Embed Size (px)
Citation preview

Structure
Article
Complex Formation and Light Activationin Membrane-Embedded Sensory Rhodopsin IIas Seen by Solid-State NMR SpectroscopyManuel Etzkorn,1,4,5 Karsten Seidel,1,4,6 Lin Li,2,4 Swetlana Martell,2 Matthias Geyer,2 Martin Engelhard,2,*and Marc Baldus1,3,*1Department for NMR-based Structural Biology, Max Planck Institute for Biophysical Chemistry, Am Fassberg 11, 37077 Gottingen, Germany2Department of Physical Biochemistry, Max Planck Institute for Molecular Physiology, Otto-Hahn-Strasse 11, 44227 Dortmund, Germany3Bijvoet Center for Biomolecular Research, Utrecht University, Padualaan 8, 3584 CH Utrecht, Netherlands4These authors contributed equally to this work5Present address: Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, 240 Longwood Avenue,
Boston, MA 02115, USA6Present address: BASF SE, 67056 Ludwigshafen, Germany
*Correspondence: [email protected] (M.E.), [email protected] (M.B.)
DOI 10.1016/j.str.2010.01.011
SUMMARY
Microbial rhodopsinsexecutediversebiological func-tions in the cellular membrane. A mechanistic under-standing of their functional profile is, however, stilllimited. We used solid-state NMR (ssNMR) spectros-copy to study structure and dynamics of a 2 3 400amino acid sensory rhodopsin/transducer (SRII/HtrII)complex from Natronomonas pharaonis in a naturalmembrane environment. We found a receptor-trans-ducer binding interface in the ground state that sig-nificantly extends beyond the available X-ray struc-ture. This binding domain involves the EF loop ofthe receptor and stabilizes the functionally relevant,directly adjacent HAMP domain of the transducer.Using 2D ssNMR difference spectroscopy, we iden-tified protein residues that may act as a functionalmodule around the retinal binding site during theearly events of protein activation. These latter pro-tein segments, the inherent plasticity of the HAMPdomain, and the observation of an extended SRII/HtrII membrane-embedded interface may be crucialcomponents for optimal signal relay efficiency acrossthe cell membrane.
INTRODUCTION
Transmission of signals across the cellular membrane is essen-
tial for life. In this process, understanding the molecular details
that determine protein structure and organization and, at the
same time, allow for structural changes in response to external
stimuli is crucial. A prominent example are microbial rhodopsins
that exert diverse biological functions including light sensing and
molecular ion transport across the cellular membrane (Klare
et al., 2008). Sensory rhodopsin II (SRII) is an important member
of this family triggering a signal transduction chain homologous
to the two-component system of eubacterial chemotaxis. Similar
Structure 18, 2
to other microbial rhodopsins, sensory rhodopsins are seven-
helix (A�G) membrane proteins containing retinal as cofactor,
which is bound to a lysine residue on helix G via a protonated
Schiff base. The color of this complex (lmax around 495nm) is
regulated by specific protein-retinal interactions. Upon light
excitation the all-trans chromophore isomerizes to a 13-cis con-
figuration, thereby inducing conformational changes, which lead
to the activation of the cognate transducer HtrII (Klare et al.,
2004; Sasaki and Spudich, 2008). X-ray crystallography has
provided detailed structural insight into rhodopsins and in partic-
ular into SRII. Structures were not only obtained of SRII in free
(Luecke et al., 2001; Royant et al., 2001) and transducer-bound
form (Gordeliy et al., 2002), but also photocycle intermediates K
and late M were characterized at the atomic level (Edman et al.,
2002; Moukhametzianov et al., 2006).
Yet, with the structures available, other biophysical evidence,
in particular regarding the involvement of SRII helices F and G
in the initial stages of activation, has been difficult to discern
(Bordignon et al., 2007; Wegener et al., 2001). In addition,
tracking structural changes involving the cognate transducer
has been complicated since all available X-ray structures are
confined to the membrane-spanning helices, equivalent to resi-
dues 23�82 in Natronomonas pharaonis HtrII (NpHtrII). Residues
beyond Leu82 could not be resolved, and the structure of the
corresponding HAMP domain—a protein module (found in histi-
dine kinases, adenylyl cyclases, methyl-accepting chemotaxis
proteins, and phosphatases) required for signal transfer into
the cytoplasmic domain—reaching from Gly84 to Glu136 is
unknown. EPR distance measurements (Doebber et al., 2008)
have suggested that this segment adopts a conformation homol-
ogous to the structure of the isolated HAMP domain fragment of
Archaeoglobus fulgidus Af1503 (Hulko et al., 2006). According to
the interpretation of available data, the HAMP domain can adopt
two states with dynamic (dHAMP) and more compact (cHAMP)
properties (Bordignon et al., 2005; Doebber et al., 2008). Similar
conclusions were drawn from electron microscopy experiments
for a chemotaxis receptor (Khursigara et al., 2008).
Structural and dynamic information that reconciles crystallo-
graphic and other biophysical evidence on the formation and
activation of the NpSRII/NpHtrII complex is hence highly
93–300, March 10, 2010 ª2010 Elsevier Ltd All rights reserved 293
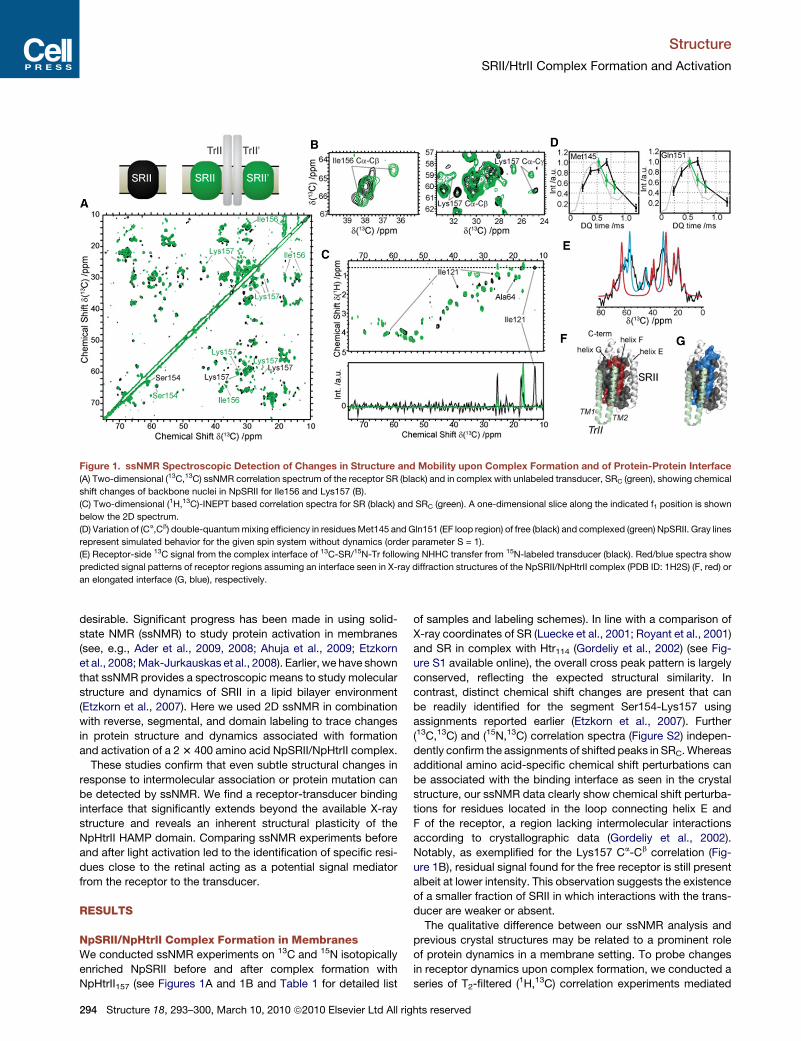
Figure 1. ssNMR Spectroscopic Detection of Changes in Structure and Mobility upon Complex Formation and of Protein-Protein Interface
(A) Two-dimensional (13C,13C) ssNMR correlation spectrum of the receptor SR (black) and in complex with unlabeled transducer, SRC (green), showing chemical
shift changes of backbone nuclei in NpSRII for Ile156 and Lys157 (B).
(C) Two-dimensional (1H,13C)-INEPT based correlation spectra for SR (black) and SRC (green). A one-dimensional slice along the indicated f1 position is shown
below the 2D spectrum.
(D) Variation of (Ca,Cb) double-quantum mixing efficiency in residues Met145 and Gln151 (EF loop region) of free (black) and complexed (green) NpSRII. Gray lines
represent simulated behavior for the given spin system without dynamics (order parameter S = 1).
(E) Receptor-side 13C signal from the complex interface of 13C-SR/15N-Tr following NHHC transfer from 15N-labeled transducer (black). Red/blue spectra show
predicted signal patterns of receptor regions assuming an interface seen in X-ray diffraction structures of the NpSRII/NpHtrII complex (PDB ID: 1H2S) (F, red) or
an elongated interface (G, blue), respectively.
Structure
SRII/HtrII Complex Formation and Activation
desirable. Significant progress has been made in using solid-
state NMR (ssNMR) to study protein activation in membranes
(see, e.g., Ader et al., 2009, 2008; Ahuja et al., 2009; Etzkorn
et al., 2008; Mak-Jurkauskas et al., 2008). Earlier, we have shown
that ssNMR provides a spectroscopic means to study molecular
structure and dynamics of SRII in a lipid bilayer environment
(Etzkorn et al., 2007). Here we used 2D ssNMR in combination
with reverse, segmental, and domain labeling to trace changes
in protein structure and dynamics associated with formation
and activation of a 2 3 400 amino acid NpSRII/NpHtrII complex.
These studies confirm that even subtle structural changes in
response to intermolecular association or protein mutation can
be detected by ssNMR. We find a receptor-transducer binding
interface that significantly extends beyond the available X-ray
structure and reveals an inherent structural plasticity of the
NpHtrII HAMP domain. Comparing ssNMR experiments before
and after light activation led to the identification of specific resi-
dues close to the retinal acting as a potential signal mediator
from the receptor to the transducer.
RESULTS
NpSRII/NpHtrII Complex Formation in MembranesWe conducted ssNMR experiments on 13C and 15N isotopically
enriched NpSRII before and after complex formation with
NpHtrII157 (see Figures 1A and 1B and Table 1 for detailed list
294 Structure 18, 293–300, March 10, 2010 ª2010 Elsevier Ltd All rig
of samples and labeling schemes). In line with a comparison of
X-ray coordinates of SR (Luecke et al., 2001; Royant et al., 2001)
and SR in complex with Htr114 (Gordeliy et al., 2002) (see Fig-
ure S1 available online), the overall cross peak pattern is largely
conserved, reflecting the expected structural similarity. In
contrast, distinct chemical shift changes are present that can
be readily identified for the segment Ser154-Lys157 using
assignments reported earlier (Etzkorn et al., 2007). Further
(13C,13C) and (15N,13C) correlation spectra (Figure S2) indepen-
dently confirm the assignments of shifted peaks in SRC. Whereas
additional amino acid-specific chemical shift perturbations can
be associated with the binding interface as seen in the crystal
structure, our ssNMR data clearly show chemical shift perturba-
tions for residues located in the loop connecting helix E and
F of the receptor, a region lacking intermolecular interactions
according to crystallographic data (Gordeliy et al., 2002).
Notably, as exemplified for the Lys157 Ca-Cb correlation (Fig-
ure 1B), residual signal found for the free receptor is still present
albeit at lower intensity. This observation suggests the existence
of a smaller fraction of SRII in which interactions with the trans-
ducer are weaker or absent.
The qualitative difference between our ssNMR analysis and
previous crystal structures may be related to a prominent role
of protein dynamics in a membrane setting. To probe changes
in receptor dynamics upon complex formation, we conducted a
series of T2-filtered (1H,13C) correlation experiments mediated
hts reserved
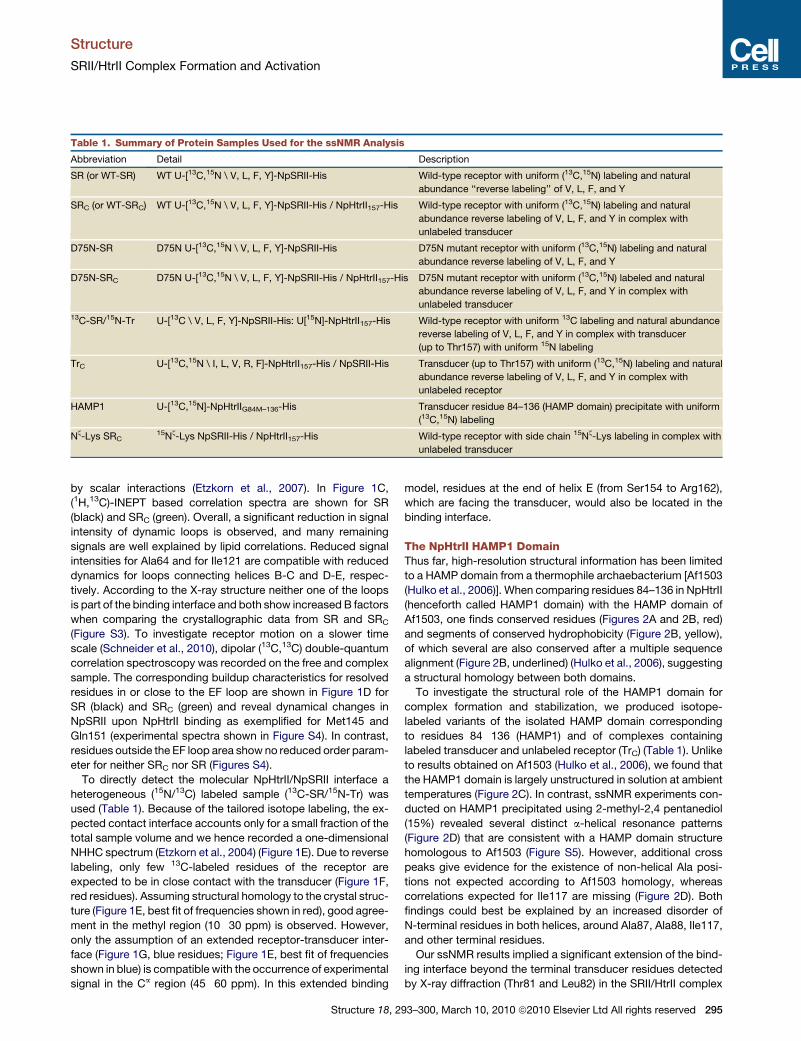
Table 1. Summary of Protein Samples Used for the ssNMR Analysis
Abbreviation Detail Description
SR (or WT-SR) WT U-[13C,15N \ V, L, F, Y]-NpSRII-His Wild-type receptor with uniform (13C,15N) labeling and natural
abundance ‘‘reverse labeling’’ of V, L, F, and Y
SRC (or WT-SRC) WT U-[13C,15N \ V, L, F, Y]-NpSRII-His / NpHtrII157-His Wild-type receptor with uniform (13C,15N) labeling and natural
abundance reverse labeling of V, L, F, and Y in complex with
unlabeled transducer
D75N-SR D75N U-[13C,15N \ V, L, F, Y]-NpSRII-His D75N mutant receptor with uniform (13C,15N) labeling and natural
abundance reverse labeling of V, L, F, and Y
D75N-SRC D75N U-[13C,15N \ V, L, F, Y]-NpSRII-His / NpHtrII157-His D75N mutant receptor with uniform (13C,15N) labeled and natural
abundance reverse labeling of V, L, F, and Y in complex with
unlabeled transducer13C-SR/15N-Tr U-[13C \ V, L, F, Y]-NpSRII-His: U[15N]-NpHtrII157-His Wild-type receptor with uniform 13C labeling and natural abundance
reverse labeling of V, L, F, and Y in complex with transducer
(up to Thr157) with uniform 15N labeling
TrC U-[13C,15N \ I, L, V, R, F]-NpHtrII157-His / NpSRII-His Transducer (up to Thr157) with uniform (13C,15N) labeling and natural
abundance reverse labeling of V, L, F, and Y in complex with
unlabeled receptor
HAMP1 U-[13C,15N]-NpHtrIIG84M–136-His Transducer residue 84–136 (HAMP domain) precipitate with uniform
(13C,15N) labeling
Nz-Lys SRC15Nz-Lys NpSRII-His / NpHtrII157-His Wild-type receptor with side chain 15Nz-Lys labeling in complex with
unlabeled transducer
Structure
SRII/HtrII Complex Formation and Activation
by scalar interactions (Etzkorn et al., 2007). In Figure 1C,
(1H,13C)-INEPT based correlation spectra are shown for SR
(black) and SRC (green). Overall, a significant reduction in signal
intensity of dynamic loops is observed, and many remaining
signals are well explained by lipid correlations. Reduced signal
intensities for Ala64 and for Ile121 are compatible with reduced
dynamics for loops connecting helices B-C and D-E, respec-
tively. According to the X-ray structure neither one of the loops
is part of the binding interface and both show increased B factors
when comparing the crystallographic data from SR and SRC
(Figure S3). To investigate receptor motion on a slower time
scale (Schneider et al., 2010), dipolar (13C,13C) double-quantum
correlation spectroscopy was recorded on the free and complex
sample. The corresponding buildup characteristics for resolved
residues in or close to the EF loop are shown in Figure 1D for
SR (black) and SRC (green) and reveal dynamical changes in
NpSRII upon NpHtrII binding as exemplified for Met145 and
Gln151 (experimental spectra shown in Figure S4). In contrast,
residues outside the EF loop area show no reduced order param-
eter for neither SRC nor SR (Figures S4).
To directly detect the molecular NpHtrII/NpSRII interface a
heterogeneous (15N/13C) labeled sample (13C-SR/15N-Tr) was
used (Table 1). Because of the tailored isotope labeling, the ex-
pected contact interface accounts only for a small fraction of the
total sample volume and we hence recorded a one-dimensional
NHHC spectrum (Etzkorn et al., 2004) (Figure 1E). Due to reverse
labeling, only few 13C-labeled residues of the receptor are
expected to be in close contact with the transducer (Figure 1F,
red residues). Assuming structural homology to the crystal struc-
ture (Figure 1E, best fit of frequencies shown in red), good agree-
ment in the methyl region (10�30 ppm) is observed. However,
only the assumption of an extended receptor-transducer inter-
face (Figure 1G, blue residues; Figure 1E, best fit of frequencies
shown in blue) is compatible with the occurrence of experimental
signal in the Ca region (45�60 ppm). In this extended binding
Structure 18, 2
model, residues at the end of helix E (from Ser154 to Arg162),
which are facing the transducer, would also be located in the
binding interface.
The NpHtrII HAMP1 DomainThus far, high-resolution structural information has been limited
to a HAMP domain from a thermophile archaebacterium [Af1503
(Hulko et al., 2006)]. When comparing residues 84–136 in NpHtrII
(henceforth called HAMP1 domain) with the HAMP domain of
Af1503, one finds conserved residues (Figures 2A and 2B, red)
and segments of conserved hydrophobicity (Figure 2B, yellow),
of which several are also conserved after a multiple sequence
alignment (Figure 2B, underlined) (Hulko et al., 2006), suggesting
a structural homology between both domains.
To investigate the structural role of the HAMP1 domain for
complex formation and stabilization, we produced isotope-
labeled variants of the isolated HAMP domain corresponding
to residues 84�136 (HAMP1) and of complexes containing
labeled transducer and unlabeled receptor (TrC) (Table 1). Unlike
to results obtained on Af1503 (Hulko et al., 2006), we found that
the HAMP1 domain is largely unstructured in solution at ambient
temperatures (Figure 2C). In contrast, ssNMR experiments con-
ducted on HAMP1 precipitated using 2-methyl-2,4 pentanediol
(15%) revealed several distinct a-helical resonance patterns
(Figure 2D) that are consistent with a HAMP domain structure
homologous to Af1503 (Figure S5). However, additional cross
peaks give evidence for the existence of non-helical Ala posi-
tions not expected according to Af1503 homology, whereas
correlations expected for Ile117 are missing (Figure 2D). Both
findings could best be explained by an increased disorder of
N-terminal residues in both helices, around Ala87, Ala88, Ile117,
and other terminal residues.
Our ssNMR results implied a significant extension of the bind-
ing interface beyond the terminal transducer residues detected
by X-ray diffraction (Thr81 and Leu82) in the SRII/HtrII complex
93–300, March 10, 2010 ª2010 Elsevier Ltd All rights reserved 295
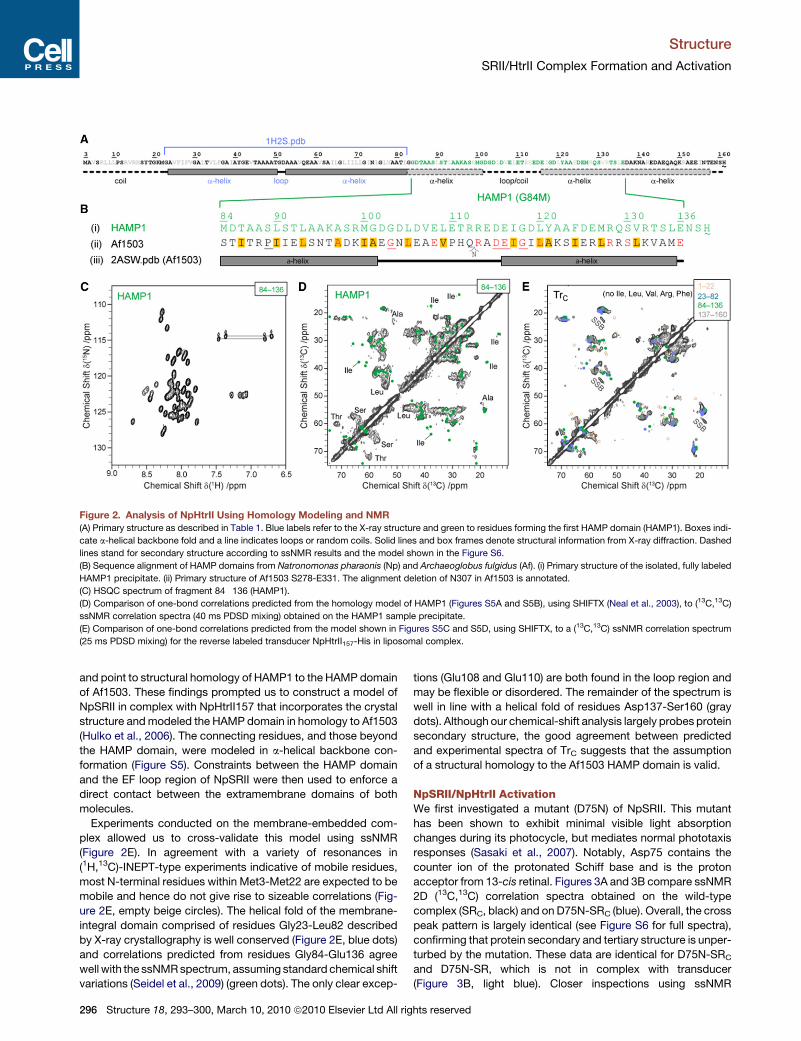
Figure 2. Analysis of NpHtrII Using Homology Modeling and NMR
(A) Primary structure as described in Table 1. Blue labels refer to the X-ray structure and green to residues forming the first HAMP domain (HAMP1). Boxes indi-
cate a-helical backbone fold and a line indicates loops or random coils. Solid lines and box frames denote structural information from X-ray diffraction. Dashed
lines stand for secondary structure according to ssNMR results and the model shown in the Figure S6.
(B) Sequence alignment of HAMP domains from Natronomonas pharaonis (Np) and Archaeoglobus fulgidus (Af). (i) Primary structure of the isolated, fully labeled
HAMP1 precipitate. (ii) Primary structure of Af1503 S278-E331. The alignment deletion of N307 in Af1503 is annotated.
(C) HSQC spectrum of fragment 84�136 (HAMP1).
(D) Comparison of one-bond correlations predicted from the homology model of HAMP1 (Figures S5A and S5B), using SHIFTX (Neal et al., 2003), to (13C,13C)
ssNMR correlation spectra (40 ms PDSD mixing) obtained on the HAMP1 sample precipitate.
(E) Comparison of one-bond correlations predicted from the model shown in Figures S5C and S5D, using SHIFTX, to a (13C,13C) ssNMR correlation spectrum
(25 ms PDSD mixing) for the reverse labeled transducer NpHtrII157-His in liposomal complex.
Structure
SRII/HtrII Complex Formation and Activation
and point to structural homology of HAMP1 to the HAMP domain
of Af1503. These findings prompted us to construct a model of
NpSRII in complex with NpHtrII157 that incorporates the crystal
structure and modeled the HAMP domain in homology to Af1503
(Hulko et al., 2006). The connecting residues, and those beyond
the HAMP domain, were modeled in a-helical backbone con-
formation (Figure S5). Constraints between the HAMP domain
and the EF loop region of NpSRII were then used to enforce a
direct contact between the extramembrane domains of both
molecules.
Experiments conducted on the membrane-embedded com-
plex allowed us to cross-validate this model using ssNMR
(Figure 2E). In agreement with a variety of resonances in
(1H,13C)-INEPT-type experiments indicative of mobile residues,
most N-terminal residues within Met3-Met22 are expected to be
mobile and hence do not give rise to sizeable correlations (Fig-
ure 2E, empty beige circles). The helical fold of the membrane-
integral domain comprised of residues Gly23-Leu82 described
by X-ray crystallography is well conserved (Figure 2E, blue dots)
and correlations predicted from residues Gly84-Glu136 agree
well with the ssNMR spectrum, assuming standard chemical shift
variations (Seidel et al., 2009) (green dots). The only clear excep-
296 Structure 18, 293–300, March 10, 2010 ª2010 Elsevier Ltd All rig
tions (Glu108 and Glu110) are both found in the loop region and
may be flexible or disordered. The remainder of the spectrum is
well in line with a helical fold of residues Asp137-Ser160 (gray
dots). Although our chemical-shift analysis largely probes protein
secondary structure, the good agreement between predicted
and experimental spectra of TrC suggests that the assumption
of a structural homology to the Af1503 HAMP domain is valid.
NpSRII/NpHtrII ActivationWe first investigated a mutant (D75N) of NpSRII. This mutant
has been shown to exhibit minimal visible light absorption
changes during its photocycle, but mediates normal phototaxis
responses (Sasaki et al., 2007). Notably, Asp75 contains the
counter ion of the protonated Schiff base and is the proton
acceptor from 13-cis retinal. Figures 3A and 3B compare ssNMR
2D (13C,13C) correlation spectra obtained on the wild-type
complex (SRC, black) and on D75N-SRC (blue). Overall, the cross
peak pattern is largely identical (see Figure S6 for full spectra),
confirming that protein secondary and tertiary structure is unper-
turbed by the mutation. These data are identical for D75N-SRC
and D75N-SR, which is not in complex with transducer
(Figure 3B, light blue). Closer inspections using ssNMR
hts reserved
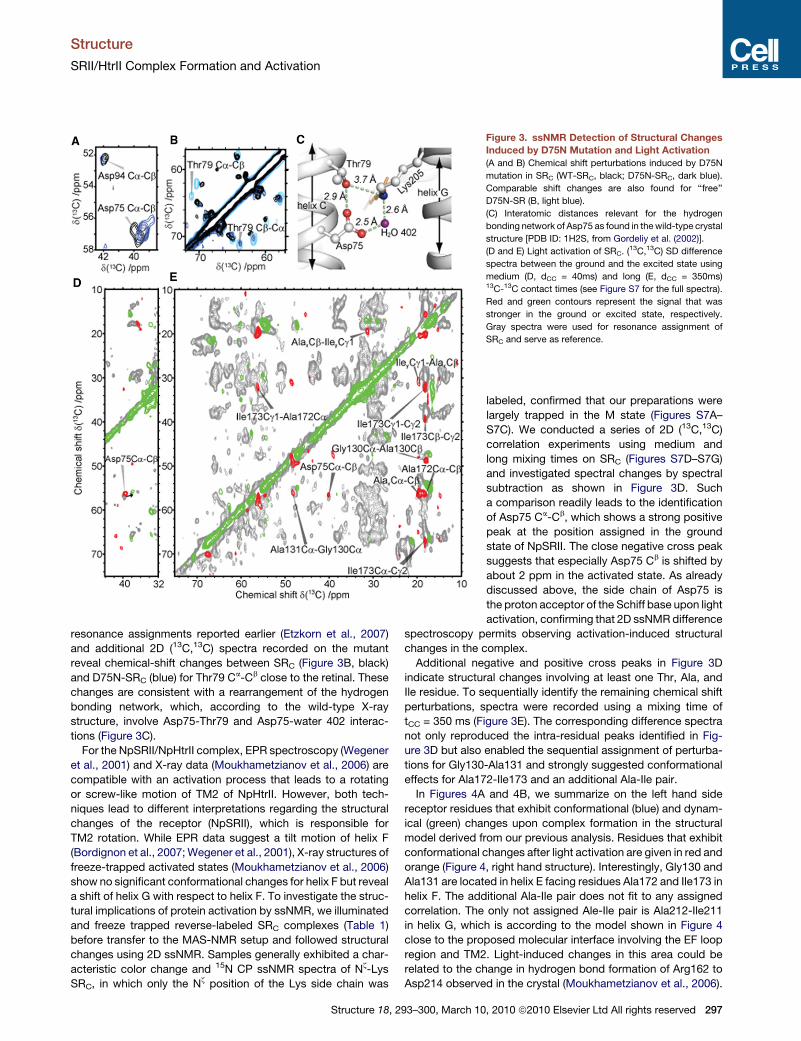
Figure 3. ssNMR Detection of Structural Changes
Induced by D75N Mutation and Light Activation
(A and B) Chemical shift perturbations induced by D75N
mutation in SRC (WT-SRC, black; D75N-SRC, dark blue).
Comparable shift changes are also found for ‘‘free’’
D75N-SR (B, light blue).
(C) Interatomic distances relevant for the hydrogen
bonding network of Asp75 as found in the wild-type crystal
structure [PDB ID: 1H2S, from Gordeliy et al. (2002)].
(D and E) Light activation of SRC. (13C,13C) SD difference
spectra between the ground and the excited state using
medium (D, dCC = 40ms) and long (E, dCC = 350ms)13C-13C contact times (see Figure S7 for the full spectra).
Red and green contours represent the signal that was
stronger in the ground or excited state, respectively.
Gray spectra were used for resonance assignment of
SRC and serve as reference.
Structure
SRII/HtrII Complex Formation and Activation
resonance assignments reported earlier (Etzkorn et al., 2007)
and additional 2D (13C,13C) spectra recorded on the mutant
reveal chemical-shift changes between SRC (Figure 3B, black)
and D75N-SRC (blue) for Thr79 Ca-Cb close to the retinal. These
changes are consistent with a rearrangement of the hydrogen
bonding network, which, according to the wild-type X-ray
structure, involve Asp75-Thr79 and Asp75-water 402 interac-
tions (Figure 3C).
For the NpSRII/NpHtrII complex, EPR spectroscopy (Wegener
et al., 2001) and X-ray data (Moukhametzianov et al., 2006) are
compatible with an activation process that leads to a rotating
or screw-like motion of TM2 of NpHtrII. However, both tech-
niques lead to different interpretations regarding the structural
changes of the receptor (NpSRII), which is responsible for
TM2 rotation. While EPR data suggest a tilt motion of helix F
(Bordignon et al., 2007; Wegener et al., 2001), X-ray structures of
freeze-trapped activated states (Moukhametzianov et al., 2006)
show no significant conformational changes for helix F but reveal
a shift of helix G with respect to helix F. To investigate the struc-
tural implications of protein activation by ssNMR, we illuminated
and freeze trapped reverse-labeled SRC complexes (Table 1)
before transfer to the MAS-NMR setup and followed structural
changes using 2D ssNMR. Samples generally exhibited a char-
acteristic color change and 15N CP ssNMR spectra of Nz-Lys
SRC, in which only the Nz position of the Lys side chain was
Structure 18, 293–300, March 10
labeled, confirmed that our preparations were
largely trapped in the M state (Figures S7A–
S7C). We conducted a series of 2D (13C,13C)
correlation experiments using medium and
long mixing times on SRC (Figures S7D–S7G)
and investigated spectral changes by spectral
subtraction as shown in Figure 3D. Such
a comparison readily leads to the identification
of Asp75 Ca-Cb, which shows a strong positive
peak at the position assigned in the ground
state of NpSRII. The close negative cross peak
suggests that especially Asp75 Cb is shifted by
about 2 ppm in the activated state. As already
discussed above, the side chain of Asp75 is
the proton acceptor of the Schiff base upon light
activation, confirming that 2D ssNMR difference
spectroscopy permits observing activation-induced structural
changes in the complex.
Additional negative and positive cross peaks in Figure 3D
indicate structural changes involving at least one Thr, Ala, and
Ile residue. To sequentially identify the remaining chemical shift
perturbations, spectra were recorded using a mixing time of
tCC = 350 ms (Figure 3E). The corresponding difference spectra
not only reproduced the intra-residual peaks identified in Fig-
ure 3D but also enabled the sequential assignment of perturba-
tions for Gly130-Ala131 and strongly suggested conformational
effects for Ala172-Ile173 and an additional Ala-Ile pair.
In Figures 4A and 4B, we summarize on the left hand side
receptor residues that exhibit conformational (blue) and dynam-
ical (green) changes upon complex formation in the structural
model derived from our previous analysis. Residues that exhibit
conformational changes after light activation are given in red and
orange (Figure 4, right hand structure). Interestingly, Gly130 and
Ala131 are located in helix E facing residues Ala172 and Ile173 in
helix F. The additional Ala-Ile pair does not fit to any assigned
correlation. The only not assigned Ale-Ile pair is Ala212-Ile211
in helix G, which is according to the model shown in Figure 4
close to the proposed molecular interface involving the EF loop
region and TM2. Light-induced changes in this area could be
related to the change in hydrogen bond formation of Arg162 to
Asp214 observed in the crystal (Moukhametzianov et al., 2006).
, 2010 ª2010 Elsevier Ltd All rights reserved 297
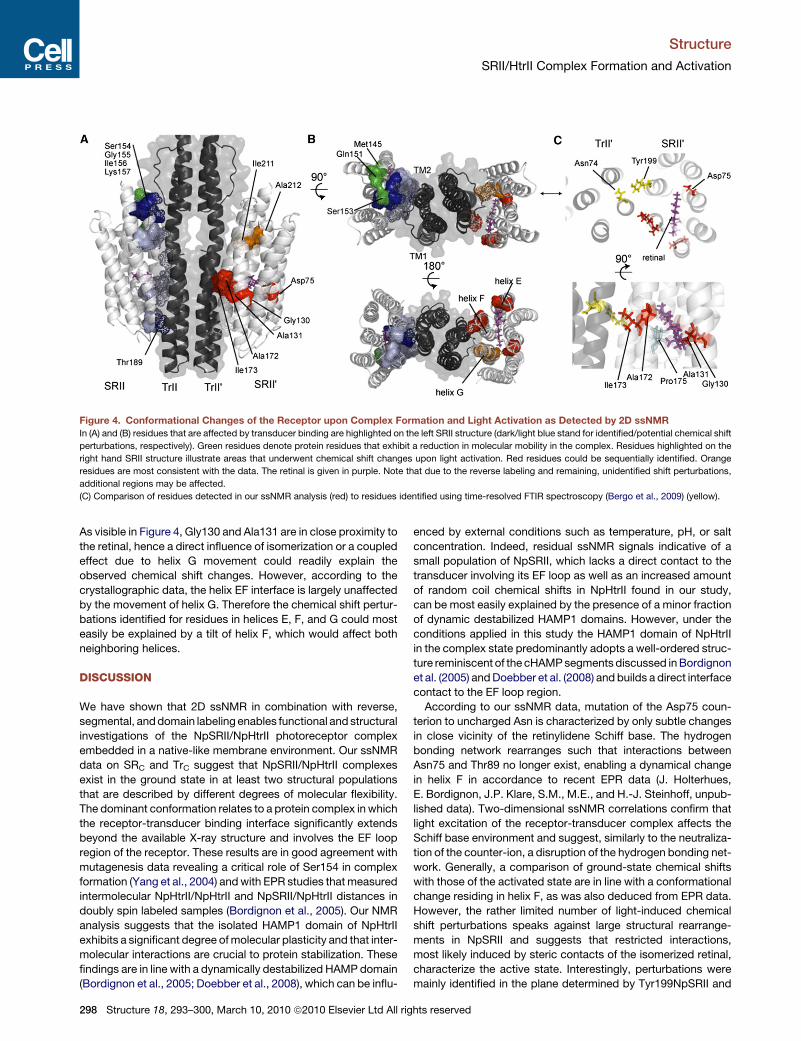
Figure 4. Conformational Changes of the Receptor upon Complex Formation and Light Activation as Detected by 2D ssNMR
In (A) and (B) residues that are affected by transducer binding are highlighted on the left SRII structure (dark/light blue stand for identified/potential chemical shift
perturbations, respectively). Green residues denote protein residues that exhibit a reduction in molecular mobility in the complex. Residues highlighted on the
right hand SRII structure illustrate areas that underwent chemical shift changes upon light activation. Red residues could be sequentially identified. Orange
residues are most consistent with the data. The retinal is given in purple. Note that due to the reverse labeling and remaining, unidentified shift perturbations,
additional regions may be affected.
(C) Comparison of residues detected in our ssNMR analysis (red) to residues identified using time-resolved FTIR spectroscopy (Bergo et al., 2009) (yellow).
Structure
SRII/HtrII Complex Formation and Activation
As visible in Figure 4, Gly130 and Ala131 are in close proximity to
the retinal, hence a direct influence of isomerization or a coupled
effect due to helix G movement could readily explain the
observed chemical shift changes. However, according to the
crystallographic data, the helix EF interface is largely unaffected
by the movement of helix G. Therefore the chemical shift pertur-
bations identified for residues in helices E, F, and G could most
easily be explained by a tilt of helix F, which would affect both
neighboring helices.
DISCUSSION
We have shown that 2D ssNMR in combination with reverse,
segmental, and domain labeling enables functional and structural
investigations of the NpSRII/NpHtrII photoreceptor complex
embedded in a native-like membrane environment. Our ssNMR
data on SRC and TrC suggest that NpSRII/NpHtrII complexes
exist in the ground state in at least two structural populations
that are described by different degrees of molecular flexibility.
The dominant conformation relates to a protein complex in which
the receptor-transducer binding interface significantly extends
beyond the available X-ray structure and involves the EF loop
region of the receptor. These results are in good agreement with
mutagenesis data revealing a critical role of Ser154 in complex
formation (Yang et al., 2004) and with EPR studies that measured
intermolecular NpHtrII/NpHtrII and NpSRII/NpHtrII distances in
doubly spin labeled samples (Bordignon et al., 2005). Our NMR
analysis suggests that the isolated HAMP1 domain of NpHtrII
exhibits a significant degree of molecular plasticity and that inter-
molecular interactions are crucial to protein stabilization. These
findings are in line with a dynamically destabilized HAMP domain
(Bordignon et al., 2005; Doebber et al., 2008), which can be influ-
298 Structure 18, 293–300, March 10, 2010 ª2010 Elsevier Ltd All rig
enced by external conditions such as temperature, pH, or salt
concentration. Indeed, residual ssNMR signals indicative of a
small population of NpSRII, which lacks a direct contact to the
transducer involving its EF loop as well as an increased amount
of random coil chemical shifts in NpHtrII found in our study,
can be most easily explained by the presence of a minor fraction
of dynamic destabilized HAMP1 domains. However, under the
conditions applied in this study the HAMP1 domain of NpHtrII
in the complex state predominantly adopts a well-ordered struc-
ture reminiscent of the cHAMP segments discussed in Bordignon
et al. (2005) and Doebber et al. (2008) and builds a direct interface
contact to the EF loop region.
According to our ssNMR data, mutation of the Asp75 coun-
terion to uncharged Asn is characterized by only subtle changes
in close vicinity of the retinylidene Schiff base. The hydrogen
bonding network rearranges such that interactions between
Asn75 and Thr89 no longer exist, enabling a dynamical change
in helix F in accordance to recent EPR data (J. Holterhues,
E. Bordignon, J.P. Klare, S.M., M.E., and H.-J. Steinhoff, unpub-
lished data). Two-dimensional ssNMR correlations confirm that
light excitation of the receptor-transducer complex affects the
Schiff base environment and suggest, similarly to the neutraliza-
tion of the counter-ion, a disruption of the hydrogen bonding net-
work. Generally, a comparison of ground-state chemical shifts
with those of the activated state are in line with a conformational
change residing in helix F, as was also deduced from EPR data.
However, the rather limited number of light-induced chemical
shift perturbations speaks against large structural rearrange-
ments in NpSRII and suggests that restricted interactions,
most likely induced by steric contacts of the isomerized retinal,
characterize the active state. Interestingly, perturbations were
mainly identified in the plane determined by Tyr199NpSRII and
hts reserved

Structure
SRII/HtrII Complex Formation and Activation
Asn74NpHtrII with Ala172-Ile173 on helix F and Gly130-Ala131
on helix E. Previous work already pointed out the importance
of the hydrogen bond between Tyr174 and Thr204 for signal
transduction (Sudo and Spudich, 2006). These protein residues
are located in the same plane, suggesting that the structural
modifications in the helix E-F interface identified here are directly
coupled to the helix F-G interface. Structural alterations around
Thr204 would also nicely explain Thr Cb-Cg chemical shift
perturbations observed in our ssNMR study after light activation
(Figure S7H). A flab-like motion of helix F would open the inter-
face between transducer and receptor, thereby inducing a rear-
rangement around Tyr199NpSRII and Asn74NpHtrII (Figure 4C).
Our findings are in remarkable agreement with recent FTIR
studies (Bergo et al., 2009) and strongly suggest a signal trans-
duction mechanism in which a TM2 rotational motion would
result from a coupled movement of helices F and G without sepa-
ration of an extended SRII/HtrII binding mode. Chimera studies
and deletion analysis showed that the EF loop plays a passive
role in signaling (Sasaki et al., 2007). These findings are in line
with our ssNMR results that do not detect any structural modifi-
cations in the EF loop after light activation, although this spectral
region is well resolved.
Interestingly, the protein region identified here as central for
SR activation also plays a crucial role in SRII/HtrII signaling
(Sudo et al., 2006) and in converting bacteriorhodopsin, another
member of the rhodopsin family, into an efficient photosignaler
after complexation with HtrII (Sudo and Spudich, 2006). These
findings suggest an efficient yet remarkably adaptable mecha-
nism that allows transport proteins of the rhodopsin family to
act as signaling units by using a central functional unit [steric
trigger (Sasaki and Spudich, 2008)] that is located close to the
retinal and by creating a tight protein-protein network that
insures a rapid and strong signal transmission to the cytoplasmic
exterior. In this process, our observation of an intrinsically
dynamic HAMP1 domain may be critical to facilitate the subse-
quent stages of signal transmission.
EXPERIMENTAL PROCEDURES
Sample Preparation
Fully uniformly (13C,15N) labeled NpSRII and NpHtrII analogs were expressed
in E. Coli grown on 13C-glucose and 15N NH4Cl. Reversely labeled samples
were obtained by adding amino acids of natural 13C and 15N isotope abun-
dance to the expression medium at concentrations of 670 mg/l for each amino
acid. Proteoliposomes were prepared as described earlier (Klare et al., 2006)
and have previously been shown to be photoactive and to contain 2:2 SRII/
HtrII complexes. (Wegener et al., 2001).
Solid-State NMR Experiments and Analysis
Samples were packed in standard 4 mm ZrO2 rotors with top spacer and Kel-F
caps. Experiments were carried out using (1H, 13C, and 15N) MAS probes at
600 to 850 MHz 1H Larmor frequency (Bruker Biospin). Magic Angle Spinning
up to urot/2p = 11 kHz was performed using low-temperature VT gas chillers at
temperatures ranging from 228 K (freeze-trapped state) to 278 K. SsNMR
studies of HAMP1 and TrC additionally used 3.2 mm rotors spinning at angular
frequencies of up to 19 kHz at 275 to 283 K.
For dipolar correlation spectra, SPC5 (Hohwy et al., 1999) recoupling, PDSD
(Bloembergen, 1949), and variants thereof (Seidel et al., 2004) were used.
Through-bond transfer involved pulse schemes described in Andronesi et al.
(2005). During dipolar and scalar-based correlation experiments, SPINAL64
(Fung et al., 2000) and GARP (Shaka et al., 1985) decoupling were applied at
83 kHz and 10 kHz, respectively. Light excitation and freeze trapping was per-
Structure 18, 2
formed within a custom-built temperature containment providing two liquid
nitrogen-cooled temperature zones of approximately 270 K and below 173 K.
To increase the light-accessible surface the sample was frozen and ground
before excitation. Excitation itself was carried out at the high temperature
zone using a projector with a color filter as light source. Excited sample was
freeze trapped using liquid nitrogen and MAS rotors were packed at the low
temperature stage, prior to transfer into the pre-cooled wide-bore 600 MHz
magnet. Two-dimensional spectral analysis was done in Sparky 3.110
(Goddard and Kneller, 2008). Secondary chemical shifts DDd = (dCAexp �
dCAav) � (dCB
exp � dCBav) (Luca et al., 2001) were calculated using average
chemical shifts from the BioMagResBank.
Prediction of Protein Interface
A Mathematica (Wolfram Research) script was used to generate a predicted
spectrum based on proton-proton distances measured in the X-ray structure.
To incorporate an additional relay step, the range of potential magnetization
transfer was set to 7 A and a detailed analysis between the peak amplitudes
and the corresponding distance was not attempted. Instead, based on the
list of possible excited frequencies in the receptor, a superposition of the
corresponding Lorentzian functions was fitted to the experimental data.
Sequence Alignment, Modeling, and Spectral Prediction
Sequence alignment of NpHtrIIG84M-136 to Af1503 (Ser278 to Glu331) was
performed using ClustalW2 (Larkin et al., 2007). Modeling of the NpSRII-His/
NpHtrII157-His complex was done with the crystallography and NMR system
(Brunger et al., 1998), using several refinement rounds with a standard simu-
lated annealing protocol, and a modified version allowing for symmetry
distance restraints. Distance and dihedral torsion angle restraints obtained
from Protein Data Bank (PDB) entry 1H2S were used to restrain the crystal
structure part of the model. Distance restraints obtained from the Af1503
structure 2ASW were used to build a homology model for the HAMP domain
by constraining backbone and Cb nuclei. Additional distance constraints were
used to assure a rotational symmetry of the dimeric complex and to assist the
arrangement of the domains. HADDOCK-like ambiguous interaction restraints
of Ser154 with transducer residues were used to close up the interface
between NpSRII and the HAMP domain (Supplemental Information).
SUPPLEMENTAL INFORMATION
Supplemental Information includes seven figures and Supplemental Refer-
ences and can be found with this article online at doi:10.1016/j.str.2010.01.011.
ACKNOWLEDGMENTS
Technical assistance by B. Angerstein is gratefully acknowledged. We thank
R. Schneider for the simulation of double-quantum buildup efficiencies. This
work was funded by Max-Planck-Gesellschaft and Deutsche Forschungsge-
meinschaft (M.E.).
Received: October 3, 2009
Revised: December 23, 2009
Accepted: January 22, 2010
Published: March 9, 2010
REFERENCES
Ader, C., Schneider, R., Hornig, S., Velisetty, P., Wilson, E.M., Lange, A., Giller,
K., Ohmert, I., Martin-Eauclaire, M.-F., Trauner, D., et al. (2008). A structural
link between inactivation and block of a K+ channel. Nat. Struct. Mol. Biol.
15, 605–612.
Ader, C., Schneider, R., Hornig, S., Velisetty, P., Vardanyan, V., Giller, K.,
Ohmert, I., Becker, S., Pongs, O., and Baldus, M. (2009). Coupling of activation
and inactivation gate in a K+-channel: potassium and ligand sensitivity. EMBO
J. 28, 2825–2834.
Ahuja, S., Hornak, V., Yan, E.C.Y., Syrett, N., Goncalves, J.A., Hirshfeld, A.,
Ziliox, M., Sakmar, T.P., Sheves, M., Reeves, P.J., et al. (2009). Helix
93–300, March 10, 2010 ª2010 Elsevier Ltd All rights reserved 299

Structure
SRII/HtrII Complex Formation and Activation
movement is coupled to displacement of the second extracellular loop in
rhodopsin activation. Nat. Struct. Mol. Biol. 16, 168–175.
Andronesi, O.C., Becker, S., Seidel, K., Heise, H., Young, H.S., and Baldus, M.
(2005). Determination of membrane protein structure and dynamics by magic-
angle-spinning solid-state NMR spectroscopy. J. Am. Chem. Soc. 127,
12965–12974.
Bergo, V.B., Spudich, E.N., Spudich, J.L., and Rothschild, K.J. (2009). Active
water in protein-protein communication within the membrane: the case of
SRII-HtrII signal relay. Biochemistry 48, 811–813.
Bloembergen, N. (1949). On the interaction of nuclear spins in a crystalline
lattice. Physica 15, 386–426.
Bordignon, E., Klare, J.P., Doebber, M., Wegener, A.A., Martell, S., Engelhard,
M., and Steinhoff, H.-J. (2005). Structural analysis of a HAMP domain: the
linker region of the phototransducer in complex with sensory rhodopsin II. J.
Biol. Chem. 280, 38767–38775.
Bordignon, E., Klare, J.P., Holterhues, J., Martell, S., Krasnaberski, A., Engel-
hard, M., and Steinhoff, H.-J. (2007). Analysis of light-induced conformational
changes of Natronomonas pharaonis sensory rhodopsin II by time resolved
electron paramagnetic resonance spectroscopy. Photochem. Photobiol. 83,
263–272.
Brunger, A.T., Adams, P.D., Clore, G.M., DeLano, W.L., Gros, P., Grosse-
Kunstleve, R.W., Jiang, J.S., Kuszewski, J., Nilges, M., Pannu, N.S., et al.
(1998). Crystallography & NMR system: a new software suite for macromolec-
ular structure determination. Acta Crystallogr. D Biol. Crystallogr. 54, 905–921.
Doebber, M., Bordignon, E., Klare, J.P., Holterhues, J., Martell, S., Mennes, N.,
Li, L., Engelhard, M., and Steinhoff, H.-J. (2008). Salt-driven equilibrium
between two conformations in the HAMP domain from Natronomonas pharao-
nis: the language of signal transfer? J. Biol. Chem. 283, 28691–28701.
Edman, K., Royant, A., Nollert, P., Maxwell, C.A., Pebay-Peyroula, E., Navarro,
J., Neutze, R., and Landau, E.M. (2002). Early structural rearrangements in the
photocycle of an integral membrane sensory receptor. Structure 10, 473–482.
Etzkorn, M., Bockmann, A., Lange, A., and Baldus, M. (2004). Probing molec-
ular interfaces using 2D magic-angle-spinning NMR on protein mixtures with
different uniform labeling. J. Am. Chem. Soc. 126, 14746–14751.
Etzkorn, M., Martell, S., Andronesi, O.C., Seidel, K., Engelhard, M., and
Baldus, M. (2007). Secondary structure, dynamics, and topology of a seven-
helix receptor in native membranes, studied by solid-state NMR spectros-
copy. Angew. Chem. Int. Ed. Engl. 46, 459–462.
Etzkorn, M., Kneuper, H., Dunnwald, P., Vijayan, V., Kramer, J., Griesinger, C.,
Becker, S., Unden, G., and Baldus, M. (2008). Plasticity of the PAS domain and
a potential role for signal transduction in the histidine kinase DcuS. Nat. Struct.
Mol. Biol. 15, 1031–1039.
Fung, B.M., Khitrin, A.K., and Ermolaev, K. (2000). An improved broadband
decoupling sequence for liquid crystals and solids. J. Magn. Reson. 142,
97–101.
Goddard, T.D., and Kneller, D.G. (2008). SPARKY 3. (http://www.cgl.ucsf.edu/
home/sparky/).
Gordeliy, V.I., Labahn, J., Moukhametzianov, R., Efremov, R., Granzin, J.,
Schlesinger, R., Buldt, G., Savopol, T., Scheidig, A.J., Klare, J.P., and Engel-
hard, M. (2002). Molecular basis of transmembrane signalling by sensory
rhodopsin II-transducer complex. Nature 419, 484–487.
Hohwy, M., Rienstra, C.M., Jaroniec, C.P., and Griffin, R.G. (1999). Fivefold
symmetric homonuclear dipolar recoupling in rotating solids: application to
double quantum spectroscopy. J. Chem. Phys. 110, 7983–7992.
Hulko, M., Berndt, F., Gruber, M., Linder, J.U., Truffault, V., Schultz, A., Martin,
J., Schultz, J.E., Lupas, A.N., and Coles, M. (2006). The HAMP domain struc-
ture implies helix rotation in transmembrane signaling. Cell 126, 929–940.
Khursigara, C.M., Wu, X., Zhang, P., Lefman, J., and Subramaniam, S. (2008).
Role of HAMP domains in chemotaxis signaling by bacterial chemoreceptors.
Proc. Natl. Acad. Sci. USA 105, 16555–16560.
Klare, J.P., Gordeliy, V.I., Labahn, J., Buldt, G., Steinhoff, H.J., and Engelhard,
M. (2004). The archaeal sensory rhodopsin II/transducer complex: a model for
transmembrane signal transfer. FEBS Lett. 564, 219–224.
300 Structure 18, 293–300, March 10, 2010 ª2010 Elsevier Ltd All rig
Klare, J.P., Bordignon, E., Doebber, M., Fitter, J., Kriegsmann, J., Chizhov, I.,
Steinhoff, H.J., and Engelhard, M. (2006). Effects of solubilization on the struc-
ture and function of the sensory rhodopsin II/transducer complex. J. Mol. Biol.
356, 1207–1221.
Klare, J.P., Chizhov, I., and Engelhard, M. (2008). Microbial rhodopsins:
scaffolds for ion pumps, channels, and sensors. Results Probl. Cell Differ.
45, 73–122.
Larkin, M.A., Blackshields, G., Brown, N.P., Chenna, R., McGettigan, P.A.,
McWilliam, H., Valentin, F., Wallace, I.M., Wilm, A., Lopez, R., et al. (2007).
Clustal W and clustal X version 2.0. Bioinformatics 23, 2947–2948.
Luca, S., Filippov, D.V., van Boom, J.H., Oschkinat, H., de Groot, H.J.M., and
Baldus, M. (2001). Secondary chemical shifts in immobilized peptides and
proteins: a qualitative basis for structure refinement under magic angle
spinning. J. Biomol. NMR 20, 325–331.
Luecke, H., Schobert, B., Lanyi, J.K., Spudich, E.N., and Spudich, J.L. (2001).
Crystal structure of sensory rhodopsin II at 2.4 angstroms: insights into color
tuning and transducer interaction. Science 293, 1499–1503.
Mak-Jurkauskas, M.L., Bajaj, V.S., Hornstein, M.K., Belenky, M., Griffin, R.G.,
and Herzfeld, J. (2008). Energy transformations early in the bacteriorhodopsin
photocycle revealed by DNP-enhanced solid-state NMR. Proc. Natl. Acad.
Sci. USA 105, 883–888.
Moukhametzianov, R., Klare, J.P., Efremov, R., Baeken, C., Goppner, A.,
Labahn, J., Engelhard, M., Buldt, G., and Gordeliy, V.I. (2006). Development
of the signal in sensory rhodopsin and its transfer to the cognate transducer.
Nature 440, 115–119.
Neal, S., Nip, A.M., Zhang, H.Y., and Wishart, D.S. (2003). Rapid and accurate
calculation of protein H-1, C-13 and N-15 chemical shifts. J. Biomol. NMR 26,
215–240.
Royant, A., Nollert, P., Edman, K., Neutze, R., Landau, E.M., Pebay-Peyroula,
E., and Navarro, J. (2001). X-ray structure of sensory rhodopsin II at 2.1-A reso-
lution. Proc. Natl. Acad. Sci. USA 98, 10131–10136.
Sasaki, J., and Spudich, J.L. (2008). Signal transfer in haloarchaeal sensory
rhodopsin-transducer complexes. Photochem. Photobiol. 84, 863–868.
Sasaki, J., Nara, T., Spudich, E.N., and Spudich, J.L. (2007). Constitutive
activity in chimeras and deletions localize sensory rhodopsin II/HtrII signal
relay to the membrane-inserted domain. Mol. Microbiol. 66, 1321–1330.
Schneider, R., Seidel, K., Etzkorn, M., Lange, A., Becker, S., and Baldus, M.
(2010). Probing molecular motion by double-quantum (13C, 13C) solid-state
NMR spectroscopy: application to ubiquitin. J. Am. Chem. Soc. 132, 223–233.
Seidel, K., Lange, A., Becker, S., Hughes, C.E., Heise, H., and Baldus, M.
(2004). Protein solid-state NMR resonance assignments from (C-13, C-13)
correlation spectroscopy. Phys. Chem. Chem. Phys. 6, 5090–5093.
Seidel, K., Etzkorn, M., Schneider, R., Ader, C., and Baldus, M. (2009).
Comparative analysis of NMR chemical shift predictions for proteins in the
solid phase. Solid State Nucl. Magn. Reson. 35, 235–242.
Shaka, A.J., Barker, P.B., and Freeman, R. (1985). Computer-optimized
decoupling scheme for wideband applications and low-level operation. J.
Magn. Reson. 64, 547–552.
Sudo, Y., and Spudich, J.L. (2006). Three strategically placed hydrogen-
bonding residues convert a proton pump into a sensory receptor. Proc. Natl.
Acad. Sci. USA 103, 16129–16134.
Sudo, Y., Furutani, Y., Kandori, H., and Spudich, J.L. (2006). Functional impor-
tance of the interhelical hydrogen bond between Thr(204) and Tyr(174) of
sensory rhodopsin II and its alteration during the signaling process. J. Biol.
Chem. 281, 34239–34245.
Wegener, A.A., Klare, J.P., Engelhard, M., and Steinhoff, H.J. (2001). Structural
insights into the early steps of receptor-transducer signal transfer in archaeal
phototaxis. EMBO J. 20, 5312–5319.
Yang, C.-S., Sineshchekov, O., Spudich, E.N., and Spudich, J.L. (2004). The
cytoplasmic membrane-proximal domain of the HtrII transducer interacts
with the E-F loop of photoactivated Natronomonas pharaonis sensory
rhodopsin II. J. Biol. Chem. 279, 42970–42976.
hts reserved