Embed Size (px)
Citation preview

ANNALS OF MICROSCOPY Vol 10, May 2010
36
Comparison of Chikungunya Virus Infectivity in Baby Hamster Kidney (BHK-21) Cell Line and C6/36, an Aedes albopictus Cell Line using Electron Microscopy
TS Saraswathy1*, Tengku Rogayah Tar1, AS Santana Raj2 , Aida Suhana R2, Teh H2 and Zainah S1
1 Virology Unit, Infectious Diseases Research Centre, Institute for Medical Research, Jalan Pahang, 50588 Kuala Lumpur2 Electron Microscopy Unit, Medical Research Resource Centre, Institute for Medical Research, Jalan Pahang, 50588 Kuala Lumpur*Corresponding author.
ABSTRACT
The infection and morphogenetic events associated with the replication of Chikungunya virus (CHIKV) in baby hamster kidney (BHK-21) and C6/36 cell cultures were compared. Confl uent cell cultures of BHK-21 and C6/36 were prepared in 25 cm2 fl asks and were inoculated with CHIKV from our laboratory stock. Infected cultures, from 24 hour post infection onwards were fi xed in buffered 2% glutaraldehyde at 24 hour intervals and processed for thin section electron microscopy. The intracellular mechanisms of CHIKV infection in these two cell cultures differed considerably. Infected BHK-21 showed cytopathic effects (CPE) after 24 hours post infection. Severe CPE with presence of large vacuoles in the cytoplasmic matrix were seen within 48 hours of post infection followed by cell lysis caused by rupture of plasma membrane. Newly assembled viruses budded out of the cell from the plasma membrane. In contrast the CPE in C6/36 culture was only observed on the 7th day. Cell monolayer integrity was preserved; however there was intense proliferation of intracellular membranous vesicles in the cytoplasmic matrix. On the 10th day post infection, electron dense virus precursors were observed in these vesicles. Our electron microscopic study has demonstrated that CHIKV have developed a delicate balance between aggressive replication in a vertebrate host and limited pathology in their mosquito host that allows effi cient vectoring to ensure continuity and fi tness of the virus.
Keywords: Chikungunya virus, ultrastructure, morphogenesis, BHK-21, C6/36
INTRODUCTION
Chikungunya virus (CHIKV) is an arbovirus that belongs to the genus alphavirus and family Togaviridae. The virus was fi rst isolated during an outbreak of dengue-like fever in Tanzania, Africa in 1952 (Johnston and Peters, 1996). Subsequently the virus was introduced into Asia and caused major outbreaks in many parts of South and South-east Asia. Recently from 2004 till 2008, re-emergence of CHIKV outbreaks were reported in several islands in the Indian Ocean, (the French Reunion Islands, Mauritius and Seychelles) as well as in southern India and South east Asia, including Indonesia, Singapore and Malaysia. Imported cases of the disease were also reported in Hong Kong, Taiwan, USA, and Europe (Lahariya and Pradhan, 2004; Pialoux et al, 2007)
The two main vectors of the virus are Aedes aegypti and Aedes albopictus; the infection is transmitted to humans through the bite of infected mosquitoes. Symptoms of CHIKV infection are characterized by acute onset of joint pain, followed by myalgia, fever, and maculo-papular rash on the trunk and limbs with recovery usually within weeks. Recent outbreaks have reported unusual severe forms of the disease which include chronic and persistant arthralgia, neurological disease and mortality among elderly with underlying conditions. Mother-to-child transmission has also been observed and is associated with severe neonatal disease such as encephalopathy (Powers and Logue, 2007).
The occurance of these outbreaks with unusual severity of the disease as well as the spread of Aedes mosquitoes in other areas of the world has highlighted an urgent need to increase our

ANNALS OF MICROSCOPY Vol 10, May 2010
37
basic understanding of CHIKV biology and pathophysiology induced in their hosts. CHIKV belong to the Alphavirus family which have been extensively studied using Semliki Forest (SF) and Sindbis (SIN) viruses as prototypes. However, there is a critical lack of knowledge on our understanding of CHIKV, in particular, the pathology of CHIKV infection in, in vitro host systems and the interaction between the CHIKV and their vectors. The virus can easily be propagated in cell culture using a number of vertebrate fi broblast and continuous cell lines, for example, Baby Hamster Kidney (BHK) and VERO cell lines and also in invertebrate cell lines, particularly, C6/36, an Aedes albopictus cell line. These cell culture systems serve as an excellent tool for studying virus infectivity.
The objective of this study was to compare CHIKV infectivity in BHK and C6/36 cell lines using transmission electron microscopy. The ultrastructural details of CHIKV morphogenesis in these infected cell cultures were compared with a literature review of other alphavirus replication in vertebrate and mosquito host systems.
MATERIALS AND METHODS
Cell lines Two cell lines, a vertebrate-derived fi broblast cell line, BHK-21 (baby hamster kidney) and
a mosquito-derived, C6/36 (Ae. albopictus) cell line were used for these studies. A monolayer culture of Aedes albopictus cells, clone C6/36 in passage 121, and BHK-21 in passage 53 was obtained from ATCC. The C6/36 cells were grown in Leibovitz medium (L-15) containing 10% inactivated fetal calf serum 100 U/mL penicillin, and 100 μg/mL streptomycin and supplemented with 10% Tryptose Phoshate Broth. The Leibovitz maintenance medium consisted of 2% fetal calf serum. BHK-21 cells were grown in Hanks Minimun Essential Medium with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 μg/mL streptomycin. Monolayer cultures were grown in plastic 25-cm2 fl asks. Stock cultures split at a 1 to 4 ratio and seeded at 2 x 105 cells per ml. BHK-21 cultures generally formed complete monolayers within 36 hours while C6/36 formed monolayers after 72 hours. BHK-21 and C6/36 mosquito cells were maintained at 37°C and 28°C, respectively.
Viruses The CHIKV was fi rst isolated in our laboratory from human serum in 2006 and has since
been passaged in BHK-21 cell line; this isolate has been sequenced and is similar genetically to Asian genotype (NCBI GenBank accession number EU 703759).
Stock viruses were produced after a single passage in BHK-21 cells maintained at 37°C in Hanks Minimun Essential Medium with 2% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 μg/mL streptomycin. Cell supernatants were harvested when 75% of the cells showed cytopathic effect (CPE 3+), aliquoted, and stored at –70°C for use in all experiments.
Virus titrations For each virus, confl uent cell monolayers in 25 cm2 fl asks were infected with a standard
250uL inoculum by rocking at room temperature for 1 hour. Viral samples harvested from cell culture were quantifi ed as tissue culture infectious dose 50 end-point titers (log10 TCID50/mL). Briefl y, 100μL samples of cell culture supernatant were pipetted into wells of the fi rst column of a 96-well plate, serially diluted in a 10-fold series, seeded with BHK-21 cells, and incubated at 37°C for 7 days.
Electron microscopy The 25 cm 2 fl asks, seeded with BHK-21 cells and C6/36 cells and infected with CHIKV
were observed for cytopathological changes every day using a phase contrast microscope. Each

ANNALS OF MICROSCOPY Vol 10, May 2010
38
day from 24 hours post infection (p.i.) onwards, the infected BHK-21 cells and infected C6/36 cells were rinsed in phosphate buffered saline (PBS), then fi xed in 2.5 % glutaraldehyde in 0.1% phosphate buffer, pH 7.4 for 2 hours at room temperature. The fi xed cells were scraped and pelleted before post-fi xing in PBS-1% osmium tetroxide for 20 minutes. If cells had detached due to advanced CPE, the cells were fi rst scraped gently, pelleted, rinsed in PBS before fi xation procedures. After rinsing, 3 changes in distilled water, the fi xed cells were pre-stained en bloc with 2% uranyl acetate for 10 minutes. After rinsing with fresh distilled water, the fi xed cells were dehydrated in an ethanol series (50% ethanol for 3 minutes, 70% ethanol for 3 minutes, 90% ethanol for 3 minutes and 2 changes of 100% ethanol for 3 minutes. This was followed by 2 changes of propylene oxide for 5 minutes each and then infi ltration with a mixture of propylene oxide and epoxy; with a fi nal infi ltration in pure resin and fi nal embedding in fresh epoxy resin. The blocks were trimmed and thin sections (0.5 micron) were stained in 4% uranyl acetate and Reynold’s lead citrate for examination using a Technai G2 Transmission Electron Microscope.
RESULTS
BHK-21 cell cultures showed fi rst signs of cytopathic effects (CPE) after 24 hours post infection (p.i.). The CPE developed rapidly in the BHK-21 cultures with subsequent total monolayer destruction within 48 hours. The initial events of CPE include rounding of cells and loss of cell to cell contacts after 24 to 36 hours. This was followed by classical features of programmed cell death or apoptosis such as nuclear condensation, membrane rupture, cytoplasmic vacuolization, cell fragmentation, and fi nally cell death.
These events were not observed in the mosquito cell line. Infected C6/36 cells became syncytia with cell to cell fusion at a late stage of the infection, after 7 days of p.i.. Some cells detached from the fl ask. In this study the sequence of events in the infected BHK-21 and C6/36 cell culture has been captured by transmission electron microscopy.
CHIKV infected BHK-21 cells (as seen in our electron micrograph image in Figure 2a) had undergone dramatic changes and exhibited unique membrane structures within the cytoplasm.
Figure 1a shows an uninfected BHK-21 cell. Figure 1b shows a BHK-21 cell after 24 hour post infection with CHIKV. After 24 hour p.i., some extracellular virions were still observed outside the plasma membrane of infected BHK-21 cells. The enlarged viral particles shown in the inset, were roughly spherical (about 58 to 65 nm in diameter) containing a densely stained inner core of about 40 nm (the icosahedral nucleocapsid) and surrounded by a translucent layer (lipid envelope) with a fi ne fuzzy fringe on the outer surface (the glycoprotein spikes). Virions were not present on the surface of the uninfected cell.
Figure 1a Figure 1b
ANNALS OF MICROSCOPY Vol 10, May 2010
39
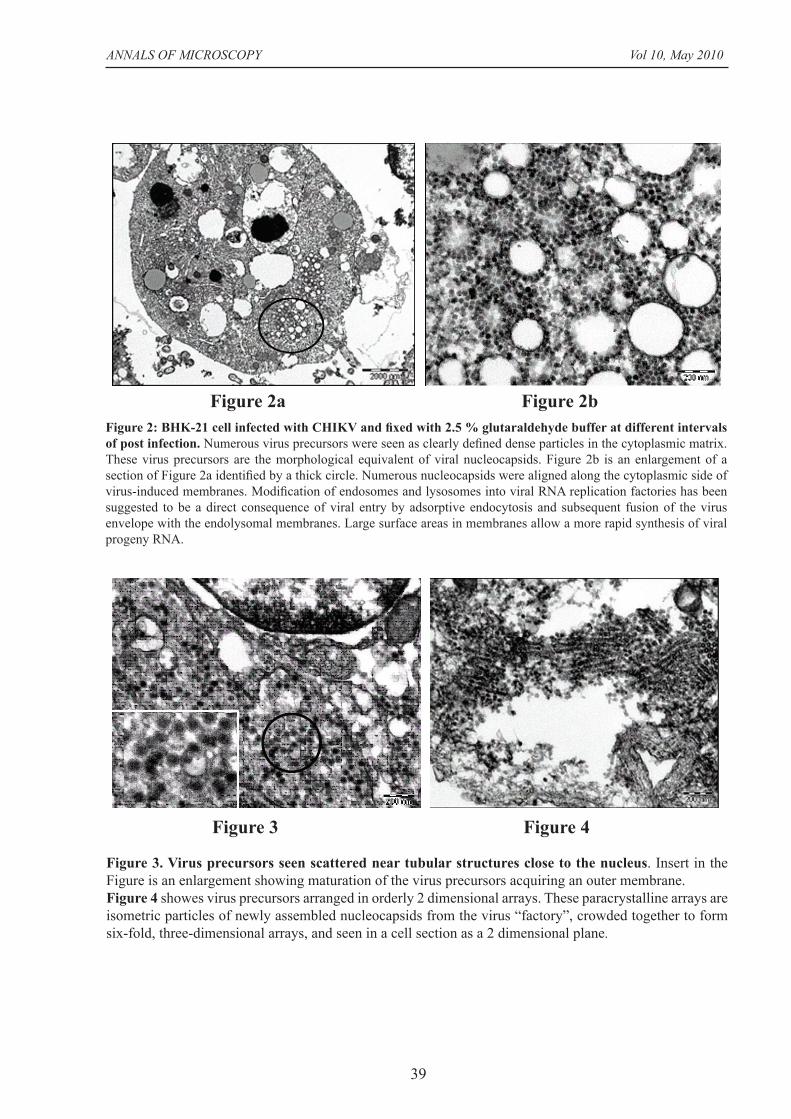
Figure 3. Virus precursors seen scattered near tubular structures close to the nucleus. Insert in the Figure is an enlargement showing maturation of the virus precursors acquiring an outer membrane. Figure 4 showes virus precursors arranged in orderly 2 dimensional arrays. These paracrystalline arrays are isometric particles of newly assembled nucleocapsids from the virus “factory”, crowded together to form six-fold, three-dimensional arrays, and seen in a cell section as a 2 dimensional plane.
Figure 3 Figure 4
Figure 2: BHK-21 cell infected with CHIKV and fi xed with 2.5 % glutaraldehyde buffer at different intervals of post infection. Numerous virus precursors were seen as clearly defi ned dense particles in the cytoplasmic matrix. These virus precursors are the morphological equivalent of viral nucleocapsids. Figure 2b is an enlargement of a section of Figure 2a identifi ed by a thick circle. Numerous nucleocapsids were aligned along the cytoplasmic side of virus-induced membranes. Modifi cation of endosomes and lysosomes into viral RNA replication factories has been suggested to be a direct consequence of viral entry by adsorptive endocytosis and subsequent fusion of the virus envelope with the endolysomal membranes. Large surface areas in membranes allow a more rapid synthesis of viral progeny RNA.
Figure 2a Figure 2b

ANNALS OF MICROSCOPY Vol 10, May 2010
40
DISCUSSION
CHIKVs belong to the genus alphavirus and like other arboviruses alternate in nature, between replication in higher vertebrates and arthropod hosts, namely, Aedes albopictus and Aedes aegypti mosquitoes. Several signifi cant differences in the infection process between the vertebrate and invertebrate systems have been described for arboviruses. Alphavirus infection of vertebrates is acute and may result in illness with symptoms of fever, arthralgia and encephalitis. The infection of adult mosquitoes with alphaviruses results in a persistent infection that lasts the lifetime of the mosquito but that leads to relatively little cytopathology and mortality. This non lethal interaction is essential to ensure continuation of transmitting the the virus to the next available vertebrate host by the insect. Viral persistence represents a complex interaction of virus and host wherein innate immune responses are suppressed to some extent allowing viral replication and specifi c pathways important to regulate co-existence with the virus are activated.
The characteristics of alphavirus infection of tissue culture cells mimic what is observed in vivo. Infection of most vertebrate cell lines results in massive cell death within 24 to 48 hours, due to shut down of host cell macromolecular synthesis. In mosquito cells, the development of infection results in a prolonged persistent phase in which virus production is maintained at low levels and the cells continue to grow and divide through many passages. The progression of the infection into persistence is accompanied by an ability of the cell to control, suppress cell death
Figure 5: Gallery of electron micrographs showing events leading to cell deathLarge vacuoles in infected BHK-21 cell precede cell rupture. CHIKV infection results in a drastic inhibition of host macromolecular synthesis, since the virus utilizes the basic cellular mechanisms and intracellular organelles for the effi cient replication of viral progeny. Severe CPE results in rupture of plasma membrane as newly assembled virus bud out of the cell. Virus progeny acquire part of plasma membrane as they egress from the cell.
ANNALS OF MICROSCOPY Vol 10, May 2010
41
and coexist with the virus.In our study, CHIKV infection in BHK-21 cells were associated with CPE of infected cells
that ultimately resulted in cell lysis, within 36 to 48 hours p.i. After 24 hours of p.i. of BHK-21 cell culture, CHIKV virions were observed outside the plasma membrane. Alphaviruses are often described as having little ultrastructural detail. The virions in our images displayed the characteristic features of alphaviruses: spherical, a size of 60–70 nm, an icosahedral-like shell of C proteins surrounded by the presence of a lipid envelope with heterodimeric glycoprotein spikes on their surface.
Alphavirus replication has been elucidated using Sindbis viruses (SINV) and Semliki Forest virus (SFV). Many types of protein receptors can be involved in alphavirus intake at the host plasma membrane. This may explain the wide host range of alphaviruses and the capability to cause infection in various tissues. Most receptors are poorly characterized, (Kääriäinen et al,
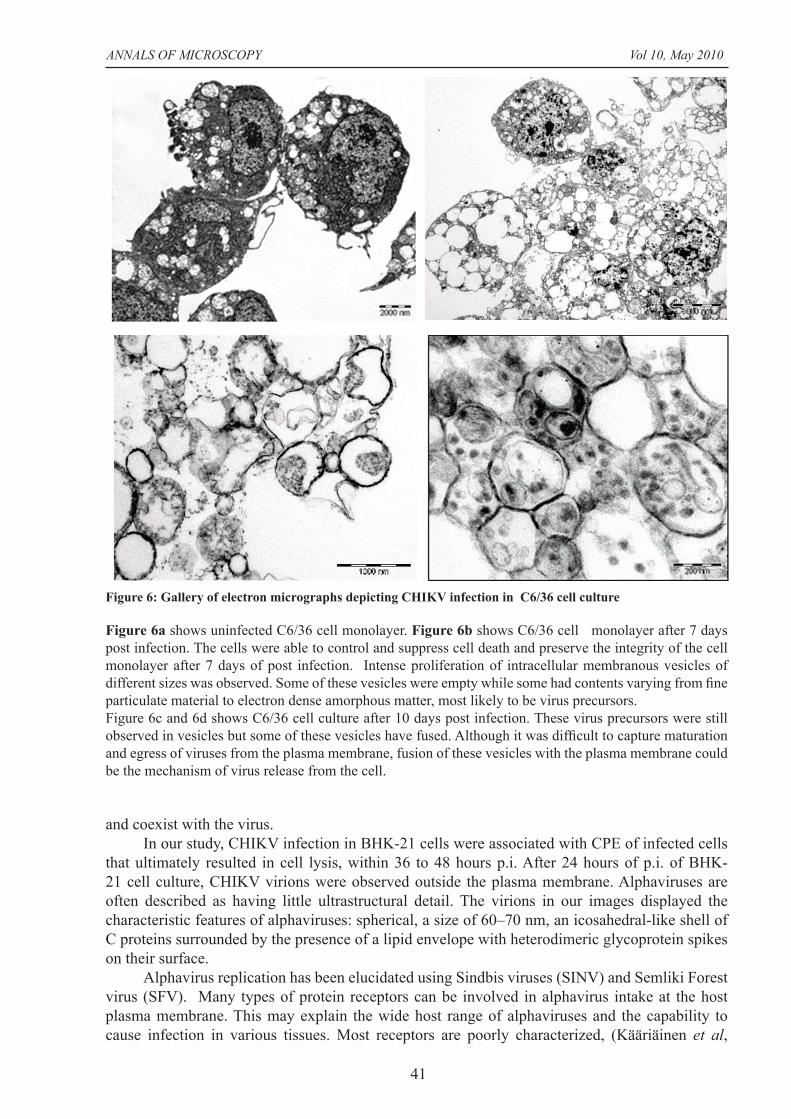
Figure 6: Gallery of electron micrographs depicting CHIKV infection in C6/36 cell culture
Figure 6a shows uninfected C6/36 cell monolayer. Figure 6b shows C6/36 cell monolayer after 7 days post infection. The cells were able to control and suppress cell death and preserve the integrity of the cell monolayer after 7 days of post infection. Intense proliferation of intracellular membranous vesicles of different sizes was observed. Some of these vesicles were empty while some had contents varying from fi ne particulate material to electron dense amorphous matter, most likely to be virus precursors.Figure 6c and 6d shows C6/36 cell culture after 10 days post infection. These virus precursors were still observed in vesicles but some of these vesicles have fused. Although it was diffi cult to capture maturation and egress of viruses from the plasma membrane, fusion of these vesicles with the plasma membrane could be the mechanism of virus release from the cell.

ANNALS OF MICROSCOPY Vol 10, May 2010
42
1987). SINV and SFV penetrate target cells through clathrin-mediated endocytosis, the low pH of the endosomal compartment promoting conformational changes of envelope glycoprotein and viral fusion. Sourisseau et al (2007), studied the role of the clathrin endocytic pathway in CHIKV replication by knocking down the expression of dynamin-2 (Dyn-2), a key protein required for the formation of clathrin-coated pits and vesicles. Their study indicated that CHIKV entry into vertebrate cells is mediated through pH dependant endocytosis.
Viruses are known to induce intracellular membrane alterations for the effi cient replication of their genomes. This is because of their limited genetic capacity, the viruses utilize the basic host cellular mechanisms for entry, protein synthesis and intracellular transport. CHIKV are enveloped, single stranded positive polarity RNA viruses. Like other alphaviruses, the viral (+) RNA genome in CHIKV has the same polarity as cellular mRNA. Therefore, the genome can be translated by the host cell translation machinery into one or multiple viral polyproteins, which are co- and post-translationally cleaved by viral and host cell proteases into proteins. CHIKV replication as in other alphaviruses takes place in the cytoplasm of infected cells in association with characteristic virus-induced cytopathic vacuoles (CPVs). CPVs are modifi ed endosomal and lysosomal structures that are 600–2,000 nm in size. The CPVs serve as sites of RNA replication and perhaps as sites of RNA translation and nucleocapsid assembly and are unique to the Togaviridae family. During alphavirus infection two main types of morphologically distinct, virus-induced cytopathic vacuoles have been described. Cytopathic vacuoles of type I (CPV-I) form in the early phase in infection (Acheson and Tamm, 1967, Grimley et al., 1968). They represent the sites of viral RNA replication. Type II (CPV-II) vacuolar structures form in the late phase in alphavirus infection and consist of numerous nucleocapsids attached to the cytoplasmic face of membranes (Grimley et al., 1972). This is due to spike proteins, which tend to accumulate in the Golgi complex late in infection and start to bind nucleocapsids. Our electron micrograph analysis (Figure 2) showed a type 2 CPV with numerous virus precursors outside these vacuoles. Protein synthesis is mediated by the cellular ribosome and endoplasmic reticulum–based translocation machinery. The virus maturation of the virus precursors were seen close to the nucleus in our images, in association with tubular structures. The assembly of matured viruses was observed to take place by a budding process from the plasma membrane into the extracellular space.
In mosquito cells, the development of infection results in a prolonged persistent phase in which virus production is maintained at low levels and the cells continue to grow and divide through many passages. The progression of the infection into persistence is accompanied by an ability of the cell to control, suppress cell death and coexist with the virus. From our study, the intracellular mechanisms of CHIKV infection in C6/36 cell cultures differed considerably compared to infection in BHK-21 cell culture. CPE was only observed on the 7th day. Cell monolayer integrity was preserved although there was intense proliferation of intracellular membranous vesicles of different sizes. Some of these vesicles were empty while some had contents varying from fi ne particulate material to electron dense amorphous matter, most likely to be virus precursors. Virus assembly occurs within these intracellular vesicles or “virus factories”. Only a small percentage of persistently infected insect cells produce infectious virus. Gliedman et al. (1975) studied SINV replication in Aedes albopictus cells and reported that free nucleocapsids were rarely seen in the cytoplasm of infected mosquito cells, and that budding of the virus from the cell surface was detected so infrequently that this process of virus production could not account for the amount of virus produced by the infected cells. The vast majority of extracellular virus was produced by the fusion of the virus-containing vesicles with the plasma membrane releasing mature virions and
membrane nucleocapsid complexes in various stages of development.
It appears that CHIKV have developed a delicate balance between aggressive replication in mammalian host and limited pathology in the mosquito host that allows persistent infection and effi cient vectoring to ensure continuity and fi tness of the virus. Our study has also demonstrated that BHK-21 is a good cell line to isolate CHIKV since these viruses cause rapid death of infected BHK-21 cells within 48 hours by apoptosis with effi cient release of viruses in the culture medium.

ANNALS OF MICROSCOPY Vol 10, May 2010
43
ACKNOWLEDGEMENTS
We are grateful to the Director General of Health, Malaysia and the Director of the Institute for Medical Research, Kuala Lumpur for permission to publish this paper.
REFERENCES
Ache son, N.H. and Tamm, I. Replication of Semliki Forest virus: An electron microscopic study. Virology. 1967; 32: 128-143.
Gliedman, J.B., Smith J.F. and Brown D.T. Morphogenesis of Sindbis virus in cultured Aedes albopictus cells. J Virol. 1975; 16(4): 913-926
Grimley, P.M., Berezesky, I.K., and Friedman, R.M. Cytoplasmic structures associated with an arbovirus infection: loci of viral ribonucleic acid synthesis. J. Virol. 1968; 2: 1326-1338.
Grimley, P.M., Levin, J.G., Berezesky, I.K., and Friedman, R.M. Specifi c membranous structures associated with the replication of group A arboviruses. J. Virol. 1972; 10: 492-503
Johnston R.E., Peters C. Alphaviruses associated primarily with fever and polyarthritis. In: Fields BN, Knipe DM, Howley PM, editors. Fields Virology, 3rd edition. New York: Raven Press. 1996, pp. 843–898.
Kääriäinen, L., Takkinen, K., Keränen, S., and Söderlund, H. Replication of the genome of alphaviruses. J. Cell Sci. Suppl. 1987; 7: 231-250.
Lahariya C., and Pradhan S.K. Emergence of chikungunya virus in the Indian subcontinent after 32 years: A review. J Vector Borne Dis. 2004; 43(4): 151-60
Pialoux G, Gauzere B.A., Jaureguiberry S, Strobel M. Chikungunya, an epidemic arbovirosis. Lancet Infect Dis. 2007; 7:319-327
Powers A.M. and Logue C.H. Changing patterns of chikungunya virus: re-emergence of a zoonotic arbovirus. J Gen Virol 2007; 88: 2363-2377
Sourisseau M., Schilte C., Casartelli N. et al. Characterization of Reemerging Chikungunya Virus. Plos Pathogens. 2007; 3(6) e89: 0804-0817



















![[Challenge:Future] Mission NO-Chikungunya](https://img.pdfslide.us/doc/110x75/55804b82d8b42ae32c8b4e06/challengefuture-mission-no-chikungunya-55848be297559.jpg)









