Embed Size (px)
Citation preview

Original Paper
Brain Behav Evol 2002;60:65–79
Comparative Anatomy of the ParatympanicOrgan (Vitali Organ) in the Middle Ear of Birdsand Non-Avian Vertebrates:Focus on Alligators, Parakeets and Armadillos
Jason A. Neesera Christopher S. von Barthelda–c
aDepartment of Physiology and Cell Biology, University of Nevada School of Medicine, Reno, Nev.,bHearing Development Laboratories, University of Washington School of Medicine, Seattle, Wash., USA;cDepartment of Neuroanatomy, University of Göttingen School of Medicine, Göttingen, Germany
Received: March 4, 2002Returned for revision: April 19, 2002Accepted after revision: June 13, 2002
Dr. Christopher S. von BartheldDepartment of Physiology and Cell BiologyUniversity of Nevada School of MedicineReno, NV 89557 (USA)Tel. +1 775 784 6022 or 4635, Fax +1 775 784 6903, E-Mail [email protected]
ABCFax + 41 61 306 12 34E-Mail [email protected]
© 2002 S. Karger AG, Basel
Accessible online at:www.karger.com/journals/bbe
Key WordsMiddle ear W Reptile W Bird W Alligator W Parakeet W
Armadillo W Comparative anatomy W Paratympanicorgan W Acoustic stapedius reflex W Elastic ligament
AbstractThe paratympanic organ (PTO) in the middle ear hasbeen described in numerous bird species, but little isknown about the distribution of this presumed lateralline remnant in other vertebrate classes. Here we pro-vide evidence for a PTO in juvenile alligators, and makethe first detailed description of its location and relation toligaments in the reptilian middle ear. The alligator PTOmeasures about 450 Ìm in diameter. The alligator PTOcontains hair cells whose cilia extend into a mucous sub-stance within the lumen. The PTO connects though aligament to the ear drum, suggesting that pressure ontothe tympanic membrane might induce fluid movementin the PTO. Labeling of innervating nerve fibers with thefluorescent dye, DiI, indicates that the alligator PTO isconnected with the vestibular brainstem. Because allbird species examined possess a PTO except for owlsand possibly parakeets, we verified the absence of a PTO
in parakeets by examination of serial sections combinedwith GABA immunolabeling for potential hair cells. Birdspecies with significant upper beak movement lack aPTO, suggesting that PTO function is incompatible withupper beak movement. We also examined the middleear of an armadillo, a mammal that has a very basal posi-tion within the eutherian phylogenetic tree. A small vesi-cle with ciliated cells was found, but did not label with ahair-cell specific marker, antibodies to myosin VIIa, andthus is not likely to represent a true PTO. Our evidencefor a PTO in a non-avian species, the alligator, togetherwith previous reports suggesting the presence of a PTOin some mammals, indicates that ancestral stem am-niotes possessed a PTO, and that the PTO was not a denovo invention of birds.
Copyright © 2002 S. Karger AG, Basel
Introduction
The paratympanic organ (PTO) is a small sensoryorgan in the middle ear of birds. More than ninety yearsafter its discovery [Vitali, 1911], the function of this senseorgan has still not been proven. The presence of a PTO

66 Brain Behav Evol 2002;60:65–79 Neeser/von Bartheld
has been demonstrated in most bird species examined [forreview, see Simonetta, 1953; von Bartheld, 1990]. Thedistribution of a PTO among non-avian species is uncer-tain. Work by Vitali [1924] and Simonetta [1953] indi-cated that a PTO-like vesicle may be present in someembryonic reptiles and mammals, and suggested that itmight also persist in a small number of juvenile and adultnon-avian species. However, the morphology was notconvincing, and the authors did not attempt to demon-strate the presence of hair cells or their innervation in theputative PTO. If a PTO can be found among non-avianvertebrates, it would indicate that this organ or its homo-log was likely present in ancestral vertebrates that gaverise to amniotes.
The PTO is thought to be a homolog of the spiracularsense organ (SSO) in fishes [Vitali, 1925; Ranzi, 1926;Barry and Boord, 1984; Barry and Bennett, 1989; vonBartheld, 1990; von Bartheld and Rubel, 1990]. All fishesexamined (including lungfishes, elasmobranchs, holo-cephali, chondrosteans, ginglymodi and cladistia), withthe exception of the teleosts, possess an SSO [reviewed bySimonetta, 1953]. In ancestral vertebrates, the spiraculumwas involved in breathing, and the hyomandibula orstapes appears to have assisted such a breathing function[Clack, 1989, 1990, 1994]. The PTO develops indepen-dently of the otic placode or otocyst [Yntema, 1944; vonBartheld et al., 1992]. Both sense organs, the SSO and thePTO, appear to be intimately related to the hyomandibulaand the columella, respectively. Thus, it is likely that withchanging functions of the hyomandibula/columella frombreathing assistance to hearing assistance, the associatedsense organ underwent its first change in function. Inbirds, the PTO appears to have undergone a second trans-formation into an organ with the possible function ofdetection of barometric pressure [Vitali, 1921, 1941; Ran-zi, 1926; Federici, 1927; Griffin, 1969; Delius and Em-merton, 1978; Jørgensen, 1984; Giannessi and Pera,1986; von Bartheld, 1994], and it is possibly used in addi-tion as an ‘altimeter’ in some birds [Griffin, 1969; Kreith-en and Keeton, 1974; von Bartheld, 1990, 1994].
In this study, we examined whether a PTO is present injuvenile alligators. We provide the first detailed descrip-tion of a PTO in a non-avian species [Neeser and von Bar-theld, 2001]. We also report on the relationship of the alli-gator PTO with ligaments in the middle ear, and on itsinnervation from the brainstem. We further examinedcorrelations of morphological and physiological parame-ters with the presence or absence of PTOs in bird species,which could give an indication of its function. We reportthe remarkable correlation of a fixed upper beak with the
presence of a PTO, and an additional correlation with theabsence of an acoustic stapedius reflex. Such data suggestthe presence of two principal mechanisms for control ofthe position and tension of the tympanic membraneamong birds, which correlate with the presence or absenceof a PTO. Finally, we examined the identity of cell typesin a putative PTO of the armadillo, a mammal thatemerged early at the Cretaceous/Tertiary boundary [Ar-nason et al., 1997; Delsuc et al., 2001], and we concludethat ciliated cells in this putative PTO lack antigenicity tomyosin VIIa, a specific hair cell marker in mammals.Such data indicate that reports of PTOs in mammals orreptiles should be regarded with caution when they aremerely based on the presence of an epithelial vesiclealone.
Materials and Methods
MaterialsTwo specimens of 2-year old alligators (Alligator mississippiensis)
were obtained from existing collections in New York (AmericanMuseum of Natural History, permit # CRP90–363, New York StateDivision of Fish and Wildlife). Two adult parakeet specimens (Me-lopsittacus undulatus) were obtained from existing collections at theUniversity of Göttingen (Germany). One adult armadillo specimen(Dasypus novemcinctus) was obtained from an existing collection atthe Mote Marine Laboratory, Sarasota, Florida. The original re-search reported herein was performed under guidelines establishedby the U.S. Department of Health, Education and Welfare [DHEWPub. No. 78-23, 1978] and all procedures were approved by localanimal care committees.
HistologyThe alligators were anesthetized with a mixture of ketamine/xyla-
zine and perfused intracardially with saline followed by half strengthKarnovsky’s fixative. The heads were then decalcified in severalchanges of EDTA (10%) and stored for 10 years at room temperature[von Bartheld et al., 1990]. Two parakeets were anesthetized withNembutal and perfused through the heart with either 4% paraformal-dehyde (PFA) or 2% PFA and 2% glutaraldehyde. Their heads weredecalcified in EDTA for 2 months. The armadillo was fixed in for-malin and decalcified and stored in 10% EDTA for 10 years. Thewhole heads of parakeet or dissected middle ears (alligator and arma-dillo) were cryoprotected in 30% sucrose, embedded in OCT com-pound (Tissue-Tek®), frozen on crushed dry ice, and frozen sectionsof 25–40 Ìm were cut in the transverse plane (alligator, parakeet) orin the horizontal plane (armadillo) on a Leica cryostat. Sections werecollected on gelatin-coated glass slides, dried, and stained with thio-nin. Slides were dehydrated, coverslipped with DPX and observedon a Nikon Eclipse E600 photomicroscope. The approximate num-ber of hair cells in the PTO was estimated by focusing through serialsections of the tissue at 100! magnification. Each hair cell bundlethat was identified was scored for quantification; no correction fordouble-counting was used, because the cilial bundle (5–7 Ìm length)was small relative to the section thickness (25–40 Ìm). Camera-luci-da drawings of serial sections were prepared to reconstruct how the

Paratympanic Organ in Birds and Reptiles Brain Behav Evol 2002;60:65–79 67
ligament adjacent to the PTO connects the PTO with the tympanicmembrane.
DiI-TracingOne side of one alligator head was used for DiI-tracing. The head
was washed 2 days in phosphate-buffered saline, pH 7.4. Crystals ofDiI (Molecular Probes) were placed in the brainstem at the entry ofthe vestibular and cochlear cranial nerves, and the tissue was storedfor 3 months in 2–4% PFA fixative at room temperature. The middleear including the peripheral ganglia and some of the brainstem tissuewas dissected out and placed in 30% sucrose containing 0.1% EDTAto minimize DiI diffusion [Hofmann and Bleckmann, 1999]. The tis-sue was frozen with dry ice in OCT embedding compound and sec-tioned in a cryostat at 30–40 Ìm. The tissue was oriented so that theknife blade cut through the peripheral tissue first, thus minimizingthe possibility of contamination by sectioning through the vicinity ofthe injection site. Serial sections were collected on gelatine-coatedglass slides and observed on a Nikon Eclipse microscope with fluores-cence optics. Some sections were coverslipped with Gelmount™(Biomeda), and images were documented on TMAX 3200 film.
ImmunocytochemistryOne of the decalcified, glutaraldehyde-fixed parakeet heads, one
decalcified alligator middle ear, and one decalcified armadillo mid-dle ear were sectioned for immunohistochemistry. They were cryo-protected as described above, embedded in OCT compound and fro-zen on crushed dry ice. Cryosections of 25–40 Ìm were collected ongelatine-coated glass slides and dried for 30–90 min. Each section tobe immunolabeled was circled on the glass slide with either a greasepen or a rim of nail polish. Sections were incubated in phosphatebuffer, followed by 5% normal goat serum. The primary antibody forGABA was a polyclonal rabbit antibody (against GABA coupled toglutaraldehyde, Incstar, now Diasorin); the primary antibody formyosin VIIa was a kind gift from Tama Hasson [Hasson et al., 1995].Standard immunocytochemical techniques were used with the pri-mary GABA antibody at 1:3,000 and the primary myosin antibody at10 Ìg/ml overnight at 4°C. After washes at room temperature, sec-ondary biotinylated goat-anti-rabbit antibody from Vector Labs(1:500 dilution) was applied for 1 h, followed by washes and strept-avidin-HRP from Vector Labs (1:500 dilution, 1 h), followed bywashes in phosphate buffer and then 50 mM Tris buffer (pH 7.6).Sections were reacted in diaminobenzidine (0.02%) and 0.015%H2O2 for 5–10 min, dehydrated in graded ethanols and coverslippedwith DPX. As a positive control for the myosin VIIa antibody, innerear hair cells of the armadillo were used. As a negative control, theprimary antibodies were omitted, resulting in minimal backgroundstaining. Sections were examined on a Nikon Eclipse microscope fit-ted with Nomarski optics and pictures taken using TechPan film. Allimages are presented without digital manipulations.
Results
Location and Morphology of a PTO in JuvenileAlligatorsThe dorsomedial margin of the alligator middle ear
contains an elongated vesicle of about 200 Ìm in length inthe rostrocaudal axis and a diameter about 450 Ìm dor-
soventrally (fig. 1A, B). This vesicle is located at a dis-tance of about 300 Ìm from the tympanic membranewhere the ear drum attaches to the dorsomedial cavity ofthe middle ear (fig. 1A). The lumen of the vesicle is in partfilled with a dark-staining amorphous mass (fig. 1B), pre-sumably mucus, as demonstrated in the avian PTO lumen[Jørgensen, 1984]. The medial side of the lumen containsan epithelium of tall cells, and the lateral wall contains anepithelium of cells with a lesser height (fig. 1B, 2A). Thisis the site where a short ligament (about 300 Ìm) attachesto the lateral wall of the PTO (fig. 2). This ligamentextends in a rostral, lateral, and dorsal direction and con-nects to the dorsalmost origin of the tympanic membrane(fig. 1A, 2). At higher magnification, it can be seen thatthe medial wall of the PTO contains a sensory epitheliumwith numerous hair cells (fig. 3A). In several of these haircells, individual stereocilia (5–7 Ìm length) can be recog-nized at higher magnification (fig. 3A, B). These hair cellsare indistinguishable at the light-microscopic (LM) levelfrom those found in the inner ear sensory epithelia in thesame tissue sections (fig. 3C). Based on the frequency ofhair cells in individual serial sections, we estimate that thealligator PTO contains at least 60 hair cells. The images ofthese cells provide strong evidence that the ciliated cells inthe reptilian middle ear are indeed hair cells, comparableto those demonstrated previously at the LM and electron-microscopic level in the avian PTO [Petrash et al., 1983;Jørgensen, 1984; Giannessi and Pera, 1986; Giannessiand Ruffoli, 1996]. Attempts to label these hair cells withGABA antibodies were not successful, due to the extraor-dinarily high background staining in the alligator tissue.The myosin VIIa antibody [Hassan et al., 1995; see below]that labels mammalian hair cells did not label either PTOor inner ear hair cells in the alligator, probably due to spe-cies-specific differences (data not shown).
Innervation of the PTO in Juvenile AlligatorsIn the avian PTO, the sensory epithelium is innervated
by afferent ganglion cells located in the facial ganglionthat project to the facial/vestibular brainstem [von Bar-theld, 1990] as well as efferent neurons in the vicinity ofthe facial motor nucleus [von Bartheld, 1994]. To deter-mine if the hair cells in the alligator PTO are innervated,the lipophilic fluorescent tracer DiI was applied to thevestibular brainstem, and 3 months later, the middle earwas sectioned and analyzed for DiI-labeled fibers and ter-minals. The injection site included the vestibular brain-stem, as evidenced by robust labeling of the vestibularnerve and its peripheral terminations in the end organswithin the inner ear (data not shown). A few aligned punc-

68 Brain Behav Evol 2002;60:65–79 Neeser/von Bartheld
Fig. 1A, B. Location of the paratympanic organ (PTO) in the alligator middle ear. A Cryosection through the regionwhere the tympanic membrane (TM) attaches to the dorsomedial wall of the middle ear. Section through decalcifiedtissue was stained with thionin. The arrow points to the PTO. Other abbreviations: E = external ear canal; M = middleear cavity; L = ligament connecting with the TM. B Higher magnification of the PTO (arrow). Note the tall epitheliumon the medial (left) side of the lumen. The bracket indicates where the ligament (L) attaches to the PTO. Scale bars:200 Ìm (A) and 50 Ìm (B).
tate labeling patterns indicative of a few labeled nervefibers were seen adjacent to the medial wall of the PTO(fig. 3D), and occasional labeled hair cells were seen in themedial epithelium (fig. 3D). The majority of the hair cellbody appeared labeled, not only the basal margin, possi-bly indicating transneuronal label across tight junctions
[Jørgensen, 1984], as reported in other systems [von Bar-theld et al., 1990; Hofmann and Bleckmann, 1999].Although the label was too sparse to trace individualfibers with certainty to their respective ganglia, these datasupport the idea of a presumptive afferent or efferentinnervation of the alligator PTO.

Paratympanic Organ in Birds and Reptiles Brain Behav Evol 2002;60:65–79 69
Fig. 2. Schematic drawing of the spatial relationship of the paratym-panic organ (PTO) in the alligator middle ear and the tympanicmembrane (TM), the columella (COL) and the short ligament (LIG)connecting the PTO with the TM. The external ear canal (E) is indi-cated. This highly schematic drawing was reconstructed from serialsections through the alligator middle ear.
PTOLIG
COL
TM
E
dorsal
lateral
Lack of a PTO in the Parakeet,Melopsittacus undulatusA PTO has been demonstrated in all adult bird species
examined to date (a total of 38 bird species), with theexception of two owl species [Vitali, 1914, 1923] and pos-sibly the parakeet, Melopsittacus undulatus [Oldenstam,1925]. Because owls and parakeets are bird species withthe most significant upper beak movement [Stellbogen,1930] and such movement in the quadratosquamosalarticulation might be incompatible with a functionalPTO, we re-examined the question of the existence of aPTO in the parakeet. We used both normal Nissl-stainedserial histological sections and a series of sections whichwere alternately stained with GABA antibodies. GABA isa sensitive and reliable marker of hair cells in the avianinner ear [Usami et al., 1987], as well as the PTO [vonBartheld, 1990]. A careful analysis of histological andimmunolabeled sections showed no evidence for a PTO in
the parakeet. One section with a thickened epithelium inNissl staining slightly resembled a rudimentary PTO(fig. 4A) as described by Oldenstam [1925], but the adja-cent GABA-labeled section did not contain GABA-labeledcells in this region (fig. 4B). Because other known GABAer-gic structures, such as the inner ear hair cells (fig. 4B) andPurkinje cells in the cerebellum (data not shown), were reli-ably labeled in the same tissue sections, we conclude that aPTO is indeed lacking in the parakeet, or that it is reducedto absolutely minimal proportions compared with otherbird species [except owls; Vitali, 1914, 1923].
Examination of a Putative PTO in the ArmadilloIt has been claimed that a small number of mammalian
species contain a PTO in their middle ears, and althoughmost of these reports are based on embryonic specimens,some PTO-like structures might persist into adulthoodamong mammals that are considered to be at the base of

70 Brain Behav Evol 2002;60:65–79 Neeser/von Bartheld
Fig. 3A–D. Details of hair cells in the paratympanic organ (PTO)and the inner ear of the alligator. A High magnification of the sensoryepithelium in the PTO showing several hair cells (arrows). Note theciliary bundles extending into the lumen. Bar = 10 Ìm. B One haircell extends its ciliary bundle into the lumen of the PTO (arrow).
Several stereocilia can be seen in focus. Bar = 10 Ìm. C Hair cells,some with a long ciliary bundle (arrow), in the inner ear of the alliga-tor. Bar = 10 Ìm. D Section through the PTO after application of thefluorescent tracer DiI to the vestibular brainstem. 25 Ìm cryosectionwas coverslipped with Gelmount™ prior to photography. Three cells

Paratympanic Organ in Birds and Reptiles Brain Behav Evol 2002;60:65–79 71
the mammalian phylogenetic tree, such as monotremes,marsupials, and xenarthans [Miyamoto and Goodman,1986]. Monotremes and marsupials may possess a PTO[table 1; Simonetta, 1953], but not the three-toed sloth, alineage that diverged more recently from anteaters [Del-suc et al., 2001]. Recent molecular data suggest the arma-dillo as the sister group of ferungulates (cetaceans) andplace the early emergence of armadillos around the Creta-ceous/Tertiary boundary [Arnason et al., 1997; Delsuc etal., 2001]. These reports render the armadillo a species ofparticular interest with regards to the presence or absenceof a PTO, but the middle ear of this species has not yetbeen examined for a PTO [Simonetta, 1953].
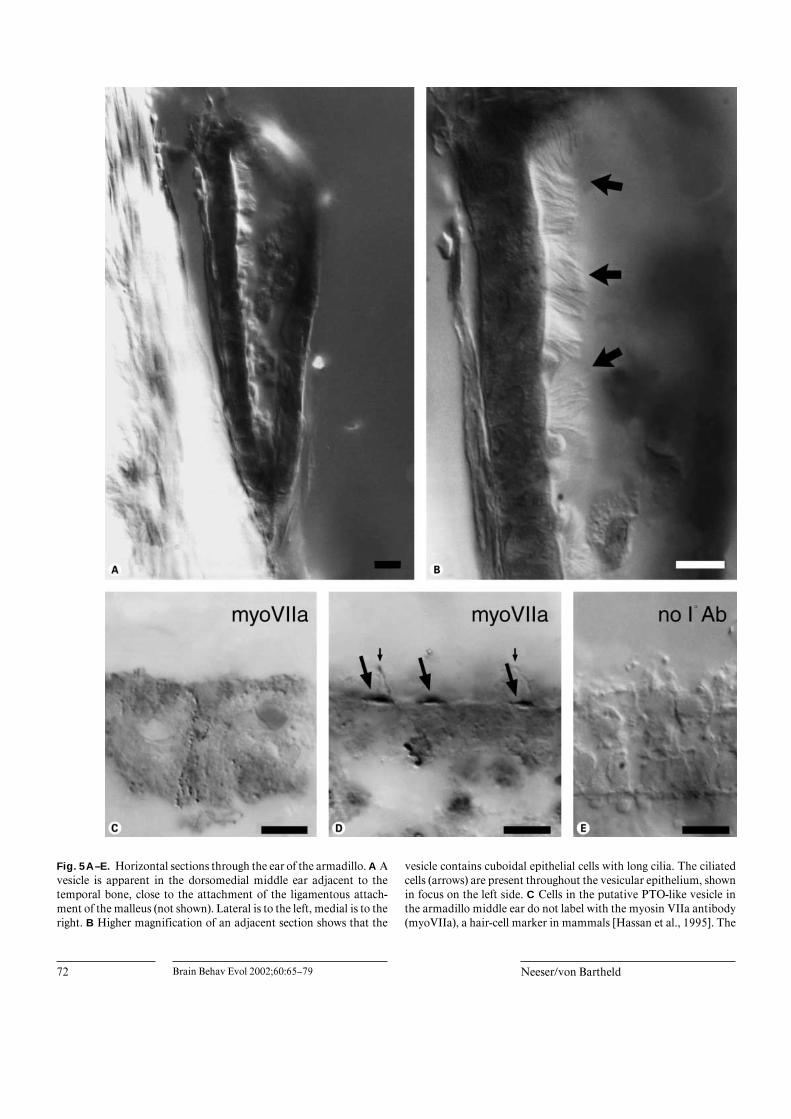
The middle ear of the armadillo contains a small vesicleabout 300 Ìm in length adjacent to the temporal bonewhere the malleus attaches with an anterolateral ligament(fig. 5A). This vesicle contains an epithelium of cuboidalcells bearing long cilia (fig. 5B). The lumen appears largelyclear. The ciliated cells do not have the appearance of typi-cal inner ear hair cells; the cilia are much longer (10–15 Ìm,fig. 5B) and resemble epidermal cilia-bearing cells reportedin the lamprey ear [Popper and Hoxter, 1987] and in thecanal pores of the lateral line in the fishes Polypterus andAcipenser [Webb and Northcutt, 1991]. To determine ifthese cells in the armadillo show the properties of hair cells,we immunolabeled sections through the putative armadilloPTO with antibodies against myosin VIIa, which arethought to be specific markers of mammalian hair cells[Hasson et al., 1995]. The ciliated cells in this vesicle didnot label with the myosin VIIa antibody (fig. 5C) althougharmadillo hair cells in the vestibular and cochlear endor-gans consistently labeled in the same tissue sections(fig. 5D). Label of inner ear hair cells was always restrictedto the apical cytoplasm underlying the base of the stereo-
(presumptive hair cells, arrows) are labeled in the PTO. In addition,at least one nerve fiber is labeled by punctate specks (marked witharrowheads) in the region adjacent to the sensory epithelium. Part ofthe lateral epithelium of the PTO broke away (stippled line) when thesection was collected on the slide. Bar = 50 Ìm.
Fig. 4A–C. Sections through the middle ear of a parakeet, Melopsittacus undulatus. A Nissl (thionin)-stained cryosec-tion through the region of the middle ear which in other bird species contains a paratympanic organ (PTO). Thethickened epithelium may represent a regressed PTO. B Adjacent section through the thickened epithelium immuno-labeled for GABA. Note the absence of GABA-labeled cells in the putative regressed PTO. C A different region fromthe same tissue section as in B showing robust immunolabel for GABA in hair cells in the vestibular sensory epithe-lium of the inner ear. Note that virtually all hair cells (arrows) are labeled. Scale bars for all panels: 10 Ìm.
72 Brain Behav Evol 2002;60:65–79 Neeser/von Bartheld
Fig. 5A–E. Horizontal sections through the ear of the armadillo. A Avesicle is apparent in the dorsomedial middle ear adjacent to thetemporal bone, close to the attachment of the ligamentous attach-ment of the malleus (not shown). Lateral is to the left, medial is to theright. B Higher magnification of an adjacent section shows that the
vesicle contains cuboidal epithelial cells with long cilia. The ciliatedcells (arrows) are present throughout the vesicular epithelium, shownin focus on the left side. C Cells in the putative PTO-like vesicle inthe armadillo middle ear do not label with the myosin VIIa antibody(myoVIIa), a hair-cell marker in mammals [Hassan et al., 1995]. The

Paratympanic Organ in Birds and Reptiles Brain Behav Evol 2002;60:65–79 73
cila, but often included one long cilium, possibly the kino-cilium (fig. 5D). No label was seen when the primary anti-body was omitted (fig. 5E). We conclude that the ‘PTO-like’ vesicle has either transdifferentiated and lost hair-celllike characteristics in this species, or that this vesicle in thearmadillo is not related at all to a PTO. These data indicatethat one has to use caution in interpreting the identity ofPTO-like vesicles when such conclusions are based entirelyon the appearance of an epithelium-lined vesicle as in pre-vious reports [Vitali, 1924; Simonetta, 1953].
Discussion
Evidence for a PTO in a ReptileAlthough the existence of a PTO among reptiles was
suggested almost half a century ago [Simonetta, 1953],this claim has not been examined or confirmed and hasreceived very little attention. Here we provide unequivo-cal evidence for a PTO in the juvenile alligator, based onfive criteria: typical morphology, conserved location,presence of hair cells, relation to ligaments, and innerva-tion. The question of whether or not a PTO exists exclu-sively among bird species is important from a compara-tive anatomical and physiological standpoint, as well asan evolutionary perspective. If a PTO is present amongreptiles, and possibly some mammals, then it appearslikely that a PTO was present among the ancestral verte-brates that gave rise to amniotes (fig. 6), as discussed inmore detail below. Simonetta [1953] did not detect a PTOin adult crocodiles, but described the organ in embryonicstages of alligators. The organ was differently developedin the 4 middle ears that were available to us from two2-year old alligators, raising the possibility that PTOs can
long cilia shown in B did not withstand the repeated washes neces-sary for the immunolabeling. D A section through the utricle showsstrong immunolabel with the myosin VIIa antibody in a disc-shapedregion within the apical cytoplasm of each hair cell (large arrows, 3hair cells in focus). Two of the hair cells also show a labeled cilium,possibly the kinocilium (small arrows). E In adjacent sectionsthrough the utricle, label was abolished when the primary antibodywas omitted. Scale bars for all panels: 10 Ìm.
Fig. 6. Schematic drawing of the lineages ofmammals and reptiles and current knowledgeabout the distribution of paratympanic organs(PTOs) among classes. The phylogenetic treewas adapted from Manley [2000] and takes intoaccount recent molecular evidence that turtlesmay be more distant to squamates (lizards andsnakes) and more closely related to archosaurs[Hedges and Poling, 1999; Zardoya and Meyer,2001]. The distribution of the PTO is shownaccording to Vitali [1920, 1924], Simonetta[1953], and the present study. ‘PTO?’ indicatesthat histology of at least one species within thislineage suggests that a PTO might be present,but conclusive evidence is lacking.
Mono-tremes
Placentals,Marsupials
Lizards,Snakes
Croco-dilesTuatara Turtles Birds
Lepidosaurs
Archosaurs
Tetrapods
Synapsids
PTO? PTO? PTO? PTO PTO
74 Brain Behav Evol 2002;60:65–79 Neeser/von Bartheld
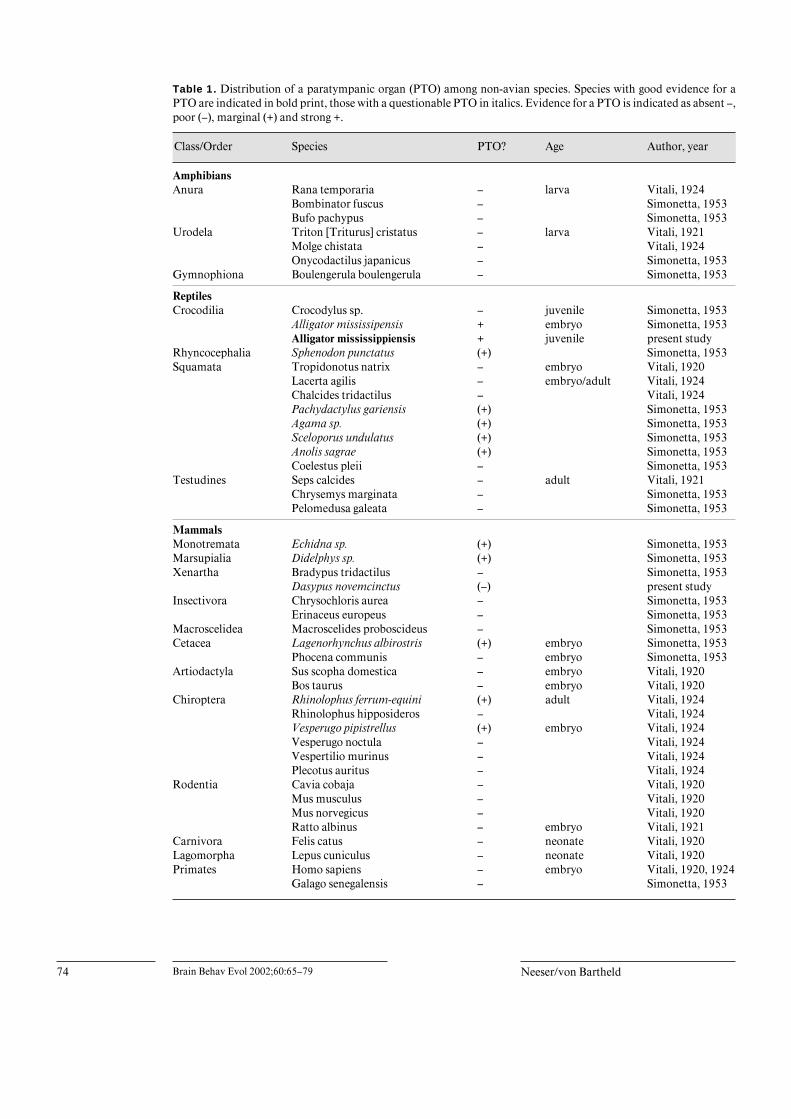
Table 1. Distribution of a paratympanic organ (PTO) among non-avian species. Species with good evidence for aPTO are indicated in bold print, those with a questionable PTO in italics. Evidence for a PTO is indicated as absent –,poor (–), marginal (+) and strong +.
Class/Order Species PTO? Age Author, year
AmphibiansAnura Rana temporaria – larva Vitali, 1924
Bombinator fuscus – Simonetta, 1953Bufo pachypus – Simonetta, 1953
Urodela Triton [Triturus] cristatus – larva Vitali, 1921Molge chistata – Vitali, 1924Onycodactilus japanicus – Simonetta, 1953
Gymnophiona Boulengerula boulengerula – Simonetta, 1953
ReptilesCrocodilia Crocodylus sp. – juvenile Simonetta, 1953
Alligator mississipensis + embryo Simonetta, 1953Alligator mississippiensis + juvenile present study
Rhyncocephalia Sphenodon punctatus (+) Simonetta, 1953Squamata Tropidonotus natrix – embryo Vitali, 1920
Lacerta agilis – embryo/adult Vitali, 1924Chalcides tridactilus – Vitali, 1924Pachydactylus gariensis (+) Simonetta, 1953Agama sp. (+) Simonetta, 1953Sceloporus undulatus (+) Simonetta, 1953Anolis sagrae (+) Simonetta, 1953Coelestus pleii – Simonetta, 1953
Testudines Seps calcides – adult Vitali, 1921Chrysemys marginata – Simonetta, 1953Pelomedusa galeata – Simonetta, 1953
MammalsMonotremata Echidna sp. (+) Simonetta, 1953Marsupialia Didelphys sp. (+) Simonetta, 1953Xenartha Bradypus tridactilus – Simonetta, 1953
Dasypus novemcinctus (–) present studyInsectivora Chrysochloris aurea – Simonetta, 1953
Erinaceus europeus – Simonetta, 1953Macroscelidea Macroscelides proboscideus – Simonetta, 1953Cetacea Lagenorhynchus albirostris (+) embryo Simonetta, 1953
Phocena communis – embryo Simonetta, 1953Artiodactyla Sus scopha domestica – embryo Vitali, 1920
Bos taurus – embryo Vitali, 1920Chiroptera Rhinolophus ferrum-equini (+) adult Vitali, 1924
Rhinolophus hipposideros – Vitali, 1924Vesperugo pipistrellus (+) embryo Vitali, 1924Vesperugo noctula – Vitali, 1924Vespertilio murinus – Vitali, 1924Plecotus auritus – Vitali, 1924
Rodentia Cavia cobaja – Vitali, 1920Mus musculus – Vitali, 1920Mus norvegicus – Vitali, 1920Ratto albinus – embryo Vitali, 1921
Carnivora Felis catus – neonate Vitali, 1920Lagomorpha Lepus cuniculus – neonate Vitali, 1920Primates Homo sapiens – embryo Vitali, 1920, 1924
Galago senegalensis – Simonetta, 1953

Paratympanic Organ in Birds and Reptiles Brain Behav Evol 2002;60:65–79 75
be in different stages of regression. Nevertheless, someregression does not preclude the possibility that the PTOstill has a functional role in reptiles. The primary role isprobably the detection of changes in barometric or hy-drostatic pressure as suggested in fishes for the spiracularsense organ (SSO) and in birds for the PTO [Ranzi, 1926;von Bartheld, 1994]. The crocodilian middle ear is notthought to flood when the head is submerged in water[Wever, 1978], similar to aquatic birds that contain well-developed PTOs, presumably for detection of hydrostaticpressure [Vitali, 1941]. The PTO should no longer beignored as a purely avian curiosity [Butler and Hodos,1996], but rather deserves to be included as a more gener-al vertebrate phenomenon in future editions and text-books of comparative vertebrate biology.
Why Do Certain Bird Species Lack a PTO?For almost a century it seemed peculiar that a small
number of bird species, which are not particularly closelyrelated, lacked a PTO [Vitali, 1914, 1923; Oldenstam,1925], while the large majority of avian species possess awell-developed PTO (table 2). We now offer a plausibleexplanation for this mystery. Birds that possess a PTO allhave an upper beak that is relatively fixed, allowing littlemotion, and only the lower beak moves [Platner, 1839;Pohlmann, 1921; Stellbogen, 1930]. Birds lacking a PTO(owls, parakeets) all show a considerable movement of theupper beak. ‘In birds with a freely movable upper bill, asin the parrot, the quadratosquamosal articulation is ...more pronouncedly a hinge than in birds with little upperbill movement, as in chicken’ [Pohlmann, 1921]. Owlsand parakeets are among the birds with the most extremeupper beak movement [Platner, 1839; Pohlmann, 1921,1922; Stellbogen, 1930], resulting in the largest movementin the quadratosquamosal articulation. This joint is invery close proximity to the PTO. A moving upper beakappears to be incompatible with a functional PTO, andthis might be the reason why the PTO regresses in thesebirds. The organ does seem to be present in embryonicowls [Vitali, 1923], indicating that the PTO initiallydevelops in all birds, but then its development is sup-pressed in those bird species with significant upper billmovement. These species also possibly develop a ring-likebony frame for attachment of the tympanic membranewhich minimizes movements of the tympanic membranewhen ‘the quadrate bone ... participates in a swingingmotion with movements of the upper bill’ [Pohlmann,1922]. Neither owls nor parakeets have a lifestyle thatwould make PTOs particularly advantageous, unlikesmall birds that rely on barometric information for weath-
er changes and upcoming (winter) storms [Buttemer,1985]. Migratory birds might require barometers forchoosing favorable weather conditions [Kreithen andKeeton, 1974] and could utilize the PTO as an altimeterto maintain altitudes when flying considerable distancesat night or in cloud cover [Griffin, 1969; Kreithen andKeeton, 1974; von Bartheld, 1994]. Absence of a PTO inowls and parakeets may present no major disadvantage,owing to the lifestyles of these birds that do not migrateand do not need to change their behavior much withupcoming winterstorms, unlike smaller birds whose lifemight depend on a functional PTO [Buttemer, 1985].
The initial suggestion that the PTO might be an ‘organof flight’, based on the observation that lesions of the PTOreduced flight [Ruffini, 1920; Vitali, 1921; Benjamins,1926], can now be layed to rest, because such observationscould not be reproduced in subsequent lesion experi-ments, navigation in homing pigeons was not impairedafter ear or PTO lesions, and some bird species lacking aPTO (owls, parakeets) can fly better than some birds withPTOs (such as chickens and penguins) [Benjamins, 1939;Wallraf, 1972; Giannessi et al., 1996].
A Role of the PTO in the Control of the StapediusMuscle and Reflex?The presence of a PTO in birds furthermore appears to
correlate with the absence of an acoustic stapedius reflex.It was noted many years ago that, unlike in mammalianspecies, the stapedius muscle of most birds is activated bynon-acoustic stimuli [Wada, 1924; Oeckinghaus andSchwartzkopff, 1975, 1983; Counter and Borg, 1979;Grassi et al., 1988; Borg and Counter, 1989]. Apparentlyan exception to this rule are owls in which an acousticstapedius reflex has been demonstrated [Golubeva, 1972;Oeckinghaus and Schwartzkopff, 1983]. An explanationfor these surprising species differences is lacking, but ithas been suggested that differences in the neural control ofthe stapedius muscle might play a role [Grassi et al., 1983;Borg and Counter, 1989]. Because the PTO appears tomediate the afferent input to the centers that control thestapedius muscle [von Bartheld, 1994], it is possible thatthere are two different types of neural circuitry amongbirds: One that utilizes acoustic stimuli to adjust the ten-sion of the tympanic membrane (owls), and another thatis sensitive to barometric pressure [Pohlmann, 1921,1922; Wada, 1924] and utilizes information conveyed tothe CNS via the PTO (most other birds). Such conclusionsmake it likely that the PTO is intimately involved in thecontrol of tympanic membrane tension [von Bartheld,1994].

76 Brain Behav Evol 2002;60:65–79 Neeser/von Bartheld
Table 2. Negative correlation of the distribution of a paratympanic organ (PTO) and an acoustic stapedius reflex among birds. + = Presence ofPTO; +++ = hypertrophied PTO; – = absence of PTO
Species Para-tympanicorgan?
Author, year Acousticstapediusreflex?
Author, year
StruthioniformesStruthio camelus + Simonetta, 1953
GalliformesGallus domesticus + Vitali, 1914; Oldenstam, 1925 – Wada, 1924; Counter and Borg, 1979Melegris gallopavo + Vitali, 1914Perdix cinerea + Vitali, 1914
AnseriformesAnas boschas + Vitali, 1914; Oldenstam, 1925 – Wada, 1924Anser anser + Simonetta, 1953 – Wada, 1924Ceronectes maccoa + Simonetta, 1953
SphenisciformesSpheniscus demersus + Simonetta, 1953’penguin’ + Benjamins, 1939
GruiformesFulica atra + Vitali, 1914
CharadriiformesPluvialis apricaria + Jørgensen, 1984Scolopax rusticola + Vitali, 1914Numenius arquata + Jørgensen, 1984‘sea gull’ + Oldenstam, 1925
ColumbiformesColumba livia + Vitali, 1914; Oldenstam, 1925;
Jørgensen, 1984– Wada, 1924
Streptopelia decaocto + Jørgensen, 1984Psittaciformes
Melopsittacus undulatus – Oldenstam, 1925; present studyFalconiformes
Accipiter nisus + Vitali, 1914Tinnunculus tinnunculus + Vitali, 1914
StrigiformesAsio otus + Golubeva, 1972Athene noctua – Vitali, 1914; 1923Strix flammea – Vitali, 1914; 1923Strix aluco + Oeckinghaus and Schwartzkopff, 1983
MicropodiformesApus apus +
PasseriformesPasser domesticus italiae + Vitali, 1911; 1914 – Wada, 1924 Parus caeruleus + Jørgensen, 1984Taeniopygia guttata + Jørgensen, 1984Serinus canaria – Wada, 1924Fringilla chloris + Oldenstam, 1925Fringilla cannabina + Oldenstam, 1925Fringilla spinus + Oldenstam, 1925Fringilla coelebs + Vitali, 1914Emberica citrinella + Oldenstam, 1925Turdus iliacus + Oldenstam, 1925Turdus musicus + Vitali, 1914Hirundo rustica + Vitali, 1914Sturnus vulgaris + Jørgensen, 1984 – Oeckinghaus and Schwartzkopff, 1983Garrulus glandarius + Vitali, 1914Coccothraustes coccothraustes + Vitali, 1914Chelidon urbica + Vitali, 1914Cipselus apus +++ Vitali, 1914

Paratympanic Organ in Birds and Reptiles Brain Behav Evol 2002;60:65–79 77
Distribution of the PTO/SSO among VertebratesIt has been suggested that the PTO is a lateral line rem-
nant that is homologous to the spiracular sense organ(SSO) in fishes [Vitali, 1925; Ranzi, 1926; Simonetta,1953; Barry and Boord, 1984; Barry and Bennett, 1989;von Bartheld, 1990; von Bartheld and Rubel, 1990]. Allfishes examined, except for teleosts, possess an SSO,including lungfishes, sharks, rays, chimaeras, sturgeons,gars, paddlefishes, bowfins and bichirs [for review, seeSimonetta, 1953]. If the PTO developed from an SSO-likeorgan, one would expect the PTO to be present amongseveral vertebrate classes, and one would further expectthe PTO to develop early in ontogeny, based on the ‘gen-eral rule’ that whatever develops early in phylogeny alsodevelops early in ontogeny [Jacobson, 1978]. Reports sug-gesting that presumptive PTOs may be more widespreadamong vertebrates during development (in Cetacean, bat,and reptilian embryos) than in adults [Vitali, 1920, 1924;Simonetta, 1953] is consistent with this hypothesis (ta-ble 1). The potential (anlage) for a PTO/SSO might beinherited from SSO-bearing ancestors, not necessarilyresulting in a fully developed PTO that is retained intoadulthood. The possible persistence of the PTO in selectspecies among non-avian vertebrates into adulthood is ofinterest. Our study shows this unambiguously for a rep-tile. Whether a PTO indeed persists in orders with a basalposition among mammals, such as prototherians (mono-tremes) and metatherians (marsupials) as suggested bySimonetta [1953], should be evaluated with modern tech-niques. Our data revealed that a vesicle in the armadillomiddle ear with some resemblence to a PTO does not con-tain true hair cells, but rather cilia-bearing epidermal cells[Popper and Hoxter, 1987; Webb and Northcutt, 1991].This underscores the necessity to treat reports on PTO-like structures in mammals and reptiles with caution.Although such structures might have regressed or trans-differentiated from a PTO or SSO, it is also possible thatthey are unrelated to the PTO in their ontogenetic andevolutionary history.
Evolution of the SSO/PTO: Implications for theEvolution of HearingThe middle ear is thought to have been crucial for the
evolutionary development of the cochlea [Manley, 2000].If it is true that the PTO (or SSO) is always associated withthe hyomandibula (stapes), this was probably also true forancestral vertebrates that gave rise to tetrapods. It is nowassumed that the hyomandibular bone or stapes in ances-tral tetrapods carried muscles operating a spiracular valve[Clack, 1994]. The early stapes controlled palatal and spi-
racular movements (for respiration) rather than support-ing a tympanum or ear drum (for hearing) [Clack, 1989,1990, 1994]. It is likely that the same is true for the func-tion of the sense organ associated with that bony struc-ture. This leaves us with the following probable scenario:The ancestral PTO was a sense organ that was part of theafferent control for spiracular movements, for the pur-poses of breathing, as the SSO possibly still functions infishes. The spiracle in early tetrapods might have beenused for inspiration when the rest of the animal’s headwas submerged [Clack, 1989]. When the hyomandibularbone changed its shape and function and eventuallybecame involved in terrestrial hearing [Clack, 1990,1994], the PTO became superfluous, and it disappearedin most species, except in birds and a few other specieswhere control of the tension of the tympanic membraneand associated functions required an afferent sensoryinput. Thus, the evolutionary history of the PTO could bemore intimately involved with the evolutionary biology ofhearing in tetrapods than previously appreciated.
Acknowledgments
This work was supported in part by NIH grant DC 00019(C.S.v.B.), DFG grants to Dietrich Meyer (Göttingen, Germany),and NIH grants to Edwin Rubel (Seattle, Wash.). We thank JeffWoodbury (American Museum of Natural History, N.Y.) for provid-ing two alligator specimens. Two adult parakeet specimens werekindly provided from the collection of Dietrich Meyer (University ofGöttingen, Germany). The Mote Marine Laboratory (Sarasota, Fla.)kindly provided an armadillo specimen from their collections. Wealso thank Nico Schellart (University of Amsterdam, The Nether-lands) for his help with the armadillo preparation, Tama Hasson (UCIrvine, Calif.) for the myosin VII antibody, Bernd Fritzsch (Creigh-ton University, Nebr.) for advice, Hanna von Bartheld (Bremen,Germany) for help in obtaining the older European PTO literature,and two anonymous reviewers for helpful comments.

78 Brain Behav Evol 2002;60:65–79 Neeser/von Bartheld
References
Arnason, U., A. Gullberg, and A. Janke (1997) Phy-logenetic analyses of mitochondrial DNA sug-gest a sister group relationship between Xenar-tha (Edentata) and Ferungulates. Mol. Biol.Evol., 14: 762–768.
Barry, M.A., and R.L. Boord (1984) The spiracularorgan of sharks and skates: Anatomical evi-dence indicating a mechanoreceptive role.Science, 226: 990–992.
Barry, M.A., and M.V.L. Bennett (1989) Special-ized lateral line receptor systems in elasmo-branchs: the spiracular organs and vesicles ofSavi. In The Mechanosensory Lateral Line:Neurobiology and Evolution (ed. by S.Coombs, P. Görner and H. Münz), Springer,New York, pp. 591–606.
Benjamins, C.E. (1926) Y a-t-il une relation entrel’organe paratympanique de Vitali et le vol desoiseaux? Arch. Neerl. Physiol., 11: 215–222.
Benjamins, C.E. (1939) L’ organe paratympaniquede Vitali, soi-disant organe du vol des oiseaux,chez le Pingouin. Acta Otolaryngol. (Stockh.),27: 266–270.
Borg, E., and S.A. Counter (1989) The middle earmuscles. Sci. Am., 261: 74–80.
Butler, A.B., and W. Hodos (1996) ComparativeVertebrate Neuroanatomy. Evolution and Ad-aptation. Wiley, New York.
Buttemer, W.A. (1985) Energy relations of winterroost-site utilization by American goldfinches(Carduelis tristis). Oecologia, 68: 126–132.
Clack, J.A. (1989) Discovery of the earliest knowntetrapod stapes. Nature, 342: 425–427.
Clack, J.A. (1990) Nos ancêtres respiraient-ils parles oreilles? (Did our ancestors breathe throughtheir ears?). Recherche, 21: 770–772.
Clack, J.A. (1994) Earliest known tetrapod brain-case and the evolution of the stapes and fenes-tra ovalis. Nature, 369: 392–394.
Counter, S.A., and E. Borg (1979) Physiologicalactivation of the stapedius muscle in Gallusgallus. Acta Otolaryngol., 403: 13–19.
Delius, J.D., and J. Emmerton (1978) Sensorymechanisms related to homing in pigeons; InAnimal Migration, Navigation and Homing(ed. by K. Schmidt-Koenig and W.T. Keeton),Springer, Berlin, pp. 35–41.
Delsuc, F., F.M. Catzeflis, M.J. Stanhope, andE.J.P. Douzery (2001) The evolution of arma-dillos, anteaters and sloths depicted by nuclearand mitochondrial phylogenies: Implicationsfor the status of the enigmatic fossil Eurota-mandua. P. Roy. Soc. Lond. Ser. B-Biol. Sci.,268: 1605–1615.
Federici, F. (1927) Über die Innervation des vonVitali entdeckten Sinnesorgans im Mittelohrder Vögel (sogen. paratympanisches Organ).Anat. Anz., 62: 241–254.
Giannessi, F., and L. Pera (1986) The ultrastruc-ture of the paratympanic organ in the domesticfowl (Gallus gallus domesticus). J. Anat., 147:191–199.
Giannessi, F., and R. Ruffoli (1996) The ultrastruc-ture of the sensory hairs of the paratympanicorgan receptor cells in chicken. Anat. Embryol.(Berl), 193: 569–575.
Giannessi, F., B. Fattori, R. Ruffoli, and A. Ga-gliardo (1996) Homing experiments on pigeonssubjected to bilateral destruction of the para-tympanic organ. J. Exp. Biol., 199: 2035–2039.
Golubeva, T.B. (1972) The reflex activity of thetympanal muscle in the owl Asio otus. Zhurn.Evol. Biochem. Fisiol., 8: 173–181. (J. Evol.Biochem. Physiol., 8: 149–155).
Grassi, S., F. Magni, and F. Ottaviani (1983) Iden-tification of the motoneurons innervating thestapedius muscle in Gallus gallus: A horserad-ish peroxidase study. Arch. Ital. Biol., 121: 37–45.
Grassi, S., F. Magni, and F. Ottaviani (1988) Mech-anisms controlling vocalization-evoked stape-dius muscle activity in chickens (Gallus gallus).J. Comp. Physiol. A, 162: 525–532.
Griffin, D.R. (1969) The physiology and geophy-sics of bird navigation. Q. Rev. Biol., 44: 255–276.
Hasson, T., M.B. Heintzelman, J. Santos-Sacchi,D.P. Corey, and M.S. Mooseker (1995) Expres-sion in cochlea and retina of myosin VIIa, thegene product defective in Usher syndrometype1B. Proc. Natl. Acad. Sci. U.S.A., 92:9815–9819.
Hedges, S.B., and L.L. Poling (1999) A molecularphylogeny of reptiles. Science, 283: 998–1001.
Hofmann, M.H., and H. Bleckmann (1999) Effectof temperature and calcium on transneuronaldiffusion of DiI in fixed brain preparations. J.Neurosci. Methods, 88: 27–31.
Jacobson, M. (1978) Developmental Neurobiology,2nd ed., Plenum Press, New York.
Jørgensen, J.M. (1984) Fine structure of the para-tympanic organ in the avian middle ear. ActaZool., 65: 89–94.
Kreithen, M.L., and W.T. Keeton (1974) Detectionof changes in atmospheric pressure by the hom-ing pigeon, Columba livia. J. Comp. Physiol.,89: 73–82.
Manley, G.A. (2000) Cochlear mechanisms froma phylogenetic viewpoint. Proc. Natl. Acad.U.S.A., 97: 11736–11743.
Miyamoto, M.M., and M. Goodman (1986) Bio-molecular systematics of eutherian mammals –phylogenetic patterns and classification. Syst.Zool., 35: 230–240.
Neeser, J.A., and C.S. von Bartheld (2001) Evi-dence for a paratympanic organ (presumed lat-eral line remnant) in the middle ear of juvenilealligators: implications for the phylogeny ofhearing. Brain Behav. Evol., 57: 229.
Oeckinghaus, H., and J. Schwartzkopff (1975)Elektrische Aktivierung des Mittelohrmuskelsbeim Star. Naturwissenschaften, 62: 582.
Oeckinghaus, H., and J. Schwartzkopff (1983) Elec-trical and acoustical activiation of the middleear muscle in a songbird. J. Comp. Physiol.,150: 61–67.
Oldenstam, R.A. (1925) Over het z.g.n. vliegeor-gaan van Vitali in het middenoor der vogels.Ph.D. dissertation. University of Groningen,Groningen.
Petrash, E.W., K.H. Andres, M. Von Düring, andJ.D. Delius (1983) Morphologische Aspektedes Vitali-Organs bei der Taube. Anat. Anz.,153: 288.
Platner, F. (1839) Bemerkungen über das Quadrat-bein und die Paukenhöhle der Vögel. GerhardFleischer, Leipzig.
Pohlman, A.G. (1921) The position and functionalinterpretation of the elastic ligaments in themiddle ear region of Gallus. J. Morphol., 35:229–262.
Pohlman, A.G. (1922) The problem of middle earmechanics. Ann. Otol. Rhinol. Laryngol., 31:1–45.
Popper, A.N., and B. Hoxter (1987) Sensory andnonsensory ciliated cells in the ear of the sealamprey, Petromyzon marinus. Brain Behav.Evol., 30: 43–61.
Ranzi, S. (1926) L’organo di senso spiracolare deiselaci. Pubbl. Staz. Zool. Napoli, 7: 37–76.
Ruffini, A. (1920) Sull’organo nervoso paratimpan-ico di G. Vitali od organo del volo degli uccelli.Boll. Sci. med. Soc. med.-chir.Bologna, pp.141–154/ Archo ital. Otol. Rhinol. Lar., 31:397–413.
Simonetta, A. (1953) L’organo di senso dello spira-colo e l’organo paratimpanico nella sistematicadei vertebrati. Arch. Ital. Anat. Embryol., 58:266–294.
Stellbogen, E. (1930) Über das äussere und mittlereOhr des Waldkauzes (Syrnium aluco, L.). Z.Morphol. Okol. Tiere, 19: 686–731.
Usami, S., M. Igarashi, and G.C. Thompson (1987)GABA-like immunoreactivity in the chick ves-tibular end organs. Brain Res., 418: 383–387.
Vitali, G. (1911) Di un interessante derivato dellaprima fessura branchiale nel passero. Anat.Anz., 39: 219–224.
Vitali, G. (1914) Di un nuovo organo nervoso disenso nell’orecchio medio degli uccelli. Ulter-iore destino dell’organo della prima fessurabranchiale. Int. Msch. Anat. Physiol., 30: 363–428.
Vitali, G. (1920) Sulla presenza nei chirotteridell’organo nervoso di senso da me scopertonell’orecchio medio degli uccelli. Atti R. Acad.d. Fisiocritici in Siena, 12: 521–524.
Vitali, G. (1921) L’organo nervoso paratimpanico ela sua funzione. Riv. Biol., 3: 302–316.
Vitali, G. (1923) Sul comportamento dell’organodella prima fessura branchiale in Athene noc-tua. Richerche Morfol., 3: 153–168.
Vitali, G. (1924) Il compartamento dell’organo del-la prima fessura branchiale negli anfibi, nei ret-tili e nei mammiferi. Richerche Morfol., 4:191–221.
Vitali, G. (1925) Il comportamento dell’organo del-la prima fessura branchiale (placode epibran-chiale) nei selaci. Monitore Zool. Ital., 36: 122–130.
Vitali, G. (1941) Alcune considerazioni sulla pub-blicazione del Prof. Benjamins della presenzadell’organo paratimpanico nel Pinguino. Anat.Anz., 91: 364–368.

Paratympanic Organ in Birds and Reptiles Brain Behav Evol 2002;60:65–79 79
von Bartheld, C.S. (1990) Development and inner-vation of the paratympanic organ (Vitali organ)in chick embryos. Brain Behav. Evol., 35: 1–15.
von Bartheld, C.S. (1994) Functional morphologyof the paratympanic organ in the middle ear ofbirds. Brain Behav. Evol., 44: 61–73.
von Bartheld, C.S., and E.W. Rubel (1990) Para-tympanic and spiracular sense organs: Phyloge-netic distribution and theories of function, in-cluding hearing. In Evolutionary Biology ofHearing (ed. by D.B. Webster, R.R. Fay andA.N. Popper), Springer, New York, pp. 582–583.
von Bartheld, C.S., D.E. Cunningham, and E.W.Rubel (1990) Neuronal tracing with DiI: Decal-cification, cryosectioning, and photoconver-sion for light and electron microscopic analysis.J. Histochem. Cytochem., 38: 725–733.
von Bartheld, C.S., W. Yang, and E.W. Rubel(1992) Normal and abnormal pathfinding offacial nerve fibers in the chick embryo. J. Neu-robiol., 23: 1021–36.
Wada, Y. (1924) Beiträge zur vergleichenden Phy-siologie des Gehörorganes. Pflüg. Arch., 202:46–69.
Wallraf, H.G. (1972) Homing of pigeons after extir-pation of their cochleae and lagenae. NatureNew Biol. (London), 236: 223–224.
Webb J.F., and R.G. Northcutt (1991) Ciliated epi-dermal-cells in non-teleost actinopterygianfish. Acta Zool. (Stockholm), 72: 107–111.
Wever, E.G. (1978) The Reptile Ear: Its Structureand Function. Princeton University Press,Princeton, N.J.
Yntema, C.L. (1944) Experiments on the origin ofthe sensory ganglia of the facial nerve in thechick. J. Comp. Neurol., 81: 147–167.
Zardoya, R., and A. Meyer (2001) The evolutiona-ry position of turtles revised. Naturwissen-schaften, 88: 193–200.