Embed Size (px)
Citation preview

Cc
Sa
b
c
d
a
AA
KRRR
1
iowlo(psci2ic
IS
sc
1h
Respiratory Physiology & Neurobiology 185 (2013) 410– 415
Contents lists available at SciVerse ScienceDirect
Respiratory Physiology & Neurobiology
jou rn al h om epa ge: www.elsev ier .com/ locate / resphys io l
ompartmental chest wall volume changes during volitional hyperpnoea withonstant tidal volume in healthy individuals
abine K. Illi a,b, Stefanie Hostettlera,b,c, Andrea Alivertid, Christina M. Spenglera,b,∗
Exercise Physiology, Institute of Human Movement Sciences, ETH Zurich, Winterthurerstrasse 190, 8057 Zurich, SwitzerlandInstitute of Physiology and Zurich Center for Integrative Human Physiology (ZIHP), University of Zurich, Winterthurerstrasse 190, 8057 Zurich, SwitzerlandSwiss Paraplegic Research, Guido A. Zäch Strasse 4, 6207 Nottwil, SwitzerlandTBM Lab, Dipartimento di Bioingegneria, Politecnico di Milano, Piazza Leonardo da Vinci 32, 20133 Milano, Italy
r t i c l e i n f o
rticle history:ccepted 23 August 2012
eywords:espiratory muscle recruitmentespiratory muscle fatigueapid shallow breathing
a b s t r a c t
Prolonged high-intensity ventilation is associated with the development of rapid shallow breathing withdecreased end-inspiratory volumes of all chest wall compartments. During respiratory muscle endurancetraining using normocapnic hyperpnoea, tidal volume (VT) is normally kept constant. The aim of thisstudy was to investigate possible changes in muscle recruitment during constant-VT hyperpnoea, toassess potential mechanisms related to rapid shallow breathing. Ten healthy subjects performed 1 h ofnormocapnic hyperpnoea at 70% of maximal voluntary ventilation. Chest wall volume changes were
assessed by optoelectronic plethysmography. End-inspiratory (1.08 ± 0.18 versus 0.96 ± 0.27 l, p = 0.017)and end-expiratory volumes (−0.13 ± 0.15 versus −0.31 ± 0.19 l, p = 0.007) of the pulmonary ribcagedecreased significantly and lung function and respiratory muscle strength were reduced (all p < 0.05).Since with forced, constant VT only the inspiratory rib cage muscles were unable to sustain end-inspiratoryvolume of their compartment, inspiratory rib cage muscles are the most likely candidate responsible forthe development of rapid shallow breathing.. Introduction
It is widely accepted that inspiratory rib cage muscles, such asnspiratory intercostals, but also scalene, sternocleidomastoid, andther muscles that insert into and expand the rib cage, take overork from the fatiguing diaphragm during exercise when venti-
ation increases. This is seen in an over-proportional increase ofesophageal compared to transdiaphragmatic pressure generationJohnson et al., 1993; Mador et al., 1993). Furthermore, an increasederception of respiratory exertion and the adoption of a rapid andhallow breathing pattern are often observed in the course of exer-ise (Kearon et al., 1991), both of which have been ascribed, at leastn part, to rib cage muscle fatigue (Ward et al., 1988; Verges et al.,006). Shallow breathing leads, however, to inefficient ventilation,
.e. increased dead space ventilation, which may compromise exer-ise performance at high intensities.
∗ Corresponding author at: University and ETH Zurich, Exercise Physiology,nstitute of Human Movement Sciences, Winterthurerstrasse 190, 8057 Zurich,witzerland. Tel.: +41 44 635 50 07; fax: +41 44 635 68 14.
E-mail addresses: [email protected] (S.K. Illi),[email protected] (S. Hostettler), [email protected] (A. Aliverti),[email protected] (C.M. Spengler).
569-9048/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.resp.2012.08.018
© 2012 Elsevier B.V. All rights reserved.
Respiratory muscle training can reduce or delay the develop-ment of respiratory muscle fatigue (Romer et al., 2002b; Vergeset al., 2007, 2009). This in turn may prevent the development ofrapid and shallow breathing as shown in some (Spengler et al.,1999; Volianitis et al., 2001; Amonette and Dupler, 2002; Romeret al., 2002a; Wylegala et al., 2007; Esposito et al., 2010) butnot in other (Kohl et al., 1997; Stuessi et al., 2001; Volianitiset al., 2001; McMahon et al., 2002; Holm et al., 2004; Griffithsand McConnell, 2007; Verges et al., 2007; Brown et al., 2010;Ray et al., 2010) studies. To gain further insights into potentialmechanisms of maintaining tidal volume after respiratory muscletraining, we investigated respiratory muscle recruitment during asingle respiratory muscle endurance training session. Normocapnichyperpnoea, which is usually performed for respiratory mus-cle endurance training, predominantly induces diaphragm fatigue(McCool et al., 1992). In a recent study of our laboratory, Renggliet al. (2008) showed an increase in the ratio of oesophageal to trans-diaphragmatic pressure generation during prolonged normocapnichyperpnoea suggesting a partial takeover of diaphragmatic work byinspiratory rib cage muscles, similar to what was observed duringexercise (Johnson et al., 1993; Mador et al., 1993).
Using a different technique to assess respiratory muscle recruit-ment, i.e. optoelectronic plethysmography (OEP), recent studiesin patients with Duchenne muscular dystrophy showed that withprogressive weakness of the diaphragm the relative contribution to

gy & N
tefctub2temamsi2vbt
2htiwcnttdmvt
2
2
rphSmat
TS
VimmW
S.K. Illi et al. / Respiratory Physiolo
idal volume of the abdominal compartment decreases (Lo Maurot al., 2010; Romei et al., 2012). Thus, specific respiratory muscleatigue, i.e. a “transient” weakness for instance of the diaphragm,ould also lead to changes in relative compartmental contributiono tidal volume. However, results of another study of our laboratorysing OEP (Illi et al., 2011), did not confirm the observation madey Renggli et al. during normocapnic hyperpnoea (Renggli et al.,008). Instead, rib cage muscles appeared to fatigue similarly tohe diaphragm, or even earlier, as indicated by the decrease in thend-inspiratory volumes of the corresponding chest wall compart-ents (Illi et al., 2011). These recent findings thus challenge the
ssumption that inspiratory rib cage muscles take over diaphrag-atic work during normocapnic hyperpnoea. However, the two
tudies differed in one important aspect: tidal volume was fixedn one study (Renggli et al., 2008) but not in the other (Illi et al.,011). Thus, the aim of the present study was to assess chest wallolume changes during 1 h of normocapnic hyperpnoea with a fixedreathing pattern similar to the study by Renggli et al. (2008) andhat used during respiratory muscle endurance training sessions.
Based on these previous studies (Renggli et al., 2008; Illi et al.,011), two hypotheses were tested: (1) during prolonged volitionalyperpnoea with fixed tidal volume, inspiratory rib cage musclesake over inspiratory work from the fatiguing diaphragm, reflectedn a change in the relative contribution of the corresponding chest
all compartments to tidal volume, i.e. an increase in upper ribage contribution and a decrease in lower rib cage and abdomi-al contribution; or (2) during prolonged hyperpnoea with fixedidal volume, inspiratory muscle recruitment does not change andhe development of global inspiratory muscle fatigue leads to aecrease in end-inspiratory volumes of all chest wall compart-ents which is either compensated by a decrease in end-expiratory
olumes, i.e. expiratory muscles take over work from the inspira-ory muscles, or results in task failure.
. Methods
.1. Subjects
Ten healthy, non-smoking subjects (3 men, 7 women) wereecruited. All had previously participated in the study on com-artmental chest wall volume changes during normocapnicyperpnoea with unconstrained breathing pattern (Illi et al., 2011).ubjects’ characteristics are given in Table 1. All subjects had nor-
al lung function and respiratory muscle strength. Subjects weresked to refrain from strenuous physical activity for two days prioro test days and from any exercise on test days. Drinking caffeinated
able 1ubjects’ characteristics.
Anthropometric dataAge [y] 36.6 ± 3.7Height [cm] 170.9 ± 8.6Body mass [kg] 63.4 ± 10.4
Lung functionFVC [% pred] 116 ± 14FEV1 [% pred] 111 ± 11PEF [% pred] 116 ± 17FIV1 [% pred] 120 ± 16MVV [% pred] 131 ± 18
Respiratory muscle strengthMIP [% pred] 137 ± 36MEP [% pred] 138 ± 47
alues are means ± SD. FVC: forced vital capacity; FEV1: forced expiratory volumen 1 s; PEF: peak expiratory flow rate; FIV1: forced inspiratory volume in 1 s; MVV:
aximal voluntary ventilation; MIP: maximal inspiratory mouth pressure; MEP:aximal expiratory mouth pressure; % pred: % predicted, reference values are fromilson et al., 1984, and Quanjer et al., 1993.
eurobiology 185 (2013) 410– 415 411
beverages on test days prior to testing was prohibited, as was foodintake 2 h before testing.
Subjects were provided with detailed information about thetesting procedures and then gave their written informed consent toparticipate. The study was approved by the local ethics committeeand performed according to the Declaration of Helsinki.
2.2. Protocol overview
Subjects reported to the laboratory on two different occasions,separated by at least 72 h and scheduled at the same time of day.The first session consisted of lung function and respiratory musclestrength measurements, as well as familiarization of subjects withthe breathing device for normocapnic hyperpnoea. In the secondtest session, lung function (forced spirometry) and respiratory mus-cle strength were assessed, followed by 5 min of quiet breathingand then 1 h of voluntary normocapnic hyperpnoea with a constantbreathing pattern. Immediately after, lung function and respiratorymuscle strength measurements were repeated, in the same orderas prior to hyperpnoea.
2.3. Lung function and respiratory muscle strength measurements
Lung function (vital capacity (VC), forced vital capacity (FVC),forced expiratory volume in 1 s (FEV1), peak expiratory flow rate(PEF), forced inspiratory volume in 1 s (FIV1), peak inspiratory flowrate (PIF), and maximal voluntary ventilation in 12 s (12 s-MVV)was assessed according to standard procedures (Miller et al., 2005)using an ergospirometric device (Quark b2, Cosmed, Rome, Italy)with a calibrated turbine for volume measurements.
Respiratory muscle strength measurements (maximal inspi-ratory mouth pressure (MIP) from residual volume, maximalexpiratory mouth pressure (MEP) from total lung capacity) wereconducted according to the current ATS/ERS statement (Greenet al., 2002) using a hand held mouth pressure meter (MicroRPM,MicroMedical, Kent, Great Britain). A minimum of five technically-adequate manoeuvres were performed, selecting the highest ofthree values within no more than 5% variability.
2.4. Normocapnic hyperpnoea
Normocapnic hyperpnoea was performed at a target ventila-tion of 70% of subjects’ individual 12 s-MVV and at a target tidalvolume of 60% of subjects’ VC with target breathing frequency cal-culated accordingly. A self-constructed partial rebreathing devicedescribed in detail elsewhere (Illi et al., 2011) was used to ensurenormocapnia, and breathing frequency was paced by a metronome.Ventilation and end-tidal CO2 partial pressure (PETCO2; assessed bya calibrated infrared absorption gas sensor) were measured con-tinuously (Quark b2). If the breathing pattern deviated from thetarget, subjects received verbal instructions to adjust either tidalvolume or breathing frequency accordingly. In addition, subjectswere asked every 5 min to rate their perceived respiratory exer-tion on a linear scale ranging from 0 (no exertion) to 10 (maximalexertion).
2.5. Optoelectronic plethysmography
Compartmental chest wall volume changes during quiet breath-ing and normocapnic hyperpnoea (all conducted in standing) wereassessed by OEP (BTS, Milan, Italy), a system described in detail byCala et al. (Cala et al., 1996). Briefly, 89 infrared reflective markers
were placed circumferentially on the chest wall between the clavi-cles and the anterior superior iliac spine. Six infrared cameras (threein front and three behind the subject) traced the three-dimensionalcoordinates of these markers at a sampling rate of 60 Hz in order
4 gy & Neurobiology 185 (2013) 410– 415
tloabomcioutAm
2
(odddifvrzt
infsahcu
3
3n
mocimefishtcv
3
g(n
Fig. 1. Relative compartmental contribution to tidal volume. �Vrc,p: relative con-tribution of the pulmonary rib cage to tidal volume; �Vrc,a: relative contribution ofthe abdominal rib cage to tidal volume; �Vab: relative contribution of the abdomento tidal volume; TLC: total lung capacity; RV: residual volume; QB: quiet breathing;start: first 3 min; middle: middle 1 min; end: last 3 min of normocapnic hyperpnoea.#p < 0.05 compared to start; *p < 0.05 compared to middle.
Table 2Absolute values for lung function and respiratory muscle strength measurementspre and post normocapnic hyperpnoea.
pre NH post NH
Lung functionFVC [l] 4.8 ± 0.9 4.7 ± 1.0FEV1 [l] 3.9 ± 0.6 3.8 ± 0.7*PEF [l s−1] 9.2 ± 1.8 8.5 ± 1.8*FIV1 [l] 4.2 ± 0.7 4.0 ± 0.8*PIF [l s−1] 6.8 ± 1.5 6.3 ± 1.6*MVV [l min−1] 164.0 ± 25.7 na
Respiratory muscle strengthMIP [cm H2O] 113 ± 21 102 ± 14*MEP [cm H2O] 151 ± 51 128 ± 41**
Values are means ± SD. NH: normocapnic hyperpnoea; FVC: forced vital capac-ity; FEV1: forced expiratory volume in 1 s; PEF: peak expiratory flow rate; FIV1:forced inspiratory volume in 1 s; PIF: peak inspiratory flow rate; MVV: maximal
12 S.K. Illi et al. / Respiratory Physiolo
o compute the thoracoabdominal volumes using surface triangu-ation. Chest wall volume was modelled as the sum of the volumesf the three compartments, i.e. the pulmonary rib cage (RCp), thebdominal rib cage (RCa), and the abdomen (AB). The boundariesetween RCp and RCa and between RCa and AB were set at the levelf the xiphoid and at the lower costal margin, respectively. Specificechanical properties, including inserting muscles, pressures, and
ompliance of a compartment, influence volume changes. Changesn volume of the pulmonary apposed RCp (�Vrc,p) reflect the actionf the inspiratory and expiratory rib cage muscles, changes in vol-me of the diaphragm apposed RCa (�Vrc,a) reflect the action ofhe diaphragm and abdominal muscles, and volume changes of theB (�Vab) also reflect the action of the diaphragm and abdominaluscles.
.6. Data analysis
OEP data of 1 min of quiet breathing, as well as of the first 3 minstart), the middle 1 min (middle), and the last 3 min (end) of norm-capnic hyperpnoea were analyzed. The following variables wereerived from OEP using a customized software program (Diamov,esigned and developed at the Politecnico di Milano, Dipartimentoi Bioingegneria, Italy): tidal volume, breathing frequency, end-
nspiratory and end-expiratory chest wall volumes (all derivedrom or measured at zero-flow points), as well as compartmentalolumes (given relative to average end-expiratory volume of theespective compartment during quiet breathing, which was set toero), and relative contribution of the different compartments toidal volume.
Data of quiet breathing and of the three time points dur-ng hyperpnoea (start, middle, end) were compared using theon-parametric Friedman test. Where a time effect was found, dif-
erences between time points were assessed using the Wilcoxonigned rank test, which was also applied to compare lung functionnd respiratory muscle strength before versus after normocapnicyperpnoea. Data are presented as mean ± SD. Statistical signifi-ance was accepted if p < 0.05. Statistical analysis was performedsing IBM SPSS Statistics 19.0 (SPSS, Chicago, IL, USA).
. Results
.1. Compartmental chest wall volume changes duringormocapnic hyperpnoea
A significant time-effect was found for the relative compart-ental contribution. As shown in Fig. 1, the relative contribution
f the abdominal compartment to tidal volume decreased signifi-antly from middle to end (p = 0.009) while the contribution of RCpncreased (p = 0.017). End-inspiratory and end-expiratory compart-
ental volumes decreased significantly in RCp (Fig. 2). The mainnd-inspiratory and end-expiratory decreases occurred within therst half of the test (p = 0.013 and 0.037, respectively). In threeubjects, breathing frequency was reduced during normocapnicyperpnoea (after 9, 25, and 30 min) as they would have reachedask failure otherwise. Their end-inspiratory and end-expiratoryompartmental volumes were included in the analysis since tidalolume was kept constant.
.2. Lung function and respiratory muscle strength
In Table 2, lung function and respiratory muscle strength areiven before and after normocapnic hyperpnoea. All variablesexcept FVC) decreased significantly after normocapnic hyperp-oea.
voluntary ventilation; MIP: maximal inspiratory mouth pressure; MEP: maximalexpiratory mouth pressure; na: not available. Significances are indicated for differ-ences between absolute values pre versus post NH. *p < 0.05, **p < 0.01.
3.3. Respiratory exertion, target ventilation, and end-tidal CO2partial pressure
Perceived respiratory exertion increased significantly duringnormocapnic hyperpnoea (start: 4.0 ± 2.6, middle: 6.1 ± 2.4, end:7.3 ± 1.8; all p < 0.01). Average ventilation (start: 108 ± 17 l min−1,middle: 106 ± 21 l min−1, end: 107 ± 21 l min−1; p = 0.741), targettidal volume (start: 2.8 ± 0.6 l, middle: 2.8 ± 0.7 l, end: 2.8 ± 0.7 l;p = 0.273) and breathing frequency (start: 40 ± 5 min−1, middle:39 ± 6 min−1, end: 39 ± 7 min−1; p = 0.497) remained constant,and PETCO2 was kept normocapnic (start: 38 ± 3 mmHg, middle:39 ± 4 mmHg, end: 39 ± 3 mmHg; p = 0.122).
4. Discussion
The present investigation of subjects performing 1 h of norm-ocapnic hyperpnoea with a constant breathing pattern suggests
that inspiratory rib cage muscles do not take over inspiratorywork from the diaphragm. Although the relative contribution ofRCp to tidal volume increased, end-inspiratory Vrc,p decreased,suggesting additional expiratory adaptations. This compensation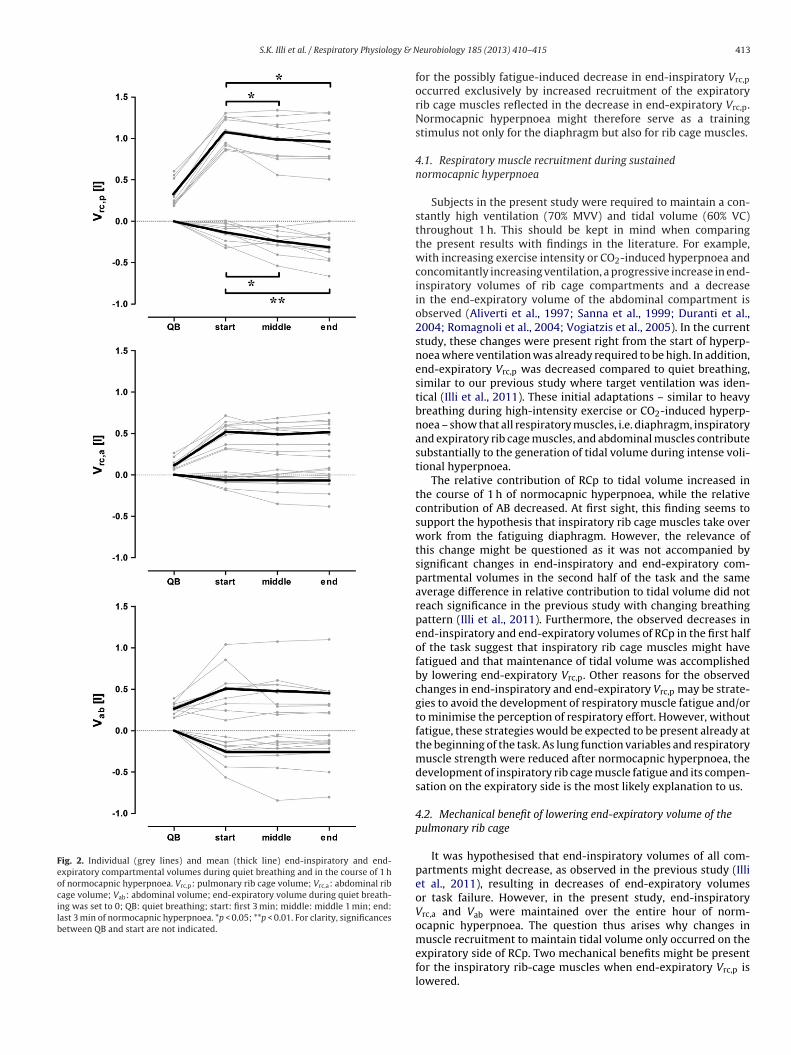
S.K. Illi et al. / Respiratory Physiology & N
Fig. 2. Individual (grey lines) and mean (thick line) end-inspiratory and end-expiratory compartmental volumes during quiet breathing and in the course of 1 hof normocapnic hyperpnoea. Vrc,p: pulmonary rib cage volume; Vrc,a: abdominal ribcage volume; Vab: abdominal volume; end-expiratory volume during quiet breath-ing was set to 0; QB: quiet breathing; start: first 3 min; middle: middle 1 min; end:last 3 min of normocapnic hyperpnoea. *p < 0.05; **p < 0.01. For clarity, significancesbetween QB and start are not indicated.
eurobiology 185 (2013) 410– 415 413
for the possibly fatigue-induced decrease in end-inspiratory Vrc,p
occurred exclusively by increased recruitment of the expiratoryrib cage muscles reflected in the decrease in end-expiratory Vrc,p.Normocapnic hyperpnoea might therefore serve as a trainingstimulus not only for the diaphragm but also for rib cage muscles.
4.1. Respiratory muscle recruitment during sustainednormocapnic hyperpnoea
Subjects in the present study were required to maintain a con-stantly high ventilation (70% MVV) and tidal volume (60% VC)throughout 1 h. This should be kept in mind when comparingthe present results with findings in the literature. For example,with increasing exercise intensity or CO2-induced hyperpnoea andconcomitantly increasing ventilation, a progressive increase in end-inspiratory volumes of rib cage compartments and a decreasein the end-expiratory volume of the abdominal compartment isobserved (Aliverti et al., 1997; Sanna et al., 1999; Duranti et al.,2004; Romagnoli et al., 2004; Vogiatzis et al., 2005). In the currentstudy, these changes were present right from the start of hyperp-noea where ventilation was already required to be high. In addition,end-expiratory Vrc,p was decreased compared to quiet breathing,similar to our previous study where target ventilation was iden-tical (Illi et al., 2011). These initial adaptations – similar to heavybreathing during high-intensity exercise or CO2-induced hyperp-noea – show that all respiratory muscles, i.e. diaphragm, inspiratoryand expiratory rib cage muscles, and abdominal muscles contributesubstantially to the generation of tidal volume during intense voli-tional hyperpnoea.
The relative contribution of RCp to tidal volume increased inthe course of 1 h of normocapnic hyperpnoea, while the relativecontribution of AB decreased. At first sight, this finding seems tosupport the hypothesis that inspiratory rib cage muscles take overwork from the fatiguing diaphragm. However, the relevance ofthis change might be questioned as it was not accompanied bysignificant changes in end-inspiratory and end-expiratory com-partmental volumes in the second half of the task and the sameaverage difference in relative contribution to tidal volume did notreach significance in the previous study with changing breathingpattern (Illi et al., 2011). Furthermore, the observed decreases inend-inspiratory and end-expiratory volumes of RCp in the first halfof the task suggest that inspiratory rib cage muscles might havefatigued and that maintenance of tidal volume was accomplishedby lowering end-expiratory Vrc,p. Other reasons for the observedchanges in end-inspiratory and end-expiratory Vrc,p may be strate-gies to avoid the development of respiratory muscle fatigue and/orto minimise the perception of respiratory effort. However, withoutfatigue, these strategies would be expected to be present already atthe beginning of the task. As lung function variables and respiratorymuscle strength were reduced after normocapnic hyperpnoea, thedevelopment of inspiratory rib cage muscle fatigue and its compen-sation on the expiratory side is the most likely explanation to us.
4.2. Mechanical benefit of lowering end-expiratory volume of thepulmonary rib cage
It was hypothesised that end-inspiratory volumes of all com-partments might decrease, as observed in the previous study (Illiet al., 2011), resulting in decreases of end-expiratory volumesor task failure. However, in the present study, end-inspiratoryVrc,a and Vab were maintained over the entire hour of norm-ocapnic hyperpnoea. The question thus arises why changes in
muscle recruitment to maintain tidal volume only occurred on theexpiratory side of RCp. Two mechanical benefits might be presentfor the inspiratory rib-cage muscles when end-expiratory Vrc,p islowered.
4 gy & N
(
4
tHvspTotoos
cwbwa1itecsm
tboiowtd
14 S.K. Illi et al. / Respiratory Physiolo
(i) Although more respiratory muscle work is required to lowerthe end-expiratory volume of the rib cage than that of theabdomen, since rib cage compliance decreases substantiallybelow functional residual capacity (Grimby et al., 1968) andinspiratory rib cage muscles develop more passive tension thanthe diaphragm when stretched beyond the optimal length foractive force development (Farkas et al., 1985), this work per-formed during expiration is stored as elastic energy and can beretrieved during inspiration in the form of passive recoil of therib cage and inspiratory rib cage muscles.
ii) The capacity of the inspiratory muscles to generate pressure isdiminished at increased shortening velocities (i.e. flows) andsuboptimal length for tension development (i.e. high lung vol-umes) and might even approach its maximum during intenseexercise and/or when respiratory muscle fatigue develops(Leblanc et al., 1988; Johnson et al., 1992). Improvements in thepressure-generating capacity of inspiratory rib cage musclescould thus be achieved by decreasing either shortening veloc-ity or operating lung volume. To avoid task failure in face of therequirement to maintain tidal volume and despite the devel-opment of inspiratory muscle fatigue, subjects needed to lowertheir operating lung volume. The present data suggest thatthis lowering of end-expiratory volume took place exclusivelyin RCp. Since inspiratory rib cage muscles are predominantlyresponsible for rib cage expanding forces while the diaphragmacts as the main flow generator (Aliverti et al., 1997), it seemsintuitive to decrease end-expiratory Vrc,p further in order toimprove the pressure-generating capacity of the inspiratory ribcage muscles.
.3. Implications on exercise hyperpnoea
In the present study seven out of ten subjects were able to main-ain ventilation at 70% MVV with tidal volume fixed to 60% VC.owever, all of these ten subjects spontaneously decreased tidalolume and increased breathing frequency while maintaining theame constant target ventilation during 1 h of normocapnic hyper-noea when breathing pattern was not imposed (Illi et al., 2011).hus, the question arises, why rapid and shallow breathing devel-ped although most of the subjects would have been able to main-ain a constant tidal volume if required to do so. This question isf particular interest for subjects exercising at very high intensitiesr at altitude where increased dead space ventilation during rapidhallow breathing may further compromise blood oxygen content.
In fact, lowering end-expiratory Vrc,p might be even more ineffi-ient than the development of rapid and shallow breathing, whichould be supported by the fact that lowering end-expiratory Vrc,p
elow functional residual capacity is associated with respiratoryork against a decreased rib cage compliance (Grimby et al., 1968)
nd increased passive tension in the rib cage muscles (Farkas et al.,985). Another reason for the development of rapid shallow breath-
ng might be an increased perception of respiratory effort whenidal volume was required to be held constant. Interestingly, how-ver, perception of respiratory effort was not different betweenonditions in the present and previous (Illi et al., 2011) study. Thisuggests that the development of rapid shallow breathing does notinimize the perception of adverse respiratory sensations.However, the development of respiratory muscle fatigue, in par-
icular of rib cage muscle fatigue (Verges et al., 2006), could haveeen responsible for the development of rapid shallow breathing inur previous study (Illi et al., 2011). Considering decreases in end-nspiratory Vrc,p, it seems that inspiratory rib cage muscle fatigue
ccurred early during the one hour of normocapnic hyperpnoeahich would in turn suggest that respiratory muscle training inhe form of normocapnic hyperpnoea would not only train theiaphragm but also the other respiratory muscles, in particular the
eurobiology 185 (2013) 410– 415
rib cage muscles. This assumption is further supported by the find-ings of Verges et al. (2009), who showed a significantly smallerdecrease in twitch oesophageal pressure during and after normo-capnic hyperpnoea after a period of respiratory muscle endurancetraining, suggesting reduced development of inspiratory rib cagemuscle fatigue. Future studies should thus assess compartmentalchest wall volume changes during normocapnic hyperpnoea andhigh-intensity exercise before and after a period of respiratory mus-cle endurance training to investigate whether or not this decreasein the development of contractile fatigue of the inspiratory rib cagemuscles affects chest wall kinematics and the ability to maintainend-inspiratory Vrc,p.
4.4. Limitations
Respiratory muscle fatigue was assessed by voluntary manoeu-vers rather than pressure measurements during phrenic nervestimulation. In addition, assessment of the development ofoesophageal and transdiaphragmatic pressures over the course ofnormocapnic hyperpnoea would be helpful to explain the discrep-ancies between the interpretation of studies calculating the ratioof the pressure-time products and the present results using chestwall volume changes. However, abstaining from invasive pressuremeasurements in the present study ensured undisturbed respira-tory muscle recruitment. Although volitional measures of fatiguemay be confounded by the presence of central fatigue, the decreasein end-inspiratory Vrc,p with no change in the other two compart-ments is considered an independent indicator of inspiratory ribcage muscle fatigue. Furthermore, the few studies assessing respi-ratory muscle fatigue after exercise with both measurement ofoesophageal and gastric pressure during nerve stimulation and dur-ing voluntary respiratory manoeuvres show a decrease in twitchpressures with smaller decreases (or no significant change) inpressures during voluntary manoeuvres (Johnson et al., 1993;Mador et al., 1993; Babcock et al., 1995a,b; Taylor et al., 2006; Kabitzet al., 2008; Taylor and Romer, 2008).
Moreover, sample size of the present study was rather low.However, post hoc power analysis revealed that in most cases,power was close to or above 80% (relative compartmental con-tributions: RCp 70%, AB 91%; end-inspiratory volume of RCp: 77%;end-expiratory volume of RCp: 80%).
Finally, there might be some concerns about whether or not thethree subjects reaching task failure before 1 h of hyperpnoea wascompleted may have influenced the results. For the following rea-sons, we are confident that this was not the case: (i) these threesubjects showed changes in relative compartmental contributionthat were similar to those of the seven subjects that were able tofulfil the task; (ii) end-inspiratory Vrc,p decreased in one subjectwhile in two subjects it did not change; (iii) end-expiratory Vrc,p
decreased in two subjects and increased in one subject.
4.5. Conclusion
In conclusion, during 1 h of normocapnic hyperpnoea witha constant breathing pattern, inspiratory rib cage muscles wereunable to sustain end-inspiratory Vrc,p. Expiratory rib cage mus-cles were increasingly recruited, lowering end-expiratory Vrc,p,thereby allowing an increased pressure-generating capacity of theinspiratory rib cage muscles. This in turn suggests that inspiratoryrib cage muscles play an important role in the development ofrapid shallow breathing when subjects are not forced to keep tidalvolume constant. If inspiratory rib cage muscle fatigue were the
reason for the lowering of end-inspiratory Vrc,p, a period of respira-tory muscle endurance training at a constant and high tidal volumecould delay or prevent the development of inspiratory rib cagemuscle fatigue and hence the onset of rapid and shallow breathing.
gy & N
D
sPi
A
ittTim
R
A
A
B
B
B
C
D
E
F
G
G
G
H
I
J
J
K
K
K
L
L
S.K. Illi et al. / Respiratory Physiolo
isclosure statement
Prof. A. Aliverti is a co-inventor of the Optoelectronic Plethy-mography system. Its patent rights are held by his Institution, theolitecnico di Milano, Italy. All other authors have no conflict ofnterest to disclose.
cknowledgements
The authors thank Dr. E. Mohler and Dr. D. Notter for intellectualnput and technical support, Dr. R. Briggs for English-editing, andhe subjects for their time and effort. The study was supported byhe Swiss National Science Foundation (SNF grant no. 32-116777).he SNF was not involved in study design, collection, analysis andnterpretation of data, writing of the report, or the decision to sub-
it the paper for publication.
eferences
liverti, A., Cala, S.J., Duranti, R., Ferrigno, G., Kenyon, C.M., Pedotti, A., Scano, G.,Sliwinski, P., Macklem, P.T., Yan, S., 1997. Human respiratory muscle actionsand control during exercise. Journal of Applied Physiology 83 (4), 1256–1269.
monette, W.E., Dupler, T.L., 2002. The effects of respiratory muscle training onVO2max, the ventilatory threshold and pulmonary function. JEPonline 5 (2),29–35.
abcock, M.A., Johnson, B.D., Pegelow, D.F., Suman, O.E., Griffin, D., Dempsey, J.A.,1995a. Hypoxic effects on exercise-induced diaphragmatic fatigue in normalhealthy humans. Journal of Applied Physiology 78 (1), 82–92.
abcock, M.A., Pegelow, D.F., McClaran, S.R., Suman, O.E., Dempsey, J.A., 1995b.Contribution of diaphragmatic power output to exercise-induced diaphragmfatigue. Journal of Applied Physiology 78 (5), 1710–1719.
rown, P.I., Sharpe, G.R., Johnson, M.A., 2010. Loading of trained inspiratory musclesspeeds lactate recovery kinetics. Medicine and Science in Sports and Exercise 42(6), 1103–1112.
ala, S.J., Kenyon, C.M., Ferrigno, G., Carnevali, P., Aliverti, A., Pedotti, A., Macklem,P.T., Rochester, D.F., 1996. Chest wall and lung volume estimation by opticalreflectance motion analysis. Journal of Applied Physiology 81 (6), 2680–2689.
uranti, R., Sanna, A., Romagnoli, I., Nerini, M., Gigliotti, F., Ambrosino, N., Scano, G.,2004. Walking modality affects respiratory muscle action and contribution torespiratory effort. Pflugers Archiv 448 (2), 222–230.
sposito, F., Limonta, E., Alberti, G., Veicsteinas, A., Ferretti, G., 2010. Effect of respi-ratory muscle training on maximum aerobic power in normoxia and hypoxia.Respiratory Physiology and Neurobiology 170 (3), 268–272.
arkas, G.A., Decramer, M., Rochester, D.F., De Troyer, A., 1985. Contractile proper-ties of intercostal muscles and their functional significance. Journal of AppliedPhysiology 59 (2), 528–535.
reen, M., Road, J., Sieck, G.C., Similowski, T., 2002. ATS/ERS statement on respiratorymuscle testing. American Journal of Respiratory and Critical Care Medicine 166(4), 528–547.
riffiths, L.A., McConnell, A.K., 2007. The influence of inspiratory and expiratorymuscle training upon rowing performance. European Journal of Applied Physi-ology 99 (5), 457–466.
rimby, G., Bunn, J., Mead, J., 1968. Relative contribution of rib cage and abdomento ventilation during exercise. Journal of Applied Physiology 24 (2), 159–166.
olm, P., Sattler, A., Fregosi, R.F., 2004. Endurance training of respiratory musclesimproves cycling performance in fit young cyclists. BMC Physiology 4, 9.
lli, S.K., Hostettler, S., Mohler, E., Aliverti, A., Spengler, C.M., 2011. Compartmentalchest wall volume changes during volitional normocapnic hyperpnoea. Respi-ratory Physiology and Neurobiology 177 (3), 294–300.
ohnson, B.D., Babcock, M.A., Suman, O.E., Dempsey, J.A., 1993. Exercise-induceddiaphragmatic fatigue in healthy humans. Journal of Physiology 460, 385–405.
ohnson, B.D., Saupe, K.W., Dempsey, J.A., 1992. Mechanical constraints on exercisehyperpnea in endurance athletes. Journal of Applied Physiology 73 (3), 874–886.
abitz, H.J., Walker, D., Prettin, S., Walterspacher, S., Sonntag, F., Dreher, M.,Windisch, W., 2008. Non-invasive ventilation applied for recovery fromexercise-induced diaphragmatic fatigue. Open Respiratory Medicine Journal 2,16–21.
earon, M.C., Summers, E., Jones, N.L., Campbell, E.J., Killian, K.J., 1991. Breathingduring prolonged exercise in humans. Journal of Physiology 442, 477–487.
ohl, J., Koller, E.A., Brandenberger, M., Cardenas, M., Boutellier, U., 1997. Effect ofexercise-induced hyperventilation on airway resistance and cycling endurance.European Journal of Applied Physiology and Occupational Physiology 75 (4),305–311.
eblanc, P., Summers, E., Inman, M.D., Jones, N.L., Campbell, E.J., Killian, K.J., 1988.Inspiratory muscles during exercise: a problem of supply and demand. Journalof Applied Physiology 64 (6), 2482–2489.
o Mauro, A., D’Angelo, M.G., Romei, M., Motta, F., Colombo, D., Comi, G.P., Pedotti,A., Marchi, E., Turconi, A.C., Bresolin, N., Aliverti, A., 2010. Abdominal volume
eurobiology 185 (2013) 410– 415 415
contribution to tidal volume as an early indicator of respiratory impair-ment in Duchenne muscular dystrophy. European Respiratory Journal 35 (5),1118–1125.
Mador, M.J., Magalang, U.J., Rodis, A., Kufel, T.J., 1993. Diaphragmatic fatigue afterexercise in healthy human subjects. American Review of Respiratory Disease148 (6 Pt 1), 1571–1575.
McCool, F.D., Hershenson, M.B., Tzelepis, G.E., Kikuchi, Y., Leith, D.E., 1992. Effectof fatigue on maximal inspiratory pressure-flow capacity. Journal of AppliedPhysiology 73 (1), 36–43.
McMahon, M.E., Boutellier, U., Smith, R.M., Spengler, C.M., 2002. Hyperpnea train-ing attenuates peripheral chemosensitivity and improves cycling endurance.Journal of Experimental Biology 205 (Pt 24), 3937–3943.
Miller, M.R., Hankinson, J., Brusasco, V., Burgos, F., Casaburi, R., Coates, A., Crapo, R.,Enright, P., van der Grinten, C.P., Gustafsson, P., Jensen, R., Johnson, D.C., MacIn-tyre, N., McKay, R., Navajas, D., Pedersen, O.F., Pellegrino, R., Viegi, G., Wanger,J., 2005. Standardisation of spirometry. European Respiratory Journal 26 (2),319–338.
Quanjer, P.H., Tammeling, G.J., Cotes, J.E., Pedersen, O.F., Peslin, R., Yernault, J.C.,1993. Lung volumes and forced ventilatory flows. Report Working Party Stan-dardization of Lung Function Tests, European Community for Steel and Coal.Official Statement of the European Respiratory Society. European RespiratoryJournal 16, Supplement 5–40.
Ray, A.D., Pendergast, D.R., Lundgren, C.E., 2010. Respiratory muscle training reducesthe work of breathing at depth. European Journal of Applied Physiology 108 (4),811–820.
Renggli, A.S., Verges, S., Notter, D.A., Spengler, C.M., 2008. Development of respi-ratory muscle contractile fatigue in the course of hyperpnoea. RespiratoryPhysiology and Neurobiology 164 (3), 366–372.
Romagnoli, I., Gigliotti, F., Lanini, B., Bianchi, R., Soldani, N., Nerini, M., Duranti, R.,Scano, G., 2004. Chest wall kinematics and respiratory muscle coordinated actionduring hypercapnia in healthy males. European Journal of Applied Physiology91 (5–6), 525–533.
Romei, M., D’Angelo, M.G., Lomauro, A., Gandossini, S., Bonato, S., Brighina, E., Marchi,E., Comi, G.P., Turconi, A.C., Pedotti, A., Bresolin, N., Aliverti, A., 2012. Low abdom-inal contribution to breathing as daytime predictor of nocturnal desaturation inadolescents and young adults with Duchenne muscular dystrophy. RespiratoryMedicine 106 (2), 276–283.
Romer, L.M., McConnell, A.K., Jones, D.A., 2002a. Effects of inspiratory muscle train-ing on time-trial performance in trained cyclists. Journal of Sports Sciences 20(7), 547–562.
Romer, L.M., McConnell, A.K., Jones, D.A., 2002b. Inspiratory muscle fatigue in trainedcyclists: effects of inspiratory muscle training. Medicine and Science in Sportsand Exercise 34 (5), 785–792.
Sanna, A., Bertoli, F., Misuri, G., Gigliotti, F., Iandelli, I., Mancini, M., Duranti, R.,Ambrosino, N., Scano, G., 1999. Chest wall kinematics and respiratory mus-cle action in walking healthy humans. Journal of Applied Physiology 87 (3),938–946.
Spengler, C.M., Roos, M., Laube, S.M., Boutellier, U., 1999. Decreased exercise bloodlactate concentrations after respiratory endurance training in humans. EuropeanJournal of Applied Physiology and Occupational Physiology 79 (4), 299–305.
Stuessi, C., Spengler, C.M., Knopfli-Lenzin, C., Markov, G., Boutellier, U., 2001. Respira-tory muscle endurance training in humans increases cycling endurance withoutaffecting blood gas concentrations. European Journal of Applied Physiology 84(6), 582–586.
Taylor, B.J., How, S.C., Romer, L.M., 2006. Exercise-induced abdominal muscle fatiguein healthy humans. Journal of Applied Physiology 100 (5), 1554–1562.
Taylor, B.J., Romer, L.M., 2008. Effect of expiratory muscle fatigue on exercise tol-erance and locomotor muscle fatigue in healthy humans. Journal of AppliedPhysiology 104 (5), 1442–1451.
Verges, S., Lenherr, O., Haner, A.C., Schulz, C., Spengler, C.M., 2007. Increasedfatigue resistance of respiratory muscles during exercise after respiratory mus-cle endurance training. American Journal of Physiology – Regulatory, Integrative,and Comparative Physiology 292 (3), R1246–R1253.
Verges, S., Notter, D., Spengler, C.M., 2006. Influence of diaphragm and rib cage mus-cle fatigue on breathing during endurance exercise. Respiratory Physiology andNeurobiology 154 (3), 431–442.
Verges, S., Renggli, A.S., Notter, D.A., Spengler, C.M., 2009. Effects of different respi-ratory muscle training regimes on fatigue-related variables during volitionalhyperpnoea. Respiratory Physiology and Neurobiology 169 (3), 282–290.
Vogiatzis, I., Aliverti, A., Golemati, S., Georgiadou, O., Lomauro, A., Kosmas, E.,Kastanakis, E., Roussos, C., 2005. Respiratory kinematics by optoelectronic ple-thysmography during exercise in men and women. European Journal of AppliedPhysiology 93 (5–6), 581–587.
Volianitis, S., McConnell, A.K., Koutedakis, Y., McNaughton, L., Backx, K., Jones, D.A.,2001. Inspiratory muscle training improves rowing performance. Medicine andScience in Sports and Exercise 33 (5), 803–809.
Ward, M.E., Eidelman, D., Stubbing, D.G., Bellemare, F., Macklem, P.T., 1988. Respi-ratory sensation and pattern of respiratory muscle activation during diaphragmfatigue. Journal of Applied Physiology 65 (5), 2181–2189.
Wilson, S.H., Cooke, N.T., Edwards, R.H., Spiro, S.G., 1984. Predicted normal values
for maximal respiratory pressures in caucasian adults and children. Thorax 39(7), 535–538.Wylegala, J.A., Pendergast, D.R., Gosselin, L.E., Warkander, D.E., Lundgren, C.E., 2007.Respiratory muscle training improves swimming endurance in divers. EuropeanJournal of Applied Physiology 99 (4), 393–404.





























