice MDA-MA-549HCT-116
BrP-LPA Dual pan-LPA antagonist/ATX inhibitor
Balb-c nu/nu mice Mouse G
ATX = autotaxin, GBM = glioblastoma multiforme, BrP = a-bromomethylene phosphonat
somehow modulates activation of LPA1 or its signalling. Interest-ingly, activation of LPA5 has recently been reported to inhibit B16melanoma cell migration [53]. This further supports the notion thatthe non-Edg (purinergic; LPA4–6) family of LPA receptors may mod-ulate the effects of the Edg (LPA1–3) family.
In addition to the reported Rho-dependent mechanisms, LPA-mediated cell migration is also thought to occur via MAPK andPI3K signalling. LPA1 has been shown to activate MAPK and PI3K/Akt signalling to promote cell migration [16,54]. Also, it has beenconfirmed that ATX induced motility in melanoma cells is medi-ated via the p110c subunit (of PI3K) and that PI3K inhibitor can in-hibit this motile response in a dose-dependent manner [41].Further, Kim et al. reported that simultaneous knockout of Akt1and Akt2 abolished LPA induced motility in MEF [54]. Only re-expression of Akt1 following the double knockout restored the mo-tile response to LPA. They also established that exposure of MEF tothe pan-PI3K inhibitor (LY294002) or the LPAR antagonist,Ki16425, also completely abolished the motile response. Thiswould make it likely that the downstream activation of Akt1 isvia PI3K activation. However, it is not completely clear whetherthe subsequent phosphorylation of Akt is via transactivation ofEGFR, or via interaction of LPAR associated Gi-protein with PI3K.
4.4. The potential role of LPA in angiogenesis
LPA signalling seems to have a minor physiological role in angi-ogenesis, as S1P normally maintains this process. However, inpathological states, the role of LPA in angiogenesis may becomemore important [12]. Lpa1
(�/�) null mice have been shown to havea small but significant incidence (2.5%) of frontal intracerebralhaemorrhages which increases substantially in Lpa1
(�/�)/Lpa2(�/�)
double-null mice to 26%. This may support a role for LPA in vesselmaturation [12]. Furthermore, Atx(�/�) null murine embryos have aprofound absence of vessel maturation that causes uniform embry-onic mortality by day 10 [16]. This points to a potential role for LPAsignalling in neoangiogenesis. In vivo murine models investigatingATX have shown that ATX plays a role in angiogenesis similar tothat of vascular endothelial growth factor [55]. Other in vivo stud-ies investigating knockouts of Lpa1–4 have not had the profound ef-fect of Atx(�/�) knockout on vascular development, but instead,G12/13-protein knockouts have been found to have effects whichparallel those of ATX. Therefore, it is possible that the LPAR mayplay redundant roles in neoangiogenesis, or the effect ATX has onvascular development may be mediated through other pathwayssuch as vascular endothelial growth factor [23].
lation
(s) Cancer Effect Reference
t2
OvarianBreast
In vivo – regression of bonemetastases
Boucharaba et al. [46]
Melanoma In vitro – reduction in LPAproduction and subsequentcell migration and invasion
Baker et al. [58]
Melanoma In vitro – reduced cellmigration and invasion
Saunders et al. [59]
B-231 Breast In vitro – reduced cellmigration and invasionIn vivo – tumour regression
Zhang et al. [57]
B-231 BreastLungColon
In vivo – tumour growth delay Xu et al. [60]
L-261 GBM In vitro – radioprotection ofglioma cell linesIn vivo – tumour growthdelay
Schleicher et al. [42]
e, LPA = lysophosphatidic acid, NA = not applicable.

W. Ng et al. / Journal of Clinical Neuroscience 21 (2014) 893–898 897
Recent evidence also suggests that LPA may bind intracellulartargets such as peroxisome proliferator-activated receptor-c(PPAR-c) to cause neointimal formation [27]. It is unclear as towhether LPA migrates across the cell membrane but the effectsof LPA induced PPAR-c activity are significantly diminished inLpa1
(�/�) knockout mice. Regardless, specific unsaturated acyl LPAinteract with PPAR-c to induce neointimal formation and are likelyproduced by phosphorylation of monoacylglycerols (mitochondrialacylglycerol kinase) or by conversion of phosphatidic acid to LPAby cytosolic phospholipase A2 [18,27,56]. The role of LPA andATX in angiogenesis no doubt complements their putative rolesin tumour growth and invasion/metastasis.
5. Discussion
Decades of glioma research are now seeing the beginnings ofmolecular and genetic profiling of gliomas. Additionally, glioma-derived cancer stem cells have been identified, providing a newframework to investigate this disease’s resistance to treatmentand subsequent recurrence. Much of the evidence for the role ofLPA in cancer is spread across different cell lineages making it diffi-cult to extrapolate some of the knowledge to gliomas. However, theemerging reports regarding LPA and gliomagenesis for the most partecho the findings in other solid tumours. The discovery of ATX’s che-motactic effects has brought the spotlight to this field, resulting ininvaluable insights into the complexities of intracellular cross-communication and pathway redundancies. It has also lead to pre-clinical trials demonstrating the potential of this pathway as a targetin malignancies (Table 2). Recently, the potential benefits of target-ing ATX-LPA signalling were shown in murine breast cancer modelsusing a combination ATX inhibitor and LPAR antagonist. Thistreatment reduced in vitro migration of breast cancer cells andcaused tumour regression in vivo [57]. This strategy is based onthe theory that the Edg (LPA1–3) receptors may be the main pro-carcinogenic mediators of cancer in dysregulated LPA signalling.
An alternative strategy to targeting LPA signalling might look toproduce agonists to the non-Edg (LPA4/5) receptor family, as there ismounting evidence that they might mitigate the pro-carcinogeniceffects of the Edg receptors. This strategy may have benefits ofbeing more selective. However, the relative underexpression or ab-sence of these receptors in some tumours may limit its value. LPA1
has been shown to be the dominant LPAR expressed in gliomas butsome glioma cell lines have also been shown to express moderatelevels of LPA4 [2]. Therefore, at least in some gliomas, there maybe merit to developing LPA4 agonists as an anti-cancer therapy.Also, an investigation into expression patterns of LPAR in differentcell subpopulations (glioma-derived cancer stem cells, progenitorcells, and recruited host cells such as microglia and reactive astro-cytes and oligodendroglia) within the glioma microenvironmentis critical to ascertain the true value of LPA modulation as a treat-ment strategy.
Conflicts of Interest/Disclosures
The authors declare that they have no financial or otherconflicts of interest in relation to this research and its publication.
Acknowledgements
This study was funded by the Neurosurgical Society ofAustralasia/LifeHealthCare scholarship (2012) and the BrainFoundation of Australia Brain Tumours Award (2012) and theVictorian State Government’s Department of Innovation, Industryand Regional Development’s Operational Infrastructure SupportProgram. AP is supported by a National Health and MedicalResearch Council Career Development Fellowship.
References
[1] Stupp R, Hegi ME, Mason WP, et al. Effects of radiotherapy with concomitantand adjuvant temozolomide versus radiotherapy alone on survival inglioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol 2009;10:459–66.
[2] Kishi Y, Okudaira S, Tanaka M, et al. Autotaxin is overexpressed in glioblastomamultiforme and contributes to cell motility of glioblastoma by convertinglysophosphatidylcholine to lysophosphatidic acid. J Biol Chem 2006;281:17492–500.
[3] Camara-Quintana JQ, Nitta RT, Li G. Pathology: commonly monitoredglioblastoma markers: EFGR, EGFRvIII, PTEN, and MGMT. Neurosurg Clin NAm 2012;23:237–46.
[4] Wen PY, Lee EQ, Reardon DA, et al. Current clinical development of PI3Kpathway inhibitors in glioblastoma. Neuro Oncol 2012;14:819–29.
[5] Pullen NA, Anand M, Cooper PS, et al. Matrix metalloproteinase-1 expressionenhances tumorigenicity as well as tumor-related angiogenesis and isinversely associated with TIMP-4 expression in a model of glioblastoma. JNeurooncol 2012;106:461–71.
[6] Gschwind A, Prenzel N, Ullrich A. Lysophosphatidic acid-induced squamouscell carcinoma cell proliferation and motility involves epidermal growth factorreceptor signal transactivation. Cancer Res 2002;62:6329–36.
[7] Tchaicha JH, Reyes SB, Shin J, et al. Glioblastoma angiogenesis and tumor cellinvasiveness are differentially regulated by b8 integrin. Cancer Res 2011;71:6371–81.
[8] D’Abaco GM, Kaye AH. Integrins: molecular determinants of glioma invasion. JClin Neurosci 2007;14:1041–8.
[9] Hoelzinger DB, Nakada M, Demuth T, et al. Autotaxin: a secreted autocrine/paracrine factor that promotes glioma invasion. J Neurooncol 2007;86:297–309.
[10] Singh SK, Clarke ID, Terasaki M, et al. Identification of a cancer stem cell inhuman brain tumors. Cancer Res 2003;63:5821–8.
[11] Hemmati HD, Nakano I, Lazareff JA, et al. Cancerous stem cells can arise frompediatric brain tumors. Proc Natl Acad Sci USA 2003;100:15178–83.
[12] Ishii I, Fukushima N, Ye X, et al. Lysophospholipid receptors: signaling andbiology. Annu Rev Biochem 2004;73:321–54.
[13] Moolenaar WH. Lysophospholipids in the limelight: autotaxin takes centerstage. J Cell Biol 2002;158:197–9.
[14] Umezu-Goto M, Kishi Y, Taira A, et al. Autotaxin has lysophospholipase Dactivity leading to tumor cell growth and motility by lysophosphatidic acidproduction. J Cell Biol 2002;158:227–33.
[15] Tokumura A, Majima E, Kariya Y, et al. Identification of human plasmalysophospholipase D, a lysophosphatidic acid-producing enzyme, as autotaxin,a multifunctional phosphodiesterase. J Biol Chem 2002;277:39436–42.
[16] Choi JW, Lee CW, Chun J. Biological roles of lysophospholipid receptorsrevealed by genetic null mice: an update. Biochim Biophys Acta 2008;1781:531–9.
[17] Tanaka M, Okudaira S, Kishi Y, et al. Autotaxin stabilizes blood vessels and isrequired for embryonic vasculature by producing lysophosphatidic acid. J BiolChem 2006;281:25822–30.
[18] Aoki J, Inoue A, Okudaira S. Two pathways for lysophosphatidic acidproduction. Biochim Biophys Acta 2008;1781:513–8.
[19] Prestwich GD, Gajewiak J, Zhang H, et al. Phosphatase-resistant analogues oflysophosphatidic acid: agonists promote healing, antagonists and autotaxininhibitors treat cancer. Biochim Biophys Acta 2008;1781:588–94.
[20] Baker DL, Morrison P, Miller B, et al. Plasma lysophosphatidic acidconcentration and ovarian cancer. JAMA 2002;287:3081–2.
[21] Westermann AM, Havik E, Postma FR, et al. Malignant effusions containlysophosphatidic acid (LPA)-like activity. Ann Oncol 1998;9:437–42.
[22] Chun J, Hla T, Lynch KR, et al. International Union of Basic and ClinicalPharmacology. LXXVIII. Lysophospholipid receptor nomenclature. PharmacolRev 2010;62:579–87.
[23] Yanagida K, Ishii S. Non-Edg family LPA receptors: the cutting edge of LPAresearch. J Biochem 2011;150:223–32.
[24] Frisca F, Sabbadini RA, Goldshmit Y, et al. Biological effects of lysophosphatidicacid in the nervous system. Int Rev Cell Mol Biol 2012;296:273–322.
[25] Anliker B. Lysophospholipid G protein-coupled receptors. J Biol Chem2004;279:20555–8.
[26] Keller JN, Steiner MR, Holtsberg FW, et al. Lysophosphatidic acid-inducedproliferation-related signals in astrocytes. J Neurochem 1997;69:1073–84.
[27] Meyer zu Heringdorf D, Jakobs KH. Lysophospholipid receptors: signalling,pharmacology and regulation by lysophospholipid metabolism. BiochimBiophys Acta 2007;1768:923–40.
[28] Prenzel N, Zwick E, Daub H, et al. EGF receptor transactivation by G-protein-coupled receptors requires metalloproteinase cleavage of proHB-EGF. Nature1999;402:884–8.
[29] Daub H, Weiss FU, Wallasch C, et al. Role of transactivation of the EGF receptorin signalling by G-protein-coupled receptors. Nature 1996;379:557–60.
[30] Daub H, Wallasch C, Lankenau A, et al. Signal characteristics of G protein-transactivated EGF receptor. EMBO J 1997;16:7032–44.
[31] Goldshmit Y, Munro K, Leong SY, et al. LPA receptor expression in the centralnervous system in health and following injury. Cell Tissue Res2010;341:23–32.
[32] Lee Z, Cheng CT, Zhang H, et al. Role of LPA4/p2y9/GPR23 in negativeregulation of cell motility. Mol Biol Cell 2008;19:5435–45.

898 W. Ng et al. / Journal of Clinical Neuroscience 21 (2014) 893–898
[33] Tabuchi S, Kume K, Aihara M, et al. Expression of lysophosphatidic acidreceptor in rat astrocytes: mitogenic effect and expression of neurotrophicgenes. Neurochem Res 2000;25:573–82.
[34] Savaskan NE, Rocha L, Kotter MR, et al. Autotaxin (NPP-2) in the brain: celltype-specific expression and regulation during development and afterneurotrauma. Cell Mol Life Sci 2007;64:230–43.
[35] Steiner MR, Holtsberg FW, Keller JN, et al. Lysophosphatidic acid induction ofneuronal apoptosis and necrosis. Ann N Y Acad Sci 2000;905:132–41.
[36] Stracke ML, Krutzsch HC, Unsworth EJ, et al. Identification, purification, andpartial sequence analysis of autotaxin, a novel motility-stimulating protein. JBiol Chem 1992;267:2524–9.
[37] Omidfar K, Shirvani Z. Single domain antibodies: a new concept for epidermalgrowth factor receptor and EGFRvIII targeting. DNA Cell Biol 2012;31:1015–26.
[38] Gan HK, Kaye AH, Luwor RB. The EGFRvIII variant in glioblastoma multiforme. JClin Neurosci 2009;16:748–54.
[39] Gschwind A, Zwick E, Prenzel N, et al. Cell communication networks:epidermal growth factor receptor transactivation as the paradigm forinterreceptor signal transmission. Oncogene 2001;20:1594–600.
[40] Hernández M, Barrero MJ, Crespo MS, et al. Lysophosphatidic acid inhibitsCa2+ signaling in response to epidermal growth factor receptor stimulation inhuman astrocytoma cells by a mechanism involving phospholipase C(gamma)and a G(alphai) protein. J Neurochem 2000;75:1575–82.
[41] Lee HY, Bae GU, Jung ID, et al. Autotaxin promotes motility via G protein-coupled phosphoinositide 3-kinase gamma in human melanoma cells. FEBSLett 2002;515:137–40.
[42] Schleicher SM, Thotala DK, Linkous AG, et al. Autotaxin and LPA receptorsrepresent potential molecular targets for the radiosensitization of murineglioma through effects on tumor vasculature. PLoS One 2011;6:e22182.
[43] Shah BH, Neithardt A, Chu DB, et al. Role of EGF receptor transactivation inphosphoinositide 3-kinase-dependent activation of MAP kinase by GPCRs. JCell Physiol 2006;206:47–57.
[44] Ren XD, Kiosses WB, Schwartz MA. Regulation of the small GTP-bindingprotein Rho by cell adhesion and the cytoskeleton. EMBO J 1999;18:578–85.
[45] Yang SY, Lee J, Park CG, et al. Expression of autotaxin (NPP-2) is closely linkedto invasiveness of breast cancer cells. Clin Exp Metastasis 2002;19:603–8.
[46] Boucharaba A, Serre CM, Guglielmi J, et al. The type 1 lysophosphatidic acidreceptor is a target for therapy in bone metastases. Proc Nat Acad Sci USA2006;103:9643–8.
[47] Harper K, Arsenault D, Boulay-Jean S, et al. Autotaxin promotes cancerinvasion via the lysophosphatidic acid receptor 4: participation of the cyclicAMP/EPAC/Rac1 signaling pathway in invadopodia formation. Cancer Res2010;70:4634–43.
[48] Frisca F, Crombie DE, Dottori M, et al. Rho/ROCK pathway is essential to theexpansion, differentiation, and morphological rearrangements of humanneural stem/progenitor cells induced by lysophosphatidic acid. J Lipid Res2013;54:1192–206.
[49] Hoelzinger DB, Demuth T, Berens ME. Autocrine factors that sustain gliomainvasion and paracrine biology in the brain microenvironment. JNCI J NatlCancer Inst 2007;99:1583–93.
[50] Clair T, Lee HY, Liotta LA, et al. Autotaxin is an exoenzyme possessing50-nucleotide phosphodiesterase/ATP pyrophosphatase and ATPase activities. JBiol Chem 1997;272:996–1001.
[51] Seasholtz TM, Radeff-Huang J, Sagi SA, et al. Rho-mediated cytoskeletalrearrangement in response to LPA is functionally antagonized by Rac1 andPIP2. J Neurochem 2004;91:501–12.
[52] Khalil BD, El-Sibai M. Rho GTPases in primary brain tumor malignancy andinvasion. J Neurooncol 2012.
[53] Jongsma M, Matas-Rico E, Rzadkowski A, et al. LPA is a chemorepellent for B16melanoma cells: action through the cAMP-elevating LPA5 receptor. PLoS One2011;6:e29260.
[54] Kim EK, Yun SJ, Do KH, et al. Lysophosphatidic acid induces cell migrationthrough the selective activation of Akt1. Exp Mol Med 2008;40:445–52.
[55] Nam SW, Clair T, Kim YS, et al. Autotaxin (NPP-2), a metastasis-enhancingmotogen, is an angiogenic factor. Cancer Res 2001;61:6938–44.
[56] Tigyi G. Aiming drug discovery at lysophosphatidic acid targets. Br J Pharmacol2010;161:241–70.
[57] Zhang H, Xu X, Gajewiak J, et al. Dual activity lysophosphatidic acid receptorpan-antagonist/autotaxin inhibitor reduces breast cancer cell migrationin vitro and causes tumor regression in vivo. Cancer Res 2009;69:5441–9.
[58] Baker DL, Fujiwara Y, Pigg KR, et al. Carba analogs of cyclic phosphatidic acidare selective inhibitors of autotaxin and cancer cell invasion and metastasis. JBiol Chem 2006;281:22786–93.
[59] Saunders LP, Ouellette A, Bandle R, et al. Identification of small-moleculeinhibitors of autotaxin that inhibit melanoma cell migration and invasion. MolCancer Ther 2008;7:3352–62.
[60] Xu X, Yang G, Zhang H, et al. Evaluating dual activity LPA receptor pan-antagonist/autotaxin inhibitors as anti-cancer agents in vivo using engineeredhuman tumors. Prostaglandins Other Lipid Mediat 2009;89:140–6.