Embed Size (px)
Citation preview

Retrospective Theses and Dissertations Iowa State University Capstones, Theses andDissertations
2002
Comparison of porcine CD4 and gamma-delta Tcell systemic and mucosal responses to thespirochete Brachyspira hyodysenteriaeRaquel Hontecillas-MagarzoIowa State University
Follow this and additional works at: https://lib.dr.iastate.edu/rtd
Part of the Immunology and Infectious Disease Commons, Medical Immunology Commons,and the Pathology Commons
This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State UniversityDigital Repository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State UniversityDigital Repository. For more information, please contact [email protected].
Recommended CitationHontecillas-Magarzo, Raquel, "Comparison of porcine CD4 and gamma-delta T cell systemic and mucosal responses to the spirocheteBrachyspira hyodysenteriae " (2002). Retrospective Theses and Dissertations. 520.https://lib.dr.iastate.edu/rtd/520

INFORMATION TO USERS
This manuscript has been reproduced from the microfilm master. UMI films
the text directly from the original or copy submitted. Thus, some thesis and
dissertation copies are in typewriter face, while others may be from any type of
computer printer.
The quality of this reproduction is dependent upon the quality of the
copy submitted. Broken or indistinct print, colored or poor quality illustrations
and photographs, print bleedthrough, substandard margins, and improper
alignment can adversely affect reproduction.
In the unlikely event that the author did not send UMI a complete manuscript
and there are missing pages, these will be noted. Also, if unauthorized
copyright material had to be removed, a note will indicate the deletion.
Oversize materials (e.g., maps, drawings, charts) are reproduced by
sectioning the original, beginning at the upper left-hand comer and continuing
from left to right in equal sections with small overlaps.
ProQuest Information and Learning 300 North Zeeb Road, Ann Arbor, Ml 48106-1346 USA
800-521-0600


Comparison of porcine CD4 and gamma-delta T cell systemic and mucosal
responses to the spirochete Brachyspira hyodysenteriae
by
Raquel Hontecillas-Magarzo
A dissertation submitted to the graduate faculty
in partial fulfillment of the requirements for the degree of
DOCTOR OF PHILOSOPHY
Major: Immunobiology
Program of Study Committee: Michael J. Wannemuehler, Major Professor
Janice E. Buss Susan J. Lamont Harley W. Moon Randy E. Sacco
Iowa State University
Ames. Iowa
2002
Copyright © Raquel Hontecillas-Magarzo. All rights reserved.

UMI Number: 3073455
Copyright 2003 by
Hontecillas-Magarzo, Raquel
All rights reserved.
UMT UMI Microform 3073455
Copyright 2003 by ProQuest Information and Learning Company. All rights reserved. This microform edition is protected against
unauthorized copying under Title 17, United States Code.
ProQuest Information and Learning Company 300 North Zeeb Road
P.O. B0X1346 Ann Arbor, Ml 48106-1346

ii
Graduate College Iowa State University
This is to certify that the doctoral dissertation of
Raquel Hontecillas-Magarzo
has met the dissertation requirements of Iowa State University
Major Professor
For the Major Program
Signature was redacted for privacy.
Signature was redacted for privacy.

iii
TABLE OF CONTENTS
ABSTRACT
CHAPTER 1. GENERAL INTRODUCTION I Background and rationale I Objectives of the dissertation 5 Dissertation organization 5 Literature review 6
Functional differences between aP and y8 T cells 6 Origin and distribution 6 Differential aspects of antigen recognition between ap and y8 T cells 7
Comparison and contrast between the (%P and yô T cell receptor 8 Structure of the T cell receptor 8 Differences between the (%P and y8 T cell receptor 12
Porcine CD4 T cells 13 Coexpression of CD8aa in mature porcine CD4T T cells 15 The CD8 coreceptor: isoforms, function and regulation of cell surface expression 16 Possible explanations for the function of mature CD4+CD8aaT cells 18
Porcine y8 T cells 19 Lymphocyte composition of the intestinal mucosa 21 Role of T cells in the development and regulation of intestinal inflammation 23
Cellular mechanisms involved in intestinal mucosa homeostasis 23 Role of CD4+ T cells in the establishment and resolution of inflammation 24 Y8 T cell involvement in the control of inflammation 27
References 29
CHAPTER 2. DIFFERENTIAL REQUIREMENTS FOR THE PROLIFERATION OF PORCINE CD4 AND yô T CELLS TO EXTRACELLULAR BACTERIAL ANTIGENS 43
Abstract 43 Introduction 44 Materials and methods 46 Results 49 Discussion 52 References 61

iv
CHAPTER 3. MODIFICATION OF THE LYMPHOCYTE COMPOSITION AND THE CYTOKINE ENVIRONMENT DURING Brachyspira hyodysenteriae-INDUCED COLITIS BY SYSTEMIC IMMUNIZATION WITH A PROTEASE DIGESTEDBACTERIN 64
Abstract 64 Introduction 65 Materials and methods 66 Results 70 Discussion 73 References 85
CHAPTER 4. GENERAL CONCLUSIONS 89
ACKNOWLEDGEMENTS 95

V
ABSTRACT
In pigs, «P CD4+ and yô T cells constitute an important fraction of the circulating
lymphocyte pool. This makes the pig an ideal animal model for functional studies of the two
cell populations. A comparative evaluation of the systemic and mucosal responses of porcine
CD4+ and yô T cells was conducted in the context of immunization and/or challenge with the
pathogen Brcichyspirci hyodysenteriae. This spirochete causes a severe colitis in pigs;
however, immunization with a pepsin-digested bacterin ameliorates clinical signs of disease.
Protection has been associated with in vitro antigen-dependent proliferation of CD41" and
yS T cells to B. hyodysenteriae antigens. A first set of experiments was conducted to evaluate
the differential functional aspects of the systemic responses to B. hyodysenteriae antigens of
CD41" and yÔ T cells. Results show that, while the proliferative responses of CD4+ were
exclusively antigen-specific, y5 T cells responded to stimuli other than B. hyodysenteriae
antigens. Proliferation of the two cell populations in vitro was dependent on the presence of
IL-2. In addition, examination of IFN-y production indicated that CD4+ T cells are the major
source of this cytokine, although stimulation with B. hyodysenteriae antigens decreased IFN-
y-producing CD4+ T cells. This effect was paralleled by an increase in yô T cells producing
IFN-y. These results suggest that, as has been demonstrated for other infectious disease
models, yô T cells may contribute to downregulate CD4+ T cell responses. A second study
describes the impact that immunization and/or challenge had on the lymphocyte composition
and cytokine environment of the colonic mucosa. Immunization decreased the colonic CD41"
T cells irrespective of the challenge status, whereas challenge with B. hyodysenteriae resulted
in depletion of epithelial yS T cells; although, vaccination ameliorated the loss of this

vi
lymphocyte subset. These cellular changes were accompanied by modulation of cytokine
expression. Particularly, TNF-a, IL-1 (3 and TGF-3i mRNA were upregulated in vaccinated
and infected pigs. The possible contribution of porcine CD4^ and yô T cells to the
development and resolution of B. hyodysenteriae-induced colitis and the mechanisms of
activation of the two subsets are discussed in this dissertation in the context of the present
findings.

1
CHAPTER 1. GENERAL INTRODUCTION
Introduction
Brachyspira hyodysenteriae is a spirochete that causes swine dysentery, a severe
mucohemorrhagic colitis (I). In addition to the etiologic agent, the presence of an appropriate
intestinal flora is required in order for the disease to develop. This was shown in experiments
involving germ-free mice or pigs, in which disease developed only after coinfection with
Bacteroides vulgatus and B. hyodysenteriae (1 ; 2).The molecular pathogenesis of swine
dysentery has not been elucidated; however, it is believed that colonization of the mucosa by
B. hyodysenteriae results in the loss of the epithelial barrier function. This is due in to the
production by the spirochete of a ^-hemolysin that is cytotoxic for enterocytes (3). This
initial phase is followed by the invasion of the mucosa by commensal bacteria from the
colon. Excessive exposure of the underlying mucosa to luminal antigens ultimately leads to a
severe, acute inflammatory response (3; 4), which was characterized in a murine model o the
disease by the upregulation of proinflammatory cytokines, such as tumor necrosis factor-
ex (TNF-a), interleukin I-(3 (IL-1(3) (5). Figure 1 depicts the most characteristic histologic
features of the disease: superficial epithelial erosion, increased thickness of the mucosa with
crypt hyperplasia, and leukocyte infiltration. The disease is basically restricted to the mucosa,
although multifocal leukocytic infiltrates may sometimes be present at the submucosa (6).

Figure 1: Photomicrographs of hematoxilin-eosin stained sections of porcine colon from a healthy (A) and a B. hyodysenteriae infected (B) pig at 160 x magnification. The most characteristic histopathologic features of the disease are increased thickness of the epithelium (indicated by arrows) with crypt hyperplasia, superficial epithelial erosions (indicated by an asterisk) and leukocytic infiltration.
Systemic immunization with a B. hyodysenteriae protease-digested bacterin provides
protection against the disease (i.e., reduced clinical signs), although it does not prevent
bacterial colonization (7; 8). The vaccine has also been used as a tool for evaluating porcine
T cell responses to extracellular bacterial antigens (8-12). A diagram showing the phenotypic
classification of porcine T cells is depicted in figure 2. Based on T cell receptor (TCR)
expression, porcine T cells can be divided into afi or yô T cells. aP T cells can be further
subdivided depending on CD4 and CD8 coreceptor expression. Besides the CD4"1" CD8" aP T
cells, three subsets of porcine CD8+ TCR aP T cells have been described: 1) CD4 CD8
(%P or cytotoxic T lymphocytes, 2) CD4+ CD8acT or memory/effector T cells, and 3) CD4"
CD8acT T cells, of unknown function (13-19). Finally, two major subsets of y5 T cells have
been characterized: 1) SWC6^ CD2", which constitute the majority of the peripheral blood
yô T cells, and 2) SWC6*CD2+ which are low in peripheral blood and preferentially migrate
to non-lymphoid organs and spleen. A fraction of the SWC6~CD2+ y5 T cells also express
CD8a (20-22).

3
Porcine T-lymphocytes
a0 T cells yS T cells
SWC6+ CD2-CD4+CD8- CD4+ CD8aa+
SWC6- CD2+CD8aa+
Figure 2: Subset classification of porcine T lymphocytes
Previous studies reported that there was an increase in the percentage of CD8cf
lymphocytes in peripheral blood, colonic lymph nodes (CLN), and colonic mucosa following
challenge of vaccinated pigs with B. hyodysenteriae (10). Also, in vitro stimulation of
peripheral blood mononuclear cells (PBMC) isolated from immunized animals resulted in
antigen-dependent proliferation of CD8cf lymphocytes, CD4+ CDSacf, CD4+ CD8aa+, and
TCR yô cell subsets. Blockade of antigen presentation with antibodies against class II major
histocompatibility complex (MHC-II) diminished in vitro proliferation of PBMC from
immunized pigs, and similar results were obtained by depleting CD4+ T cells (9). Therefore,
vaccination with a B. hyodysenteriae bacterin induces a MHC-II-dependent CD41" T cell
response. However, because significant proliferative responses of yô T cell are also induced
following in vitro antigenic stimulation of unfractionated PBMC, this immunization regime
can be utilized for the study of the differential mechanisms of activation of CD4+ and yô T
cells.

4
The identification of yô T cells is relatively recent compared to a|3 T lymphocytes
(23). Although both aP and yô T cells have the capacity to specifically recognize antigens
through their respective TCR, the type of antigens recognized and the process of antigen
recognition are different between the two subsets (23). In fact, knowledge on yS T cell
biology has greatly increased in recent years, making it evident that their function in the
immune system is different from that of ap T cells (23). The tissue distribution pattern of
aP and yô T cells is a first indicator of their differential role. In mice and humans, y5 T cells
are almost absent from peripheral blood and organized lymphoid tissues. In other species
such as poultry, cattle or pigs, yô T cells constitute a major proportion of the pool of
circulating lymphocytes, and they are present in secondary lymphoid organs although in low
numbers (24-26). In both cases, yS T cells are abundant at mucosal sites (27). On the other
hand, the T cell population within the spleen and lymph nodes are predominantly aP T cells.
These are the compartments of the immune system where naïve T cells are first exposed to
antigens processed and presented by professional antigen presenting cells (APC) and after
antigen presentation, naïve T cells undergo clonal expansion and differentiation into
effector/memory cells (28). Because yS T cells are not retained at these sites, it could be
predicted that their involvement in the immune response is different from that of ap T cells.
Questions relating to the process of antigen presentation to yÔ T cells (e.g., MHC-restriction,
requirements for APC or costimulation) and whether or not they become memory cells are
currently under investigation (27).
yô and CD41" T cells are the two T cell subsets responding following immunization
with a B. hyodysenteriae bacterin (10). During the clinical period of swine dysentery, the
bacteria invade the colonic mucosa without spreading systemically (6). Thus, by utilizing

5
different routes of antigenic exposure, this model offers the possibility of evaluating the local
response of CD4+ and y5 T cells at the colonic mucosa (i.e., following challenge),
systemically (i.e., by intramuscular immunization), or as a result of the interaction between
immunization and challenge.
Objectives of the dissertation
The objectives of the experiments conducted for this dissertation were to evaluate the
impact that immunization and /or challenge with B. hyodysenteriae had on porcine CD4~ and
yS T cells. More specifically, we investigated in vitro differential aspects of the responses
(i.e.. proliferation and cytokine production) of CD4+ and yS PBMC to B. hyodysenteriae
antigens following immunization. At the colonic mucosal level, we examined the changes
that immunization and/or challenge induced on the numbers and distribution of these subsets
at the colonic mucosa.
Dissertation organization
This dissertation is organized in four chapters: a General Introduction (chapter 1), two
journal style manuscripts (chapters 2 and 3) and a General Conclusion (chapter 4).

6
LITERATURE REVIEW
Functional differences between af) and y8 T cells
T cells derive from pluripotent precursors that follow the lymphoid lineage of
differentiation (29; 30). Their designation as "T cells" comes from their "thymus-dependent"
origin. Although this terminology has been maintained, the existence of extrathymically-
derived T lymphocytes has been known for a long time (31-33). By definition, all T cells are
lymphocytes that in mature state express a T cell receptor (TCR) (28). The TCR is a
heterodimeric molecule that results from the association of a and P (TCRaP), or y and
5 (TCRyô) polypeptide chains. Thus, two major subsets of T cells exist based on the type of
TCR expressed on the cell surface: (%P and yô T cells (34). Concepts related to T cell biology,
such as MHC restriction, immunological memory or acquired immunity resulted from studies
on «P T cells. yS T cells were identified more recently than aP T cells, and initial studies
demonstrated that some principles of aP T cell biology would not apply to them. There is
enough evidence now to support that, rather than being exceptions, y8 T cells have different
and non-overlapping roles in comparison to (%P T cells within the immune system (23; 35).
Origin and tissue distribution of C# and yS T cells
Although both cell types originate from a common lymphoid precursor, they diverge
early in ontogeny (36). During lymphocyte development, thymocytes commit to either the
aP or yô T cell lineage. Thymocytes that successfully rearrange a TCR, either aP or yS,
become naïve mature T cells (30). A difference between aP and y8 T cell development is that
y8 T cells are considered mature lymphocytes from the moment they express a functional
TCR (30; 37). At this point, they migrate to the periphery (27). Conversely, thymocytes that
have committed to the aP T cell lineage go through several checkpoints before becoming

7
mature cells. The first checkpoint occurs during the process known as |3 selection (i.e., in-
frame rearrangement and successful expression of the TCRP chain) (38), afterwards, a(3 TCR
thymocytes capable of recognizing self-MHC through the rearranged TCR are rescued from
apoptosis or are positively selected (39). Finally, the recognition of self-peptides through the
TCR generates strong signals that activate apoptosis. This phenomenon, known as negative
selection, prevents auto-reactive T cells from migrating to the periphery (40; 41).
A second aspect that differentiates a(3 and yÔ T cells is their trafficking patterns.
Naïve a(3 T cells migrate from the thymus to secondary lymphoid organs where, eventually,
they will be exposed to antigen and differentiate into effector or memory
cells. Effector/memory a(3 T cells then home to non-lymphoid tissues where they sample for
their cognate antigen. In contrast, yô T cells directly accumulate in non-lymphoid tissues,
especially at mucosae, without the need of a previous antigenic exposure (27; 42; 43). Most
challenges to the immune system of environmental origin, (e.g., infectious agents or
allergens), occur at mucosal sites. While yô T cells are ready to respond to an antigenic
challenge, aP T cells must have been previously exposed to the antigen in a secondary
lymphoid organ. Because this process requires some time for aP T cells, it has been proposed
that yô T cells constitute a first line of defense (44).
Differential aspects of antigen recognition between af) and y6 T cells
Antigen presentation to naïve aP T cells is a function of dendritic cells (DC) that
have migrated from the periphery to secondary lymphoid tissues (45). Because yô T cells are
not retained at these sites, the question then is how does antigen presentation to yô T cells
occur? aP T cells recognize peptides resulting from antigenic processing, presented in
association with MHC-class I or II molecules on the surface of target cells or APC to CD8+

8
(i.e., cytotoxic) or CD4+ (helper) T cells (28). The yô TCR is MHC-I and II-unrestricted;
instead, non-classical MHC molecules, namely T10/T22(46), its murine homologue retinoic
acid early inducible Rae (47) antigen or CD I (48) are ligands for the yô TCR. In addition,
previous research showed that inhibition of antigen processing and presentation by classical
MHC molecules did not affect y5 TCR activation (46). These results lead to the conclusion
that yô T cells recognize unprocessed antigens. However, in similar action to a(3 T cells, cell
to cell interaction and costimulation are required for yô T cell activation through the TCR
(49). Therefore, yô T cells are capable of recognizing unprocessed antigens directly on
tissues without the mediation of a professional APC (27).
MHC-I and II molecules are specialized in presenting peptidic epitopes to T cells
(50). In addition to recognizing protein-derived antigens (e.g.,glycoprotein 1 from Herpes
simplex virus) (51), yô T cells recognize non-peptidic molecules. Special attention has been
given to the capacity of phosphoantigens, low molecular mass molecules rich in phosphate
groups abundant in bacteria (e.g., Mycobacterium spp, Escherichia coli), to stimulate human
yÔ T cells (52-55). In addition, it has been shown that yÔ T cells can recognize stress-induced
self-molecules such as MHC-class I related chain A (MICA) (56), its murine homologue
retinoic acid early inducible Rae (47) antigen (56), and transformed cells (57).
Comparison and contrast between the a0 and the y8 TCR
Structure of the TCR
Studies on the structure of the TCR have provided a basis for some of the differences
between a(3 and yô of T cells. As already mentioned, the TCR is a heterodimer formed by
two disulphide-linked polypeptide chains; a-fi or y-ô . It is always expressed along with the

9
CDS complex, thus phenotypically, T cells are designated as CD3+. The CD3 complex is the
association of three different polypeptide chains (i.e., CD3y, CD3S and CD3e), with an
extracellular immunoglobulin-like (Ig-like) domain and a short cytoplasmic tail. Also, most
of the TCR-CD3 complexes are associated with a homodimer formed by two Ç chains, or a
heterodimer formed by Ç and its alternative spliced form, x\. The latter proteins have a very
short extracellular domain and a long cytoplasmic tail that very actively participate in TCR
signal transduction (58). The composition of the TCR complex is not constant and varies
depending on developmental or activation state of the cell (28; 59; 60).
Based on genetic features and protein structure, the TCR has been classified within
the immunoglobulin superfamily. Figure 3 depicts the structure of a TCR. Each polypeptide
chain results from the combination of one invariant, or constant (C) (i.e., Ca and CP in the
TCRaP, or Cy and CÔ in the TCRyô), and one variable (V) region (i.e., Va and VP or Vy
and Vô) (28; 30; 61-63). The C portion of the protein has an Ig-like domain followed by a
hinge region, a transmembrane region and a short cytoplasmic tail. Cystein residues at the
hinge region are involved in dimer formation between a and P or y and Ô chains. The
transmembrane domain facilitates the association between the TCR and the CD3 signaling
complex. In the case of antibodies, different isoforms exist for the C region (i.e., isotypes). In
contrast, isotypic variation at the TCR C region is lower (28). Two isoforms of the Cp genes
have been described in the mouse with apparently no significant functional differences (64)
and three isoforms of the Cy have been identified in the pig (65).
The variable domain of the TCR results from the somatic recombination of germline
encoded gene segments (29). The great advantage that this unique system shared by the TCR,
B cell receptor (BCR) and antibodies, is the generation of receptor diversity. This allows for

10
the specific recognition of the great variety of antigens to which the immune system is
exposed. TCR diversity is achieved by recombination of different segments that will form the
V chain. The a and y V chains result from the recombination of V and joining (J) segments,
(i.e., VJ). In the P and S chains, an additional diversity (D) segment is incorporated into the V
chain (i.e., VDJ). The existence of several V, D and J genes that can be recombined in the V
chain is what brings about receptor diversity (66).
Amino acid sequence comparisons of V domains of each of the polypeptide chains
that form the TCR indicated that within the V chain there are regions of hypervariability
(HV) (67). The HV regions of the a-P or y-S chains form the complementary determining
regions (COR) on the TCR. The CDR1 and CDR2 domains interact with MHC-I or II for aP
T cells, or a non-classical MHC molecule in the case of yô T cells. The CDR3 is the region of
the TCR that is directly implicated in antigen recognition (68; 69). This feature is indicated in
figure 3.

Il
Structural differences between the C regions of the aP and yô TCR suggest that the two types of
receptors form different signaling complexes.
CDR3 has the same length in Va and Vp, but it is longer in Vô than in Vy.
This feature is also present in antibody molecules. From this finding it was interpreted that yô T cells recognize
epitopes in a similar fashion to immunoglobulins.
Figure 3. Crystal structure of an aP T cell receptor (TCR) interacting with a major histocompatibility complex (MHC-l) molecule loaded with a peptide (P1-P8). The complementary determining regions (CDR) are the loops represented in colors at the are area of contact between the TCR and the MHC-peptide complex. The major structural differences between the ap and the yô TCR, and the functional implications that they have are indicated in boxes (with permission from LA, Wilson) (70).

12
Differences Between the (*P and y5 TCR
Comparative studies examining the structure and function of the a(3 and yô TCR
showed that the CDR3 of VS is longer than the CDR3 of Vy (70). This structural feature
results from the rearrangement of up to three D segments and the addition of N-nucleotides
in VÔ (71). In contrast to this, the CDR3 of the a and p chains are of the same length (70).
Besides, the number of amino acid residues that form the CDR3 is larger in the yô than in the
aP TCR. These structural differences have important functional implications (70). T cells
recognize peptides of 6 to 12 aminoacids in length that result from antigen processing and are
presented by MHC molecules on APC (28). In turn, yô T cells can recognize antigens of
different chemical structure other than proteins (e.g., phosphoantigens or lipoproteins) (52-
55; 72; 73). In fact, based on the basis of the CDR3, the yÔ TCR closely resembles
antibodies, which have longer CDR3 in the heavy than in the light chain (70). These
structural similarities between immunoglobulins and the yôTCR suggest that yÔ T cells can
recognize unprocessed antigens (46; 74-76). However, in contrast to antibodies, antigen
recognition by yô TCR requires cell to cell contact (49).
The first crystallographic model of a complete human yô TCR (i.e., Vy9 VÔ2) was
recently published (63). Vy9 VÔ2 T cells constitute the majority of the circulating y5 T cells
in humans (77-79). In this specific case, the CDR3 regions are of the same length in the y and
Ô chains. The most significant differences with the aP TCR are found in the C chain. The
comparison of both the structure and shape of the «P and yô TCR reveals that they form
different signaling complexes. First, within the C chain Ig-like domain, the polypeptide
domain that in the «P TCR apparently associates with the CD3E (i.e., FG loop), is longer
than the corresponding region in the yô TCR. In addition, both receptors differ in their hinge

13
regions, with a maximum of 7 versus 23 residues between the end of the Ig-like domain and
the interchain disulphide bond in the ap and yÔ TCR, respectively. These particularities
indicate that a different type of association exists between the CD3 complex and the a(3 or
yS TCR and that the receptor on yS T cells has higher flexibility (63). As already mentioned,
three Cy isoforms have been described in the pig (65; 80). It is interesting that the three
isoforms are different from one another in the hinge region and in the cytoplasmic tail.
Again, these differences suggest that yô T cells form different signaling complexes and,
therefore, distinct signal transduction pathways may regulate activation of the different yô T
cell subsets.
In summary, both the a(3 and the yô TCR belong to the immunoglobulin superfamily,
however, modifications of the C and V domains account for their differential biological
properties. Mainly, the type of antigen that they recognize, the mechanisms of antigen
recognition, and the association with signaling complexes are characteristic for each receptor.
The overall result is that the two cell types have different roles within the immune system.
Porcine aP CD4 T cells
An important role of the CD4+ T cells is to coordinate acquired immune responses.
They are involved in both cellular and humoral responses by serving as intermediates
between antigen presenting cells (APC) and B cells, for the production of antibodies, or
cytotoxic T cells (28). T helper 1 (Thl) and Th2 cells regulate immune responses involved in
the elimination of foreign antigens of microbial origin (81). CD4+ T cells are implicated in
the discrimination of self from non-self are equally important. In this regard, CD4+

14
regulatory T cells control other lymphocytes capable of recognizing self-antigens preventing
autoimmune diseases (82).
CD4 is a glycoprotein that possesses 4 Ig-like domains. It interacts with a non-
polymorphic region on the (32 domain of the MHC-II molecule stabilizing antigen
recognition by the TCR. In addition, its cytoplasmic tail associates with Lck, a tyrosine
kinase of the Src family that participates in proximal signal transduction events (83-85).
Although no monoclonal antibodies that recognize the a or p chains of the porcine TCR are
available, CD4+ T cells are CD3+ and are considered TCRaP cells (14; 86; 87). The Thl/Th2
paradigm has not been demonstrated for porcine T cells. However, it is assumed that porcine
CD4+ T cells have similar roles to CD4T T cells from other species, and that they use the
similar effector mechanisms (87; 88). CD4 T cells provide help for the generation of
alloantigen-specific responses (89). The expression of MHC-II antigens is unique to a subset
of porcine CD4+ T cells (i.e., CD44" CD8acO- Although the significance is unknown, it
seems that it does not confer them with the ability to present antigens, because the absence of
other accessory APC abrogated alloantigen-specific responses (89). It has also been
suggested that CD4"1* T cells interact with B-cells for the generation of humoral responses
(90).
Most of the information currently available with regards to the biology of porcine
CD4+ T cells has been obtained in the context of the cellular immune responses to infectious
agents, mainly of viral origin. As it can be expected, infections of bacterial, viral or parasitic
etiology induce detectable in vivo changes in the CD4+ subset (91-96). In vitro, CD4+T cells
proliferate in response to viral or bacterial antigens and these antigen-specific responses are

15
MHC-II restricted (9; 88; 97). In addition, CD4+ T cells become CD8aa+ during antigen-
specific recall responses (18).
Coexpression of CD8aa in mature porcine CD4 T cells
The simultaneous expression of the CD8aa coreceptor on peripheral mature CD4+T
cells has been emphasized as one of the distinctive characteristics of the porcine immune
system. However, other species have also detectable numbers of CD4+ CD8aa+ T cells (e.g.,
monkeys and chickens) (17). The CD8 molecule is coexpressed on mature CD4+ T cells in
low levels and always as a CD8aa homodimer, which differentiates it from the CD8«P
heterodimer on CD8+ cytotoxic T cells (9; 14; 98). CD4+CD8aa+T cells are normal
components of the gut mucosa (99). In healthy individuals conditions, a small percentage of
peripheral lymphocytes are CD4+CD8aa+T cells. However, the presence of this subset is
associated with certain diseases such as neoplasia (100) or rheumathoid arthritis (99).
It is has been accepted that porcine CD4^CD8acTT cells represent a fraction of
memory/activated cells within the CD4"1" pool (18). This assumption is based on the lack of
expression of CD1 (101), a molecule present on immature T cells, and the fact that the
numbers of CD4 CD8aa T cells increase with age, suggesting that this is the result of
antigenic exposure (17; 18). Ontogenetically CD4+CD8aa+T lymphocytes are undetectable
until day 90 of gestation. The peripheral blood of newborn piglets posses approximately I %
of CD4+CD8aa+T cells (102). This number increases to 30-55 % of the total PBMC in 3
year old pigs (18).
In vitro experiments showed that sorted CD4+ T cells from pigs immunized against
pseudorabies virus (PRV) became CD8acT after ex vivo stimulation with PRV antigens (18).
Moreover, most of the CD4 CDScxct T cells also coexpressed CD29, a phenotypic marker of

16
memory T cells (18). Neither the mechanisms regulating CD8aa expression on porcine T
cells, nor its functional role on these cells have been investigated to date.
The CDS coreceptor: isoforms, function and regulation of cell surface expression
CD8 is a glycoprotein of the immunoglobulin superfamily. There are two isoforms:
CD8a and CD8P; the association of two a chains or an a and a P chain form the
homodimeric or heterodimeric forms of the coreceptor, respectively (103; 104). The CD8aP
form is characteristically expressed on cytotoxic T cells (104; 105), while CDSaa can be
expressed on dendritic cells (106), natural killer cells, yô T cells and on mature CD4T T cells
(107). The CD8 coreceptor facilitates the interaction with APC. Both forms of the coreceptor
recognize non-variable regions on the MHC-l molecule (108) and, in addition, CDSaa
recognizes non-classical MHC antigens as has been reported for thymus leukemia (TL)
antigens expressed by enterocytes ( 109).
There has been some controversy regarding which form of CDS is more efficient as a
coreceptor. Soluble CDSaa and CDSaP molecules have equal binding affinity for MHC-I
(110), however, cell-associated CDSaP is more efficient coreceptor than CDSaa (111). The
clearest difference between the coreceptors is in their contribution to T cell activation. CDSa
has a long cytoplasmic tail that associates with Lck (85). As it has been indicated for the CD4
coreceptor, this interaction is important in early signal transduction events mediated by the
TCR. CD8P has a short cytoplasmic tail and does not associate with Lck (112); however, it
has a domain that is necessary for palmitoylation and for the recruitment of CDSaP into lipid
rafts, where most of the cellular Lck is found (113). Deletion of the palmitoylation residue on
CDSP abrogated its ability to partition into lipid rafts, and chemical inhibition of
palmitoylation decreased T cell activation (114; 115). Thus, CDSP is crucial role for the

17
formation of efficient signaling complexes and activation of cytotoxic lymphocytes through
the TCR. On the other hand, although CD8aa can associate with Lck, the lack of a
palmitoylation residue suggests that it cannot come in contact with the TCR, and therefore, it
does not participate in T cell activation in the same way that CD8«P would (115).
In the mouse, the genes that code for CD8a and CD8(3 are in close proximity on
chromosome 6(116). Both transcriptional and postranscriptional mechanisms for the control
of the expression of these genes have been described. Within the transcriptional mechanisms,
there are four enhancers (E8t-E8iv) that are tissue-, lineage-, and developmentally-specific
(117; 118). For instance, E8, regulates CD8a expression in T cells and mature single positive
(SP) thymocytes, but it has no effect in double positive (DP) thymocytes. Also, it controls the
expression of CD8a in NK cells, intestinal TCRyô CD8aa and TCRaP CD8aa T cells but
not in TCRaP CD8a|3 intraepithelial lymphocytes (IEL) (118-120). Neither CD8a nor
CD8P genes of the pig have been sequenced, and no information is available on how its
expression is regulated. It could be anticipated that an element similar to E8, exists which
regulates the expression of porcine CD8a gene, because the homodimeric form of CD8a is
expressed in equivalent cell subsets. With regards to postranscriptional regulation, mature
human CD4+ T cells transcribe CD8a, although the mRNA is very unstable and the protein is
never produced (121). However, it was reported that phorbol myristate acetate (PMA)-
activated human CD4+ T cell clones were able to synthesize and coexpress CD8a in the
presence of interleukin-4 (IL-4). The synthesis of CD8a was transient and downregulated by
IL-2 (122). Although these are in vitro studies, certain human diseases such as neoplasia
(100) or rheumathoid arthritis (99) are associated with an increase in recirculating mature

18
CD4+CD8TT cells. Therefore, molecular mechanisms that stabilize CD8a mRNA could
enhance the expression of CD8a on the surface of CD4+ T cell.
Possible explanations for the function of mature CD4+CD8aa+T cells
A question that remains to be answered is what are the functional differences between
CD4""T cells and CD4+CD8acf T cells. It has been hypothesized that human CD4+CD8acT
are regulatory cells. Consistent with this hypothesis, cloned CD4+CD8+aaT cells from
lepromatous leprosy patients inhibited activated T cells (124). In line with the findings in
human lymphocytes, porcine CD4TCD8+aaT cells isolated from peripheral blood were
shown to express high levels of IL-10 mRNA (92). IL-IO is a cytokine associated with
regulatory responses (125; 126). However, a different study showed that peripheral blood
CD4^CD8aa*T cells produced high levels of IFN-y when stimulated with PMA and
ionomycin (127).
A second theory proposes that circulating human CD4TCD81aaT cells have
cytotoxic activity. In support to this theory, addition of anti-CD8a monoclonal antibodies
blocked anti-CD3-induced cytotoxicity of human CD4+CD8+aaclones (123). Porcine CD4T
CD8+aaT cells isolated from PRV-immunized pigs contributed to MHC-class [-restricted,
antigen-specific cytotoxic activity of CTLs (128). It has also been suggested that porcine
CD4+CD8+aaT cells could themselves be cytotoxic (129). It is improbable that the
mechanism of cytotoxicity depends on dual MHC-I/MHC-II restriction of CD4+CD8aoTT
cells, because the CD4+ T cell precursors express a TCR specific for a MHC-II-dependent
epitope. Although this rules out the dual MHC-I/MHC-II-restriction of porcine CD41"
CD8aa+T cells, they could still induce cytotoxicity of target cells by alternative mechanisms
(e.g., Fas-FasL interaction). This hypothesis, however, has not been tested. Studies on the

19
localization of CD8aa on the cell membrane of CD4+CD8aa+T cells and the determination
of the molecules that associate with the cytoplasmic tail would help in elucidating if
CD8aa contributes to TCR-mediated activation of CD4+T cells. In addition, the
identification of the factors that stimulate CD8aa expression would provide some insights
into the possible role of porcine CD4+CD8aa+T cells as memory/effector T cells.
Porcine yfi T cells
yô T cells were discovered during the identification and sequencing of the
genes for the a and (3 chains of the TCR (130-132). One of the principal characteristics of y5
T cells is that they tend to accumulate in non-organized lymphoid tissues, especially the skin
and intestinal mucosa (27). However, species-specific differences have been reported, with
mice and humans having very low numbers of circulating yô T cells while in pigs, ruminants
and chickens they constitute an important fraction of the PBMC (24-26). When monoclonal
antibodies became available for some of the cell surface markers of porcine lymphocytes, it
was observed that a high proportion of PBMC were negative for either B or T cell surface-
expressed proteins (i.e., CD4, CD8 or slgM). These "non-B, non-T cells" were subsequently
phenotypically characterized as y8 T lymphocytes (133).
The classification of mouse and human yô T cells is based on their variable TCR gene
usage (35) while the pig it is based on their phenotypic characteristics. There are two main
subsets of porcine y5 T cells. One is the analogue of bovine WCl^yô T cells and is
phenoty pically defined by the expression of SWC6 and CC101 antigens. Bovine WC1 is a
member of the scavenger receptor cysteine-rich family and, antibodies against CC101
recognize a related molecule expressed by pig yô T cells. This subset is also CD2~ and CD8a.

20
SWC61" TCRyô cells constitute the majority of the yô T cells in PBMC (26; 134). The second
subset can be phenotypically defined as SWC6" CD2+, of which a small fraction expresses
CD8a. Cells from this subset have been detected in the spleen and lymph nodes and as
recirculating lymphocytes (134; 135).
Protein analyses of the yS TCR yield a unique form of the 8 chain of 38 kDa, and 3
isoforms of the y chain of 37, 38 and 46 kDa. The 37 and 38 kDa isoforms are expressed in
CD2CD8" (i.e., SWC6*) y5 T cells. These two y chains are subset-specific, as the 38 kDa
iso form is only present in circulating SWC6+ cells that also express the 86D molecule, while
the 37 kDa form is characteristic of circulating but also tissue-resident cells. Finally, the 46
kDa iso form was attributed to CD21" CD8*(SWC6") cells present in secondary lymphoid
tissues (81). Cloning of the porcine Cy chain revealed the existence of 3 iso forms different in
their hinge regions. Two of them (G10 and G15) are similar to each other with longer hinge
regions that have 2 additional cysteine residues in comparison with the G4 isoform. The third
iso form (04) differs from G10 and G15 in its shorter hinge region and in the cytoplasmic
tail. G10 and G15 were identified as the 37 and 38 kDa isoforms of the y chain. The 46 kDa
isoform, however, has not been matched to any Cy sequence (65). A possible explanation for
this is that PBMC were used as a source of T cells for cloning the Cy, when cells bearing the
46 kDa chain are found in lymphoid tissue.
Very little information is available regarding functional aspects of porcine y5 T cells.
There are reports describing yô T cells from peripheral blood proliferating in vitro to viral or
bacterial antigens (9-11; 89; 98). However, they appear to have very low capacity of IFN-y
production in vitro (127). Porcine CD6* lymphocytes are functionally characterized by having
non-specific cytotoxic activity (128). The SWC6+ subset of porcine yS T cell is CD6

21
and has NK-like activity (127; 135). In addition, it was shown that this cell type
preferentially migrated to sites of cutaneous inflammation induced by administration of
phytohemagglutinin (136; 137). There are contradictory results with regard to the SWC6"
yÔ T cells. They have been described as both CD6" (I28)and CD6+ (133) by different authors.
Thus, their function remains unclear.
Lymphocyte composition of the intestinal mucosa
The intestinal mucosa is the most inner layer of the gut wall. It is divided in two
functional and structurally different compartments: lamina propria and epithelium. The
intestinal epithelium is a single cell layer of polarized cells exposed to the lumen, thus, in
direct contact with the intestinal contents (138). Lymphocytes constitute an important portion
of both the lamina propria (i.e., lamina propria lymphocytes, LPL) and the epithelium (i.e.,
intraepithelial lymphocyes, IEL) (139; 140). However, the lymphocyte composition of the
mucosa is not random, but characteristic of the different anatomical regions of the intestine
(e.g., small versus large intestine), and of each of the two compartments (i.e., epithelium
versus lamina propria) (141; 142). In addition, due to the specific functions that the immune
system has at this site, lymphocytes from the intestine are different in several aspects (138).
In regards to the phenotype, they have an atypical pattern of expression of cell surface
molecules compared to conventional lymphocytes at the systemic/central lymphoid system.
First of all, the gut associated lymphoid tissue (GALT) is rich in y5 T cells (143) and, within
the aP T cell fraction, cells expressing CD4 (TCRaP CD4+CD8"), CD8 (TCRaP CD4*
CD84), or neither of the coreceptors (TCRaP CD4~CD8~) have been described (139; 144). In
addition, the expression of the CD8aa homodimer is common among cells of both the aP

and yô T cell lineage (145; 146). In the mouse, it has been proven that the CD8aa I EL
develop extrathymically while the rest are of thymic origin and migrate from the periphery to
colonize the intestine (147; 148). This separation in origin has not been elucidated in other
species
Lymphocytes of the epithelium and lamina propria constitute the diffuse lymphoid
tissue of the GALT as opposed to isolated follicles, Peyer's patches, lymphocyte-filled villi
and cryptopatches, which constitute the organized lymphoid tissue (140; 149). Functionally,
diffuse and oganized lymphoid structures are effector and inductor sites, respectively (140).
In the porcine small intestine, the crypt areas of the lamina propria are rich in plasma cells
while T cells, most them ofCD4+ phenotype although there are also some CD8T, localize
within the villi. The CD4+ cells are in close association with the epithelium, as they are
located right underneath the basement membrane (150). The IEL constitute approximately 20
percent of the epithelial cells and are distributed mainly in the villus. In a recent study, it was
reported that in adult pigs approximately 50 % of the IEL were CD2+ CD4" CD8+ and were
localized in the proximity of the basement membrane. The remaining, CD2+ CD4" CD8" IEL,
were aligned with the enterocyte nucleus except for a small percentage located towards the
apical side (151). This study did not include additional markers, therefore, it cannot be
concluded to which T cell lineage (i.e.,oc(3 oryÔ) these cells belong. When the lymphocyte
composition of the intestinal mucosa was first evaluated, monoclonal antibodies to the
yd TCR were not available yet. For this reason the classification has been incomplete until
recently. yS T cells have been found in both epithelium and lamina propria (152). Currently,
there are no publications describing the lymphocyte composition of the large intestine of the
pig. Immunohistochemical analyses done in our laboratory have shown similar distribution of

23
CD4\ CDS and y§ cells in the colon: CD4+ cells are only present as LPL and, CD8T and
TCRyS localize to both compartments (unpublished data). However, the exact percentages of
and numerical evaluation of each cell type has not been completed yet.
Role of T cells in the development and regulation of intestinal inflammation
Cellular mechanisms involved in intestinal mucosa homeostasis
Cells of the systemic/central immune system are constrained within a sterile
environment and will react to any antigen recognized as non-self. However, the G ALT has
evolved to be in a non-sterile environment, under constant antigenic challenge. Several
mechanisms operate for maintaining tolerance towards antigens from food origin and from
the enteric normal flora. At the same time, the G ALT has to keep its capacity to react to
pathogenic microorganisms (144). Thus, establishment of equilibrium between active
responses and tolerance ensures the maintenance of the barrier function across the mucosa
(153).
Anergy, or lack of reactivity, is one of the mechanisms by which tolerance towards
food antigens or intestine-associated commensal flora is maintained (144). Several theories
have been proposed to explain how anergy is established. It has been hypothesized that
antigen presentation in the absence "danger signals" leads to anergy (154; 155). Such signals
originate from bacterial products referred to as pathogen-associated molecular patterns
(PAMPs) (e.g., lipopolysaccharide or peptidoglycan) which are specifically recognized by
pattern recognition receptors (PRR), such as toll-like receptors (TLR) on the surface of APC
(156; 157). Signaling through TLR results in the surface upregulation of costimulatory

24
molecules thus, in the absence of PAMPs, APC will not provide sufficient costimulation,
which will result in incomplete T cell activation, or even apoptosis (i.e., clonal deletion).
The route of antigenic exposure is an important parameter that contributes to the
maintenance of tolerance. In normal conditions, antigens from the intestinal lumen are
sampled through M-cells (i.e., at the small intestine), or through the apical side of the
absorptive epithelium (138; 140). Enterocytes are polarized cells (i.e., their plasma
membrane is divided into domains with different properties). TLR are expressed by
absorptive enterocytes, but only at the basolateral side (158). Tight junctions prevent
paracellular transport across the epithelium (159-161), therefore, this physical separation
between luminal contents and the basolateral side of enterocytes blocks recognition of
bacterial products by TLR or other PRR. However, whenever the integrity of the intestinal
epitheluim is compromised, the basolateral side of the enterocyte will be exposed to luminal
bacteria, which will ultimately lead to an inflammatory response (162; 163).
Several experimental models have been developed for the study of intestinal
inflammation (153; 164). Irrespective of the etiopathogenesis for each model, a common
denominator is that inflammation results from a break in tolerance and/or an imbalance in the
production of molecules that maintain intestinal homeostasis (153). Cellular interactions at
the intestinal mucosa are extremely complex, however, because of the focus of this
dissertation, only CD4+and yÔT cell involvement in the development and regulation of
intestinal inflammation will be discussed below.
Role of CD4 in the establishment and resolution of inflammation
CD4+T cells have been implicated both in the development of colitis and in the
maintenance of tolerance. Following antigen presentation to naïve CD4+T cells, they

25
differentiate into either Thl or Th2 effector cells (165; 166). The Thl phenotype is
associated with cellular responses modulated by proinflammatory cytokines, mainly IFN-y
and DC-derived IL-12 (167; 168). On the other hand, Th2 polarized responses are
characterized by the predominance of cytokines that favor humoral responses. Increased
production of IL-4 or IL-5 is an indicator of an ongoing Th2 response (166). The Thl/Th2
paradigm was first investigated in vitro using differentiated T cell clones. Usually, in vivo
both Thl and Th2 cells are present and there is a reciprocal regulation between cells of each
type (169). However, genetic and environmental factors may alter this equilibrium resulting
in a biased Thl or Th2 effector response (170).
Even though both polarized Thl and Th2 responses can be involved in the induction
of intestinal inflammation, this pathology is more commonly associated with Thl than with
Th2 responses (171). It has been speculated that the reason for this is that bacteria of the
normal flora, which play a critical role in the pathogenesis of inflammation, trigger the
production of IL-12 by DC, and therefore, the differentiation of naïve T cells into effector
Thl cells (153; 172-174). In line with this mechanism of pathogenesis, the cellular
mechanism underlying B. hyodysenteriae-induced colitis would be activation of intestinal
resident DC by B. hyodysenteriae and members of the normal flora. These activated DC
migrate to colonic lymph nodes, where they present antigens to naïve T cells in a
microenvironment influenced by IL-12 that favors the differentiation of naïve T cells into
Thl cells. Subsequently, bacterial-sensitized T cells would home to the colonic mucosa,
establishing a predominant Thl profile.
Cytokines produced in the context of a polarized Thl response, mainly IL-12 and.
IFN-y, inhibit the development of Th2 cells (175; 176). It has been proposed that the absence

26
of an active Thl response favors the differentiation of Th2 cells (168), while IL-10 and TGF-
P, produced by T reg cells, inhibit the production of IL-12, and therefore the development of
Thl cells (177; 178). During an inflammatory response driven by Thl cells, Th2 cells
migrating into the mucosa could block the effects of activated Thl effector cells responding
to members of the normal flora. This model could explain why vaccination with a B.
hyodysenteriae bacterin protects against swine dysentery. Basically, whereas challenge
would induce a Thl-type response, parenteral immunization would favor the development of
Th2 effector cells. Following infection, Th2 cells could migrate either to the colonic lymph
nodes and block the differentiation of naïve T cells into Thl cells, or directly to the mucosa
and inhibit effector Thl cells. Studies in mice have shown that infection with B.
hyodysenteriae increased IFN-y-producing cells in mesenteric lymph nodes and spleen, and
favored immunoglobulin isotype switching to IgG2a, which is characteristic of a Thl
response. However, cytokine analysis on vaccinated mice did not show increased production
of IL-4, which would indicate a predominant Th2 response. The lack of IL-4 and increased
production of IL-10 and TGF-(3 suggest that, alternatively, vaccination could stimulate
regulatory T cells (T reg), rather than inducing a Th-2-type response (179).
T reg cells have been identified in mice as memory CD4+ CD45RB10 cells that
provided protection against colitis induced by naïve CD4* CD45RBhl cells, when both were
transferred into a T cell-deficient SCID mouse (180). Functionally, T reg cells are
characterized by the production of TGF-P and IL-10, but not IL-4 (181). Additionally, T reg
cells have been shown to inhibit auto-reactive T cells, therefore, preventing autoimmunity
(82; 182). It could be hypothesized that immunization with B. hyodysenteriae stimulates T

27
reg cells, which would block the effect of Thl cells reacting to antigens of the normal flora
by producing TGF-P and IL-10.
yS T cell involvement in the control of inflammation
In regards to the involvement of yô T cells in intestinal inflammatory disorders, it
appears that they do not have a crucial role in the development of inflammation (153).
However, it has been suggested that they contribute to the process of epithelial restitution
that follows severe inflammation (72; 183; 184) and to downregulation of inflammatory
responses. Several models of infectious disease in which inflammation is present indicate
that a(3 T cells are increased initially, whereas y5 T cells are elevated later in the course of
the disease (185-188). The use of mice deficient in yô T cells indicated that they are
specifically implicated in the resolution of inflammatory damage. For instance, lesions
associated with infection with Listeria monocytogenes were more persistent in mice depleted
ofyô T cells (189; 190). It has been reported that during L. monocytogenes infection, yÔ T
cells shut down inflammation by inducing apoptosis of activated macrophages (191).
Another observation of these studies is that yô T cell responses are not specific for the
microorganism causing the disease (185). The fact that a subset of murine intraepithelial,
skin resident, yô T cells, (i.e., dendritic epidermal T cells (DETC)), were activated in vitro by
transformed or injured, but not healthy keratinocytes, suggested that stress-induced antigens,
rather that bacterial products might activate yô T cells (192; 193).
Although yS T cells are present both in the intestinal lamina propria and epithelium,
special emphasis has been given to the contribution of yô IEL to the resolution of
inflammation (194). yô IEL produce chemokines (e.g., RANTES or lymphotactin) for the
recruitment of lymphocytes and macrophages (e.g., MCP-1) (195). Also, neutrophil

28
recruitment into bacterial-induced pulmonary lesions was shown to be dependent on the
presence ofyô T cells, and correlated with protection from pneumonia (196; 197).
Additionally, yÔ IEL produce growth factors that promote proliferation and differentiation of
epithelial cells (198). The possibility of keratinocyte growth factor (KGF) being produced by
yS T cells was initially investigated in the context of the close interaction between epidermal
keratinocytes and DETC (198; 199). KGF belongs to the fibroblast growth factor family
(200). KGF was initially believed to be exclusively produced by stromal cells, mainly
fibroblasts, and act in a paracrine fashion on epithelial cells (201). Expression of KGF has
been detected in epithelial cells in vivo, and in DETC stimulated with anti-CD3 in vitro (202-
204). Also, murine DETC isolated from wounded skin expressed KGF mRNA, whereas
DETC obtained from healthy skin did not. In addition, skin lesions in mice lacking yô T cells
healed two to three days later than in wild-type mice (205).
A direct correlation between the production of KGF during intestinal epithelium
restitution by intraepithelial yô T cells had not been demonstrated until very recently. In a
murine model of dextran sodium sulfate (DSS) -induced colitis the number of yô IEL were
increased at the colonic mucosa during the process of tissue repair. These cells were
associated with damaged areas and were shown to produce KGF by reverse-transcriptase
polymerase chain reaction. In addition, both KGF- and TCRô-deficient mice had a delay in
recovery from DSS-induced colitis (206).
In summary, three basic mechanisms have been reported in regard to the participation
of yô T cells in mucosal protection. First, by recruiting cells which contribute to control of
infection. Second, by eliminating activated cells that otherwise would perpetuate
inflammation, such as activated macrophages or CD4+ T cells; and third, by producing

29
growth factors that contribute to regeneration of the intestinal epithelium and to restoration of
the epithelial cell barrier (27).
References
1. Harris, D. L., Alexander, T. J., Whipp, S. C., Robinson, I. M., Clock, R. D., & Matthews, P. J. (1978). Swine dysentery: studies of gnotobiotic pigs inoculated with Treponema hyodysenteriae, Bacteroides vulgatus, and Fusobacterium necrophorum. J. Am. Vet. Med. Assoc. 172: 468-471. 2. Joens, L. A., Robinson, I. M., Clock, R. D., & Matthews, P. J. (1981). Production of lesions in gnotobiotic mice by inoculation with Treponema hyodysenteriae. Infect. I mm un. 31: 504-506. 3. Hutto, D. L., & Wannemuehler, M. J. (1999). A comparison of the morphologic effects of Serpulina hyodysenteriae or its ^-hemolysin on the murine cecal mucosa. Vet. Pathol. 36: 412-422. 4. Hsu, T., Hutto. D. L., Minion, F. C., Zuerner, R. L., & Wannemuehler, M. J. (2001). Cloning of a ^-hemolysin gene of Brachysp/ra (Serpulina) hyodysenteriae and its expression in Escherichia coli. Infect. Immun. 69:706-711. 5. Li, X. (1998). Characterization of acute proinflammatory cytokine responses within the cecal mucosa following infection with Serpulina hyodysenteriae. MSc thesis. Iowa State University. 6. Kinyon, J. M., Harris, D. L., & Clock, R. D. (1977). Enteropathogenicity of various isolates of Treponema hyodysenteriae. Infect. Immun. 15:638-646. 7. Hontecillas, R., Wannemuelher, M. J., Zimmerman, D. R., Hutto, D. L., Wilson, J. H., Ahn, D. U., & Bassaganya-Riera, J. (2002). Nutritional regulation of porcine bacterial-induced colitis by conjugated linoleic acid. J. Nutr. 132: 2019-2027. 8. Waters, W. R., Sacco, R. E., Dorn, A. D., Hontecillas, R., Zuckermann, F. A., & Wannemuehler, M. J. (1999). Systemic and mucosal immune responses of pigs to parenteral immunization with a pepsin-digested Serpulina hyodysenteriae bacterin. Vet. Immunol. Immunopathol. 69: 75-87. 9. Waters, W. R., Hontecillas, R., Sacco, R. E., Zuckermann, F. A., Harkins, K. R., Bassaganya-Riera, J., & Wannemuehler, M. J. (2000). Antigen-specific proliferation of porcine CD8aa cells to an extracellular bacterial pathogen. Immunology 101: 333-341. 10. Waters, W. R., Pesch, B. A., Hontecillas, R., Sacco, R. E„ Zuckermann, F. A., & Wannemuehler, M. J. (1999). Cellular immune responses of pigs induced by vaccination with either a whole cell sonicate or pepsin-digested Brachyspira (Serpulina) hyodysenteriae bacterin. Vaccine 18: 711-719.

30
11. Bassaganya-Riera, J., Hontecillas, R., Zimmerman, D. R., & Wannemuehler, M. J. (2002). Long-term influence of lipid nutrition on the induction of CD8+ responses to viral or bacterial antigens. Vaccine 20: 1435-1444. 12. Bassaganya-Riera, J., Hontecillas, R., Zimmerman, D. R., & Wannemuehler, M. J. (2001). Dietary conjugated linoleic acid modulates phenotype and effector functions of porcine CD8+ lymphocytes. J. Nutr. 131: 2370-2377. 13. Yang, H., & Parkhouse, R. M. (1997). Differential expression of CD8 epitopes amongst porcine CD8-positive functional lymphocyte subsets [published erratum appears in Immunology 1997 Dec;92(4):587], Immunology 92: 45-52. 14. Yang, H., & Parkhouse, R. M. (1996). Phenotypic classification of porcine lymphocyte subpopulations in blood and lymphoid tissues. Immunology 89: 76-83. 15. Saalmuller, A., Pauly, T., & Pfaff, E. (1998). [Phenotypic and functional characterization of porcine T-lymphocytes]. Zentralbl. Chir. 123: 798-802. 16. Lunney, J. K.., & Pescovitz, M. D. (1987). Phenotypic and functional characterization of pig lymphocyte populations. Vet. Immunol. Immunopathol 17: 135-144. 17. Zuckermann, F. A. (1999). Extrathymic CD4/CD8 double positive T cells. Vet. Immunol. Immunopathol. 72: 55-66. 18. Zuckermann, F. A., & Husmann, R. J. (1996). Functional and phenotypic analysis of porcine peripheral blood CD4/CD8 double-positive T cells. Immunology 87: 500-512. 19. Pescovitz, M. D., Sakopoulos, A. G., Gaddy, J. A., Husmann, R. J., & Zuckermann, F. A. (1994) Porcine peripheral blood CD4+/CD8+ dual expressing T-cells. Vet. Immunol. Immunopathol. 43: 53-62. 20. Reddehase, M. J., Saalmuller, A., & Hirt, W. (1991). yô T-lymphocyte subsets in swine. Curr. Top. Microbiol. Immunol. 173: 113-117. 21. Saalmuller, A., Hirt, W., & Reddehase, M. J. (1990). Porcine yô T lymphocyte subsets differing in their propensity to home to lymphoid tissue. Eur. J. Immunol. 20:2343-2346. 22. Hirt, W., Saalmuller, A., & Reddehase, M. J. (1990). Distinct yÔ T cell receptors define two subsets of circulating porcine CD2*CD4"CD8" T lymphocytes. Eur. J. Immunol. 20: 265-269. 23. Chien, Y. H., Jores, R., & Crowley, M. P. (1996). Recognition by yÔ T cells. Ann. Rev. Immunol. 14: 511-532. 24. Mackay, C. R., & Hein, W. R. (1991). Marked variations in yô T cell numbers and distribution throughout the life of sheep. Curr. Top. Microbiol. Immunol. 173: 107-111. 25. Hein, W. R., & Mackay, C. R. (1991). Prominence of yô T cells in the ruminant immune system. Immunol. Today 12: 30-34. 26. Carr, M. M., Howard, C. J., Sopp, P., Manser, J. M., & Parsons, K. R. (1994). Expression on porcine yô lymphocytes of a phylogenetically conserved surface antigen previously restricted in expression to ruminant yô T lymphocytes. Immunology 81:36-40. 27. Hayday, A. C. (2000). yÔ cells: a right time and a right place for a conserved third way of protection. Ann. Rev. Immunol. 18:975-1026. 28. Janeway, C. A., Travers, P., Walport, M., and Capra, J.D. (2001). Immunobiology: the immune system in health and disease. Elsevier Science London, New York.

31
29. von Boehmer, H. (1988). The developmental biology of T lymphocytes. Ann. Rev. Immunol. 6: 309-326. 30. Fehling, H. J., Gilfillan, S., & Ceredig, R. (1999). ap/yô lineage commitment in the thymus of normal and genetically manipulated mice. Adv. Immunol. 71: 1-76. 31. Poussier, P., & Julius, M. (1994). Thymus independent T cell development and selection in the intestinal epithelium. Ann. Rev. Immunol. 12: 521-553. 32. Rocha, B., Guy-Grand, D., & Vassalli, P. (1995) Extrathymic T cell differentiation. Curr. Opin. Immunol. 7: 235-242. 33. Klein, J. R. (1996). Whence the intestinal intraepithelial lymphocyte? J. Exp. Med. 184: 1203-1206. 34. Saito, T., Hochstenbach, F., Marusic-Galesic, S., Kruisbeek, A. M., Brenner, M„ & Germain, R. N. (1988). Surface expression of only yô and/or a(3 T cell receptor heterodimers by cells with four (a, (3, y, Ô) functional receptor chains. J. Exp. Med. 168: 1003-1020. 35. Born, W., Cady, C., Jones-Carson, J., Mukasa, A., Lahn, M., & O'Brien, R. (1999). Immunoregulatory functions ofyS T cells. Adv. Immunol. 71: 77-144. 36. Petrie, H. T., Scollay, R., & Shortman, K. (1992). Commitment to the T cell receptor-a(3 or -yô lineages can occur just prior to the onset of CD4 and CD8 expression among immature thymocytes. Eur. J. Immunol. 22: 2185-2188. 37. Kang, J., Fehling, H. J., Laplace, C., Malissen, M., Cado, D., & Raulet, D. H. (1998). T cell receptor y gene regulatory sequences prevent the function of a novel TCRy/pTa pre-T cell receptor. Immunity 8: 713-721. 38. Michie, A., & Zuniga-Pflucker, J. (2002). Regulation of thymocyte differentiation: pre-TCR signals and beta-selection. Semin. Immuno.l 14: 311. 39. Hunig, T., Torres-Nagel, N., Mehling, B., Park, H. J., & Herrmann, T. (2001). Thymic development and repertoire selection: the rat perspective. Immunol. Rev. 184: 7-19. 40. Kisielow, P., & von Boehmer, H. (1995). Development and selection of T cells: facts and puzzles. Adv. Immunol. 58: 87-209. 41. Jameson, S. C., Hogquist, K. A., & Bevan, M. J. (1995). Positive selection of thymocytes. Ann. Rev. Immunol. 13: 93-126. 42. Butcher, E. C., Williams, M., Youngman, K., Rott, L., & Briskin, M. (1999). Lymphocyte trafficking and regional immunity. Adv. Immunol. 72: 209-253. 43. Itohara, S., Farr, A. G., Lafaille, J. J., Bonneville, M., Takagaki, Y., Haas, W., & Tonegawa, S. (1990). Homing of a gamma delta thymocyte subset with homogeneous T-cell receptors to mucosal epithelia. Nature 343: 754-757. 44. Haas, W., Pereira, P., & Tonegawa, S. (1993). yô cells. Ann. Rev. Immunol. 11: 637-685. 45. Steinman, R. M., Pack, M., & Inaba, K. (1997). Dendritic cells in the T-cell areas of lymphoid organs. Immunol. Rev. 156:25-37. 46. Schild, H., Mavaddat, N., Litzenberger, C., Ehrich, E. W., Davis, M. M., Bluestone, J. A., Matis, L., Draper, R. K., & Chien, Y. H. (1994). The nature of major histocompatibility complex recognition by yô T cells. Cell 76:29-37.

32
47. Girardi, M., Oppenheim, D. E., Steele, C. R., Lewis, J. M., Glusac, E., Filler, R., Hobby, P., Sutton, B., Tigelaar, R. E., & Hayday, A. C. (2001). Regulation of cutaneous malignancy by yô T cells. Science 294: 605-609. 48. Spada, F. M., Grant, E. P., Peters, P. J., Sugita, M., Melian, A., Leslie, D. S., Lee, H. K.., van Donselaar, E., Hanson, D. A., Krensky, A. M., Majdic, O., Porcelli, S. A., Morita, C. T., & Brenner, M. B. (2000). Self-recognition of CD1 by yS T cells: implications for innate immunity. J. Exp. Med. 191: 937-948. 49. Morita, C. T., Beckman, E. M., Bukowski, J. F., Tanaka, Y., Band, H., Bloom, B. R., Golan, D. E.. & Brenner, M. B. (1995). Direct presentation of nonpeptide prenyl pyrophosphate antigens to human yS T cells. Immunity 3: 495-507. 50. Bjorkman, P. J. (1997). MHC restriction in three dimensions: a view of T cell receptor/ligand interactions. Cell 89: 167-170. 51. Johnson, R. M., Lancki, D. W., Sperling, A. I., Dick, R. F., Spear, P. G., Fitch, F. W., & Bluestone, J. A. (1992). A murine CD4", CD8" T cell receptor-yS T lymphocyte clone specific for herpes simplex virus glycoprotein I. J. Immunol. 148: 983-8 52. Tanaka, Y., Morita, C. T., Nieves, E., Brenner, M. B., & Bloom, B. R. (1995). Natural and synthetic non-peptide antigens recognized by human yô T cells. Nature 375: 155-8 53. Schoel, B., Sprenger, S.. & Kaufmann, S. H. (1994). Phosphate is essential for stimulation of Vy9 V52 T lymphocytes by mycobacterial low molecular weight ligand. Eur. J. Immunol. 24: 1886-92. 54. Constant, P., Davodeau, F., Peyrat, M. A., Poquet, Y., Puzo, G.. Bonneville, M., & Fournie, J. J. (1994). Stimulation of human yô T cells by nonpeptidic mycobacterial ligands. Science 264: 267-70. 55. Feurle, J., Espinosa, E., Eckstein, S., Pont, F., Kunzmann, V., Fournie, J. J., Herderich, M., & Wilhelm, M. (2002). Escherichia coli produces phosphoantigens activating human yÔ T cells. J. Biol. Chem. 277: 148-154. 56. Groh, V., Steinle, A., Bauer, S., & Spies, T. (1998). Recognition of stress-induced MHC molecules by intestinal epithelial yô T cells. Science 279: 1737-40 57. Janeway, C. A., Jr., Jones, B., & Hayday, A. (1988). Specificity and function of T cells bearing yÔ receptors. Immunol. Today 9: 73-76. 58. Bromley, S. K., Burack, W. R., Johnson, K. G., Somersalo, K., Sims, T. N., Sumen, C., Davis, M. M., Shaw, A. S., Allen, P. M., & Dustin, M. L. (2001). The immunological synapse. Ann. Rev. Immunol. 19: 375-396. 59. Weiss, A., & Littman, D. R. (1994). Signal transduction by lymphocyte antigen receptors. Cell 76: 263-274. 60. Hayes, S. M., & Love, P. E. (2002). Distinct structure and signaling potential of the yô TCR complex. Immunity 16: 827-838. 61. Garcia, K. C., Teyton, L., & Wilson, I. A. (1999). Structural basis of T cell recognition. Ann. Rev. Immunol. 17:369-397. 62. Loh, E. Y., Lanier, L. L., Turck, C. W., Littman, D. R., Davis, M. M., Chien, Y. H., & Weiss, A. (1987). Identification and sequence of a fourth human T cell antigen receptor chain. Nature 330: 569-572.

33
63. Allison, T. J., Winter, C. C„ Fournie, J. J., Bonneville, M., & Garboczi, D. N. (2001). Structure of a human yô T-cell antigen receptor. Nature 411: 820-824. 64. Kronenberg, M., Siu, G., Hood, L. E., & Shastri, N. (1986). The molecular genetics of the T-cell antigen receptor and T-cell antigen recognition. Annu. Rev. Immunol. 4: 529-591. 65. Thome, A., Saalmuller, A., & Pfaff E. (1993). Molecular cloning of porcine T cell receptor a, (3, y and 5 chains using polymerase chain reaction fragments of the constant regions. Eur. J. Immunol. 23: 1005-1010. 66. Tonegawa, S. (1988). Nobel lecture in physiology or medicine-1987. Somatic generation of immune diversity. In Vitro Cell. Dev. Biol. 24: 253-265. 67. Wu, T. T., & Kabat, E. A. (1970). An analysis of the sequences of the variable regions of Bence Jones proteins and myeloma light chains and their implications for antibody complementarity. J. Exp. Med. 132: 211-250. 68. Davies, D. R., & Cohen, G. H. (1996). Interactions of protein antigens with antibodies. Proc. Natl. Acad. Sci. USA 93: 7-12. 69. Wilson, I. A., & Stanfield, R. L. (1994). Antibody-antigen interactions: new structures and new conformational changes. Curr. Opin. Struct. Biol. 4: 857-867. 70. Garcia, K. C., Degano, M., Stanfield, R. L., Brunmark, A., Jackson, M. R., Peterson, P. A., Teyton, L., & Wilson, I. A. (1996). An aP T cell receptor structure at 2.5 A and its orientation in the TCR-MHC complex. Science 274: 209-219. 71. Rock, E. P., Sibbald, P. R., Davis, M. M., & Chien, Y. H. (1994). CDR3 length in antigen-specific immune receptors. J. Exp. Med. 179: 323-328. 72. Davis, M. M.. & Bjorkman, P. J. (1988). T-cell antigen receptor genes and T-cell recognition. Nature 334: 395-402. 73. Vincent, M. S., Roessner, K., Sellati, T., Huston, C. D., Sigal, L. H., Behar, S. M., Radolf, J. D., & Budd, R. C. (1998). Lyme arthritis synovial yS T cells respond to Borrelia burgdorferi lipoproteins and lipidated hexapeptides. J. Immunol. 161: 5762-5771. 74. Allison, T. J., & Garboczi, D. N. (2002). Structure of yô T cell receptors and their recognition of non-peptide antigens. Mol. Immunol. 38: 1051-1061. 75. Davis, M. M., & Chien, Y. (1995). Issues concerning the nature of antigen recognition by «P and yS T-cell receptors. Immunol. Today. 16: 316-318. 76. Schild, H., & Chien, Y. H. (1994). The recognition of MHC molecules by yS T cells. Behring Inst. Mitt: 113-123. 77. Havran, W. L., Chien, Y. H., & Allison, J. P. (1991). Recognition of self antigens by skin-derived T cells with invariant yS antigen receptors. Science 252: 1430-1432. 78. Davodeau, F., Peyrat, M. A., Hallet, M. M., Houde, L, Vie, H., & Bonneville, M. (1993). Peripheral selection of antigen receptor junctional features in a major human yô subset. Eur. J. Immunol. 23: 804-808. 79. Davodeau, F., Peyrat, M. A., Houde, I., Hallet, M. M., De Libera, G., Vie, H., & Bonneville, M. (1993). Surface expression of two distinct functional antigen receptors on human yô T cells. Science 260: 1800-1802. 80. Davodeau, F., Peyrat, M. A., Hallet, M. M., Gaschet, J., Houde, L, Vivien, R., Vie, H., & Bonneville, M. (1993). Close correlation between Daudi and mycobacterial antigen

34
recognition by human yS T cells and expression of V9JPCly/V2DJC5-encoded T cell receptors. J. Immunol 151: 1214-1223. 81. Thome, M., Hirt, W„ Pfaff, E., Reddehase, M. J., & Saalmuller, A. (1994). Porcine T-cell receptors: molecular and biochemical characterization. Vet. Immunol. Immunopathol 43: 13-18.
82. Mosmann, T. R., & Sad, S. (1996). The expanding universe of T-cell subsets: Thl, Th2 and more. Immunol. Today 17: 138-146. 83. Itoh, M., Takahashi, T., Sakaguchi, N., Kuniyasu, Y„ Shimizu, J., Otsuka, F., & Sakaguchi, S. (1999). Thymus and autoimmunity: production of CD25+CD4+ naturally anergic and suppressive T cells as a key function of the thymus in maintaining immunologic self-tolerance. J. Immunol. 162: 5317-5326. 84. Veillette, A., Sleckman, B. P., Ratnofsky, S., Bolen, J. B., & Burakoff, S. J. (1990). The cytoplasmic domain of CD4 is required for stable association with the lymphocyte-specific tyrosine protein kinase p561ck. Eur. J. Immunol. 20: 1397-1400. 85. Veillette, A., Bookman, M. A., Horak, E. M., Samelson, L. E., & Bolen, J. B. (1989). Signal transduction through the CD4 receptor involves the activation of the internal membrane tyrosine-protein kinase p561ck. Nature 338: 257-259. 86. Veillette, A., Bookman, M. A., Horak, E. M., & Bolen, J. B. (1988). The CD4 and CD8 T cell surface antigens are associated with the internal membrane tyrosine-protein kinase p561ck. Cell 55: 301-308. 87. Saalmuller, A. (1998). Antigen-specific immune response of porcine T lymphocytes to various pathogens. Rev. Sci. Tech. 17: 71-83. 88. Samsom, J. N., de Bruin, T. G., Voermans, J. J., Meulenberg, J. J., Pol, J. M„ & Bianchi, A. T. (2000). Changes of leukocyte phenotype and function in the broncho-alveolar lavage fluid of pigs infected with porcine reproductive and respiratory syndrome virus: a role for CD8+ cells. J. Gen. Virol. 81:497-505. 89. Saalmuller, A., Pauly, T., Hohlich, B. J., & Pfaff, E. (1999). Characterization of porcine T lymphocytes and their immune response against viral antigens. J. Biotechnol. 73: 223-233. 90. Sundt, T. M., 3rd, Am, J. S., & Sachs, D. H. (1992). Patterns of T cell-accessory cell interaction in the generation of primary alloresponses in the pig. Transplantation 54:911-916. 91. Ober, B. T., Summerfield, A., Mattlinger, C., Wiesmuller, K. H., Jung, G., Pfaff, E., Saalmuller, A., & Rziha, H. J. (1998). Vaccine-induced, pseudorabies virus-specific, extrathymic CD4+CD8+ memory T-helper cells in swine. J. Virol. 72:4866-4873. 92. Hernandez, J., Reyes-Leyva, J., Zenteno, R„ Ramirez, H., Hernandez-Jauregui, P., & Zenteno, E. (1998). Immunity to porcine rubulavirus infection in adult swine. Vet. Immunol. Immunopathol. 64: 367-381. 93. Shimizu, M., Yamada, S., Kawashima, K., Ohashi, S., Shimizu, S., & Ogawa, T. (1996). Changes of lymphocyte subpopulations in pigs infected with porcine reproductive and respiratory syndrome (PRRS) virus. Vet. Immunol. Immunopathol. 50: 19-27.

35
94. Summerfield, A., McNeilly, F., Walker, I., Allan, G., Knoetig, S. M., & McCullough, K. C. (2001). Depletion of CD4+ and CD8hlgh+ T-cells before the onset of viraemia during classical swine fever. Vet. Immunol. Immunopathol. 78: 3-19. 95. Markowska-Daniel, L, Pejsak, Z., Winnicka, A., & Collins, R. A. (1999). Phenotypic analysis of peripheral leukocytes in piglets infected with classical swine fever virus. Res. Vet. Sci. 67: 53-57. 96. Sarli, G., Mandrioli, L., Laurenti, M., Sidoli, L., Cerati, C., Rolla, G., & Marcato, P. S. (2001). Immunohistochemical characterisation of the lymph node reaction in pig post-weaning multisystemic wasting syndrome (PMWS). Vet. Immunol. Immunopathol. 83: 53-67. 97. Solano Aguilar, G. I., Beshah, E., Vengroski, K. G., Zarlenga, D., Jauregui, L., Cosio, M., Douglass, L. W., Dubey, J. P., & Lunney, J. K. (2001). Cytokine and lymphocyte profiles in miniature swine after oral infection with Toxoplasma gondii oocysts. Int. J. Parasitol. 31: 187-195. 98. Lopez Fuertes, L„ Domenech, N., Alvarez, B., Ezquerra, A., Dominguez, J., Castro, J. M., & Alonso, F. (1999). Analysis of cellular immune response in pigs recovered from porcine respiratory and reproductive syndrome infection. Virus. Res. 64: 33-42. 99. Zuckermann, F. A., Pescovitz, M. D., Aasted, B., Dominguez, J., Trebichavsky, I., Novikov, B., Valpotic, I., Nielsen, J., Am, S., Sachs, D. H., Lunney, J. K., Boyd, P., Walker, J., Lee, R., Davis, W. C.. Barbosa, I. R., & Saalmuller, A. (1998). Report on the analyses of mAb reactive with porcine CD8 for the second international swine CD workshop. Vet. Immunol. Immunopathol. 60: 291-303. 100. De Maria, A., Malnati, M., Moretta, A., Pende, D., Bottino, C., Casorati, G., Cottafava, F., Melioli, G., Mingari, M. C., Migone, N., & et al. (1987). CD3+4"8"WT3V(T cell receptor Y*) cells and other unusual phenotypes are frequently detected among spontaneously interleukin 2-responsive T lymphocytes present in the joint fluid in juvenile rheumatoid arthritis. A clonal analysis. Eur. J. Immunol. 17: 1815-1819. 101. Bagot, M., Echchakir, H., Mami-Chouaib, F., Delfau-Larue, M. H., Charue, D., Bemheim, A., Chouaib, S., Boumsell, L., & Bensussan, A. (1998). Isolation of tumor-specific cytotoxic CD4+ and CD4+CD8dim+ T-cell clones infiltrating a cutaneous T-cell lymphoma. Blood 91:4331-4341. 102. Pescovitz, M. D., Hsu, S. M., Katz, S. L, Lunney, J. K., Shimada, S., & Sachs, D. H. (1990). Characterization of a porcine CD 1-specific mAb that distinguishes CD4/CD8 double-positive thymic from peripheral T lymphocytes. Tissue Antigens 35: 151-156. 103. Sinkora, M., Sinkora, J., Rehakova, Z., Splichal, L, Yang, H., Parkhouse, R. M., & Trebichavsk, I. (1998). Prenatal ontogeny of lymphocyte subpopulations in pigs. Immunology 95: 595-603. 104. Shiue, L., Gorman, S. D., & Pames, J. R. (1988). A second chain of human CD8 is expressed on peripheral blood lymphocytes. J. Exp. Med. 168: 1993-2005. 105. Zamoyska, R. (1994). The CD8 coreceptor revisited: one chain good, two chains better. Immunity 1: 243-246.

36
106. Janeway, C. A., Jr. (1992). The T cell receptor as a multicomponent signalling machine: CD4/CD8 coreceptors and CD45 in T cell activation. Ann. Rev. Immunol. 10: 645-674. 107. Suss, G., & Shortman, K. (1996). A subclass of dendritic cells kills CD4 T cells via Fas/Fas-ligand-induced apoptosis. J. Exp. Med. 183: 1789-1796. 108. Moebius, U., Kober, G., Griscelli, A. L., Hercend, T., & Meuer, S. C. (1991). Expression of different CD8 iso forms on distinct human lymphocyte subpopulations. Eur. J. Immunol. 21: 1793-1800. 109. Garcia, K. C., Scott, C. A., Brunmark, A., Carbone, F. R„ Peterson, P. A., Wilson, I. A., & Teyton, L. (1996). CD8 enhances formation of stable T-cell receptor/MHC class I molecule complexes. Nature 384: 577-581. 110. Leishman, A. J., Naidenko, O. V.. Attinger, A., Koning, F., Lena, C. J., Xiong, Y., Chang, H. C„ Reinherz, E., Kronenberg, M., & Cheroutre, H. (2001). T cell responses modulated through interaction between CD8aa and the nonclassical MHC class I molecule, TL. Science 294: 1936-1939. 111. Kern, P., Hussey, R. E., Spoerl, R., Reinherz, E. L., & Chang, H. C. (1999). Expression, purification, and functional analysis of murine ectodomain fragments of CD8aa and CD8a(3 dimers. J. Biol. Chem. 274: 27237-27243. 112. Luescher, I. F., Vivier, E., Layer, A., Mahiou, J., Godeau, F„ Malissen, B., & Romero, P. (1995). CD8 modulation of T-cell antigen receptor-ligand interactions on living cytotoxic T lymphocytes. Nature 373: 353-356. 113. Ravichandran, K. S., Pratt, J. C., Sawasdikosol, S., Irie, H. Y., & Burakoff, S. J. (1995). Coreceptors and adapter proteins in T-cell signaling. Ann. N. Y. Acad. Sci. 766: 117-133. 114. Irie, H. Y., Ravichandran, K. S., & Burakoff, S. J. (1995). CD8 p chain influences CD8 a chain-associated Lck kinase activity. J. Exp. Med. 181: 1267-1273. 115. Arcaro, A., Grégoire, C., Bakker, T. R., Baldi, L., Jordan, M., Goffin, L., Boucheron, N., Wurm, F., van der Merwe, P. A., Malissen, B., & Luescher, I. F. (2001). CD8P endows CD8 with efficient coreceptor function by coupling T cell receptor/CD3 to raft-associated CD8/p56(lck) complexes. J. Exp. Med. 194: 1485-1495. 116. Arcaro, A., Grégoire, C., Boucheron, N„ Stotz, S., Palmer, E., Malissen, B., & Luescher, I. F. (2000). Essential role of CD8 palmitoylation in CD8 coreceptor function. J. Immunol 165:2068-2076. 117. Gorman, S. D., Sun, Y. H., Zamoyska, R., & Pames, J. R. (1988). Molecular linkage of the Ly-3 and Ly-2 genes. Requirement of Ly-2 for Ly-3 surface expression. J. Immunol. 140: 3646-3653. 118. Ellmeier, W., Sawada, S., & Littman, D. R. (1999). The regulation of CD4 and CD8 coreceptor gene expression during T cell development. Ann. Rev. Immunol. 17: 523-554. 119. Ellmeier, W., Sunshine, M. J., Losos, K., & Littman, D. R. (1998). Multiple developmental stage-specific enhancers regulate CD8 expression in developing thymocytes and in thymus-independent T cells. Immunity 9: 485-496. 120. Ellmeier, W., Sunshine, M. J., Losos, K., Hatam, F., & Littman, D. R. (1997). An enhancer that directs lineage-specific expression of CD8 in positively selected thymocytes and mature T cells. Immunity 7: 537-547.

37
121. Zhang, X. L., Seong, R., Piracha, R., Larijani, M., Heeney, M., Pâmes, J. R., & Chamberlain, J. W. (1998). Distinct stage-specific cis-active transcriptional mechanisms control expression of T cell coreceptor CD8a at double- and single-positive stages of thymic development. J. Immunol. 161: 2254-2266. 122. Gao, M. H., Walz. M., & Kavathas, P. B. (1996). Post-transcriptional regulation associated with control of human CD8A expression of CD4+ T cells. Immunogenetics 45: 130-135. 123. Paliard, X., Malefijt, R. W., de Vries, J. E., & Spits, H. (1988). Interleukin-4 mediates CD8 induction on human CD4+ T-cell clones. Nature 335: 642-644. 124. Ottenhoff, T. H., Elfe rink, D. G., Klatser, P. R., & de Vries, R. R. (1986). Cloned suppressor T cells from a lepromatous leprosy patient suppress Mycobacterium leprae reactive helper T cells. Nature 322: 462-464. 125. Asseman, C., Mauze, S.. Leach, M. W., Coffman, R. L., & Powrie, F. (1999). An essential role for interleukin 10 in the function of regulatory T cells that inhibit intestinal inflammation. J. Exp. Med. 190: 995-1004. 126. Trinchieri, G. (2001). Regulatory role of T cells producing both interferon y and interleukin 10 in persistent infection. J Exp Med 194: F53-57. 127. Rodriguez-Carreno, M. P., Lopez-Fuertes, L., Revilla, C., Ezquerra, A., Alonso. F., & Dominguez, J. (2002). Phenotypic characterization of porcine IFN-y-producing lymphocytes by flow cytometry. J. Immunol. Methods. 259: 171-179. 128. de Bruin, T. G., van Rooij, E. M., de Visser, Y. E., Voermans, J. J., Samsom, J. N., Kimman, T. G., & Bianchi, A. T. (2000). Discrimination of different subsets of cytolytic cells in pseudorabies virus-immune and naive pigs. J. Gen. Virol. 81: 1529-1537. 129. De Bruin, T. G., Van Rooij, E. M., De Visser, Y. E., & Bianchi, A. T. (2000). Cytolytic function for pseudorabies virus-stimulated porcine CD4+ CD8dull+ lymphocytes. Viral Immunol. 13: 511-520. 130. Saito, H., Kranz, D. M., Takagaki, Y., Hayday, A. C., Eisen, H. N., & Tonegawa, S. (1984). Complete primary structure of a heterodimeric T-cell receptor deduced from cDNA sequences. Nature 309: 757-762. 131. Saito, H., Kranz, D. M., Takagaki, Y., Hayday, A. C., Eisen, H. N., & Tonegawa, S. (1984). A third rearranged and expressed gene in a clone of cytotoxic T lymphocytes. Nature 312:36-40. 132. Hayday, A. C., Saito, H., Gillies, S. D., Kranz, D. M., Tanigawa, G., Eisen, H. N., & Tonegawa, S. (1985). Structure, organization, and somatic rearrangement of T cell yô. Cell 40: 259-269. 133. Davis, W. C., Zuckermann, F. A., Hamilton, M. J., Barbosa, J. I., Saalmuller, A., Binns, R. M., & Licence, S. T. (1998). Analysis of monoclonal antibodies that recognize yS T/null cells. Vet Immunol. Immunopathol. 60: 305-316. 134. Davis, W. C., Haverson, K., Saalmuller, A., Yang, H., Lunney, J. K., Hamilton, M. J., & Pescovitz, M. D. (2001). Analysis of monoclonal antibodies reacting with molecules expressed on y5 T-cells. Vet. Immunol. Immunopathol. 80: 53-62.

38
135. Yang, H., & Parkhouse, R. M. (2000). Characterization of the porcine yô T-cell receptor structure and cellular distribution by monoclonal antibody PPT27. Immunology 99: 504-509. 136. de Bruin, M. G., van Rooij, E. M., Voermans, J. J., de Visser, Y. E., Bianchi, A. T., & Kimman, T. G. (1997). Establishment and characterization of porcine cytolytic cell lines and clones. Vet. Immunol. Immunopathol. 59: 337-347. 137. Whyte, A., Haskard, D. O., & Binns, R. M. (1994). Infiltrating yô T-cells and selectin endothelial ligands in the cutaneous phytohaemagglutinin-induced inflammatory reaction.. Vet. Immunol. Immunopathol. 41: 31-40 138. Whyte, A., Licence, S. T., Robinson, M. K., & van der Lienden, K. (1996). Lymphocyte subsets and adhesion molecules in cutaneous inflammation induced by inflammatory agonists: correlation between E-selectin and yô TcR+ lymphocytes. Lab Invest 75: 439-449. 139. Hershberg, R. M., & Mayer, L. F. (2000). Antigen processing and presentation by intestinal epithelial cells - polarity and complexity. Immunol. Today 21: 123-128. 140. Shanahan, F. (1997). A gut reaction: lymphoepithelial communication in the intestine. Science 275: 1897-1898. 141. Kraehenbuhl, J. P., & Neutra, M. R. (1992). Molecular and cellular basis of immune protection of mucosal surfaces. Physiol. Rev. 72: 853-879. 142. Camerini, V., Panwala, C., & Kronenberg, M. (1993). Regional specialization of the mucosal immune system. Intraepithelial lymphocytes of the large intestine have a different phenotype and function than those of the small intestine. J. Immunol. 151: 1765-1776. 143. Beagley, K. W., Fujihashi, K., Lagoo, A. S., Lagoo-Deenadaylan, S„ Black, C. A., Murray, A. M., Sharmanov, A. T., Yamamoto, M., McGhee, J. R., Elson, C. O., & et al. (1995). Differences in intraepithelial lymphocyte T cell subsets isolated from murine small versus large intestine. J. Immunol. 154: 5611-5619. 144. Hayday, A., Theodoridis, E„ Ramsburg, E., & Shires, J. (2001). Intraepithelial lymphocytes: exploring the Third Way in immunology. Nat. Immunol. 2: 997-1003. 145. Czerkinsky, C., Anjuere, F., McGhee, J. R., George-Chandy, A., Holmgren, J., Kieny, M. P., Fujiyashi, K., Mestecky, J. F., Pierrefite-Carle, V., Rask, C., & Sun, J. B. (1999). Mucosal immunity and tolerance: relevance to vaccine development. Immunol. Rev. 170: 197-222. 146. Lin, T., Matsuzaki, G., Kenai, H., & Nomoto, K. (1994). Progenies of fetal thymocytes are the major source of CD4 CD8 aa intestinal intraepithelial lymphocytes early in ontogeny. Eur. J. Immunol. 24: 1785-1791. 147. Rocha, B., Vassalli, P., & Guy-Grand, D. (1991). The V(3 repertoire of mouse gut homodimeric a CD8+ intraepithelial T cell receptor a(3 + lymphocytes reveals a major extrathymic pathway of T cell differentiation. J. Exp. Med. 173:483-486. 148. Rocha, B., Vassalli, P., & Guy-Grand, D. (1994). Thymic and extrathymic origins of gut intraepithelial lymphocyte populations in mice. J. Exp. Med 180:681-686. 149. Guy-Grand, D., Rocha, B., Mintz, P., Malassis-Seris, M,, Selz, F., Malissen, B., & Vassalli, P. (1994). Different use of T cell receptor transducing modules in two populations of gut intraepithelial lymphocytes are related to distinct pathways ofT cell differentiation. J. Exp. Med. 180: 673-679.

39
150. Pabst, R., & Rothkotter, H. J. (1999). Postnatal development of lymphocyte subsets in different compartments of the small intestine of piglets. Vet. Immunol. Immunopathol. 72: 167-173. 151. Vega-Lopez, M. A., Telemo, E., Bailey, M., Stevens, K., & Stokes, C. R. (1993). Immune cell distribution in the small intestine of the pig: immunohistological evidence for an organized compartmentalization in the lamina propria. Vet. Immunol. Immunopathol. 37:49-60. 152. Vega-Lopez, M. A., Arenas-Contreras, G., Bailey, M., Gonzalez-Pozos, S., Stokes, C. R., Ortega, M. G., & Mondragon-Flores, R. (2001). Development of intraepithelial cells in the porcine small intestine. Dev. Immunol. 8: 147-158. 153. Solano-Aguilar, G. I., Vengroski, K. G., Beshah, E„ Douglass, L. W., & Lunney, J. K. (2001). Characterization of lymphocyte subsets from mucosal tissues in neonatal swine. Dev. Comp. Immunol. 25: 245-263. 154. Strober, W., Fuss, I. J., & Blumberg, R. S. (2002). The immunology of mucosal models of inflammation. Ann. Rev. Immunol. 20:495-549. 155. Matzinger, P. (2002). The danger model: a renewed sense of self. Science 296: 301-305. 156. Gallucci, S., & Matzinger, P. (2001). Danger signals: SOS to the immune system. Curr. Opin. Immunol. 13: 114-119. 157. Hayashi, F., Smith, K. D., Ozinsky, A.. Hawn, T. R., Yi, E. C., Goodlett, D. R., Eng, J. K., Akira, S., Underbill, 0. M., & Aderem, A. (2001). The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 410: 1099-1103. 158. Schnare, M.. Holt, A. C., Takeda, K.. Akira, S., & Medzhitov, R. (2000). Recognition of CpG DNA is mediated by signaling pathways dependent on the adaptor protein MyD88. Curr. Biol. 10: 1139-1142. 159. Philpott, D. J., Girardin, S. E., & Sansonetti, P. J. (2001). Innate immune responses of epithelial cells following infection with bacterial pathogens. Curr. Opin. Immunol. 13:410-416. 160. Cereijido, M., Valdes, J., Shoshani, L., & Contreras, R. G. (1998). Role of tight junctions in establishing and maintaining cell polarity. Ann. Rev. Physiol. 60: 161-177. 161. Kazmierczak, B. I., Mostov, K., & Engel, J. N. (2001). Interaction of bacterial pathogens with polarized epithelium. Ann. Rev. Microbiol. 55:407-435. 162. Mitic, L. L., & Anderson, J. M. (1998) Molecular architecture of tight junctions. Ann. Rev. Physiol. 60: 121-142. 163. Hershberg, R. M. (2002). The epithelial cell cytoskeleton and intracellular trafficking. V. Polarized compartmentalization of antigen processing and Toll-like receptor signaling in intestinal epithelial cells. Am. J. Physiol. Gastrointest Liver. Physiol 283: G833-839. 164. Gewirtz, A. T., Navas, T. A., Lyons, S., Godowski, P. J., & Madara, J. L. (2001). Cutting edge: bacterial flagellin activates basolaterally expressed TLR5 to induce epithelial proinflammatory gene expression. J. Immunol. 167: 1882-1885. 165. Kim, H. S., & Berstad, A. (1992). Experimental colitis in animal models. Scand J. Gastroenterol. 27: 529-537.

40
166. Mosmann, T. R., Cherwinski, H., Bond, M. W., Giedlin, M. A., & Coffman, R. L. (1986). Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. J. Immunol. 136: 2348-2357. 167. Mosmann, T. R., & Coffman, R. L. (1989). TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Ann. Rev. Immunol. 7: 145-173. 168. Mosmann, T. R., & Moore, K. W. (1991). The role of IL-10 in crossregulation of TH1 and TH2 responses. Immunol. Today 12: A49-53. 169. Moser, M., & Murphy, K.. M. (2000). Dendritic cell regulation of TH1-TH2 development. Nat. Immunol. 1: 199-205. 170. Rissoan, M. C., Soumelis, V., Kadowaki, N., Grouard, G., Briere, F., de Waal Malefyt, R., & Liu, Y. J. (1999). Reciprocal control of T helper cell and dendritic cell differentiation. Science 283: 1183-1186. 171. Constant, S. L., & Bottomly, K. (1997). Induction of Thl and Th2 CD4+ T cell responses: the alternative approaches. Ann. Rev. Immunol. 15:297-322. 172. Powrie, F. (1995). T cells in inflammatory bowel disease: protective and pathogenic roles. Immunity 3: 171-174. 173. O'Garra, A. (1998). Cytokines induce the development of functionally heterogeneous T helper cell subsets. Immunity 8: 275-283. 174. Trinchieri. G. (1995). Interleukin-12: a proinflammatory cytokine with immunoregulatory functions that bridge innate resistance and antigen-specific adaptive immunity. Ann. Rev. Immunol. 13: 251-276. 175. Maldonado-Lopez, R.. De Smedt, T., Michel, P., Godfroid, J.. Pajak, B., Heirman, C., Thielemans, K., Leo, O., Urbain. J., & Moser, M. (1999). CD8a+ and CD8a" subclasses of dendritic cells direct the development of distinct T helper cells in vivo. J. Exp. Med. 189: 587-592. 176. McKnight, A. J., Zimmer, G. J., Fogelman, I., Wolf, S. F., & Abbas, A. K. (1994). Effects of IL-12 on helper T cell-dependent immune responses in vivo. J. Immunol. 152: 2172-2179. 177. Hofstra, C. L., Van Ark, I., Hofman, G., (Cool, M., Nijkamp, F. P., & Van Oosterhout, A. J. (1998). Prevention of Th2-like cell responses by coadministration of IL-12 and IL-18 is associated with inhibition of antigen-induced airway hyperresponsiveness, eosinophilia, and serum IgE levels. J. Immunol. 161: 5054-5060. 178. Moore, K. W., de Waal Malefyt, R., Coffman, R. L., & O'Garra, A. (2001). Interleukin-10 and the interleukin-10 receptor. Ann. Rev. Immunol. 19: 683-765. 179. Letterio, J. J., & Roberts, A. B. (1998). Regulation of immune responses by TGF-(3. Ann. Rev. Immunol. 16: 137-161. 180. Khalifeh, M. S. (1999). The role ofT cell mediated immune response in the development and resolution of colitis induced by Serpulina hyodysenteriae. MSc Thesis. Iowa State University. Iowa State University. 181. Morrissey, P. J., Charrier, K., Braddy, S., Liggitt, D., & Watson, J. D. (1993). CD4+ T cells that express high levels of CD45RJB induce wasting disease when transferred into

41
congenic severe combined immunodeficient mice. Disease development is prevented by cotransfer of purified CD4T T cells. J. Exp. Med. 178: 237-244. 182. Singh, B., Read, S., Asseman, C., Malmstrom, V., Mottet, C., Stephens, L. A., Stepankova, R., Tlaskalova, H., & Powrie, F. (2001). Control of intestinal inflammation by regulatory T cells. Immunol. Rev. 182: 190-200. 183. Takahashi, T., Kuniyasu, Y., Toda, M., Sakaguchi, N„ Itoh, M., Iwata, M., Shimizu, J„ & Sakaguchi, S. (1998). Immunologic self-tolerance maintained by CD25+CD4+naturally anergic and suppressive T cells: induction of autoimmune disease by breaking their anergic/suppressive state. Int. Immunol. 10: 1969-1980. 184. Vincent, M. S., Roessner, K., Lynch, D., Wilson, D., Cooper, S. M., Tschopp, J., Sigal, L. H., & Budd, R. C. (1996). Apoptosis of Fashlgh CD4+ synovial T cells by borrelia-reactive Fas-ligand hlgh yô T cells in Lyme arthritis. J. Exp. Med. 184: 2109-2117. 185. Carding, S. R., Allan, W., Kyes, S., Hayday, A., Bottomly, K., & Doherty, P. C. (1990). Late dominance of the inflammatory process in murine influenza by yô+ T cells. J. Exp. Med. 172: 1225-1231. 186. Doherty, P. C., Allan. W., Eichelberger, M., & Carding, S. R. (1992). Roles of a|3 and yÔ T cell subsets in viral immunity. Ann. Rev. Immuno.l 10: 123-151. 187. Skeen, M. J., & Ziegler, H. K. (1993). Induction of murine peritoneal yô T cells and their role in resistance to bacterial infection. J. Exp. Med. 178:971-984. 188. Skeen, M. J., & Ziegler, H. K. (1993). Intercellular interactions and cytokine responsiveness of peritoneal a|3 and yô T cells from L is ter ici- i n fee ted mice: synergistic effects of interleukin 1 and 7 on yô T cells. J. Exp. Med. 178:985-996. 189. van der Heyde, H. C., Manning, D. D., & Weidanz, W. P. (1993). Role of CD4+ T cells in the expansion of the CD4*, CD8" yô T cell subset in the spleens of mice during blood-stage malaria. J. Immunol. 151:6311 -6317. 190. Mombaerts, P., Amoldi, J., Russ, F., Tonegawa, S., & Kaufmann, S. H. (1993). Different roles of and yô T cells in immunity against an intracellular bacterial pathogen. Nature 365: 53-56. 191. Fu, Y. X., Roark, C. E., Kelly, K., Drevets, D., Campbell, P., O'Brien, R., & Bom, W. (1994). Immune protection and control of inflammatory tissue necrosis by yô T cells. J. Immunol. 153:3101-3115. 192. Egan, P. J., & Carding, S. R. (2000). Downmodulation of the inflammatory response to bacterial infection by yô T cells cytotoxic for activated macrophages. J. Exp. Med. 191: 2145-2158. 193. Huber, H., Descossy, P., van Brandwijk, R., & Knop, J. (1995). Activation of murine epidermal TCR-yô T cells by keratinocytes treated with contact sensitizers. J. Immunol. 155: 2888-2894. 194. Huber, H., Descossy, P., Regier, E., van Brandwijk, R., & Knop, J. (1995). Activation of phenotypically heterogeneous murine T cell receptor yô+ dendritic epidermal T cells by self-antigen(s). Int. Arch. Allergy Immunol. 107:498-507. 195. Boismenu, R., Chen, Y., & Havran, W. L. (1999). The role of intraepithelial yô T cells: a gut-feeling. Microbes Infect. 1: 235-240.

42
196. Boismenu, R., Feng, L., Xia, Y. Y., Chang, J. C„ & Havran, W. L. (1996). Chemokine expression by intraepithelial y8 T cells. Implications for the recruitment of inflammatory cells to damaged epithelia. J. Immunol. 157:985-992. 197. King, D. P., Hyde, D. M., Jackson, K. A., Novosad, D. M., Ellis, T. N., Putney, L., Stovall, M. Y., Van Winkle, L. S., Beaman, B. L„ & Ferrick, D. A. (1999). Cutting edge: protective response to pulmonary injury requires y5 T lymphocytes. J. Immunol. 162: 5033-5036. 198. Tam, S., King, D. P., & Beaman, B. L. (2001). Increase of y§ T lymphocytes in murine lungs occurs during recovery from pulmonary infection by Nocardia as te ro ides. Infect. Immun. 69: 6165-6171. 199. Boismenu, R., & Havran, W. L. (1994). Modulation of epithelial cell growth by intraepithelial yô T cells. Science 266: 1253-1255. 200. Boismenu, R., Hobbs, M. V., Boullier, S., & Havran, W. L. (1996). Molecular and cellular biology of dendritic epidermal T cells. Semin. Immunol. 8: 323-331. 201. Rubin, J. S., Bottaro, D. P., Chedid, M., Miki, T., Ron, D., Cheon, G., Taylor, W. G., Fortney, E., Sakata, H., Finch, P. W., & et al. (1995). Keratinocyte growth factor. Cell. Biol. Int. 19: 399-411. 202. Rubin, J. S., Bottaro, D. P., Chedid. M., Miki, T., Ron, D., Cunha, G. R., & Finch, P. W. (1995). Keratinocyte growth factor as a cytokine that mediates mesenchymal-epithelial interaction. Exs. 74: 191-214. 203. Boismenu, R., & Havran, W. L. (1998). yô T cells in host defense and epithelial cell biology. Clin. Immunol. Immunopathol. 86: 121-133. 204. Ka, H., Spencer, T. E., Johnson, G. A., & Bazer, F. W. (2000). Keratinocyte growth factor: expression by endometrial epithelia of the porcine uterus. Biol. Reprod. 62: 1772-1778. 205. Ka. H., Jaeger, L. A., Johnson, G. A., Spencer, T. E., & Bazer, F. W. (2001). Keratinocyte growth factor is up-regulated by estrogen in the porcine uterine endometrium and functions in trophectoderm cell proliferation and differentiation. Endocrinology 142: 2303-2310. 206. Jameson, J., Ugarte, K., Chen, N., Yachi, P., Fuchs, E., Boismenu, R., & Havran, W. L. (2002). A role for skin yô T cells in wound repair. Science 296: 747-749. 207. Chen, Y., Chou, K., Fuchs, E., Havran, W. L., & Boismenu, R. (2002). Protection of the intestinal mucosa by intraepithelial yô T cells. Proc. Natl. Acad. Sci. U S A 99: 14338-14343.

43
CHAPTER 2. DEFERENTIAL REQUIREMENTS FOR THE PROLIFERATION OF
PORCINE CD4+ AND yS T CELLS TO EXTRACELLULAR BACTERIAL
ANTIGENS
A manuscript to be submitted to Immunology
Raquel Hontecillas, Josep Bassaganya-Riera, and Michael J. Wannemuehler
Abstract
dp and yô T cells have different mechanisms of epitope recognition and are
stimulated by antigens of different chemical nature. Humans and mice have very low
numbers of circulating yô T cells, whereas in ruminants, chickens, and pigs this T cell subset
constitutes a significant fraction of the peripheral blood mononuclear cells (PBMC). Both
peripheral blood CD4+ and yô T cells from pigs immunized with a pepsin-digested bacterin of
the spirochete Brachyspira hyodysenteriae proliferate in vitro to stimulation with spirochetal
antigens. This antigenic preparation was used to assess the requirements for the proliferation
of each of the two subsets in mixed lymphocyte cultures. While CD4+ T cells only responded
to stimulation with B. hyodysenteriae antigens, but not to exogenous recombinant porcine
1L-2 whereas yô T cells proliferated when cultures were stimulated with either bacterial
antigens or exogenous recombinant IL-2. PBMC that had proliferated after 5 days in culture
expressed high levels of IL-2Ra and neutralization of IL-2 at the beginning of the culture
period was more efficient in blocking yô T cell than CD4+ T cell proliferation. After 5 days in
culture, CD4+ cells were the major source of IFN-y, while only a small number of yô T cells
had the capacity to produce this cytokine. Together, these results indicate that porcine CD4+
and yô T cells differ in the mechanisms that drive their proliferation, and in their effector
functions.

44
Introduction
Porcine lymphocytes, and more specifically T cells, are characterized by the
heterogeneous expression of the CD8aa coreceptor and by their diffuse distribution into the
different compartments of the immune system (I; 2). Although some murine or human T
cells also express CD8aa, numerically, the CD8aa+ subsets are not as abundant as in pigs.
In addition, whereas in humans and mice certain cell types are only present at specific
anatomic locations (e.g., murine yÔ CD8GCGC+ T cells within the intestinal epithelium), this
clear association between phenotype and tissue distribution has not been demonstrated in the
pig. Particularly, porcine peripheral blood mononuclear cells (PBMC) are characterized by
the presence of mature CD41" CD8aa" T cells (3) and by having very high numbers of yÔ T
cells (2).
yô T cells differ from afi T cells in several aspects. Through the TCR, a(5 T cells
recognize peptides presented in association with MHC-class I or II molecules on the surface
of target cells (i.e., CD8+ cytotoxic T cells) or APC (i.e., CD4+). yô T cells react to antigens
with chemical composition other than proteins (e.g., phosphoantigens) (4-6) and, although
costimulation is necessary for cell activation (7), antigen recognition does not require
processing and presentation by professional APC (8). In addition, the yô TCR is MHC-I and
II-unrestricted; instead, non-classical MHC molecules, such as T10 and T22 (8), or CD1 (9)
are ligands for the yô TCR.
Functional studies of porcine CD4+ CD8acT T cells have indicated that this cell type
represents memory/effector T helper cells. Moreover, in vitro experiments showed that

45
antigen-experienced CD4+ CD8aa~T cells from peripheral blood upregulate
CDSaa following a subsequent antigenic exposure (10; 11). Thus, it is currently accepted
that CD4^ CDSaa-T cells are the precursors of CD4+ CDSaa* T cells. With regard to yô T
cells, two major subsets have been identified based on the expression of the molecule SWC6
(12). Porcine SWC6+ cells constitute the majority of the circulating yô T cells and, although
their function is currently unknown, they are capable of inducing nonspecific cytotoxicity in
vitro (13; 14). The SWC6' yÔ T cells are phenotypically characterized as CD2+ CD6+ (15),
and a fraction of this subpopulation additionally expresses CDSaa (16). SWC6" yô T cells
are found in spleen, secondary lymphoid organs and they are also detectable as circulating
lymphocytes, although in lower numbers than SWC6+ yô T cells (15)
Porcine CD4+ and yÔ T cells are thought to contribute to cell-mediated immune
responses to extracellular bacterial antigens (17). Previous experiments demonstrated that
immunization with a pepsin-digested bacterin of Brachyspira hyodysenteriae, a gram-
negative spirochete, increases the pool of circulating CD8aa+ PBMC (18), which also
proliferate to B. hyodysenteriae specific antigen in vitro. Further analysis, indicated that
CD4+ and yÔ subsets constitute the majority of the T cells proliferating in vitro to antigenic
stimulation. In addition, CD4+ T cell depletion or blockade of antigen presentation through
MHC-II greatly diminished proliferative responses of the PBMC (17). Based on these results,
immunization with this bacterin induces a MHC-II-dependent CD41" T cell response.
However, because the vaccine affects both CD4 and yô" T cells, additional mechanisms of
activation might exist besides antigen presentation through MHC-II. We chose this
immunization regime as a model to investigate differential functional aspects of the two cell
types in the pig. In the present study, it was determined that the requirements for the

46
proliferation of porcine CD4+ and yô T cells are different: while CD4+ T cell responses were
strictly antigen-dependent, yô T cells proliferated, in addition to specific antigens, to
stimulation with exogenous IL-2. It is also shown that most of the IFN-y produced in culture
originates from CD4+ T cells, whereas, in spite of being one of the most numerous cell type
among PBMC, yô T cells have very reduced capacity of IFN-y production in culture.
Materials and methods
Animals and immunization
Four-week-old cross-bred pigs were immunized with either a squalene-adj uvanted
protease-digested preparation of Brachyspira hyodysenteriae, or adjuvant alone as previously
described (18-21). B. hyodysenteriae strain B204 was grown in tripticase soy broth (Becton
Dickinson; Cockesville, Maryland) supplemented with 5% horse serum (Hyclone, Logan,
Utah), 0.5% yeast extract (Difco, Detroit, Michigan) and VPI salts under anaerobic
conditions. Lyophilized bacterial cells were digested by incubation with pepsin (10"3 g, per g
of lyophilized B. hyodysenteriae) at pH 1.9-2.2 for 25 h at 37° C. The vaccine was prepared
by mixing at 1:1 (v:v) the pepsin-digested B. hyodysenteriae with the adjuvant (10% of
squalene/pluronic acid (80:20 v:v) in 2% Tween 80-Saline. Pigs received 3 intramuscular
doses of either vaccine or adjuvant alone at 15 day intervals. All animal experiments were
approved by the Iowa State University Committee on Animal Care.
Isolation of peripheral blood mononuclear cells
One week after the last immunization PBMC were obtained from whole blood by
gradient centrifugation. Briefly, a 1:4 dilution of blood in PBS was overlaid onto 4 mL of
1.077 lymphocyte separation media (Mediate, VA), and centrifuged at 800 x g for 40

47
minutes. PBMC from the plasma/ media interface were aspirated with a pasteur pipete,
washed 3 times in PBS, and resuspended in complete RPMI 1640 medium (10 % fetal calf
serum (Hyclone, UT), 25 mM HEPES buffer (Sigma., St. louis, MO), 100 units/ml penicillin
(Sigma), 0.1 m/ml streptomycin (Sigma), 5 x 10*5 2-mercaptoethanol (Sigma), ImM sodium
pyruvate (Sigma), 1 mM nonessential aminoacids (Sigma), and 2 mM essential amino acids
(Mediatech). Cells were enumerated with a Coulter Z1 Single Particle Counter (Beckman
Coulter, Miami, Florida).
Proliferation assay
A total of 20 x 106 PBMC isolated following the procedure described above were
labeled with the green fluorescent dye PKH67 (Sigma) following manufacturer instructions.
Briefly, PBMC were aliquoted into 15 mL polystyrene conical tubes and centrifuged at 400 x
g for 5 min. After eliminating the supernatant, cells were resuspended in 1 mL of Diluent C
(Sigma). The cell suspension was immediately transferred to a 15 mL polystyrene conical
tube containing 1 mL of a 2 mM solution of PKH67 dye in Diluent C (Sigma). Cells were
incubated with the dye for 5 min and the reaction was stopped by the addition of 2 mL of
Fetal Bovine Serum (Hyclone) and, after 1 min, 4 mL of RPMI 1640. To eliminate excess
fluorescent dye, cells were washed 3 times in RPMI 1640 and resuspended in complete
RPMI to a final 2xl06 cells/mL.
PBMC at 106 cells/ mL were seeded in 96 well flat-bottom microliter plates and
incubated with complete RPMI only, the pepsin digested preparation of B. hyodysenteriae
B204 antigen at 5 gg/mL, Concanavalin A at 5 ng/mL (Sigma) or 5 ng/mL of recombinant
porcine IL-2 (Biosource, MA), in a 200 p.L final volume. After 5 d in culture, PBMC were
harvested and labeled for the analysis cell surface marker expression by flow cytometry as

48
previously described ( 17). For IL-2 neutralization experiments, polyclonal anti-porcine IL-2
(R&D Systems, MN) was added at the beginning of the culture period at 3 gg/mL.
The optimal dose of recombinant IL-2 and the inhibitory concentration of polyclonal
anti-porcine IL-2 were determined in a titration experiment in which PBMC were incubated
with increasing amounts of recombinant IL-2, ranging from 0 to 20 ng/mL. Neutralizing IL-
2 antibodies were added at 0, 0.75, 1.5 or 3 mg/mL to triplicate cultures of the various
amounts of recombinant IL-2. Proliferation was assessed by titriated thymidine
incorporation. Results showed that addition of more than 5 ng/mL of recombinant IL-2 did
not result in stronger stimulation and that 3|ig/mL of polyclonal anti-IL-2 completely
inhibited exogenous IL-2-induced proliferation.
Antibodies used in flow cytometry
The following antibodies were used in the proliferation assay for the detection of cell
surface marker expression: mouse IgG2b anti-porcine CD4 (74-12-1), biotinylated mouse
IgG2a anti-porcine CD8a (76-2-11), mouse IgGlanti-porcine TCR5 chain (PGBL22A,
VMRD, Pullman, WA), mouse IgGl anti-porcine CD3 (8E6, VMRD), mouse IgG2a anti-
porcine CD8(3 (PG145A, VMRD), and mouse IgGl anti-porcine CD25 (PGBL25A, VMRD).
Intracellular IFN-y detection
PBMC were incubated in 96 well flat-bottom microliter plates at 106 cells/ mL with
either medium alone, or with the pepsin digested preparation of B. hyodysenteriae B204
antigen 5 |ig/mL, in a final volume of 200 |AL. On d 5, cultures were stimulated with 50
ng/mL of phorbol myristate acetate (PMA) (Sigma) and 100 ng/mL ionomycin (Sigma).
Golgi transport was blocked by the addition of 4 |xL of Golgi Stop (Pharmingen, San Diego,
CA) for every 6 mL of cell culture. After CD4/CD8a and TCRyô/CDSa cell surface staining,

49
cells were fixed and permeablized with the Cytofix-cytoperm kit (Pharmingen) following
manufacturer's instructions. PE-labeled mouse IgGl anti-porcine IFN-y (P2G10,
Pharmingen) was used for intracellular cytokine detection. Equally stained non-PMA and
ionomycin-stimulated cultures were used as negative controls, and mouse IgGl-PE (Sigma)
was used as isotype control.
Statistical analysis
Data were analyzed as a randomized complete block design. Analysis of variance (ANOVA)
was used to investigate the main effects of vaccination. ANOVA was performed using the
general linear model procedure (GLM) of SAS. Differences with P < 0.05 were considered
significant.
Results
Lymphocyte activation and proliferation in vitro
Previous studies demonstrated that immunization with a B. hyodysenteriae bacterin
induced antigen-specific T cell proliferation (21). In order to further investigate these
responses, PBMC from control and immunized animals were cultured with medium alone, or
with pepsin-digested whole cell B. hyodysenteriae antigens. As it had been reported before, T
cells from immunized animals proliferated in vitro in the presence of the pepsin digested B.
hyodysenteriae antigens as determined by the decrease in PKH67 mean fluorescence
intensity of CD3+ PBMC (figure 1 A). The same results were obtained by 3H-thymidine
incorporation (data not shown). In addition, a fraction of CD3" cells from immunized pigs
(5.6 ± 0.04 vs. 1.3 ± 0.34 % in non-immunized) also proliferated to antigenic stimulation,
although we did not phenotypically characterize the responding CD3~ lymphocytes.

50
Anti-CD25 staining of PBMC from immunized pigs cultured with B. hyodysenteriae
antigens (figure IB) indicates that antigenic stimulation upregulated cell-surface IL-2Ra .
Based on the expression of CD25 on cells from immunized pigs that had proliferated, IL-
2Ra upregulation correlated with cell division. However, by d 5, a small proportion of non-
stimulated PBMC from immunized pigs (6.09 ± 0.9), or PBMC from control pigs cultured
with either medium (7.5 ± 2.7) or B. hyodysenteriae antigens (8.9 ± 0.02) stained positive for
IL-2Ra, although these cells failed to proliferate.
Subset analysis of T cells responding to B. hyodysenteriae
In order to dissect the pool of T cells that had proliferated, CD4T(i.e., helper a(3 T
cells), CD8afT (i.e., cytotoxic a(3 T cells), and yô T cells were individually analyzed. An
additional T cell subset (i.e., TCR CD4~ CD8aaT) has been identified in porcine PBMC;
however, due to the lack of monoclonal antibodies specific for either the porcine TCRa or (3
chains, it was not possible to analyze this subset individually. Proliferation results show that
6.4 ± 2.3 % of CD4+ and 8.3 ± 3.97 % ofy5 T cells isolated from immunized pigs
proliferated in vitro to B. hyodysenteriae antigens. Thus, these two subsets account for the
vast majority of the T cells responding to antigenic stimulation (figure 2). No proliferative
responses were detected in CD8a|3+ T cells. Additionally, when non-specific T cell responses
were detected (i.e., PBMC from immunized pigs cultured with medium or PBMC from
controls cultured with B. hyodysenteriae antigens), it was due to proliferation of yô T cells.
Both CD4+ and yô T cells can be further subdivided based on the expression of
CD8a. In order to determine differential responses within CD4+ or y8 T cell subsets, cells
were stained with both anti-CD4 and anti-CD8a, or anti-TCRS chain and anti- CD8a for the
phenotypic analysis of cells that had proliferated after 5 d in culture. Results show that when

51
PBMC from immunized pigs were stimulated with B. hyodysenteriae antigens, 9.9 ± 3.9 % of
CD4* cells were CD4+ CDSaa" PKH67 10 and 12.4 ±6 % were CD4+ CDSaa*
PKH67 lo. With regard to the yô T cell subset, 17.13 ± 8.0 of the cells that proliferated were
yÔ CDSaa", wheras only a small fraction of TCRyS CDSaa* (1.4 ± 0.5 %) proliferated in the
presence of B. hyodysenteriae antigens (Table I). Also, non-specific background
proliferation (7.4 ± 2.4) was observed for TCRyô CDSaa" T cells isolated from non-
immunized animals in cultures where bacterial antigens were added.
Differential requirements for proliferation of CD4* and y& T cells
Lymphocytes that had proliferated during the 5 d in culture upregulated and
maintained cell surface CD25 expression (figure I). In order to determine the IL-2
requirements for activation and proliferation of the different cell types responding to
antigenic stimulation, PBMC were cultured with either B. hyodysenteriae pepsin-digested
antigen or medium supplemented with recombinant porcine IL-2. This cytokine alone was an
insufficient stimulus for a(3 T cells (i.e., CD4+ and CD8a0*) to proliferate (figure 3A).
However, in some instances, CD4+ T cells weakly responded (data not shown). Cell
phenotype based on CDSaa expression indicated that only CD4*CD8aa~, but not
CD4*CD8aa* T cells could undergo cell division following stimulation with exogenous IL-
2. In contrast to a(3 T cells, a significant proportion of yô T cells (13.21 ±5.7) responded to
the addition of this cytokine (figure 3 A). The majority of the yô T cells that proliferated were
CDSaa" as well. No significant differences were found between PBMC isolated from control
or B. hyodysenteriae-immunized pigs in their proliferation to the addition of exogenous IL-2.
Treatment of cultures with neutralizing anti-IL-2 antibodies at the beginning of the
culture period, blocked proliferation of yô CDSaa" T cells to exogenous IL-2, and resulted in

52
50 % reduction of the proliferative response of this cell type when PBMC from immunized
pigs where stimulated with antigen (figure 3 B). With regards to the CD4+ pool, proliferation
of CD4+CD8aa+ T cells to B. hyodysenteriae diminished from 21.01 ± 4.58 to 11.9 ± 3.6 %
by the addition of lL-2-neutralizing antibodies (figure 3B).
CD4* T cells are the major source of IFN-y
Previously, we reported the presence of IFN-y-producing cells in B. hyodysenteriae-
stimulated PBMC from vaccinated pigs. Additionally, we showed that the IFN-y response
was CD4+T cell-dependent because in vitro CD4T cell depletion decreased the numbers of
IFN-y-producing cells (18). In the present study, intracellular cytokine staining results
indicate that CD4+T cells are the major source of IFN-y in PBMC (figure 4). In addition,
when CD4* T cells were stimulated with B. hyodsyenteriae, the percent IFN-y+-producing
cells decreased (2.53±1.09 to l.07±0.1) along with the mean fluorescence intensity of the
cytokine staining (1758.7±114 to 436±18.). This effect was observed in all pigs, irrespective
of the in vivo treatment, although it was more marked in CD4+ cells from immunized pigs.
On the other hand, very few yô T were able to produce IFN-y in vitro. However, the percent
of cytokine-positive yÔ T cells increased in both vaccinated (from 0.687± 0.158 to l.52±0.82)
and controls (from 0.69±0.19 to 1.157 ±0.827) after stimulation with B. hyodysenteriae
antigens.
Discussion
Our results show that both a(3 and yô T cells respond in vitro following antigenic
stimulation, however, the requirements for activation are different for the two T cell subsets.
The fact that cells that had divided after 5 d in culture (i.e., lower intensity of PKH67

53
staining) expressed high levels of CD25, and that IL-2 neutralization blocked proliferation,
suggest that IL-2 is necessary for porcine T cell proliferation. However, based on the
responses to exogenous IL-2, the addition of this cytokine alone is insufficient to induce
CD4+ and CD8aP T cell division. The antigen used in vitro in these experiments was the
pepsin-digested preparation of B. hyodysenteriae. As it would be expected for an
extracellular antigen. CD4+ (i.e, T helper) but not CD8aP T cells (i.e., cytotoxic T cells)
responded to antigenic stimulation. Thus, both IL-2Ra and TCR-derived signals were
necessary for «P T cell activation as measured by proliferation assays.
In contrast to aP T cells, yô T cells proliferated in response to specific antigen as well
as to addition of exogenous IL-2. Earlier studies have shown that blockade of antigen
presentation through MHC-II. or culture of PBMC depleted from CD4+ T cells diminished
yô T cell proliferation to B. hyodysentariae antigens (17). It could be interpreted that yô T
cell proliferation is secondary to CD41"T cell activation. In this regard, it could be proposed
that B. hyodysenteriae-antig&n specific cells are within the CD4™ pool, and yô T cell
proliferation is bystander to the activation of antigen-specific CD4+ T cells. A similar
interaction between CD4+and yô T cells was described for in vitro pseudorabies virus-
stimulated PBMC from immunized pigs (22). It remains unknown whether, in addition to IL-
2, TCR signals are needed for proliferation of porcine yô T cells.
Yang et al. reported that, in contrast to (%P T cells, porcine yô T cells failed to
proliferate in vitro following stimulation with a panel of anti-CD3 monoclonal antibodies.
These results suggest that signal transduction events that lead to TCR activation are distinct
for porcine aP and yô T cells (23). Comparative studies of the structure of the human aP and
yô TCR have shown differences that constitute the basis for some of the biological properties

54
of each the two cell populations. Among these, the fact that the hinge region of the yô TCR
has additional amino-acid residues, and that the domain that has been proposed to interact
with CD3E is shorter in the <*P TCR, suggest that aP and yô TCR form different signaling
complexes (24). In fact, the molecular composition of the TCR-CD3 complex in human and
murine yô T cells differs from aP T cells in that the first lacks CD3S subunits. However, at
the functional level, the threshold for TCR activation appears to be lower in yS than in (%P T
cells based on calcium mobilization, extracellular signal-regulated kinase (ERK.) activation,
and proliferation following stimulation with anti-CD3 (25).
Our results do not provide sufficient evidence to support the concept that all yô T
cells that proliferated in B. hyodysenteriae-stimulated cultures responded solely to IL-2-
induced stimulation. A portion of yô T cells could have responded independently from CD4T
T cells and, equally to ctp T cells, also require TCR activation for proliferation. This could be
explained by non-MHC-II-restricted specific recognition of B. hyodysenteriae epitopes by yô
T cells. This hypothesis is supported by the fact that yô T cell proliferation at background
levels was observed when PBMC from non-immunized pigs were cultured with bacterial
antigens. Also, because yô T cells proliferated in the absence of B. hyodysenteriae antigens
but in the presence of exogenous IL-2, an additional CD4* and B. hyodysenteriae antigen-
independent mechanism must exist for the activation of yô T cells. A CD41"-independent
pathway of yô T cell activation in vitro by an unknown ligand present on monocytes and
granulocytes has been described in cattle (26). Porcine and bovine yô T cells have similar
phenotypic characteristics. It is also probable that they also have functional aspects in
common. A similar ligand might also exist in porcine cells, however, while stimulation with

55
this ligand induced IFN-y production by bovine cells, in our experiments yô T cells failed to
produce this cytokine.
Several in vivo models have shown an increase in yô T cells following activation of
a(3 T cells (27-29). Such changes in the yô T cell population initially depend on the
production of IL-2 (30; 31). It has been suggested that a negative feed back loop is
established between yô and a(3 T cells by which, antigen-specific a(3 T cell responses are
inhibited by the expanded yô T cells (28). Activated yô T cells express high levels of Fas
ligand and can induce apoptosis of Fas* CD4+ T cells (32). Porcine yô T cells have non
specific cytotoxic activity in vitro, although the molecular basis has not been investigated.
Our results show that B. hyodysenteriae antigen stimulates CD4* CDSaa™ and CD4*
CDSaa* from immunized pigs to proliferate. In addition, antigenic stimulation decreased the
number of CD4+ cells producing IFN-y. It is possible that the activated yô T cells
proliferating in B. hyodysenteriae-stimulated cultures downregulated CD4+
CDSaa" and CD4+ CDSaa* responses (i.e., less IFN-y production).
The magnitude of in vitro T cell proliferation in antigen recall responses is a
functional parameter frequently used to evaluate the efficacy of a cellular immune response.
T cell proliferative responses to an antigen indicate the presence of antigen-specific cells, and
therefore the generation of immunological memory. We have shown in a mixed lymphocyte
culture of porcine PBMC that vaccination with a gram-negative spirochete bacterin generated
antigen-dependent CD4 T cell responses. Additionally, yô CDSaa" T cells proliferated in
vitro. We analyzed yô CDSaa* T cells separately because we had reported that CDSaa*
PBMC from B. hyodysenteriae-immunized pigs were increased in vivo, and proliferated in
antigen recall responses. In these experiments, yô CDSaa* T cells failed to respond to either

56
antigen or IL-2 stimulation. Future studies should address the subset composition (i.e.,
SWC6+ or SWC6") of the responding yô T cells.
Immunization with this bacterin protects pigs against B. hyodysenteriae-indxiced
colitis. We are currently investigating the function of CD4T and yô T cells at the colonic
mucosa during infection. Very few studies have addressed the role of porcine yÔ T cells in
vivo. SWC6+ yS T cells (i.e., CD2" CDSaa") were recruited to skin inflammatory lesions
induced phytohemagglutining injection (33; 34). In several murine models, yô T cells have
been reported to downregulate inflammation (28). If these responses were beneficial, it
would be of interest to elucidate the basis of the CD4T-yô T cell interactions and whether this
mechanism should be exploited for the treatment or prevention of inflammatory diseases.
Table 1. Cell surface phenotype of proliferating peripheral blood T cell subsets from control or immunized pigs '
Medium2 B. hyodysenteriae antigen
Phenotype3 Control Immunized Control Immunized
CD4* CDSaa" PKH671"4
CD4* CDSaa1- PKH67'0
yô CDSaa" PKH67loS
yô CDSaa' PKH6710
2.228 ± 0.998 1.263 ±0.709 3.804 ±2.061 0.438 ±0.279
2.244 ±0.826 0.961 ±0.261 7.226 ± 4.226 0.326 ±0.161
3.345 ± 1.422 1.831 ±0.891 7.366 ±2.414 0.689 ±0.401
9.894 ±3.971*6
12.394 ±6* 17.131 ±8.063* 1.383 ±0.487
1 Results are expressed as mean (n=5) percentage of CD4*or TCR5* cells that had proliferated (i.e., PKH6710) after 5 d in culture. " Peripheral blood mononuclear cells isolated from control and B. hyodysenteriae-immumzsd pigs were labeled with PK.H67 and cultured with either complete RPMI medium or 5 gg/mL of B. hyodysenteriae pepsin-digested antigenic preparation. 3 Cells were harvested on day 5 and double stained with monoclonal antibodies specific for CD4 and CD8a or TCRS and CD8a, and analyzed by 3 color flow cytometry. 10,000 events within the viable cell gate were acquired. 4 CD4* cells within the mononuclear viable gate were analyzed by plotting CD8a versus RK.H67. 5 TCR5* cells within the monocyte viable gate were analyzed by plotting CD8a versus PK.H67. 6 *, f-value < 0.05 when compared to the non-immunized control group.

57
Medium Medium
B
mn 0.7
1 i :-•» ft.
PKH67
Immunized Control
Figure 1. Proliferation (A) and CD25 expression (B) of PBMC from an immunized (left panel) and a non-immunized control pig (right panel) after 5 d in culture with either RPMI medium or B. hyodysenteriae antigens. PBMC were isolated one week after three intramuscular immunizations with a pepsin-inactivated B. hyodysenteriae bacterin. CD3 staining (top row) indicates that T cells from immunized pigs specifically responded to stimulation with spirochetal antigens. The forward versus side scatter plot shows the cells included in the analysis (i.e., live mononuclear cells). Plots are representative from 2 experiments with a total of 5 pigs per group.

58
Medium
O
ÎÏS16
PKH67
Medium B. hyo
I 0.2 26.6
a: 0.3 25.5
|*r
1.63 ' 0.6 - • £*:
•* rfl
0.3
Immunized Control
Figure 2. Proliferation of CD41", CD8afT and yô T cells from an immunized (left panel) and a non-immunizad control pig (right panel) after 5d in culture with either RPMI medium or B. hyodysenteriae antigens. PBMC were isolated one week after three immunizations with a pepsin-inactivated B. hyodysenteriae bacterin. CD4+ and y5 T cells from vaccinated pigs account for the majority of the CD3+ proliferating cells. The forward versus side scatter plot shows the cells included in the analysis (i.e., live mononuclear cells). Plots are representative from 2 experiments with a total of 5 pigs per group.

59
PKH67
B. hyo rpIL-2
Anti-lL-2: • + •
TCRS
PKH67
Figure 3. IL-2 requirement for the proliferation of porcine T cells.
A) y§ T cells proliferated to the addition of 5 ng/mL of exogenous porcine recombinant IL-2 while neither CD4 nor CD8«P+ T cells responded to stimulation with this cytokine alone. Plots correspond to 5 d cultured PBMC from a representative non-immunized control pig. Two experiments were conducted with a total of 4 pigs per group. Cells included in the analysis correspond to live mononuclear cells gated as indicated in the forward versus side scatter plot.
B) IL-2 neutralization blocked proliferation of y8 CD8~ T cells stimulated with exogenous porcine IL-2 (lower row in right panel). In PBMC cultures from immunized pigs stimulated with B. hyodysenteriae antigen, the addition of anti-porcine IL-2 antibodies at the beginning of the culture period diminished proliferation of both CD4+CD8aot+ (top row in left panel) and yô CDS" T cells (bottom row in left panel), although proliferative responses of the latter were more affected by IL-2 neutralization. CD4+ or TCR§+ T cells within the viable gate were analyzed by plotting CD8a versus PKH67 as indicated in the CD4 versus CDSa or TCR5 versus CDSa plots, respectively. Plots are representative of one experiment with 3 pigs per group.

60
I Medium B. hyo FSC ] 2:3
i 1 :
Z ! be. ;
0.3 2 1-1
4 0:5 < i
4
Medium B. hyo
1 0.6
•32.2 Iliiêf'
3.4 1.7
Immunized
O.R. 0.5 -
0.6 . . 0.5 Control
w : u CD4 TCR-Ô
Figure 4. IFN-y production by CD4+ and yS T cells from an immunized (top row) and a non-immunized control pig (bottom row). PBMC were isolated from controls and B. hyodysenteriae pepsin-digested-immunized pigs one week after the third immunization. After 5d in culture with either medium alone or B. hyodysenteriae antigens, cells were stimulated with PMA and ionomycin for 4 hours for the detection of intracellular IFN-y by flow cytometry. CD41" T cells are the major source of IFN-y, and stimulation with B. hyodysenteriae antigens modulates IFN-y production by CD41" and yô T cells. Initial gate was set on the mononuclear cell population as indicated in the forward versus side scatter plot. Plots are representative from one experiment of 3 pigs per treatment.

61
References
1. Yang, H., & Parkhouse, R. M. (1996). Phenotypic classification of porcine lymphocyte subpopulations in blood and lymphoid tissues. Immunology 89: 76-83. 2. Binns, R. M. (1994). The Null/yô TCR+ T cell family in the pig. Vet. Immunol. Immunopathol. 43: 69-77. 3. Pescovitz, M. D., Sakopoulos, A. G., Gaddy, J. A., Husmann, R. J., & Zuckermann, F. A. (1994). Porcine peripheral blood CD4+/CD8+ dual expressing T-cells. Vet. Immunol. Immunopathol. 43: 53-62. 4. Schoel, B., Sprenger. S., & Kaufmann, S. H. (1994). Phosphate is essential for stimulation of Vy9 V52 T lymphocytes by mycobacterial low molecular weight ligand. Eur. J. Immunol. 24: 1886-92. 5. Constant, P., Davodeau, F., Peyrat, M. A., Poquet, Y., Puzo, G., Bonneville, M., & Fournie, J. J. (1994). Stimulation of human yô T cells by nonpeptidic mycobacterial ligands. Science 264: 267-70 6. Tanaka, Y., Mori ta, C. T., Nieves, E., Brenner, M. B., & Bloom, B. R. (1995). Natural and synthetic non-peptide antigens recognized by human yô T cells. Nature 375: 155-8. 7. Sperling, A. I., Linsley, P. S., Barrett, T. A., & Bluestone, J. A. (1993). CD28-mediated costimulation is necessary for the activation of T cell receptor-yô+ T lymphocytes. J. Immunol. 151: 6043-50
8. Schild, H., Mavaddat, N., Litzenberger, C., Ehrich, E. W., Davis, M. M., Bluestone, J. A., Matis, L., Draper, R. K., & Chien, Y. H. (1994). The nature of major histocompatibility complex recognition by yô T cells. Cell 76: 29-37. 9. Spada, F. M., Grant, E. P., Peters, P. J., Sugita, M., Melian, A., Leslie, D. S., Lee, H. K., van Donselaar, E., Hanson, D. A., Krensky, A. M., Majdic, O., Porcelli, S. A., Mori ta, C. T., & Brenner, M. B. (2000). Self-recognition of CD1 by y/ô T cells: implications for innate immunity. J. Exp. Med. 191:937-948. 10. Zuckermann, F. A., & Husmann, R. J. (1996). Functional and phenotypic analysis of porcine peripheral blood CD4/CD8 double-positive T cells. Immunology 87: 500-512. 11. Zuckermann, F. A. (1999). Extrathymic CD4/CD8 double positive T cells. Vet. Immunol. Immunopathol. 72: 55-66. 12. Carr, M. M., Howard, C. J., Sopp, P., Manser, J. M., & Parsons, K. R. (1994). Expression on porcine yô lymphocytes of a phylogenetically conserved surface antigen previously restricted in expression to ruminant yÔ T lymphocytes. Immunology 81: 36-40. 13. Pauly, T., Weiland, E., Hirt, W., Dreyer-Bux, C., Maurer, S., Summerfield, A., & Saalmuller, A. (1996). Differentiation between MHC-restricted and non-MHC-restricted porcine cytolytic T lymphocytes. Immunology 88:238-46. 14. de Bruin, T. G., van Rooij, E. M., de Visser, Y. E., Voermans, J. J., Samsom, J. N., Kimman, T. G., & Bianchi, A. T. (2000). Discrimination of different subsets of cytolytic cells in pseudorabies virus-immune and naive pigs. J. Gen. Virol. 81: 1529-1537.

62
15. Yang, H., & Parkhouse, R. M. (2000). Characterization of the porcine yô T-cell receptor structure and cellular distribution by monoclonal antibody PPT27. Immunology 99: 504-9 16. Yang, H., & Parkhouse, R. M. (1997). Differential expression of CDS epitopes amongst porcine CDS-positive functional lymphocyte subsets. Immunology 92: 45-52. 17. Waters, W. R., Hontecillas, R., Sacco, R. E., Zuckermann, F. A., Harkins, K.. R., Bassaganya-Riera, J., & Wannemuehler, M. J. (2000). Antigen-specific proliferation of porcine CDSaa cells to an extracellular bacterial pathogen. Immunology 101: 333-341. 18. Waters, W. R., Sacco, R. E., Dorn, A. D., Hontecillas, R., Zuckermann, F. A., & Wannemuehler, M. J. (1999). Systemic and mucosal immune responses of pigs to parenteral immunization with a pepsin-digested Serpulina hyodysenteriae bacterin. Vet. Immunol. Immunopathol. 69: 75-87. 19. Bassaganya-Riera, J., Hontecillas, R., Zimmerman, D. R., & Wannemuehler, M. J. (2002). Long-term influence of lipid nutrition on the induction of CDS* responses to viral or bacterial antigens. Vaccine 20: 1435-1444. 20. Bassaganya-Riera, J., Hontecillas, R., Zimmerman, D. R., & Wannemuehler, M. J. (2001) Dietary conjugated linoleic acid modulates phenotype and effector functions of porcine CDS* lymphocytes. J. Nutr. 131:2370-2377. 21. Waters, W. R., Pesch, B. A., Hontecillas, R., Sacco, R. E., Zuckermann, F. A., & Wannemuehler, M. J. (1999). Cellular immune responses of pigs induced by vaccination with either a whole cell sonicate or pepsin-digested Brachyspira (Serpulina) hyodysenteriae bacterin. Vaccine 18: 711-719. 22. Summerfield, A.. & Saalmuller, A. (1998). Interleukin-2 dependent selective activation of porcine yô T lymphocytes by an extract from the leaves of Acanthospermum hispidum. Int. J. Immunopharmacol. 20: 85-98. 23. Yang, H., & Parkhouse, R. M. (1998). Differential activation requirements associated with stimulation of T cells via different epitopes of CD3. Immunology 93:26-32. 24. Allison, T. J., Winter, C. C„ Fournie, J. J., Bonneville, M., & Garboczi, D. N. (2001). Structure of a human yô T-cell antigen receptor. Nature 411: 820-824. 25. Hayes, S. M., & Love, P. E. (2002). Distinct structure and signaling potential of the yô TCR complex. Immunity 16: 827-838. 26. Sathiyaseelan, T., Naiman, B., Welte, S., Machugh, N., Black, S. J., & Baldwin, C. L. (2002). Immunological characterization of a yô T-cell stimulatory ligand on autologous monocytes. Immunology 105: 181-189. 27. Carding, S. R., Allan, W., Kyes, S., Hayday, A., Bottomly, K., & Doherty, P. C. (1990). Late dominance of the inflammatory process in murine influenza by y/ô * T cells. J. Exp. Med. 172: 1225-1231. 28. Born, W., Cady, C., Jones-Carson, J., Mukasa, A., Lahn, M., & O'Brien, R. (1999). Immunoregulatory functions ofyô T cells. Adv. Immunol. 71:77-144. 29. Doherty, P. C., Allan, W., Eichelberger, M., & Carding, S. R. (1992). Roles of a(3 and yô T cell subsets in viral immunity. Annu. Rev. Immunol. 10:123-151. 30. Haque, A., Echchannaoui, H., Seguin, R., Schwartzman, J., Kasper, L. H., & Haque, S. (2001). Cerebral malaria in mice: interleukin-2 treatment induces accumulation of yô T cells

63
in the brain and alters resistant mice to susceptible-like phenotype. Am. J. Pathol. 158: 163-172. 31. Choi, K. D., & Lillehoj, H. S. (2000). Role of chicken IL-2 on yS T-cells and Eimeria acervulina-induced changes in intestinal IL-2 mRNA expression and yô T-cells. Vet. Immunol. Immunopathol. 73: 309-321. 32. Vincent, M. S., Roessner, K., Lynch, D., Wilson, D., Cooper, S. M., Tschopp, J., Sigal, L. H., & Budd, R. C. (1996). Apoptosis of Fashlgh CD4T synovial T cells by borrelia-reactive Fas-ligandhlgh yô T cells in Lyme arthritis. J. Exp. Med. 184: 2109-2117. 33. Whyte, A., Haskard, D. O., & Binns, R. M. (1994). Infiltrating yô T-cells and selectin endothelial ligands in the cutaneous phytohaemagglutinin-induced inflammatory reaction.PG. Vet. Immunol. Immunopathol. 41: 31-40. 34. Whyte, A., Licence, S. T., Robinson, M. K.., & van der Lienden, K. (1996). Lymphocyte subsets and adhesion molecules in cutaneous inflammation induced by inflammatory agonists: correlation between E-selectin and yÔ TcR* lymphocytes. Lab. Invest. 75: 439-449.

64
CHAPTER 3. MODIFICATION OF THE LYMPHOCYTE COMPOSITION AND
CYTOKINE ENVIRONMENT DURING Brachyspira hyodysenteriae-INDUCED
COLITIS BY SYSTEMIC IMMUNIZATION WITH A PROTEINASE DIGESTED
BACTERIN
A manuscript to be submitted to Immunology
Raquel Hontecillas, Josep Bassaganya-Riera, Jennifer Wilson. David L. Hutto, and Michael
J. Wannemuehler.
Abstract
Systemic immunization has proven to be of limited efficacy in preventing bacterial
mucosal infections. Here, we have examined the mechanism by which an enteric disease can
be ameliorated by parenteral immunization with a protease-digested bacterin of the
spirochete Brachyspira hyodysenteriae. We have previously shown that systemic
immunization with B. hyodysenteriae induced proliferation of CDSaa*, CD4* and yÔ* T cells
to specific B. hyodysenteriae antigens. Here, we have examined how the colonic profiles of
immunoregulatory and pro-inflammatory cytokines as well as the lymphocyte subset
distribution were influenced by immunization and/or challenge with B. hyodysenteriae.
Following immunization with the protease-digested B. hyodysenteriae bacterin, colonic
inflammation was triggered by challenging the pigs with B. hyodysentariae. Colonic mucosal
lesions and lymphocyte subset distribution were evaluated by histology and
immunohistochemistry. Immunoregulatory and pro-inflammatory cytokines were assayed in
colonic tissue. Immunization up-regulated TNF-a and IL-1 (5 mRNA expression. In addition,
TGF-(3l mRNA was increased in infected pigs previously immunized. Finally, vaccination
diminished the loss of yô T cells observed following the bacterial challenge and favored a

65
decrease in numbers of lamina proprial CD4+ cells. Therefore, we report a unique case of
parenteral immunization that regulates colonic lymphocyte subset distribution and cytokine
expression. This model of immunoregulation of colitis may have important applications in
the treatment of inflammatory bowel disease in humans.
Introduction
Enteric inflammatory processes associated with inflammatory bowel disease (IBD)
have been studied using a number of murine models of coltis (IBD) (I). Antigens expressed
by the intestinal normal flora contribute to the pathogenesis of IBD by triggering abnormal
immune and/or inflammatory responses (1; 2). Excessive activity of T helper (Th) cells (i.e.,
CD4*), especially polarized Thl, induces a cytokine imbalance often observed in Crohn's
disease patients (3-6). On the other hand, Th cells are controlled by regulatory T cells (i.e.,
Th3. Trl or CD4+ CD25+), mainly through the production of suppressor cytokines such as
transforming growth factor-^ (TGF-P) or IL-10 (7-10). Thus, defects in the regulation of T
cell responses contribute to the pathogenesis of inflammation by negatively affecting
mucosal homeostasis (1).
Brachyspira hyodysenteriae is a bacterial enteric pathogen that causes swine
dysentery, a severe hemorrhagic colitis (11). The bacteria invade the colonic mucosa without
spreading systemically (12). Similarly to IBD, the normal intestinal flora is involved in the
disease pathogenesis (13). One of the virulence factors of B. hyodysenteriae is a low
molecular mass P-hemolysin that was shown to induce apoptosis of intestinal epithelial cells
(14). Following loss of the epithelial cell barrier, excessive exposure of the underlying
mucosa to luminal antigens triggers a severe inflammatory response.

66
Parenetral immunization of pigs with a pepsin-inactivated bacterin of B.
hyodysenteriae ameliorates clinical and histopathological signs associated with swine
dysentery (15). CD8act\ CD4+ and yô T cells isolated from peripheral blood of immunized
pigs respond in vitro to stimulation with B. hyodysenteriae antigens (16-19); however, the
effect that vaccination has on these cell subsets in the colonic mucosa during infection has
not been investigated. In the present study it was shown that both vaccination and infection
modulated lymphocyte composition of the colonic mucosa. Particularly, it is shown that the
protection against disease induced by immunization correlated with a decrease in lamina
proprial CD4+ cells. Furthermore, although infected pigs had lower yÔ intraepithelial
lymphocytes (IEL), those that were previously immunized maintained higher numbers of this
lymphocyte subset. These phenotypic changes were accompanied by an increase in TGF-
P mRNA in colonic tissue recovered from immunized pigs. Immunization also increased the
expression of IL-1(3 and TNF-a. Thus, on the basis of these results, systemic immunization
modified the outcome of colonic inflammatory and immune responses.
Materials and methods
Experimental design
Immunization: a total of 12 four-week old pigs were immunized with either a squalene-
adjuvanted, pepsin-digested preparation of Brachyspira hyodysenteriae (n=6) or adjuvant
alone (n=6). The vaccine was prepared as previously described (20). Pigs received 3
intramuscular doses at 15 days intervals.
Challenge: two weeks after the third immunization, half of the control or immunized pigs
were orally challenged with two doses of 1010 CFU of B. hyodysenteriae strain B204
given on consecutive days. All cultures used for infection studies were >90% motile and

67
had been passed in vitro 23 to 25 times. After challenge, pigs were monitored daily for
clinical signs of disease (e.g., diarrhea, depression, anorexia, abdominal pain, and
dehydration). Stools were observed and scores were based on consistency: (0), no diarrhea,
stools formed: (1) mild, (2) moderate, and (3) severe diarrhea. Stools were also observed for
the presence of blood and/or mucus. Clinical response scores were calculated daily following
challenge by adding the diarrhea score (i.e., 0 to 3) plus a 1 for the presence of blood and/or a
1 for the presence of mucus. All animal related procedures were approved by the Iowa State
University Animal Care and Use Committee.
Necropsy: 15 days post-challenge pigs were anesthetized by administration of Rompun
(Bayer; Shawnee, KS)/Telazol (Fort Dodge Laboratories; Fort Dodge, IA) intramuscularly
and euthanized via electrocution. At necropsy, swabs of cecal and spiral colon contents were
streaked onto modified BJ blood agar plates containing antibiotics for isolation of B.
hyodysenteriae (21). Sections of spiral colon and cecum were obtained, fixed in 10%
buffered formalin, later embedded in paraffin, and then sectioned for histological
examination. Samples of colon and mesenteric lymph nodes were embedded in RNA Later
(Ambion Inc., Austin, TX) for isolation of total RNA and analysis of cytokine m RNA
expression. For immunohistochemistry, samples of colonic tissue were placed in tissue
freezing medium (Triangle Biomedical Sciences, Durham, NC) and snap frozen in
ethanol/dry ice bath.
Histopathological evaluation of colonic samples.
Hematoxilin-eosin (H & E)-stained sections from colon and cecum were labeled with
accession numbers lacking any reference to immunization or infection status (i.e., blinded to
evaluator). Histological evaluation was conducted on the basis of gland depth (i.e., expressed

68
as crypt depth as a function of crypt width), metaplasia/hyperplasia (i.e., increased cell
turnover and basal cell basofilia, or goblet cell metaplasia), lamina propria vascular change
(i.e., capillary dilation and lamina propria hemorrhage), inflammation and submucosal
change (i.e.. inflammatory cells infiltrates at the lamina propria or submucosa), and epithelial
erosions (i.e., 1 ) no erosion, 2) mild erosion and 3) severe erosion).
Determination of the lymphocyte subset composition of the mucosa by
immunohistology.
Frozen colonic tissue embeded in OCT tissue freezing medium were cut at 5 to 7 |im-
thick sections on a cryostat at - 18 °C. Sections were placed on poly-L-lysine coated slides,
fixed in 95% methanol for 2 minutes and soaked in cryopreservative solution (0.5 M sucrose,
0.006 M MgCh, 50 % glycerol) for 10 minutes. Slides were stored at - 20 °C until processed
for immunohistchemistry. Prior to staining tissues with monoclonal antibodies, slides were
warmed to room temperature and rehydrated in 0.5 M Tris solution. Endogenous peroxidase
activity was blocked by adding 0.3 % hydrogen peroxide for 10 minutes. Non-specific
binding was blocked by adding immunohistochemistry buffer containing 5 % normal goat
Serum/ 3 % bovine serum albumin/tris buffer (NGS/BSA/Tris) solution at room temperature
for 2 hours. Slides were incubated with the primary antibody solution overnight at 4 °C.
Primary antibodies were diluted in NGS/BSA/Tris. Sections were stained with anti-porcine
CD4 (74-12-1 hybridoma supernatant), anti-porcine-CD8a (76-2-11 hybridoma supernatant),
anti- porcine TCRÔ (PGBL22A, VMRD, Pullman, WA), or anti-porcine CD3 (8E6, VMRD).
Prior to the incubation with the secondary antibodies, slides were rinsed with Tris solution.
Peroxidase-conjugated goat anti-mouse IgG (H+L) (Jackson ImmunoResearch, West Grove,
PA) diluted at 1:300 in NGS/BSA/Tris was used for the detection of anti-porcine CD4, anti-

69
porcine CD3, and anti-porcine TCR8. Biotinylated goat anti-mouse IgG2a F(ab')i fragments
(Southern Biotechnologies Associates Inc. Birmingham, AL) were used for the detection of
anti-porcine CD8a primary antibody diluted in NGS/BSA/Tris at 1:250. All secondary
antibodies were incubated for 2 hours at RT. Slides were then treated with peroxidase-
conjugated streptavidin, diluted in Tris solution (1:500) and incubated for 1 hour (RT).
Positive antibody binding was visualized with diaminobenzediene (Biomedia Corporation,
Foster City. CA). Tissue sections were counterstained with Instant Hematoxylin (Shandon,
Pittsburg, PA) and coversliped with Immu-mount (Shandon). The numbers of CD4+, CD8a+,
CD3T, and TCRyfT cells in either epithelium or lamina propria were enumerated for each pig
and antibody staining. Data are presented as the sum of positive-staining cells in 5 randomly
chosen fields of 0.375 mm2 area, observed at 400 x magnification.
Isolation of total RNA.
Total RNA from full-thickness colonic tissue recovered with a 3 mm diameter punch
biopsy 15 days after challenge was isolated using the total RNA isolation MiniKit (Qiagen,
Valencia. CA) following manufacturer's instructions. RNA was diluted in 0.02% diethyl
pyrocarbonate (DEPC)-treated water (Qiagen). RNA concentration and purity were
determined using a spectrophotometer at an optical density (OD)2eo and OD26o/OD2so ratios,
respectively. All samples had OD260/OD280 ratios above 1.80 corresponding to 90-100% pure
nucleic acid. Samples were stored at —70°C until processed.
Ribonuclease protection assay
A custom template set (Pharmingen, San Diego, CA) was used for the synthesis of
antisense-RNA probes for the detection of porcine IL-4, IL-12p40, TNFa, IL-lfî, TGF-
Pt, IL-6, IFN-y, L32 and GAPDH mRNAs. [32P]UTP (Amersham Pharmacia)-labeled probes

70
were synthesized with an in vitro transcription kit (Pharmingen) following manufacturer's
instructions. Target RNA was hybridized overnight to the antisense RNA probes. Single-
stranded unbound RNA and free probe were subjected to RNases A and Tl digestion.
Protected double-stranded RNA duplexes were purified and resolved on a urea-
polyacrylamide denaturing gel. The gel was dried for I h at 80 C and bands were visualized
and quantified by phosphorimaging.
Statistical analysis
Post-challenge, data were analyzed as a 2 X2 factorial arrangement of treatments
within a split-plot design. Analysis of variance (ANOVA) was used to investigate the main
effects of vaccination and infection as well as the interaction between infective status and
vaccination. ANOVA was performed using the general linear model procedure (GLM) of
SAS. Differences with P < 0.05 were considered significant.
Results
Vaccination ameliorates the severity of B. hyodysenteriae-induced colitis
Following challenge, pigs were monitored daily for the development of clinical signs
associated with B. hyodysenteriae-induced colitis. No signs were observed until 4 days post
infection (figure 1). Disease progression from this point was similar for both of the groups
that were challenged although severity was milder in the group previously immunized with
the B. hyodysenteriae bacterin, as shown by the lower daily average score. Examination of
colonic sections recovered on day 15 post-infection revealed the typical microscopic lesions
associated with B. hyodysenteriae;-induced colitis. A total lesion score was generated based
on grading changes in gland depth, hyperplasia/metaplasia, lamina proprial vascular change,

71
submucosal change, inflammation, and superficial epithelial erosions (Table 1), as described
in the materials and methods section. No differences in total average cecal scores were found
between the different treatment groups (data not shown). In the colon, the average total score
of immunized and infected pigs was lower (15.6± 5.7) than that of the corresponding
challenged pigs not receiving immunization (19.6± 4.8). All infected pigs were culture
positive for B. hyodysenteriae the day they were necropsied. The histopathological score in
the non-infected groups was equal (11.3± 2.4and 11.33± 1.8) for the non-vaccinated and non-
infected and vaccinated and non-infected pigs.
Severe inflammation was manifested by increased mucosal thickness due to a
pronounced hyperplasia of crypt epithelial cells, which was accompanied by increased
cellular basophilia, and increased mitotic figures. This lesion was associated with multifocal
inflammatory cell infiltrates across the lamina propria muscularis (figure 2H). On the other
hand, pigs recovering from disease (as determined by clinical evaluation) showed goblet cell
metaplasia with excess mucus production (figure 2G). Non-vaccinated and infected pigs had
a 2.33 ± 0.44 hyperplasia/metaplasia average score, with 13 ± 2.67 gland depth whereas the
score for infected and vaccinated pigs were 1.66 ± 1.11 for hyperplasia/metaplasia, and
10.31 ±3.1 for gland depth. Thus, vaccinated and non-vaccinated groups differed in the
intensity rather than in the type of lesions developed following challenge.
Lymphocyte distribution in the colonic mucosa
Studies of the lymphocyte composition of the intestinal mucosa have revealed
regional specialization between the small and large intestine, and between lamina propria and
epithelium. Also, species-specific differences have been reported. Phenotypic
characterization of the small intestine of pigs revealed the absence of CD4+ cells within the

72
epithelium. Immunohistochemical analysis of colonic sections showed the same distribution
of CD4+ cells (figure 3B). yô T cells are present in both compartments of the colonic mucosa,
as lamina propria lymphocytes (LPL) and as intraepithelial lymphocytes (IEL) (figure 3C).
CDScT cells follow the same distribution as y8 T cells (not shown).
While yô LPL were dispersed in the tissue, clusters of CD4+ cells were present across
the lamina propria. These aggregates were more abundant in tissues recovered from B.
hyodysenteriae infected pigs (figure 4A-4C). In addition, immunohistological analysis of
longitudinal sections revealed that the multifocal cellular infiltrates found in the lamina
propria muscularis (figure 2H) were mainly composed of CD4+ T cells. yÔ T cells were also
detected in these lesions although in much smaller numbers (figures 4D-4F).
Changes in the lymphocyte subset composition of the colonic mucosa induced by
immunization and/or challenge.
Peripheral blood CD4^and yô T cells from immunized pigs respond in vitro to B.
hyodysenteriae antigens. In order to investigate whether immunization affects the
lymphocyte composition of the mucosa, CD3+, CD4+, yÔ T cells and CD8a* from the colonic
epithelium or lamina propria were enumerated. Immunization decreased the numbers of
CD4+ cells in the lamina propria (figure 5A). This effect was found irrespective of challenge
status. Changes were also detected at the epithelium, with lower numbers of epithelial T cells
(i.e., CD3+), consistent with a decrease in yô IEL. Vaccination, however, attenuated the loss
of this T cell subset (figure 5B).
Proinflammatory cytokine expression
A ribonuclease protection assay was utilized to investigate how infection and/or
immunization modulated the expression of cytokines in vivo (Figure 6). Transcriptional

73
activity of TNF-a and IL-1(3 was increased in the group of vaccinated and infected pigs when
compared to those pigs infected only and non-infected pigs. Immunization also upregulated
the expression of IL-1(3. In addition, vaccinated and infected pigs had higher expression of
TGF-(3, mRNA. Infection also stimulated the expression of IFN-y and IL-6 without any
significant differences between immunized and non-immunized pigs (data not shown).
Finally, transcriptional activity of IL-4 or IL-12p40 was undetectable (not shown).
Discussion
Protection induced by systemic immunization with a proteinase-digested bacterin of
B. hyodysenteriae correlates with changes in the lymphocytic composition of the colonic
mucosa and in the proinflammatory cytokine expression. B. hyodysenteriae is a non-invasive
bacterium which disrupts the colonic epithelium (14). Lymphocytes are part of the
population of leukocytes infiltrating the lamina propria of inflammatory lesions induced by
B. hyodysenteriae. Our results show that immunization decreased the numbers of CD4+ cells
in both infected and non-infected pigs. In the former group, these changes were accompanied
with a milder disease manifestation and less severe microscopic lesions. Interestingly, the
expression of the proinflammatory cytokines TNF-a and IL-1(3 m RNA was higher in
vaccinated pigs.
The cytokine profile of colonic tissue recovered from infected pigs is suggestive of a
predominant Thl response. In the majority of murine models of colitis, lesions result from
aberrant Thl responses, irrespective of the factor triggering the process (1). These pathogenic
CD4+ T cells are specific for antigens expressed by members of the normal flora (2; 4). The
presence of members of the normal colonic flora is also necessary for the development of B.

74
hyodysenteriae-induced colitis because disease was not detected in germ-free/gnotobiotic
pigs or when mice were selectively depleted of gram negative bacteria via antibiotics prior to
challenge (22-25). Antibiotic treatment has also proven to be effective in decreasing
intestinal inflammation in murine models of IBD (26; 27). Thus, part of the LP CD4^ cells
might be responding to antigens from the colonic flora.
An interesting finding is that vaccination alone modified the expression of
proinflammatory cytokines at the colonic mucosa. B. hyodysenteriae produces a
lipopolysaccharide (LPS) that stimulates the production of IL-1(3 and TNF-a by
macrophages (28; 29). However, because the vaccine was administered parenterally (i.e.,
intramuscular), the increase in proinflammatory cytokine expression is secondary to the
cellular immune response induced by vaccination. Peripheral blood CD8aar, CD4+, and yôT
cells from immunized pigs respond in vitro to B. hyodysenteriae antigens (17-19; 30). Also,
the production of IFN-y in antigen-recall responses to B. hyodysenteriae was shown to be
CD4-dependent (20). Thus, it is possible that immunization favors differentiation of naïve
cells into Thl cells that home to the colonic mucosa where they activate macrophages to
synthesize IL-1(3 and TNF-a. However, cytokine upregulation induced by previous
vaccination may contribute to epithelial regeneration. Specifically, IL-IJ3 is considered a
restitution-promoting cytokine (31). Skin keratinocytes-derived IL-1(3 stimulates the
production of keratinocyte growth factor (K.GF) by fibroblasts which, in turn favor
differentiation and proliferation of epithelial cells (32). This mechanism is dependent on the
transcription factors c-fos and c-jun, which are transcribed as early response genes during
epithelial restitution and modulate the expression of genes involves in cell migration (33; 34).

75
TGF-P, mRNA expression was increased in immunized and infected pigs. The
production of this cytokine by T reg cells has been associated with the downregulation of Th
I responses (7; 35). It has been shown that the same antigens that activate effector cells, can
also prime regulatory T cells (36; 37). Thus, it is possible that both Thl effector cells and T
reg cells are present at the colonic mucosa following immunization and challenge.
Additionally, yô T cells have been implicated in the downregulation of inflammatory
responses (38-40). Based on immunohistochemical analysis, the numbers of yô T cells were
not increased in vaccinated pigs, although the loss of yô IEL was less severe than in the non-
immunized and challenged group. It has been shown that yÔ T cells downregulate
inflammation by inducing apoptosis of activated aP T cells and activated macrophages (41;
42). Also yô IEL are important in maintaining epithelial homeostasis (43; 44). This subset in
addition contributes to the process of epithelial wound healing (45). The possible
contribution of yô T cells in protection against B. hyodysenteriae-induced colitis should be
addressed in future experiments. Finally, it has been shown that NK and NK-T cells provided
protection in murine models of colitis (46; 47). NK cells have been characterized in the pig
as CD3" CD8aa+. In addition, a subset of TCRotP CD8aa* has been identified as part of the
circulating lymphocyte pool (48; 49). Although CD8aa+ cells were analyzed during the
course of this study, several cell types within the mucosa express this coreceptor; thus, a
more specific phenotypic analysis should be performed in order to determine whether the
composition of NK cells or CD3" CD8aa+ subsets is modulated by infection and/or
vaccination.
Mechanisms involved in downregulation of inflammation are impaired in IBD. The
present study describes an immunization treatment that modulates the course of an
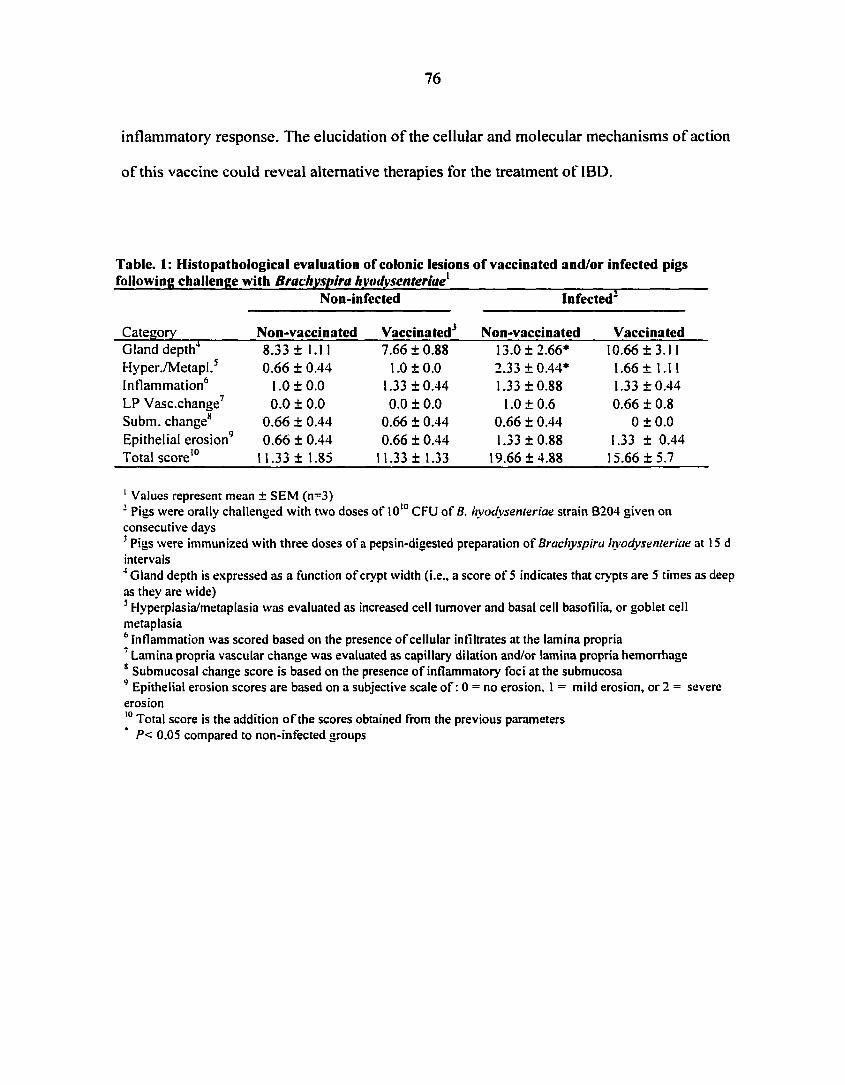
76
inflammatory response. The elucidation of the cellular and molecular mechanisms of action
of this vaccine could reveal alternative therapies for the treatment of IBD.
Tabic. I: Histopathological evaluation of colonic lesions of vaccinated and/or infected pigs following challenge with Brachyspira hyodysenteriae'
Category
Non-infected Infected*
Non-vaccinated Vaccinated Non-vaccinated Vaccinated Gland depth
HyperVMetapl.5
Inflammation6
LP Vasc.change7
Subm. change8
Epithelial erosion9
Total score 10
8.33 ± 1.11 0.66 ± 0.44
1.0 ± 0.0 0.0 ± 0.0
0.66 ± 0.44 0.66 ± 0.44
11.33 ± 1.85
7.66 ± 0.88 1.0 ±0.0
1.33 ±0.44 0.0 ± 0.0
0.66 ± 0.44 0.66 ±0.44
11.33 ± 1.33
13.0 ±2.66* 2.33 ±0.44* 1.33 ±0.88
1.0 ±0.6 0.66 ± 0.44 1.33 ±0.88
19.66 ±4.88
10.66 ±3.11 1.66 ± 1.11 1.33 ±0.44 0.66 ± 0.8
0±0.0 1.33 ± 0.44 15.66 ±5.7
' Values represent mean ± SEM (n=3) 2 Pigs were orally challenged with two doses of 1010 CPU of B. hyodysenteriae strain B204 given on consecutive days 3 Pigs were immunized with three doses of a pepsin-digested preparation of Brachyspira hyodysenteriae at 15 d intervals 4 Gland depth is expressed as a function of crypt width (i.e., a score of 5 indicates that crypts are 5 times as deep as they are wide) 5 Hyperplasia/metaplasia was evaluated as increased cell turnover and basal cell basofilia, or goblet cell metaplasia 6 inflammation was scored based on the presence of cellular infiltrates at the lamina propria 7 Lamina propria vascular change was evaluated as capillary dilation and/or lamina propria hemorrhage 8 Submucosal change score is based on the presence of inflammatory foci at the submucosa 9 Epithelial erosion scores are based on a subjective scale of : 0 = no erosion, 1 = mild erosion, or 2 = severe erosion 10 Total score is the addition of the scores obtained from the previous parameters
P< 0.05 compared to non-infected groups

77
e ti C«
4-
3-
2 -
1-
TAT
o-
jA rti111
7 12 Days post-infection
Non-vacc., non-inf ... Vacc., non-infected - Vacc., inf. — Non-vacc., inf.
Figure 1. Clinical scores of non-vaccinated and non-infected, vaccinated and non-infected, vaccinated and infected, and non-vaccinated and infected pigs. Vaccinated pigs received 3 doses of a pepsin-digested bacterin of B. hyodysenteriae. Pigs were challenged with 2 doses of 1010 CPU of B. hyodysenteriae given intragastrically on consecutive days. Diarrhea associated with the development of swine dysentery was monitored daily after challenge. Scores are based on stool consistency: (0), no diarrhea, stools formed; (1) mild, (2) moderate, and (3) severe diarrhea. Clinical scores were calculated by adding diarrhea score (i.e., 0 to 3) plus a 1 for the presence of blood and/or a 1 for the presence of mucus represents significant differences (P<0.05) between infected groups attributed to the effect of vaccination. Values are means ± SEM (n=3), from day 1 to day 13 post infection.

Figure 2. Photomicrographs of colonic mucosa stained with hematoxylin and eosin. Tissues were recovered from non-vaccinated and non-infected (A and E), vaccinated and non-infected (B and F), vaccinated and infected (C and G), and non-vaccinated and infected pigs (D and H) on day 15 post-challenge with B. hyodysenteriae. Inflammation was more severe in non-vaccinated and infected pigs (D and H). Protection induced by vaccination resulted in lesser mucosal thickening when compared to non-immunized pigs (C and G). Original magnification, top panel 63 X, bottom panel 160 X.

Non-vaccinated, non-infected

80
•S Figure 3. Photomicrographs of colonic sections from a representative non-vaccinated and non-infected pig stained for CD3, CD4 and TCR8 chain by immunohistochemistry. T cells are present, both in the lamina proprial and epithelial compartments (A). However, while CD4+ cells exist only as lamina propria lymphocytes (B), yô T cells are present in both epithelium and lamina propria (C).

Figure 4. Photomicrographs of colonic sections recovered on day 15 post-challege to depict CD4+ T cell distribution in the mucosa. Top panel: CD4+ T cell clusters in the lamina propria of non-vaccinated and infected pigs (C) were more abundant than in non-vaccinated and non-infected (A) or vaccinated and infected pigs (B) (indicated with arrowheads). Original magnification, 200 X. Bottom panel: hematoxylixn and eosin staining (D) and immunohistochemical staining (E and F) of the basal lamina propria revealed multifocal leukocytic infiltrates composed mainly of CD4+ T cells (E). Very few yô T cells were present at these inflammatory foci (F). Original magnification, 400 X.


83
Lamina propria
ë s a
I !
rr in c
« U
B
200
150-
100-
50-
Non-vacc., non-inf.
Vacc., non-inf. Vacc, inf . Non-vacc, inf „
i i r CD3 CD4 TCRyS CD8a
Epithelium
Non-vacc., non-inf.
Vacc., non-inf. Vacc, inf. Non-vacc, inf.
i r CD3 CD4 TCRyS CD8a
Phenotype
Figure 5. Changes in lymphocyte composition induced by vaccination and/or infection with Brcichyspira hyodysenteriae. Colonic sections recovered from non-vaccinated and non-infected, vaccinated and non-infected, vaccinated and infected, or non-vaccinated and infected pigs were analyzed by immunohistochemistry for the presence of CD3+, CD4+, TCR5+, and CD8a+cells in the lamina propria (A) and epithelium (B). Vaccination reduced the numbers of CD4+ lamina propria lymphocytes (*P<0.05). Infected pigs had lower numbers of y5 intraepithelial lymphocytes (5 P<0.05), although previous immunization ameliorated the loss of this subset. Values are means + SEM (n=3).

84
1 2 3 4 S 6 7 8 9 10 II 12 13 14 IS 16
******
« * * «* •
* 8 î = * | l #
TNF-a
IL-1P
TGF-P
L32
GAPDH
Figure 6. Detection of cytokine mRNA from colonic tissue using a ribonuclease protection assay. Full thickness colonic samples were obtained with a 3 mm-diameter punch biopsy from non-vaccinated and infected (1-4), vaccinated and infected (5-8), vaccinated and non-infected (9-12) and non-vaccinated and non-infected pigs (13-16) at day 15 post-challenge.

85
References
1. Strober, W„ Fuss, I. J., & Blumberg, R. S. (2002) The immunology of mucosal models of inflammation. Annu. Rev. Immunol. 20:495-549. 2. Cong, Y., Weaver, C. T., Lazenby, A., & Elson, C. O. (2000) Colitis induced by enteric bacterial antigen-specific CD4+ T cells requires CD40-CD40 ligand interactions for a sustained increase in mucosal IL-12. J. Immunol. 165: 2173-2182. 3. Czerkinsky, C., Anjuere, F., McGhee, J. R., George-Chandy, A., Holmgren, J., Kieny, M. P., Fujiyashi, K., Mestecky, J. F., Pierrefite-Carle, V., Rask, C., & Sun, J. B. (1999) Mucosal immunity and tolerance: relevance to vaccine development. Immunol Rev 170: 197-222. 4. Iqbal, N., Oliver, J. R., Wagner, F. H.. Lazenby, A. S., Elson, C. O., & Weaver, C. T. (2002) T helper 1 and T helper 2 cells are pathogenic in an antigen-specific model of colitis. J. Exp. Med. 195: 71-84. 5. Parronchi, P., Romagnani, P., Annunziato, F., Sampognaro, S., Becchio, A., Giannarini, L., Maggi, E., Pupilli, C., Tonelli, F., & Romagnani, S. (1997) Type I T-helper cell predominance and interleukin-12 expression in the gut of patients with Crohn's disease. Am. J. Pathol. 150: 823-832. 6. Romagnani, S. (1997) The Thl/Th2 paradigm. Immunol Today 18: 263-266. 7. Singh, B., Read, S.. Asseman, C.. Malmstrom, V., Mottet, C., Stephens, L. A., Stepankova, R., Tlaskalova, H., & Powrie, F. (2001) Control of intestinal inflammation by regulatory T cells. Immunol. Rev. 182: 190-200. 8. Groux, H., O'Garra, A., Bigler, M., Rouleau, M., Antonenko, S., de Vries, J. E., & Roncarolo, M. G. (1997) A CD4+ T-cell subset inhibits antigen-specific T-cell responses and prevents colitis. Nature 389: 737-742. 9. Chen, Y., Kuchroo, V. K., Inobe, J., Hafler, D. A., & Weiner, H. L. (1994) Regulatory T cell clones induced by oral tolerance: suppression of autoimmune encephalomyelitis. Science 265: 1237-1240. 10. Takahashi, T., Kuniyasu, Y., Toda, M., Sakaguchi, N., Itoh, M., Iwata, M., Shimizu, J., & Sakaguchi, S. (1998) Immunologic self-tolerance maintained by CD25+CD4+ naturally anergic and suppressive T cells: induction of autoimmune disease by breaking their anergic/suppressive state. Int. Immunol. 10: 1969-1980. 11. Harris, D. L.. & Glock, R. D. (1972) Swine dvsentery. J. Am. Vet. Med. Assoc. 160: 561-565. 12. Kennedy, M. J., Rosnick, D. K., Ulrich, R. G., & Yancey, R. J., Jr. (1988) Association of Treponema hyodysenteriae with porcine intestinal mucosa. J. Gen. Microbiol. 134 ( Pt 6): 1565-1576. 13. Harris, D. L., Alexander, T. J., Whipp, S. C., Robinson, I. M., Glock, R. D., & Matthews, P. J. (1978) Swine dysentery: studies of gnotobiotic pigs inoculated with Treponema hyodysenteriae, Bacteroides vulgatus, and Fusobacterium necrophorum. J. Am. Vet. Med. Assoc. 172: 468-471. 14. Hutto, D. L., & Wannemuehler, M. J. (1999) A comparison of the morphologic effects of Serpidina hyodysenteriae or its ̂ -hemolysin on the murine cecal mucosa. Vet. Pathol. 36: 412-422.

86
15. Hontecillas, R., Wannemuelher, M. J., Zimmerman, D. R., Hutto, D. L., Wilson, J. H., Ahn, D. U., & Bassaganya-Riera, J. (2002) Nutritional regulation of porcine bacterial-induced colitis by conjugated linoleic acid. J. Nutr. 132: 2019-2027. 16. Bassaganya-Riera, J., Hontecillas-Magarzo, R., Bregendahl, K., Wannemuehler, M. J., & Zimmerman, D. R. (2001) Effects of dietary conjugated linoleic acid in nursery pigs of dirty and clean environments on growth, empty body composition, and immune competence. J. Anim. Sci. 79: 714-721. 17. Bassaganya-Riera, J., Hontecillas, R., Zimmerman, D. R., & Wannemuehler, M. J. (2002) Long-term influence of lipid nutrition on the induction of CD8+ responses to viral or bacterial antigens. Vaccine 20: 1435-1444. 18. Waters, W. R., Pesch, B. A., Hontecillas, R„ Sacco, R. E., Zuckermann, F. A., & Wannemuehler, M. J. (1999) Cellular immune responses of pigs induced by vaccination with either a whole cell sonicate or pepsin-digested Brachyspira (Serpulina) hyodysenteriae bacterin. Vaccine 18: 711-719. 19. Waters, W. R., Hontecillas, R., Sacco, R. E„ Zuckermann, F. A., Harkins, K. R., Bassaganya-Riera, J., & Wannemuehler, M. J. (2000) Antigen-specific proliferation of porcine CD8aa cells to an extracellular bacterial pathogen. Immunology 101: 333-341. 20. Waters, W. R., Sacco, R. E., Dorn, A. D., Hontecillas, R., Zuckermann, F. A., & Wannemuehler, M. J. (1999) Systemic and mucosal immune responses of pigs to parenteral immunization with a pepsin-digested Serpulina hyodysenteriae bacterin. Vet. Immunol. Immunopathol. 69: 75-87. 21. Kunkle, R. A., & Kinyon, J. M. (1988) Improved selective medium for the isolation of Treponema hyodysenteriae. J Clin Microbiol 26: 2357-2360. 22. Hayashi, T., Suenaga, I., Komeda. T., & Yamazaki, T. (1990) Role of Bacteroides uniformis in susceptibility of Ta:CF# 1 mice to infection by Treponema hyodysenteriae. Zentralbl. Bakteriol. 274: 118-125. 23. Joens, L. A., Robinson, I. M., Glock, R. D., & Matthews, P. J. (1981) Production of lesions in gnotobiotic mice by inoculation with Treponema hyodysenteriae. Infect. Immun. 31:504-506. 24. Li, X. (1998) Characterization of acute proinflammatory cytokine responses within the cecal mucosafollowing infection with Serpulina hyodysenteriae. MSc Thesis. Iowa State University. 25. Nibbelink, S. K. (1992) Abrogation of cecal lesions, but not colonization, in conventional mice experimentally challenged with Serpulina hyodysenteriae and maintained on oral antibotics. Ph.D. dissertation. Iowa State University. 26. Steinhart, A. H., Feagan, B. G., Wong, C. J.. Vandervoort, M., Mikolainis, S., Croitoru, K., Seidman, E., Leddin, D. J., Bitton, A., Drouin, E., Cohen, A., & Greenberg, G. R. (2002) Combined budesonide and antibiotic therapy for active Crohn's disease: a randomized controlled trial. Gastroenterology 123: 33-40. 27. Farrell, R. J., & LaMont, J. T. (2002) Microbial factors in inflammatory bowel disease. Gastroenterol. Clin. North. Am. 31:41-62. 28. Sacco, R. E., Nibbelink, S. K., Baarsch, M. J., Murtaugh, M. P., & Wannemuehler, M. J. (1996) Induction of interleukin (IL)-ip and IL-8 mRNA expression in porcine macrophages by lipopolysaccharide from Serpulina hyodysenteriae. Infect. Immun. 64:4369-4372.

87
29. Greer, J. M., & Wannemuehler, M. J. (1989) Pathogenesis of Treponema hyodysenteriae: induction of interleukin-1 and tumor necrosis factor by a treponemal butanol/water extract (endotoxin). Microb. Pathog. 7: 279-288. 30. Bassaganya-Riera, J., Hontecillas, R., Zimmerman, D. R., & Wannemuehler, M. J. (2001) Dietary conjugated linoleic acid modulates phenotype and effector functions of porcine CD8+
lymphocytes. J. Nutr.131: 2370-2377. 31. Podolsky, D. K. (1997) Healing the epithelium: solving the problem from two sides. J. Gastroenterol. 32: 122-126. 32. Maas-Szabowski, N., Szabowski, A., Stark, H. J., Andrecht, S., fColbus, A., Schorpp-Kistner, M., Angel, P., & Fusenig, N. E. (2001) Organotypic cocultures with genetically modified mouse fibroblasts as a tool to dissect molecular mechanisms regulating keratinocyte growth and differentiation. J. Invest. Dermatol. 116: 816-820. 33. Angel, P., Szabowski, A., & Schorpp-Kistner, M. (2001) Function and regulation of AP-1 subunits in skin physiology and pathology. Oncogene 20: 2413-2423. 34. Angel, P., & Szabowski, A. (2002) Function of AP-1 target genes in mesenchymal-epithelial cross-talk in skin. Biochem Pharmacol 64:949-956. 35. Letterio, J. J., & Roberts, A. B. (1998) Regulation of immune responses by TGF-|3. Annu. Rev. Immunol. 16: 137-161. 36. McGuirk, P., & Mills, K.. (2002) Pathogen-specific regulatory T cells provoke a shift in the Thl/Th2 paradigm in immunity to infectious diseases. Trends. Immunol. 23: 450. 37. McGuirk, P., McCann, C., & Mills, K. H. (2002) Pathogen-specific T regulatory 1 cells induced in the respiratory tract by a bacterial molecule that stimulates interleukin 10 production by dendritic cells: a novel strategy for evasion of protective T helper type 1 responses by Bordetella pertussis. J. Exp. Med. 195:221-231. 38. Bagot, M., Echchakir, H., Mami-Chouaib, F., Delfau-Larue, M. H., Charue, D., Bernheim, A., Chouaib, S., Boumsell, L„ & Bensussan, A. (1998) Isolation of tumor-specific cytotoxic CD41" and CD4+CD8dim+ T-cell clones infiltrating a cutaneous T-cell lymphoma. Blood 91:4331-4341. 39. Born, W., Cady, C., Jones-Carson, J., Mukasa, A., Lahn, M., & O'Brien, R. (1999) Immunoregulatory functions of yô T cells. Adv. Immunol. 71: 77-144. 40. Hayday, A. C. (2000) yÔ T cells: a right time and a right place for a conserved third way of protection. Annu. Rev. Immunol. 18: 975-1026. 41. Vincent, M. S., Roessner, K., Lynch, D., Wilson, 0., Cooper, S. M., Tschopp, J., Sigal, L. H., & Budd, R. C. (1996) Apoptosis of Fash'8h CD4+ synovial T cells by borrelia-reactive Fas-ligandhlgh yô T cells in Lyme arthritis. J. Exp. Med. 184: 2109-2117. 42. Egan, P. J., & Carding, S. R. (2000) Downmodulation of the inflammatory response to bacterial infection by yô T cells cytotoxic for activated macrophages. J. Exp. Med. 191 : 2145-2158. 43. Boismenu, R., & Havran, W. L. (1994) Modulation of epithelial cell growth by intraepithelial yô T cells. Science 266: 1253-1255. 44. Boismenu, R., & Havran, W. L. (1998) yô T cells in host defense and epithelial cell biology. Clin. Immunol. Immunopathol. 86: 121-133. 45. Chen, Y., Chou, K., Fuchs, E., Havran, W. L., & Boismenu, R. (2002) Protection of the intestinal mucosa by intraepithelial yô T cells. Proc. Natl. Acad. Sci. U S A 99: 14338-14343.

88
46. Fort, M. M., Leach, M. W., & Rennick, D. M. (1998) A role for NK cells as regulators of CD4* T cells in a transfer model of colitis. J. Immunol. 161: 3256-3261. 47. Saubermann, L. J„ Beck, P., De Jong, Y. P., Pitman, R. S., Ryan, M. S., Kim, H. S„ Exley, M., Snapper, S., Balk, S. P., Hagen, S. J., Kanauchi, O., Motoki, K., Sakai, T., Terhorst, C., Koezuka, Y., Podolsky, D. K., & Blumberg, R. S. (2000) Activation of natural killer T cells by a-galactosylceramide in the presence of CD Id provides protection against colitis in mice. Gastroenterology 119: 119-128. 48. Yang, H., & Parkhouse, R. M. (1997) Differential expression of CD8 epitopes amongst porcine CD8-positive functional lymphocyte subsets.PG - 45-52. Immunology 92. 49. de Bruin, M. G., van Rooij, E. M., Voermans, J. J., de Visser, Y. E., Bianchi, A. T., & Kimman, T. G. (1997) Establishment and characterization of porcine cytolytic cell lines and clones. Vet. Immunol. Immunopathol. 59: 337-347.

89
CHAPTER 4. GENERAL CONCLUSIONS
The major focus of this dissertation has been the comparison between porcine CD4+
and yô T cells. However, the experiments conducted for this aim should be considered within
the long-term objective of achieving a better understanding of the immune regulation of
mucosal inflammation. The major strength of utilizing Brachyspira hyodysenteriae-induced
colitis is the possibility of evaluating systemic and mucosal responses resulting from
vaccination or infection, respectively. In addition, the fact that swine is among the species
with large numbers of circulating yô T cells, made it an ideal animal model for conducting
these studies.
Intramuscular administration of the B. hyodysenteriae bacterin does not prevent
infection and, in most of the cases it does not prevent dysentery. However, immunization
results in a milder disease manifestation as reflected by the clinical signs and microscopic
lesions described in chapter 3. Due to the functional compartmentalization of the mucosal
immune system, oral immunization is a requirement in order to mount a protective immune
response against mucosal pathogens (e.g., production of neutralizing IgA) (1; 2). Of course
this is not the case here: the route of immunization is not the appropriate to induce this type
of response, and colonization by the spirochete is not blocked by vaccination. Then, is
vaccination inducing an anti-inflammatory response rather than an anti-fi. hyodysenteriae
response? The major effects of vaccination reported here are I) a decrease in CD4+ T cells at
the colonic mucosa, 2) the 5. hyodysenteriae-induced proliferation of CD4+ and yô T cells in
vitro, and 3) the upregulation of IL-ip and, TNF-a and TGF-P mRNA expression following

90
infection, at the colonic mucosa. It is therefore expected that these cellular and molecular
events contribute to the mechanisms of protection.
Results presented in chapter 2 suggest that a cross-talk between CD4+ and yô T cells
occurs following stimulation with B. hyodysenteriae antigens, in which IL-2 appears to have
a synergistic effect. This type of interaction was described in vivo in murine models of
influenza and listeriosis. In both cases, yô T cells were necessary for downregulating
inflammation (3-6). Thus it appears that several infectious diseases with an inflammatory
component trigger this response. Very interestingly, in Lyme disease, which is caused by
another spirochete, Borrelia burgdorferi, responses of CD41* T cells isolated from synovial
fluid were downregulated by yô T cells. Lipoproteins from B. burgdorferi were later shown
to activate yÔ T cells (7; 8). It is evident that CD4+ T cell proliferation described in chapter 2
is B. hyodysenteriae-spccific. The specificity of the yô T cells is unclear, although it is
possible that yÔ T cells are responding to B. hyodysenteriae antigens recognized through a
different mechanism than CD4+. Thus, the capacity of B. hodysenteriae antigens to stimulate
yô T cells in the absence of CD4+ T cells should be investigated in the future.
yô T cells induce apoptosis of activated CD4+T cells and macrophages. The
selectivity for activated cells has been explained on the basis of the high expression level of
Fas on target cells, whereas yô T cells constitutively express FasL (7; 9). Because
immunohistochemical analysis did not reveal a significant increase of yô T cells at the
colonic mucosa of B. hyodysenteriae vaccinated pigs, initially this does not appear to be the
mechanism that explains the decrease of CD4+ LPL. Alternatively it could be possible that

91
this response took place soon after challenge. A time course study would be necessary to test
this hypothesis.
The decrease of CD4^ T cells in protected animals was a preliminary observation
from previous experiments, which was confirmed here. As it was discussed in chapters 1 and
2, CD4+ T cells specific for intestinal flora antigens are involved in the pathogenesis of IBD.
Therefore vaccination had the beneficial effect of lowering a cell type putatively involved in
the pathogenesis of swine dysentery. Intestinal colonization by B. hyodysenteriae alone is
insufficient for the development of swine dysentery as it was demonstrated in experiments
with gnotobiotic pigs, or when mice were treated with antibiotics that eliminated specific
bacteria from the colonic commensal flora (10-12). Besides the possible interaction described
above, two additional hypotheses can explain the effect that immunization has on CD4+ LPL.
First stimulation of regulatory T cells by vaccination. Second, vaccination could partially
block inflammation (i.e., lower epithelial damage), or accelerate the process of epithelial
restitution. The upregulation of TGF-Pi mRNA expression is consistent with the stimulation
of regulatory T cells (13; 14). These results are in agreement with those reported in a murine
model in which vaccination and infection with B. hyodysenteriae increased antigen-specific
TGF-P protein levels in mesenteric lymph nodes. Measurement of bioactive TGF-P protein
in the colon would further support this hypothesis (15).
Vaccination also upregulated the expression of IL-ip and TNF-a mRNA. The
implications of these results are intriguing. These cytokines are a hallmark of
proinflammatory responses, yet they also have a role in epithelial wound healing (16; 17). It
was previously reported that mice challenged with B. hyodysenteriae upregulated colonic
TNF-a m RNA 24 hours post-infection. The increased expression of IL-ip did not occurr

92
until 5 days post-infection (12). This is the first study that evaluates the interaction between
vaccination and infection in regard to the in vivo modulation of cytokine expression. Again, a
time-course study seems to be necessary in order to determine whether these changes are
beneficial or, in the contrary, this is a negative effect of the vaccine. The increased
transcriptional activity of these cytokines, especially of IL-1(3, is most likely due to B.
hyociysenteriae lipopolysaccharide. Stimulation of murine peritoneal exudate cells or porcine
alveolar macrophage macrophage with B. hyociysenteriae LPS increased the expression of
IL-1(3 (18). Evaluation of protection against B. hyociysetitericie-'induced colitis in pigs
immunizd with a preparation of B. hyociysenteriae LPS would clarify whether the
upregulation of IL-1(3 mRNA has a beneficial effect.
Because of their complexity, the use of porcine models for immunological studies has
the disadvantage of the difficulty in analyzing the role of specific factors within a cellular or
molecular mechanism. In the case of immunization and or vaccination with B.
hyociysenteriae, the evaluation of immune responses at the single cell level would aid in
elucidating the mechanism of action of the vaccine, or the molecular pathogenesis of with B.
hyociysenteriae-induced colitis. Specifically, in vitro experiments with purified
subpopulations of yô or CD4+T cells will be necessary for the elucidation of the exact
mechanism of activation by B. hyociysenteriae antigens or by other stimuli (i.e., IL-2). In
regards to the regulation of inflammation associated with swine dysentery, there are two
aspects of major priority: the identification of the cellular source of cytokines involved in
immune regulation and the expression of molecules that participate in cell-cell interactions at
the mucosa.

93
References
1. Czerkinsky, C., Anjuere, F., McGhee, J. R., George-Chandy, A., Holmgren, J., Kieny, M. P., Fujiyashi, K., Mestecky, J. F., Pierrefite-Carle, V., Rask, C., & Sun, J. B. (1999). Mucosal immunity and tolerance: relevance to vaccine development. Immunol. Rev. 170: 197-222. 2. Kraehenbuhl, J. P., & Neutra, M. R. (1992). Molecular and cellular basis of immune protection of mucosal surfaces. Physiol. Rev. 72: 853-879. 3. Doherty, P. C., Allan, W., Eichelberger, M., & Carding, S. R. (1992). Roles of a(3 and yô T cell subsets in viral immunity. Ann. Rev. Immunol. 10: 123-151. 4. S keen, M. J., & Ziegler, H. EC (1993). Intercellular interactions and cytokine responsiveness of peritoneal a(3 and yô T cells from Listeria-infected mice: synergistic effects of interleukin I and 7 on yô T cells. J. Exp. Med. 178: 985-996. 5. Skeen, M. J., & Ziegler, H. K. (1993). Induction of murine peritoneal yô T cells and their role in resistance to bacterial infection. J. Exp. Med. 178: 971-984. 6. Carding, S. R., Allan, W., McMickle, A., & Doherty, P. C. (1993). Activation of cytokine genes in T cells during primary and secondary murine influenza pneumonia. J. Exp. Med. 177: 475-482. 7. Vincent, M. S., Roessner, K., Lynch, D., Wilson, D., Cooper, S. M., Tschopp, J., Sigal, L. H„ & Budd, R. C. (1996). Apoptosis of Fash'8h CD4+ synovial T cells by borrelia-reactive Fas-ligandh'eh yô T cells in Lyme arthritis. J. Exp. Med. 184: 2109-2117. 8. Vincent, M. S., Roessner, K.., Sellati, T., Huston, C. D., Sigal, L. H., Behar, S. M., Radolf, J. D., & Budd, R. C. (1998). Lyme arthritis synovial yô T cells respond to Borrelia burgdorferi lipoproteins and lipidated hexapeptides. J. Immunol. 161: 5762-5771. 9. Egan, P. J., & Carding, S. R. (2000). Downmodulation of the inflammatory response to bacterial infection by yô T cells cytotoxic for activated macrophages. J. Exp. Med. 191 : 2145-2158. 10. Hayashi, T.. Suenaga, L, Komeda. T., & Yamazaki, T. (1990). Role of Bacteroides uniformis in susceptibility of Ta:CF# 1 mice to infection by Treponema hyodysenteriae. Zentralb.l Bakteriol. 274: 118-125. 11. Nibbelink, S. K. (1992). Abrogation of cecal lesions, but not colonization, in conventional mice experimentally challenged with Serpulina hyodysenteriae and maintained on oral antibotics. Ph.D. dissertation. Iowa State University. 12. Li, X. (1998). Characterization of acute proinflammatory cytokine responses within the cecal mucosafollowing infection with Serpulina hyodysenteriae. MSc thesis. Iowa State University. 13. Singh, B., Read, S., Asseman, C., Malmstrom, V., Mottet, C., Stephens, L. A., Stepankova, R., Tlaskalova, H., & Powrie, F. (2001). Control of intestinal inflammation by regulatory T cells. Immunol. Rev. 182: 190-200. 14. Letterio, J. J., & Roberts, A. B. (1998). Regulation of immune responses by TGF-0. Ann. Rev. Immunol. 16: 137-161. 15. Khalifeh, M. S. (1999). The role of T cell mediated immune response in the development and resolution of colitis induced by Serpulina hyodysenteriae. MSc Thesis. Iowa State University. Iowa State University.

94
16. Strober, W., Fuss, I. J., & Blumberg, R. S. (2002). The immunology of mucosal models of inflammation. Ann. Rev. Immunol. 20:495-549. 17. Podolsky, D. K. (1997). Healing the epithelium: solving the problem from two sides. J. Gastroenterol. 32: 122-126. 18. Sacco, R. E., Nibbelink, S. K., Baarsch, M. J., Murtaugh, M. P., & Wannemuehler, M. J. (1996). Induction of interleukin (IL)-l (3 and IL-8 mRNA expression in porcine macrophages by lipopolysaccharide from Serpulina hyodysenteriae. Infect. Immun. 64: 4369-4372.

95
ACKNOWLEDGEMENTS
The author wishes to express her most sincere appreciation to Dr. Michael J.
Wannemuehler, her major professor for sharing his ideas, talent and mentorship with the
author throughout this doctoral dissertation.
Acknowledgements are also extended to Dr. Jan Buss, Dr. Susan Lamont, Dr. Harley
W. Moon and Dr. Randy E. Sacco for their assistance and advice while serving as members of
this Ph.D. committee at Iowa State University.
The author is very grateful to Dr. Josep Bassaganya-Riera, whose friendship,
constructive criticisms, positive intellectual interactions and assistance in experimental work
have converted immunology research into an enjoyable task and the author's graduate
program into a highly valuable learning experience.
The author deepest and kindest appreciation is expressed to her family for their
support and encouragement.





















![Porcine Epidemic Diarrhea [Autosaved]](https://img.pdfslide.us/doc/110x75/577c808c1a28abe054a92a69/porcine-epidemic-diarrhea-autosaved.jpg)