Embed Size (px)
Citation preview

Camp. Biochem. Physiol. Vol. 97A, No. 2, pp. 153-158, 1990 0300-9629/90 $3.00 + 0.00 Printed in Great Britain 0 1990 Pergamon Press plc
COMPARATIVE STUDY OF NUTRITIONAL REQUIREMENTS IN TWO AXENICALLY
REARED ARTEMIA STRAINS
ARANTXA HERNANDOBBNA
Laboratoire du Museum National d’Histoire Naturelle, CERS 64200 Biarritz, France. Telephone: 240259
(Received 16 January 1990)
Abstract-l. A new artificial nutritive medium developed for the axenic culture of Artemiu was used to compare the dietary RNA requirement of the American bisexual strain from Utah with that of the Spanish parthenogenetic strain from La Mata and to study the effect of an adaptation to low salinity on the dietary RNA requirement.
2. The effects of dietary RNA on Artemiu development are strain- and salinity-dependent. 3. Results raise unsolved questions regarding the developmental strategy of Artemiu and the problem
of the evolutionary divergence in Artemiu sibling species.
INTRODUCTION
A new simplified nutritive medium has been devel- oped to rear the Artemiu franciscana strain from Utah under axenic culture conditions. The medium includes Torula yeast RNA to satisfy the essential requirement of this crustacea for a dietary source of purine (Hernandorena, 1990a). This unusual require- ment arises as a consequence of an incapacity of Artemia from Utah to synthesize the purine ring de nova (Clegg et al., 1967; Warner and McClean, 1968). According to Rotllan et al. (1987) the apparent inability of Artemiu for de novo purine biosynthesis could be related to a high salinity effect, as the operation of a complete classical pathway of de novo purine biosynthesis was demonstrated in the Artemia nauplii from the San Francisco Bay strain adapted to low salinity (0.1 M NaCl). The dietary purine require- ment of Artemiu is of special interest because Utah larvae reared in purine-deficient nutritive media de- velop into abnormal adults with supernumerary gen- ital appendages on otherwise apodous abdominal segments (Hernandorena, 1970, 1977, 1979, 1987a). The new artificial nutritive medium was used to compare the dietary RNA requirement of the American bisexual strain from Utah to that of the Spanish parthenogenetic strain from La Mata and to study the effect of an adaptation to low salinity on the dietary RNA requirement.
MATERIALS AND METHODS
La Mata cysts contrary to Utah cysts are not commer- cially available. They were kindly supplied by F. Amat (Instituto de Acuicultura, Planta Pilot0 de Torre la Sal, Ribera de Cabanes, Castellon, Spain).
The techniques to produce, handle and rear Artemia under axenic culture conditions, and to prepare the nutritive medium, have been previously detailed (Hernandorena, 199Oa). La Mata nauplii take longer to hatch than Utah nauplii. Newly hatched nauplii obtained 24 hr after cyst hydration for the Utah strain and 32 hr after cyst hydration for the La Mata strain, were transferred in groups of five
into each culture tube containing 10ml of the autoclaved nutritive medium. The composition of medium C is detailed in Table 1. During these experiments, the RNA concen- tration of medium C was varied from 0 to 400 mg/lOO ml of media, and the sea-water concentration from 8Oml (28%0 salinity) to 20 ml (7% salinity) per 100 ml of media. RNA- deficient media were supplemented with inosinic acid (IMP) or with cytidylic acid (CMP) using 50mg nucleotide per 1OOml of media. The number of nauplii reared for each experimental condition is indicated in Tables 3 and 4. Their growth rate was measured by the numerical growth index defined by Provasoli and d’Agostino (1969). The morpho- genesis of adult individuals was checked by a microscopical examination of animals anesthesized with chloroform. A few of them were measured.
RESULTS
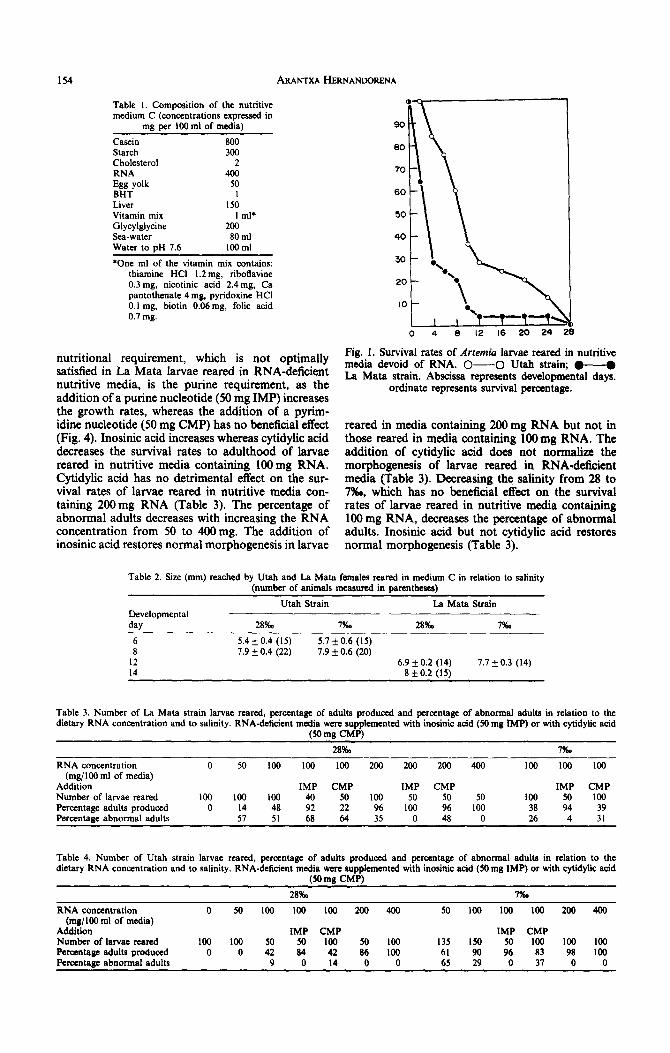
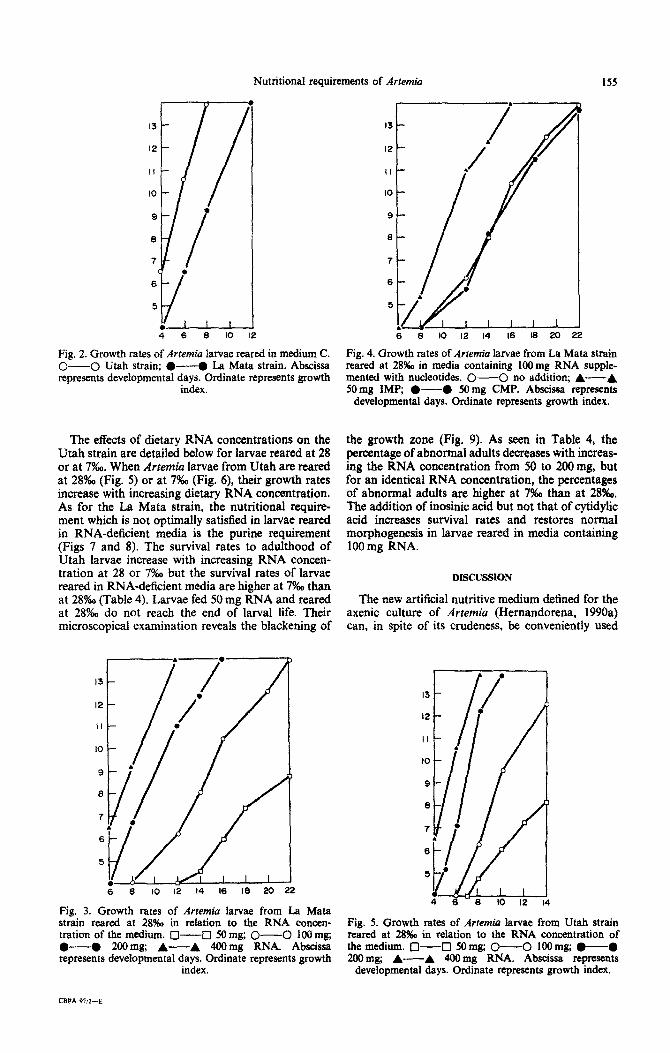
When Artemia larvae are reared in medium C devoid of RNA, they do not grow beyond the stage reached by non-fed larvae (growth index 3), but Utah larvae survive better than La Mata larvae (Fig. 1). When Artemia larvae are reared in medium C con- taining 400 mg RNA (data on the concentration of nutrients are given per 100 ml of media), Utah larvae grow faster than La Mata larvae (Fig. 2). As seen in Table 2, females of about the same size are obtained in 8 days with Utah larvae (7.9 + 0.4 mm) and in 14 days with La Mata larvae (8 f 0.2 mm). Decreas- ing the external salinity from 28 to 7!%0, which has no effect on the Utah strain larvae, increases the growth rates of the La Mata larvae. Artemia larvae from Utah and from La Mata reared in media contain- ing 400 mg RNA develop into normal adults at 28 and 7%0.
The effects of dietary RNA concentrations on Artemiu growth survival and morphogenesis are de- tailed below, starting with the La Mata strain larvae reared at 28X. The growth (Fig. 3) and survival rates (Table 3) of La Mata larvae increase with increasing the dietary RNA concentration. La Mata larvae do not grow faster when the RNA concentration is increased up to 500 mg (result not shown). The
153
ARANTXA HERNANWRENA 154
Table I. Composition of the nutritive medium C (concentrations expressed in
mg per 100 ml of media)
Casein 800 Starch 300 Cholesterol 2 RNA 400 Egg yolk 50 BHT I Liver 150 Vitamin mix 1 ml* Glycylglycine 200 Sea-water 80 ml Water to pH 7.6 lOOmI
*One ml of the vitamin mix contains: thiamine HCI 1.2 mg, riboflavine 0.3 mg, nicotinic acid 2.4mg, Ca pantothenate 4 mg, pyridoxine HCI 0.1 mg, biotin 0.06 mg, folic acid 0.7 mg.
nutritional requirement, which is not optimally satisfied in La Mata larvae reared in RNA-deficient nutritive media, is the purine requirement, as the addition of a purine nucleotide (50 mg IMP) increases the growth rates, whereas the addition of a pyrim- idine nucleotide (50 mg CMP) has no beneficial effect (Fig. 4). Inosinic acid increases whereas cytidylic acid decreases the survival rates to adulthood of larvae reared in nutritive media containing 1OOmg RNA. Cytidylic acid has no detrimental effect on the sur- vival rates of larvae reared in nutritive media con- taining 200mg RNA (Table 3). The percentage of abnormal adults decreases with increasing the RNA concentration from 50 to 400 mg. The addition of inosinic acid restores normal morphogenesis in larvae
I I T-T t T --\ o 4 6 12 16 20 24 : 3
Fig. 1. Survival rates of Arremia larvae reared in nutritive media devoid of RNA. 0-O Utah strain; l -@ La Mata strain. Abscissa represents developmental days.
ordinate represents survival percentage.
reared in media containing 200 mg RNA but not in those reared in media containing 1OOmg RNA. The addition of cytidylic acid does not normalize the morphogenesis of larvae reared in RNA-deficient media (Table 3). Decreasing the salinity from 28 to 7960, which has no beneficial effect on the survival rates of larvae reared in nutritive media containing 100 mg RNA, decreases the percentage of abnormal adults. Inosinic acid but not cytidylic acid restores normal morphogenesis (Table 3).
Table 2. Size (mm) reached by Utah and La Mata females reared in medium C in relation to salinity (number of animals measured in oarenthescs)
Developmental day
Utah Strain La Mata Strain
28% 7% 28% 7%0
6 5.4 f 0.4 (15) 5.7 + 0.6 (IS) 8 7.9 + 0.4 (22) 7.9 f 0.6 (20)
12 6.9 f 0.2 (14) 7.7 & 0.3 (14) 14 8 k 0.2 (15)
Table 3. Number of La Mata strain larvae reared, percentage of adults produced and percentage of abnormal adults in relation to the dietary RNA concentration and to salinity. RNAdeficient media were supplemented with inosinic acid (50 mg IMP) or with cytidylic acid
(50 ma CMP)
28G 7%
RNA concentration 0 50 100 100 100 200 200 200 400 100 100 100 (mg/lOO ml of media)
Addition Number of larvae reared Percentage adults produced Percentaae abnormal adults
IMP CMP IMP CMP IMP CMP 100 100 100 40 50 100 50 50 50 100 50 100
0 14 48 92 22 96 100 96 100 38 94 39 57 51 68 64 35 0 48 0 26 4 31
Table 4. Number of Utah strain larvae reared, percentage of adults produced and percentage of abnormal adults in relation to the dietary RNA concentration and to salinity. RNA-deficient media were supplemented with inosinic acid (50 mg IMP) or with cytidylic acid
(50 me CMP)
28% 7540
RNA concentration (mg/lOO ml of media)
Addition Number of larvae mated Percentage adults produced Percentage abnormal adults
0 50 100 100 100 200 400 50 100 100 100 200 400
IMP CMP IMP CMP 100 100 50 50 100 50 100 135 I50 so 100 100 100
0 0 42 84 42 86 100 61 90 96 83 98 100 9 0 14 0 0 65 29 0 37 0 0
Nutritional requirements of Artemia 155
Fig. 2. Growth rates of Artemia larvae reared in medium C. O- 0 Utah strain; 0-O La Mata strain. Abscissa represents developmental days. Ordinate represents growth
index.
The effects of dietary RNA concentrations on the Utah strain are detailed below for larvae reared at 28 or at 7%. When Artemiu larvae from Utah are reared at 28% (Fig. 5) or at 7% {Fig. 6), their growth rates increase with increasing dietary RNA concentration. As for the La Mata strain, the nu~tional require- ment which is not optimally satisfied in larvae reared in RNA-deficient media is the purine requirement (Figs 7 and 8). The survival rates to adulthood of Utah larvae increase with increasing RNA concen- tration at 28 or 7% but the survival rates of larvae reared in RNA-deficient media are higher at 7%0 than at 28% (Table 4). Larvae fed 50 mg RNA and reared at 28% do not reach the end of larval life. Their micros~pi~l examination reveals the blackening of
Fig. 3. Growth rates of Artemia larvae from La Mata strain reared at 28%~~ in relation to the RNA concen- tration of the medium. U---L3 50 mg; 0-O 100 mg; +--O 200mg; A-A 400mg RNA. Abscissa represents deveIopmen~1 days. Ordinate represents growth
index.
6 6 IO 12 14 I6 I6 20 22
Fig. 4. Growth rates of Artemia larvae from La Mata strain reared at 28% in media containing 1OOmg RNA supple- mented with nucleotides. O-0 no addition; &---A 5Omg IMP; a--- 0 50mg CMP. Abscissa represents
developmental days. Ordinate represents growth index.
the growth zone (Fig. 9). As seen in Tabb 4, the percentage of abnormal adults decreases with increas- ing the RNA concentration from 50 to 200 mg, but for an identical RNA concentration, the percentages of abnormal adults are higher at 7%0 than at 28%. The addition of inosinic acid but not that of cytidylic acid increases survival rates and restores normal mo~hogen~is in larvae reared in media containing 1OOmg RNA.
DISCUSSION
The new artificial nutritive medium defined for the axenic culture of Artemia (Hemandorena, 1990a) can, in spite of its crudeness, be conveniently used
7
6
Fig. 5. Growth rates of Artemiu larvae from Utah strain reared at 2% in relation to the RNA concentration of the medium. o-0 50 mg; 0-O 100 mg; l -# 200mg; A-- A 4OOmg RNA. Abscissa represents
developmental days. Ordinate represents growth index.
CBPA 9712-E

156 ARANTXAHERNANDORENA
Fig. 6. Growth rates of Artemiu larvae from Utah strain reared at 7% in relation to the RNA concentration of the medium. q --_O 50mg; 0-O 1OOmg; 0-O 200mg; A-A 400mg RNA. Abscissa rep- resents developmental days. Ordinate represents growth
index.
to compare the nutritional requirements of differ- ent Artemiu strains. It is more efficient than the original medium “100” developed by Provasoli and d’Agostino (1969) for the axenic culture of the Utah strain which does not support the growth of partho- genetic strains (Provasoli and Pintner, 1980). The comparative study of the dietary RNA requirement is of special interest because larvae reared in RNA- deficient media develop into abnormal adults. Results raise unsolved questions regarding the developmental strategy of Artemia (Hernandorena, 1990b) and regarding the evolutionary divergence of Artemia sibling species.
The developmental strategy of Artemia differs from that of Drosophila in that several inductive events involved in the formation of abdominal
Fig. 7. Growth rates of Artemia larvae from Utah reared at 28% in media containing 100 mg RNA supplemented with nucleotides. 0-O No addition; A----A 50mg IMP, 0-a 50 mg CMP. Abscissa represents developmental
days. Ordinate represents growth index.
Fig. 8. Growth rates of Artemiu larvae from Utah reared at 7% in media containing 1OOmg RNA supplemented with nucleotides. 0-O No addition; A-_-A 50mg IMP; l -a 50 mg CMP. Abscissa represents developmental
days. Ordinate represents growth index.
segments and in the specification of genital and abdominal segment identities take place during post- embryonic development; they can be upset by a dietary RNA deficiency.
The developmental anomalies induced by a dietary RNA deficiency are strain- and salinity-dependent. In larvae reared at 28%, they correspond to homeotic- like transformations: inductive events involved in the normal establishment of the identity of the first abdominal segment can be upset in the Utah and the La Mata strains; those involved in the normal estab- lishment of the genital segment identity can be upset in the La Mata strain. At 7%, inductive events involved in the formation of abdominal segments can be upset in the Utah strain. Artemiu is not one single species but a complex of sibling species. Artemiu
Fig. 9. Blackening of the growth zone in a Utah larvae reared at 28% in media containing 50 mg RNA.

Nutritional requirements of Arfemiu 157
franciscana (Utah) and Artemia parthenogenetica (La Mata) are no longer sibling species when reared in RNA-deficient media.
We have information regarding the amount of genetic variation which exists between Artemia populations, as revealed by electrophoretic studies and a large selection of morphological characters with which we can distinguish them, but we know little about the events which underlie speciation (Abreu-Grobois, 1987). As underlined by Barigozzi (1980) the morphological characters considered to distinguish Artemia species are more-or-less strongly influenced by environment and not very reliable to describe systematic entities. The comparative study of the dietary RNA requirement initiated in the present work with a Spanish and an American strain, may contribute to a better understanding of the unsolved questions raised by the evolutionary divergence in Artemia sibling species from the Old and the New World. In order to develop normally, Artemia larvae from both strains must be reared in RNA-rich nutri- tive media, but the RNA concentration required for normal development is higher in larvae from La Mata (> 200 mg RNA) than in larvae from Utah (> 100 mg RNA). The La Mata and Utah strains are both diploid. It will be interesting to determine the RNA concentration required for normal development in polyploid Artemia strains.
Results show that the essentiality of the dietary RNA requirement is not restricted to the Utah strain larvae, nor to larvae reared at 28%. It can be extended to the La Mata strain larvae which do not grow in media devoid of RNA. We already knew that Utah larvae reared in medium “100” devoid of a dietary source of purine do not grow but can survive for long periods (10% survival at day 24) (Hernandorena, 1974). Identical results are obtained with Utah larvae reared in the new medium devoid of RNA (12% survival at day 24, Fig. 1). Differences in the survival rates of Utah and La Mata larvae reared in media devoid of RNA could be explained by differences in their provision of maternally sup- plied purines, that is, by maternal purine nutri- tion. Artemia nauplii are equipped with maternally supplied metabolites including a provision of diguanosine-5’-tetraphosphate which remains con- stant throughout pre-emergence development and is extensively used after hatching (Warner and Finamore, 1967). Differences in the diguanosine polyphosphate levels in Artemia cysts from different strains are documented (Sonnenfeld and Warner, 1989) which could explain that without dietary RNA, survival rates are strain-dependent and that in RNA-deficient media, the induced developmental anomalies are strain-dependent (Hernandorena, l990b). In both strains, the growth and survival rates increase with increasing the dietary RNA concentration.
The essentiality of the dietary RNA requirement can be extended to larvae reared at low salinity. If the inability to synthesize the purine ring de nova was a high salinity effect, as suggested by Rotllan et al. (1987) an adaptation to 7% should increase growth and survival rates and decrease the percentage of abnormal adults. In fact, the dietary RNA deficiency is experienced differently at low salinity since the
developmental anomalies induced in the Utah strain are salinity-dependent, but it is detrimental at 28 and 7%0. We already know that the effect of an adaptation to high salinity on the form of Artemia body is strain-dependent (Gilchrist, 1960) and is purine- mediated (Hernandorena, 1975). Biochemical and nutritional data suggest that it is the ability of larvae to synthesize thymidylate and not that to synthesize the purine ring de novo which is decreased at high salinity. Utah larvae reared at 28% do not reach adulthood when fed 50mg RNA. They do not sur- vive to the blackening of the growth zone. Such a blackening of the growth zone has already been described for Artemia larvae from Utah treated with 5 FUdR, an inhibitor of thymidylate synthetase (Hernandorena, 1972). Utah larvae reared at 28X& and fed 50 mg RNA do not survive because they are incapable of synthesising thymidylate. We already know that when Utah larvae are reared in medium “100” under nutritional conditions supposed to interfere with purine nucleotide interconversion, their ability to reduce folic acid for thymidylate and hence DNA biosynthesis becomes circumstantial (Hemandorena, 1987b). It is reduced for larvae fed adenine or hypoxanthine but not for those fed guanine-deficient media. Higher survival rates and higher percentages of abnormal adults are obtained with larvae fed hypoxanthine or guanine than with larvae fed adenine. For comparable survival rates, higher percentages of abnormal adults are obtained with larvae fed guanine than with larvae fed hypox- anthine (Hemandorena, 1987a). The most efficient inducers of super-numerary abdominal appendages are the guanine compounds (guanine, guanosine and guanylic acid) found to be the least efficient at meeting the growth requirements (Hernandorena, 1985).
Results show that the ability of larvae to develop into abnormal adults and their ability to synthesize thymidylate depend on the quality and the quantity of dietary purines. According to Rotllan et al. (1987) the production of adenine nucleotides is reduced in Artemia nauplii adapted to low salinity (0.1 M NaCl). If the metabolic adaptation to 0.1 M NaCl interferes with purine nucleotide interconversion, the higher survival rates and higher percentage of abnormal adults obtained with Utah larvae fed RNA-deficient media, when adapted to 7%0, may result from a reduced adenine nucleotide production, resulting in an increased ability to synthesize thymidylate. As already underlined (Hemandorena, 1990b), the sig nificance of the role of purine nutrition on Artemia development remains unclear. Torula yeast RNA constitutes a convenient nutrient, providing Artemia larvae with a mixture of purine nucleotides and a mixture of pyrimidine nucleotides. The nutritional requirement which is not optimally satisfied in Artemia larvae reared in RNA-deficient media is the purine requirement since normal development is re- stored in adults developing from larvae reared in RNA-deficient media supplemented with a purine nucleotide. Medium C with Torula yeast RNA included can be used to compare the nutritional requirements of different strains but further exper- iments on the effect of salinity should be performed with medium C, including a chemically defined

158 ARANTU Hw
mixture of purine and pyrimidine compounds in place of Torula yeast RNA.
Acknowledgemenfs-Part of this work was financially supported by the So&e Nationale Elf Aquitaine.
REFERENCES
Abreu-Grobois F. R. (1987) A review of the genetics of Artemia. In Artemia Research and its Applications, Vol. 1, Morphology, Genetics, Strain characterization, Toxicology (Edited by Sorgeloos P., Bengtson D. A., Deeleir W. and Jaspers E.), pp. 61-99. Universa Press, Wetteren, Belgium.
Barigozzi C. (1980) Genus Artemia: problems of systemat- its. In The Brine Shrimp Artemia, Vol. 1, Morphology, Genetics, Radiobiology, Toxicology (Edited by Persoone G.. Soraeloos P., Roels 0. and Jaspers E.), pp. 147-153. Universa Press, Wetteren, BelgiumY
Clenz J. S.. Warner A. H. and Finamore F. J. (1967) Gdence ‘for the function of Pi, P’diguanosme-5’: tetraphosphate in the development of Artemia salina. J. biol Chem. 242, 1938-1967.
Gilchrist B. (1960) Growth and form of the brine shrimp Artemia salina (L.). Proc. 2001. Sot. Land. 134,221-235.
Hemandorena A. (1970) Obtention de morphogen&ses appendiculaires abortives et sumum6raires chez Artemia salina (L.) (Crustad Branchiopode) par carences alimen- taires de base pyrimldique et de nuclCotide purique. C.R. Acad. Sci. Paris 271, 14061409.
Hernandorena A. (1972) Signification morphogenetique du besoin alimentaire en acides nucl6iques chez Artemia salina. I. Besoin en thymidinc. Arch. 2001. exp. gen. 113, 425432.
Hemandorena A. (1974) Besoin alimentaire en acide adenylique, croissance et morphogenese chez Artemia salina (L.) (Crustac& Branchiopode). Ann. Nutr. Alim. 28, 65-82.
Hemandorena A. (1975) Metabolic significance in nucleic acid metabolism and protein synthesis of dietary AMP requirement in Artemia salina (L.). Biol. Bull. 148, 416-428.
Hernandorena A. (1977) Action antagoniste de m&o- tides purique et pyrimidique alimentaires sur la morpho- get&e appendiculaire d’drtemia salina (L.) (CrustacC Branchiopode). CR. Acad. Sci. Paris 284, 1337-1339.
Hemandorena A. (1979) Relationships between purine and pyrimidine dietary tequirements and Artemia salina morphogenesis. Comp. Biochem. Physiol. 62B, 7-12.
Hemandorena A. (1985) Purine-mediated phosphate inilu- ence on Artemia morphogenesis. Reprod. Nutr. D&e/. 2!3, 883893.
Hemandorena A. (1987a) Circumstances atIect.ing the efficiency of dietary purines in Artemia. Comp. Biochem. Physiol. 86B, 397-4Q3.
Hemandorena A. (1987b) An increase dietary tryptophan requirement induced by interference with purine nucleo- tide interconversion in Artemia. In Artemia Research and its Application, Vol. 2, Physiology, Biochemistry, Molecu- lar Biolonv Edited bv Decleir W.. Moens L.. Sleaers H., Jaspers ET and Sorgeloos P.). U&versa Press, Witteren, Belgium. 562 pp.
Hemandorena A. (199Oa) Development of a new simplified nutritive medium for the axenic culture of Artemia. (submitted).
Hemandorena A. (199Ob) Purine nutrition and Artemia development (submitted).
Provasoli L. and d’Agostino A. (1969) Development of artificial media for Artemia salina. Biol. BUN. 136, 434453.
Provasoli L. and Pintner I. J. (1980) Biphasic particulate media for the parthenogenetic Arfemia of S&e. In The Rrine Shrimp Artemia, Vol. 2, Physiology, Biochemistry, Molecular Biology (Edited by Persoone G., Sorgeloos P., Roels 0. and Jaspers E.), pp. 231-238. Universa Press, Wetteren, Belgium.
Rotllan P., Liras A. and Llorente P. (1987) Biosynthesis of nucleotides from exogenous purines in Artemia nauplii. In Artemia Research and its Applications, Vol. 2, Physiology, Biochemistry, Molecular Biology (Edited by Decleir W., Moens L., Slegers H., Jaspers E. and Sorgeloos P.), pp. 275-281. Universa Press, Wetteren, Belgium.
Sonnenfeld M. J. and Warner A. H. (1989) Comparison of diguanosine polyphosphate levels in various populations of Artemia cysts. In Cell and Molecular Biology of Artemia deoelopment (Edited by Warner A. H., MacRae T. H. and Bagshaw J. C.), p. 425. Plenum Press, New York.
Warner A. H. and Finamore F. J. (1967) Nucleotide metab- olism during brine shrimp embryogenesis. J. biol. Chem. 242, 19334937.
Warner A. H. and McClean D. IL (1968) Studies on the biosynthesis and role of diguanosine tetraphosphate during growth and development of Arfemia salina. De01 Biol. 18, 278-293.







![The occurrence of the brine shrimp, Artemia franciscana ... · America [25], other Artemia species include: Artemia persimilis of South America [26], Artemia salina in the Mediterranean](https://img.pdfslide.us/doc/110x75/5c4b6a7093f3c3117d72c1b0/the-occurrence-of-the-brine-shrimp-artemia-franciscana-america-25-other.jpg)





















