Embed Size (px)
Citation preview

Coexpression of Functional P2X and P2YNucleotide Receptors in Single CerebellarGranule Cells
Cristina Hervas, Raquel Perez-Sen, and MaTeresa Miras-Portugal*Biochemistry Department, Veterinary Faculty, Complutense University of Madrid, Madrid, Spain
The present study describes the presence and expres-sion of functional nucleotide receptors, both ionotropicand metabotropic, in highly purified cultures of cerebellargranule neurons. Microfluorimetric experiments havebeen carried out to record specific [Ca2�]i transients inindividual granule neurons after challenge with diversenucleotides. Although great heterogeneity was found innucleotide responses in single cells, these responses allbecame modified during the course of granule cell dif-ferentiation, not only at the level of the number of re-sponding cells, but also in the magnitude of the responseto nucleotides. These in vitro developmental changeswere more significant in metabotropic responses to py-rimidine nucleotides, UTP and UDP, which were down-and upregulated, respectively, during the time in culture.At least two types of ADP-specific receptors seem ex-pressed in different granule cell subpopulations respond-ing to 2MeSADP, as the specific P2Y1 antagonist MRS-2179 inhibited Ca2� responses in only one of thesepopulations. The great diversity of metabotropic re-sponses observed was confirmed by the RT-PCR ex-pression of different types of P2Y receptors in granulecell cultures: P2Y1, P2Y4, P2Y6, and P2Y12. Similarly,ionotropic nucleotide responses were confirmed by thepresence of specific messengers for different P2X sub-units, and by immunolabeling studies (P2X1, P2X2, P2X3,P2X4 and P2X7). Immunolabeling reflected great varietyin the P2X subunit distribution along the granule neuroncytoarchitecture, with P2X2, P2X3 and P2X4 present atsomatodendritic locations, and P2X1, P2X7, and P2X3,located at the axodendritic prolongations. The punctu-ated labeling pattern obtained for P2X3 and P2X7 sub-units is particularly notable, as it presents a high degreeof colocalization with synaptophysin, a specific marker ofsynaptic vesicles, suggesting specialized localizationand function in granule neurons. © 2003 Wiley-Liss, Inc.
Key words: granule neurons; purinergic receptors; mi-crofluorimetry
Purine and pyrimidine nucleotides, once released atthe extracellular space, play an important role as signalingmolecules, and large variability exists within their receptorfamilies and signaling mechanisms. They have been
grouped in two broad families: P2Y receptors that corre-spond to metabotropic receptors coupled to G proteins,and P2X receptors that are ionotropic (for review, seeAbbracchio and Burnstock, 1994; Ralevic and Burnstock,1998). The excitatory neurotransmitter role of ATP is wellestablished at neuromuscular junctions and in peripheraland central synapses (Benham and Tsien 1987; Edwards etal., 1992; Vulchanova et al., 1997; Pankratov et al., 1998).The ATP ionotropic receptor, P2X, is made up of homo-or heterooligomeric associations of seven subunits,P2X1–7, the distribution and relative abundance of whichdiffer according to brain areas and neuronal topology (forreview, see Norenberg and Illes, 2000; North, 2002). P2Xreceptor presence and relevance at the presynaptic level ofidentified central synapses has also been assessed clearly(Pintor and Miras Portugal, 1995; Gomez-Villafuertes etal., 2001; Dıaz-Hernandez et al., 2002; Lundy et al.,2002).
Purinergic neurotransmission seems to play an im-portant role in cerebellum, as high expression levels fordifferent P2X subunits have been described, especially inPurkinje cells, and to some extent, at the granular layer(Collo et al., 1996; Kanjhan et al., 1996; Vulchanova et al.,1996; Collo et al., 1997; Loesch and Burnstock, 1998).The cerebellum contains relatively few neuronal types,and its circuitry is well understood. Cerebellar granuleneurons, which represent the major cell population incerebellum, are densely packed, small neurons located inthe granular layer of cerebellar cortex. They are the maintargets of afferent mossy fibers, and they in turn contactsynaptically the dendrites of Purkinje neurons via parallelfibers. Postnatal development of cerebellum allows aneasier, highly purified culture and differentiation of gran-ular cells in culture. These cells are isolated from the early
Contract grant sponsor: Spanish Ministry of Science and Technology;Contract grant number: BF12002-03626.
*Correspondence to: Ma Teresa Miras-Portugal, Dept. Bioquımica y Bio-logıa Molecular IV, Facultad de Veterinaria, Universidad Complutense deMadrid, Avda. Puerta de Hierro s/n 28040 Madrid, Spain.E-mail: [email protected]
Received 4 February 2003; Revised 16 April 2003; Accepted 18 April 2003
Journal of Neuroscience Research 73:384–399 (2003)
© 2003 Wiley-Liss, Inc.

postnatal period, can be cultured in defined media, andcan be maintained for long periods. They exhibit gluta-matergic nature and characteristic shape, with soma andextensive neuritic networks, characteristic of the maturephenotype (Torres-Aleman et al., 1994). High potassiumconcentration in the culture medium, required for granuleneuron survival, elevates resting intracellular calcium con-centration and protects the cell against the cytotoxic effectof massive calcium entry through NMDA receptor acti-vation (Gallo et al., 1987; Stewart et al., 1995). Under theabove conditions, granule neurons develop and maturatein vitro. Several studies have demonstrated good correla-tion between cerebellum development and in vitro gran-ule cell development; thus, this is a suitable model to usein the study of neurotrophic factors related to develop-ment, differentiation, or survival of individual cell popu-lations of the central nervous system (CNS). In granuleneurons, changes in expression patterns of receptors andother structural or functional proteins have been reported,such as those for the GABAA receptor subunits (Thomp-son et al., 1996; Hevers and Luddens, 2002). In addition,the validity of this culture has been largely demonstrated,as similar glutamate response patterns have been found forgranule neurons in culture or in cerebellar slices (Courtneyet al., 1990; Kirischuk et al., 1996).
The significant expression pattern found for nucleo-tidic receptors in cerebellum extends beyond Purkinjecells to the molecular layer level where parallel fibersextend, and to the granular layer, where the granular cellsomas are located. This suggests that nucleotidic receptorsplay a key role in granule neuron physiology, although therole is not yet well understood (Kanjhan et al., 1996;Vulchanova et al., 1996; Loesch and Burnstock, 1998;Amadio et al., 2002). The purpose of this work was todemonstrate the presence and variety of both families ofP2 receptors, the P2X and P2Y, and their functionality inlong-term cultures of cerebellar granule neurons.
MATERIALS AND METHODS
Culture of Granule Cells
All experiments carried out at the Universidad Com-plutense de Madrid followed the guidelines of the InternationalCouncil for Laboratory Animal Science (ICLAS). Cerebellarcultures were prepared following procedures described byMeyer-Franke et al. (1995) and modified by Pons et al. (2001).Cerebella from three Wistar rat pups (P7) were removed asep-tically, washed once in Earl’s balanced salt solution (EBSS;Gibco BRL), cut into small 1-mm pieces, and transferred to a50-ml screw cap tube. Tissue fragments were allowed to settle,excess EBSS was aspirated, and 4 ml of EBSS containing100 U/ml DNase (Worthington, Lake Wood NJ), 1 mMCaCl2, and 1 mM MgCl2 were added. One hundred units ofpapain (Worthington) was added after it was preactivated(30 min at 37°C) in 1 ml of activation buffer containing EBSS,5 mM L-Cys (Sigma), 2 mM EDTA (Sigma), and 0.067 mM�-mercaptoethanol (Gibco BRL). Air in the tube was displacedwith 95% O2-5% CO2 and the tube was incubated at 37°C for1 hr on a shaking platform. Undigested fragments were allowed
to settle and the supernatant was centrifuged at 800 rpm for5 min. The pellet was resuspended in 3 ml of EBSS containing3 mg of ovomucoid protease inhibitor (Worthington) layeredonto an albumin cushion, which consisted of 5 ml of EBSScontaining ovomucoid protease inhibitor (Worthington) andovoalbumin (Worthington) at 10 mg/ml each, and the resus-pended pellet was centrifuged at 800 rpm for 5 min. Theresulting pellet was finally resuspended in neurobasal medium(Gibco BRL), and the cell number and viability was assessed. Onglass coverslips precoated with 0.1 mg/ml poly-L-lysine (Bio-chrom AG, Berlin), cells were plated at a density of 100,000cells/cm2 in neurobasal medium supplemented with B27 (GibcoBRL) and containing 20 mM KCl, 2 mM glutamine (Sigma),and 50�g/ml gentamicin (Gibco BRL), and were maintained ina humidified incubator at 37°C in 5% CO2. After 24 hr, 10 �MAraC (Sigma) was added to avoid glial cell proliferation. Culturemedium was replaced every 4 days.
Immunocytochemical Assays
For the immunofluorescence assay, coverslips were fixedwith 4% PFA (w/v; Sigma) for 15 min, washed twice in PBSand incubated for 1 hr in PBS containing 3% BSA (w/v; Sigma),0.1% Triton X-100 (v/v), and 5% normal goat serum (v/v;Sigma). Primary antibodies, diluted in PBS/BSA, recognized thespecified rat proteins: rabbit anti-glial fibrillar acid protein(GFAP, 1:200; Sigma), goat anti-�6 GABAA R (1:200; SantaCruz Biotechnology, Santa Cruz, CA), mouse anti-synaptophysin (1:200; Sigma), rabbit anti-VGLUT1 (1:1,000;Synaptic Systems), rabbit anti-VGLUT2 (1:500; Synaptic Sys-tems), rabbit anti-P2X1 (1:100; Chemicon, Temecula, CA),rabbit anti-P2X7 (1:100; Chemicon), guinea-pig anti-P2X2,(1:100, Chemicon), guinea-pig anti-P2X3 (1:100; Chemicon),rabbit anti-P2X4 (1:100; Alomone Labs, Jerusalem), rabbit anti-P2X5 (1:500; a gift from M. Voigt), and rabbit anti-P2X6(1:500; a gift from F. Soto). Cells were washed twice andincubated with primary antibody for 1 hr at 37°C. Cells werethen washed three times and incubated for 1 hr at 37°C with theappropriate secondary antibodies: goat anti-mouse IgG fluores-cein conjugated (1:500; Sigma), goat anti-rabbit IgG rhodamineconjugated (1:500; Sigma), goat anti-guinea pig IgG rhodamineconjugated (1:500), and donkey anti-goat IgG fluorescein con-jugated (1:500). After incubation, cells were washed three timesand mounted following standard procedures. To mark nuclei,4�,6-diamidino-2-phenylindole (DAPI) was added (1 �M for5 min) before mounting the covers. Controls were preparedaccording to the same protocol, but primary antibodies werereplaced by the same volume of PBS/BSA solution.
Coverslips were viewed with a NIKON TE-200 micro-scope and an Ultrapix 2000 Mono CCD camera controlled byUltraview PC software (Perkin Elmer Life Sciences, Cambridge,UK).
Calcium Imaging
At least 1 hr before the experiment, cells attached to glasscoverslips were washed, under normal K� conditions, withLocke’s solution (in mM: NaCl, 140; KCl, 4.5; CaCl2, 2.5;KH2PO4, 1.2; MgSO4, 1.2; glucose, 5.5; and HEPES [acid], 10;pH 7.4] supplemented with 1 mg/ml BSA. The cells were thenloaded with 5 �M Fura-2 AM (Molecular Probes, Eugene, OR)
P2X and P2Y Receptors in Granule Neurons 385

for 45 min at 37°C. The coverslip was placed in a small super-fusion chamber and superfused with Locke’s medium containing2.5 mM CaCl2. Cells were imaged through a NIKON TE-200microscope with a Plan Fluor 20�/0.5. Emitted light wasisolated by a dichroic mirror (430 nm) and a 510 nm band passfilter (Omega Optical). The wavelength of the incoming lightwas selected with the aid of a monochromator (12 nm band-width, Perkin Elmer Life Sciences) set at 340 and 380 nm. Thesewavelengths correspond to the fluorescence peaks of Ca2�-saturated and Ca2�-free Fura-2 solutions. Twelve-bit imageswere obtained using an Ultrapix 2000 Mono CCD cameracontrolled by Ultraview PC software (Fig. 2). Exposure timewas 50 msec and change in the wavelength of incoming lightfrom 340 to 380 nm is carried out in less than 5 msec. Timecourse data represent the average light intensity in a smallelliptical region within each cell.
Background and autofluorescence components were sub-tracted at each wavelength and the ratio 340/380 was calibratedinto [Ca2�] i values using Grynkyevicz’s equation (Grynkyeviczet al., 1985). The variables Rmax, Rmin , and � were calculatedin vitro from the spectra of small Fura-2 droplets in Ca2�-saturated solution (composition in mM: KCl, 100; NaCl, 10;MgCl2, 1; Tris, 10; MOPS, 10; CaCl2, 2; and Fura-2, [Molec-ular Probes] 100 �M) and Ca2�-free solution (composition inmM: KCl, 100; NaCl, 10; MgCl2, 1; Tris, 10; MOPS, 10;CaCl2, 2; and EGTA, 2), both determined empirically in oursystem (Dıaz-Hernandez et al., 2001).
For microfluorimetric studies, ATP, 2-methylthio-adenosine triphosphate (2MeSATP), 2-methylthioadenosinediphosphate (2MeSADP), UTP, UDP, glutamate, and 2�-deoxy-N6-methyladenosine 3�-5�-bisphosphate (MRS-2179)were all obtained from Sigma.
RT-PCR Experiments
From granule cell cultures maintained at 7 and 14 days invitro (DIV), total RNA was isolated using the RNAeasy system(Midi-kit; Qiagen) based on silica gel chromatography, whichemploys a lysis buffer containing guanidine isothiocyanate and�-mercaptoethanol (10 �g/ml). After isolation, RNA samples
underwent DNase digestion (RQ1 RNase-free DNase; Pro-mega, Madison, WI) for 1 hr at 37°C, and were cleaned againusing the Qiagen kit. RNA needed for semiquantitative PCRwas quantified accurately using the RiboGreen kit (RNAQuantitation Reagent, Molecular Probes), a highly sensitivefluorescent method based on the RiboGreen probe.
First-strand cDNA synthesis was carried out using theMultiScribe Reverse Transcriptase (1.25 U/ml; Taq ManRReverse Transcription Reagents, Applied Biosystems), in a re-action containing 2.5 �M oligo(dT)16, 0.5 U/�l RNase inhib-itor, 5.5 mM MgCl2, dNTP mixture (0.5 mM each), and 1 �gof purified RNA in a final volume of 25 �l. The reactionproceeded for 30 min at 48°C, followed by a 5-min step at 95°Cto achieve reverse transcriptase inactivation. To evaluate thelevel of contaminating genomic DNA, parallel RT reactions foreach RNA sample were carried out in the absence of the reversetranscriptase enzyme.
PCR reactions were carried out using 0.625 Ampli TaqGold DNA polymerase (Applied Biosystems), in the presence of1.5 mM MgCl2, dNTP mixture (0.2 mM each), specific P2Yand P2X oligonucleotide primers (300 nM each), and 1–5 �l ofthe RT product. After a 10-min hold at 95°C (required forAmpli Taq Gold DNA polymerase activation), the amplificationwas run for 40 cycles (94°C, 30 sec; 60°C, 30 sec; 72°C, 60 sec).Control reactions (H2O control) were done in the absence ofRT product (template) to avoid cross contamination.
Specific oligonucleotide primers for P2Y receptors wereobtained due to a collaboration with the group of Dr. J.M.Boeymaens (Erasme Hospital, Universite Libre de Bruxelles,Brussels, Belgium; Jimenez et al., 2000) (Table I), and primersfor P2Y12 were obtained from Jin et al. (2001). P2X subunitprimers, with specificity demonstrated by Southern blot analysis(Garcıa-Lecea et al., 2001), were designed in our laboratory(Table I), with the exception of P2X3 and P2X6, which wereobtained from Luo et al. (1999). Positive controls for P2X andP2Y gene expression were obtained with RNA from P7 ratcerebellum and cerebellar astrocyte cultures, respectively.
Semiquantitative PCR was approached by adjusting thenumber of cycles in linear phase of the amplification. For the
TABLE I. Oligonucleotide Primers for P2Y Receptors and P2X Subunits*
Forward primer Reverse primer Size
P2Y1 CATTCTTTATTTCTTGGCTGGAGAT GAGTCATTTCTTCACTCTTGGATTGTA 100 bpP2Y2 AGCATCCTCTTCCTCACCTGCATCAGC CGGGTGATCTTATACGCCATGTTGATG 514 bpP2Y4 TGGGTGTTTGGTTGGTAGTA GTCCCCCGTGAAGAGATAG 464 bpP2Y6 CGTGAGGATTTCAAGCGACTG CCAAACGACTCCACATACCA 371 bpP2Y12 CAGGTTCTCTTCCCATTGCT CAGCAATGATGATGAAAACC 674 bpP2X1 TTCCTCTATCACAAGATTCAAACAC GCAAAiCTGAAGTTGTAiCCTGG 229 bpP2X2 TTCACATTTGATCAGGACTCTGAC GCAAAiCTGAAGTTGTAiCCTGA 250 bpP2X3 TTGAGGGTAGGGGATGTGGT GCTGATAATGGTGGGGATGA 326 bpP2X4 TTCATTTACAATGCTCAAACGGAT GCAAAiCTGAAGTTGTAiCCAGG 250 bpP2X5 TTCCACTTCAGCTCCACAAATCTC GCAAAiCTGAAGTTGTAiCCAGA 232 bpP2X6 CTGTGGGATGTGGCTGACTT TCAAAGTCCCCTCCAGTCAT 484 bpP2X7 TTCACCTTTCACAAGACTTGGAAC GCAAAiCTGAAGTTGTAiCCAGG 232 bp
*P2Y receptors primer pairs were obtained from specific sequences of the different rat P2Y cDNAs: P2Y1, P2Y2, and P2Y4 from Jimenez et al. (2000);P2Y12 from Jin et al. (2001). Primer pairs for P2X subunits were designed based on specific rat P2X sequences located between the two transmembranedomains: P2X1, P2X2, P2X4, P2X5, and P2X7 from Garcıa-Lecea et al. (2001); P2X3 and P2X6 obtained from Luo et al. (1999).
386 Hervas et al.

P2Y PCR fragments, the number was around 28–32 cycleswhen using 5 �l of the RT product. In parallel PCR reactions,the housekeeping gene 18S was also amplified (Quantum RNA18S internal standards; Ambion, Austin, Texas), but only for18–20 cycles. PCR amplification products were electropho-resed on a 2% (wt/vol) agarose gel and stained with SYBR Goldprobe (Molecular Probes). Relative densities of PCR bandswere quantified by a gel imaging system (Fluo-S MultiImager;Bio-Rad, Hercules, CA).
RESULTS
Control and Characterization of Rat CerebellarGranule Neurons in Culture
Cerebellar granule neurons isolated and purifiedfrom 7- to 8-day-old Wistar rat pups were cultured asdescribed above. For the cell cultures, seeded at lowdensity and in the absence of fetal serum, the presenceof contaminating astrocytes was not detected, as shownin Figure 1B, where granule cells with DAPI-labelednuclei did not exhibit GFAP labeling. A GFAP-positivecontrol with cultured cerebellar astrocytes is shown inFigure 1A. The �6 subunit of the GABAA receptor iswell accepted as a good marker for mature cerebellargranule neurons, and the presence of this receptor sub-unit in the cultured granule cells is shown in Figure 1C(Thompson and Stephenson, 1994). The glutamatergicnature of these neurons has also been studied, combin-ing the specific vesicular neuronal marker, synaptophy-sin, with vesicular glutamate transporters, which definethe nature of the transmitter to be stored and released.The presence of the two most relevant vesicular gluta-mate transporters, VGLUT1 and VGLUT2, are shownin Figure 1E and 1H, respectively, and as occurs inmature cerebellum, VGLUT1 is present at a higherdensity. Under our culture conditions, granule cellsexhibited a pronounced neuronal morphology; the dis-tribution of glutamatergic vesicles matching the synap-tophysin labeling along an axon-like prolongation isshown under higher magnification in Figure 1J and 1K.The neuron-like morphology of cerebellar granulecells, with pronounced neurodendritic extensions, ap-peared early in the first week in culture and was welldeveloped at Day 8, when the first response studieswere carried out. The cytoarchitectural characteristicsof axodendritic prolongations become increasinglycomplex with the passing of days in culture.
For functional studies, cultured granule cells wereused from Day 8 in culture (DIV, days in vitro) to Day 20;longer culture periods result in significant loss of neuronalresponses and viability.
Nucleotide-Activated [Ca2�]i Signals in CerebellarGranule Cells
Cultured granule cells were challenged with variousnucleotides to elicit [Ca2�]i increase. Single cell responsesfollowed by microfluorimetry are shown in Figure 2. Inour culture conditions, all cells exhibited a good responseto K� ions (100%), and most cells to 100 �M glutamate
(90–95%, results not shown); however, not all cells re-sponded to ATP. The prominent calcium entrance re-sponse induced by K� ions was an additional control todemonstrate the neuronal nature of the single cell understudy. It is necessary to consider that we are only measur-ing responses to added nucleotides, not to the hydrolysisproducts (mainly adenosine) that could appear due toectonucleotidase action. The continuous superfusion ofthe cell chamber avoids concentration of any hydrolyticproduct in the extracellular medium; the same reasoningapplies to GABA and glutamate release that could beinduced by ATP action.
As demonstrated in Figure 3, there exists a largevariety of cellular responses to nucleotides in both nu-cleotide specificity and calcium response amplitude.Analysis of the most frequent response distributionshows that all nucleotide-responding cells can be stim-ulated by ATP, and their calcium responses are reducedsystematically, but not abolished, in the absence of Ca2�,such as when the ion is sequestered by the addition ofEGTA (Fig. 3A–D). This behavior corresponds with thepresence of both ionotropic and metabotropic P2 recep-tors on the same single cell under analysis. Results ob-tained with the ATP analogue 2MeSATP were similar,and 2MeSATP has similar agonistic effects on P2X recep-tors and on some of the P2Y family, such as P2Y1 andP2Y11. A different situation exists concerning the responseto uridine nucleotides. First, not all ATP-responsive gran-ule cells were able to respond to UTP and UDP. Further-more, frequency distribution and response intensity variedover the days in culture (DIV), as summarized in Table IIand Figure 4A and B. The presence of pyrimidine recep-tors, such as P2Y2, P2Y4, and most clearly P2Y6, is sup-ported firmly by the abundance of cells responding toUTP and UDP.
The number of days in culture of granule cells is arelevant factor to be considered when analyzing thenucleotide responses distribution. The number of cellsresponding to ATP, 2MeSATP, and UDP shows acontinuous increase until DIV 20, as demonstrated bythe number of cells exhibiting response pattern A and D(Table II) and Figure 4A. There is a clear parallelbetween the ATP and the 2MeSATP responses; bothpresent the same percent distribution in the cell popu-lation. Conversely, the total cells that responded toUTP were reduced markedly as time in culture pro-gressed. Even the Type B responses, in which cellsrespond to UTP but not UDP, disappeared completelyby the end of the culture period. Cells responding toUTP or UDP and K�, but not to ATP, were very rarelyobserved (�0.5%).
The increase in the [Ca2�]i response was anotherrelevant aspect of the analysis of calcium responses ingranule cells. Notably, the response magnitude increasedsignificantly as time in culture progressed for all nucleo-tides studied, and responses were maximal at the end of theculture period (Fig. 4B).
P2X and P2Y Receptors in Granule Neurons 387

Fig. 1. Identification by immunolabeling of cultured granule cells.A: Fluorescence image of cerebellar astrocytes labeled with rabbitanti-GFAP antibody, and tagged with goat anti-rabbit IgG rhodamineconjugate. Nuclei are marked with DAPI. B: Typical granule cellculture (17 DIV), showing negative staining with rabbit anti-GFAPantibody, and nuclei labeled with DAPI. C: Granule cell culture (17DIV) presents positive labeling with goat anti-�6 GABAAR, taggedwith donkey anti-goat IgG fluorescein conjugate. D: Granule cells (14DIV) labeled with mouse anti-synaptophysin antibody, and tagged withgoat anti-mouse IgG fluorescein conjugate. E: The same field as in (E)
labeled with rabbit anti-VGLUT1, and tagged with goat anti-rabbitIgG rhodamine conjugate. F: The same field as in (D,E) marked withDAPI. G: Another view of cerebellar granule cells (14 DIV) labeledwith synaptophysin as in (D). H: The same field presented in (G)labeled with rabbit anti-VGLUT2, and tagged with goat anti-rabbitIgG rhodamine conjugate. I: The same field as in (G,H) marked withDAPI. J: Amplified image of synaptophysin-labeled cerebellar cellaxodendritic prolongation (17 DIV). K: The same field presented in (J)labeled with rabbit anti-VGLUT1, as in (E). Scale bar � 10 �m (A–I)and 2 �m (J,K).

In the presence of EGTA, individual granule cellresponses to 2MeSADP confirmed the responses throughP2Y1 or other receptors responding to ADP and deriva-tives in cultured granule cells (Fig. 5A,B). A high percent-
age of cells (almost 60%) exhibited reduced calcium re-sponse with the addition of the specific P2Y1 antagonist,MRS-2179 (10 �M), thus confirming the presence offunctional P2Y1 receptors. The calcium responses of the
Fig. 2. Single cerebellar granule cells responses to ATP and KCl.A: View-field image of cerebellar granule cells (14 DIV) loaded withFURA-2 dye. Fluorescence at 380 nm in grayscale color is presented.Left: basal nonstimulated field; right: in the presence of 100 �M ATP.B: The sequence of images show an amplified field from A, corre-sponding to the selected square showing two cerebellar granule cells.Cell in the right is an ATP-responsive cell (both are responding to K�).
Both (A) and (B) correspond to fluorescence emission (510 nm) afterexcitation at 380 nm. C: Time-course of fluorescence changes recordedfrom the ATP-responsive cell. Agonist was challenged for 30 sec asindicated by the solid horizontal bars. Upper panel represents the ratiocalculated as the quotient F340/F380. Lower panel: Record showingcalcium increase after calibration according to Grynkyevicz’s equation.
P2X and P2Y Receptors in Granule Neurons 389

remaining 40% of cells were affected to a more limitedextent, if at all, by the presence of the inhibitor (Fig. 5B).The results seem to suggest that ADP-sensitive receptorswith inducible calcium responses, other than P2Y1 recep-
tors, are also present in cultured granule cells; the identityof these other receptors needs to be defined. Although theP2Y12 receptor has been described as being coupledmainly to Gi proteins and inhibition of the cAMP path-
Fig. 3. Typical calcium responses to nucleotides in cerebellar granulecells. Cells were stimulated sequentially with 100 �M ATP, 100 �M2MeSATP, 100 �M UTP, and 100 �M UDP. The nucleotides ATPand 2MeSATP were also tested in EGTA-buffered medium containing100 nM free Ca2�. Agonists were applied by superfusion for timeindicated by solid bars, after which cells were allowed to rest for at least5 min between consecutive challenges. Cells were always challenged
with KCl (30 mM) at the end of the experiment (as shown in Fig. 2);only cells showing healthy responses to KCl were selected. Recordingswere interrupted between stimulations to avoid excessive dye photo-bleaching. Each single cell record, (A–D), is representative of cellpopulations named (A–D), which exhibit characteristic expression pat-terns in response to nucleotides. These four populations were found inall studied cultures.
390 Hervas et al.

way, the possibility of coupling to other effector systemscannot be excluded.
P2Y Receptor Expression in Cultured CerebellarGranule Neurons
The responses found to various types of nucleotidicagonists was indicative of a great variety of ionotropic and
metabotropic receptors, and RT-PCR approach was em-ployed to assess specific nucleotidic receptor expression incultured granule neurons.
RT-PCR was carried out at two differentiationstages: 7 DIV, when granule cells have matured and ex-hibit a pronounced neuron-like morphology, as shown byGABAA �6 subunit-positive immunolabeling; and 14DIV, when responses to ATP and nucleotides becomeapparent. As shown in Figure 6A, significant levels ofexpression are found for P2Y1, P2Y4, P2Y6, and P2Y12. Apositive control for P2Y receptor expression was obtainedfrom purified P7 rat cerebellar astrocyte cultures, in whichdifferent P2Y receptors (P2Y1, P2Y2, P2Y4, and P2Y6)have been characterized pharmacologically and function-ally in a previous work (Jimenez et al., 2000). Astrocytesexhibited similar expression levels of different P2Y recep-tors, with the low band intensity corresponding to theP2Y2 amplification product (Fig. 6A), which indicates thatgranule neurons seem to lack P2Y2 receptors. Our cultureswere based on low-density, highly purified granule neu-rons, and the addition of AraC assured the absence ofproliferating glial cells. Thus, the low level of P2Y2 re-ceptor expression observed could be attributed to a smallcontribution of contaminating astrocytes in the culture.These results indicate that complete control of purity ofgranular neuron cultures is required for a precise evalua-tion of receptors.
Figure 6A also confirms the presence of P2Y12,which belongs to the family of ADP-responsive receptors.The presence of P2Y12 could explain the lack of MRS-2179-induced inhibition observed in 40% of the cellsresponding to 2MeSADP (Fig. 5); this possibility will beanalyzed in the discussion.
A growing number of cells responding to UDP wereobserved by microfluorimetry, as was an increase in re-sponse magnitude as the time in culture progressed. Thiscorrelates well with changes at the level of P2Y 6 expres-sion, as seen in the semiquantitative approach, in whichthe P2Y6 messenger level increased by a factor of 1.52 0.08 between 7 and 14 DIV (Fig. 6B).
P2X Receptor Expression in Cultured CerebellarGranule Neurons
A significant percentage of the total [Ca2�]i re-sponses to ATP in granule cells is due to ionotropicresponses. The presence of ionotropic nucleotide recep-tors was confirmed by the expression of different P2Xsubunits, such as P2X1, P2X2, P2X3, P2X4, P2X6, andP2X7, in purified granule cultures, as shown in Figure 7.The exception seems to be the P2X5 subunit, whoseexpression levels seemed not relevant, and is in agreementwith that observed in P7 whole rat cerebellum, which istaken as a positive control of P2X subunit expression(Garcıa-Lecea 2001). The contribution of P2X subunitexpression in astrocytes to total P2X expression in granulecells cultures is expected to be insignificant, as is thepresence of astrocytes in these cultures. In addition, thephysiological significance of P2X subunits in astrocytesremains under discussion, as no ionotropic responses to
TABLE II. Distribution of Nucleotide [Ca2�]i Responses inCultured Cerebellar Granule Cells*
13 DIV (%) 17 DIV (%) 21 DIV (%)
Type A 25 35.1 18.3Type B 25 19.3 0Type C 18.7 17.5 45Type D 31.3 28.1 36.7
*The table shows the percentage of each response type shown in Figure 3.A total of 372 cells from four cultures have been tested.
Fig. 4. Nucleotide responses evolution in cerebellar granule cells withthe days in culture. Granule cells were stimulated with 100 �M ATP,100 �M 2MeSATP, 100 �M UTP, and 100 �M UDP. [Ca2�]iincreases were calculated as described in the methods. A: Percentage ofcells responding to various nucleotides. B: Corresponding intracellularcalcium increases. Values represent the means SEM of four experi-ments in duplicate (372 cells in total were tested)
P2X and P2Y Receptors in Granule Neurons 391

nucleotides have been observed in previous studies (Jime-nez et al., 2000).
Immunocytochemical studies confirmed the pres-ence of several P2X subunits in granule neurons (Figs.8,9), except for the P2X6 subunit. P2X6 subunit expres-sion was clearly detectable at the messenger level, but notby immunolabeling. In addition, all granule cells exhibiteda marked synaptophysin labeling, in agreement with theirneural nature, which gives key information about P2Xsubunit localization throughout granule cell architecture.
Great heterogeneity was found in P2X subunit pres-ence and distribution in granule neurons. P2X1 labeling
was restricted to granule cell axodendritic prolongations,and presented a diffuse, uniform staining pattern that wasdistributed homogeneously along cell extensions (Fig. 8A–C). In contrast, P2X2, P2X3, and P2X4 immunolabelingwere present at somatodendritic locations (Fig. 8D,G,J),although staining for P2X2 and P2X4 does not occur ingeneral for the whole granule cell population. Moreover,whereas P2X2 seemed to be present in some dendritic-likestructures, P2X3 was distributed widely and discontinu-ously along the majority of cellular processes, and showeda great degree of colocalization with synaptophysin (Fig.8I). Figure 9 shows P2X7 subunit labeling that exhibits a
Fig. 5. Effect of the P2Y1 antagonist MRS-2179 on 2MeSADP-evoked [Ca2�]i increases. A: MRS-2179 was introduced 2 min beforechallenge with 2MeSADP. Both compounds were tested at 10�M in afree-Ca2� medium. MRS-2179 almost completely inhibits the [Ca2�]i
increase elicited by 2MeSADP in 60% of cerebellar granule cells (atypical record is shown). B: Typical record presented for 40% ofgranule cells that were insensitive to MRS-2179. Distribution percent-age corresponds to analysis of 200 cells from four separate experiments.
392 Hervas et al.

specific punctuated pattern with the most significant co-localization with synaptophysin, as indicated in the ampli-fied view in Figure 9D–F.
DISCUSSIONThe present study demonstrates the existence of both
ionotropic and metabotropic functional nucleotidic recep-tors in rat cerebellar granule cells in culture, and therelation to days in vitro. Due to the abundance of metabo-tropic responses to purine and pyrimidine nucleotides incerebellar astrocytes, and their almost ubiquitous presencein cultures of cerebellar origin, an extensive effort wasmade to completely avoid their presence in our neuronalcultures (Jimenez et al., 1999, 2000, 2002). The absence ofa control in this regard results in difficult-to-reproduce,nonreliable data. In our culture conditions, no cells ex-hibited the glial marker protein GFAP, but all contained
the neuronal vesicular marker synaptophysin. Moreover,as mature granular cells, all exhibited a glutamatergic na-ture and were labeled with antibody to the vesicularglutamate transporter, VGLUT-1, and to a lesser extentwith antibody to the VGLUT-2 (Herzog et al., 2001). Itis to highlight the exact matching between both vesicularmembrane proteins, the VGLUT-1 and synaptophysin,always observed in our double immunolabeling, as alsotheir presence in well developed axon-like structures aftersome days in culture. This distribution contrast to that ofthe GABAA receptor subunit, �6, which has been de-scribed as a granule cell maturation marker, and exhibits abroad and preferential presence on granule cell somas(Thompson and Stephenson, 1994; Hevers and Ludens,2002).
Under the specific quality control conditions main-tained, all granule cells responded to K� ions and mostresponded to glutamate, which is an additional control forneuronal and granular nature, respectively. In contrast, thecultured granule cells exhibited a large variability in re-sponses to nucleotides over the days in culture. Theybegan to give significant and functional responses to nu-cleotides only after the second week; however, mRNA fornucleotidic receptors was detected earlier, perhaps preced-ing the functional expression of the receptor’s protein.Single-cell microfluorimetric techniques reported here al-lowed us to demonstrate that heterogeneity was the prin-cipal characteristic of granule cell response to nucleotides,
Fig. 6. Expression of P2Y receptors in purified cerebellar granule cellcultures. A: Typical RT-PCR carried out with three different cultures.Granule cell cultures maintained at 7 and 14 DIV were analyzed forP2Y receptor expression. The expression pattern of each P2Y receptoris shown in individual panels. Besides the H2O control, carried out inthe absence of template, additional negative controls were carried out inthe absence of reverse transcriptase reaction (RT) for each RNAsample and each primer pair. Only the RT control for RNA obtainedat 7 DIV is shown. M, 100-bp DNA ladder. P2Y receptor expressionin cerebellar astrocytes (Ast) was used as positive control for P2Y primerspecificity. B: Representative gel of amplified products obtained forP2Y6 and 18S proteins at two different culture stages, 7 and 14 DIV.Amplification was carried out for 32 and 20 cycles, for P2Y6 and 18S,respectively, when using 5 �l of the RT reaction. Data obtained at eachculture period were normalized with respect to the amplification of the18S protein, and shown as the expression ratio 14 DIV/7 DIV. Data arethe means SD of three experiments carried out from three differentcultures.
Fig. 7. Expression of different P2X subunits in purified cerebellargranule cell cultures. Typical RT-PCR results obtained from threedifferent cultures. As above, P2X subunit expression was analyzed incultures at 7 and 14 DIV. Only results obtained at 14 DIV are shown.Negative controls (H2O control, RT control) were the same as thosefor P2Y receptors. Controls for primer efficiency and specificity foreach P2X product were obtained from P7 whole rat cerebellum RNA.GN, granule neuron; M, 100-bp DNA ladder.
P2X and P2Y Receptors in Granule Neurons 393
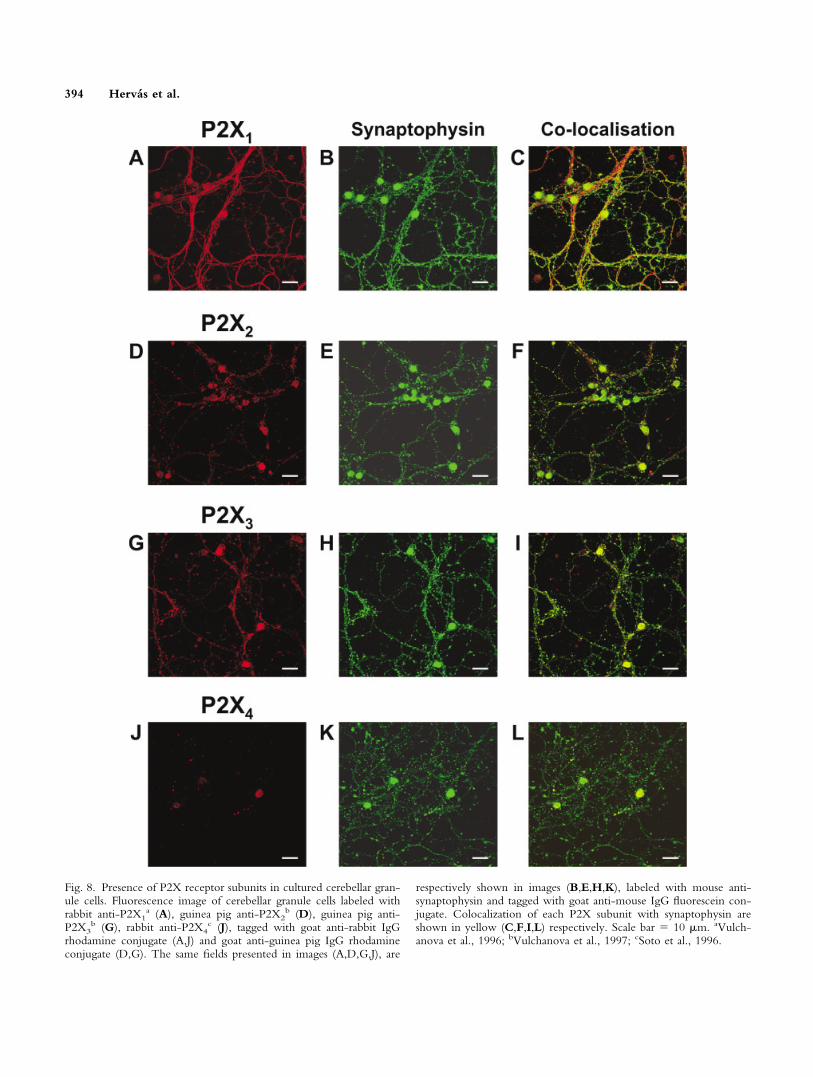
Fig. 8. Presence of P2X receptor subunits in cultured cerebellar gran-ule cells. Fluorescence image of cerebellar granule cells labeled withrabbit anti-P2X1
a (A), guinea pig anti-P2X2b (D), guinea pig anti-
P2X3b (G), rabbit anti-P2X4
c (J), tagged with goat anti-rabbit IgGrhodamine conjugate (A,J) and goat anti-guinea pig IgG rhodamineconjugate (D,G). The same fields presented in images (A,D,G,J), are
respectively shown in images (B,E,H,K), labeled with mouse anti-synaptophysin and tagged with goat anti-mouse IgG fluorescein con-jugate. Colocalization of each P2X subunit with synaptophysin areshown in yellow (C,F,I,L) respectively. Scale bar � 10 �m. aVulch-anova et al., 1996; bVulchanova et al., 1997; cSoto et al., 1996.
394 Hervas et al.

showing that different cell populations express differentP2X and P2Y receptors. Indeed, major differences arefound with respect to the P2Y receptor type, enablingclassification of cell populations with respect to pyrimidineresponses.
Metabotropic responses characteristic of the presenceof a P2Y2/P2Y4 receptor type were clear. In contrast to aprevious work (Amadio et al., 2002), however, P2Y2expression was not found in our culture conditions, anapparent discordance that can be explained by the absenceof contaminating astrocytes, which would account forP2Y2 expression. This assumption is based on previouswork by our group, which demonstrated that Type Icerebellar astrocytes coexpressed different types of P2Yreceptors (Jimenez et al., 1999). Moreover, consideringthat P2Y2 and P2Y4 were present to a similar extent incerebellar astrocytes, the nondetection of P2Y2 in granule
cells argues against the idea that P2Y2 and P2Y4, bothresponding to ATP and UTP, were the same receptor, ashas been suggested previously.
Interestingly, the UTP response pattern in granulecells decreases gradually with time in culture, completelydisappearing at the end of the culture period. This couldbe explained by changes at the level of P2Y4 receptorexpression, as was suggested to be responsible for UTPresponses. The opposite is observed for other pyrimidinereceptors present in granule cells, as is the case for P2Y6,the expression of which is upregulated during culturemaintenance. Indeed, UDP responses seem to be a delayedevent linked to granule cell maturation. In the same neu-ronal model, other responses of a metabotropic naturehave also been reported to become modified, such as thoseof metabotropic glutamate and adenosine A1 receptors(Kirischuk et al., 1996; Sanz et al., 1996).
Fig. 9. Presence of P2X7 receptor subunits in cultured cerebellargranule cells. A: Fluorescence image of cerebellar granule cells labeledwith rabbit anti-P2X7 and tagged with secondary antibody goat anti-rabbit IgG rhodamine conjugate. B: The same field as in A labeled withmouse anti-synaptophysin and tagged with goat anti-mouse IgG fluo-rescein conjugate. C: Colocalization of P2X7 subunit and synaptophy-sin marked in yellow. D: Amplified image of cerebellar granule cell
axodendritic prolongation labeled with rabbit anti-P2X 7 as in A.E: Same field as in D marked with synaptophysin antibody. F: Colo-calization of P2X7 subunit and synaptophysin. Specificity of P2X7
antibody was tested by Western blot, employing different proteinsreconstituted by in vitro transcription-translation from different ratP2X cDNAs. Scale bar � 10 �m (A–C) and 2 �m (D and F).
P2X and P2Y Receptors in Granule Neurons 395

These results indicate that P2Y receptor expression ispossibly linked to several important growth and differen-tiation signals in granule neurons. Whether these signalsare triggered early, as pre-programmed factors, before theculture is established, or whether the signals appear underthe conditions of culture, remains to be investigated. Inthis respect, developmental expression of GABAA �6 re-ceptor subunits is an event initiated by an environmentalfactor received by granule neurons in an early develop-mental stage (Thompson et al., 1996). Another example isthe GABAA �2 receptor subunit expression, which hasbeen described as dependent on the presence of a specificneuregulin, Ig-NRG (Rieff et al., 1999). On the otherhand, NMDA receptors are downregulated in granuleneurons by other endogenously produced factors involvedin neurite outgrowth, such as brain-derived neurotrophicfactor (Brandoli et al., 1998).
In regard to purine metabotropic nucleotide recep-tors, the major population of granule cells is equally re-sponsive to ATP and 2MeSATP in the absence of externalCa2�. Considering that ATP is P2Y1 and P2Y2 receptoragonist and that 2MeSATP is more specific for P2Y1, butnot for P2Y2, these results confirm the absence in granuleneurons of P2Y2 receptors, a receptor that exhibits thesame affinity for ATP and UTP. In addition, this is evi-dence that UTP responses in these cells can be attributeddefinitively to P2Y4 receptor activation. Moreover, resultsobtained with the specific antagonist MRS-2179 confirmthe predominant presence of P2Y1 receptors in granuleneurons. In agreement with this, P2Y1-specific messengerwas observed in our cultures, and P2Y1 protein localiza-tion at the granular and molecular cerebellum layers hasbeen reported by others (Moore et al., 2000; Morann-Jimenez and Matute, 2000).
Part of the ATP-responsive cell population also re-sponds to 2MeSADP in the absence of extracellular cal-cium. This indicates that ADP receptors, including P2Y1and others, could play an important role in granule neu-rons. Indeed, the fact that in 40% of these 2MeSADP-responding cells, calcium increase is unaffected by theP2Y1-specific antagonist suggests the presence of an ADP-specific receptor type other than P2Y1. This other recep-tor type could possibly be P2Y12 or P2Y13, or an ADPreceptor family member that has not yet been character-ized (Hollopeter et al., 2000; Communi et al., 2001). Thedifferential characteristic of P2Y12 and P2Y13 is that al-though they are both coupled to adenylate cyclase inhi-bition via Gi protein, to date only P2Y13 has been re-ported to induce a rise in [Ca2�]i. P2Y13 has not beencloned yet from rat, however, and the finding of P2Y12receptor-specific mRNA in our cultures suggests thatP2Y12 is the candidate for this type of behavior in granuleneurons, as has been proposed for the receptor found incells of neural origin, the PC12 cells (Uterberger et al.,2002). These considerations suggest that, in granule neu-rons, P2Y12 receptors could be linked to alternative sig-naling mechanisms.
The coexistence of ionotropic and metabotropic re-ceptors of the same neurotransmitter is a generalized eventin neural cells and has been described for glutamate andGABA receptors in cerebellar granule neurons (Courtneyet al., 1990; Huston et al., 1995; Hevers and Luddens,2002). We have demonstrated here that this occurs also fornucleotide receptors in this cellular model, similar to thatobserved previously in neurochromaffin cells (Castro etal., 1995). Expression of several P2X subunits, namelyP2X1, P2X2, P2X3, P2X4, and P2X7, which can be re-sponsible for the observed ionotropic nucleotidic re-sponses in granule neurons, was confirmed by RT-PCRand immunolabeling studies, and peculiar topological lo-calization was exhibited. The abundant presence of P2X2and P2X3 subunits at the cell somas is notable and couldaccount for ionotropic nucleotide responses observedmainly at this level. At the axodendritic-like structures, thepresence of P2X1, P2X3, and P2X7 is prominent. More-over, although P2X1 exhibits a diffuse and extended dis-tribution across the plasma membrane of granular cellprocesses, the clear punctuated pattern of P2X3 and P2X7indicates specialized localization.
P2X subunits P2X4 and P2X6 seem to contribute, toa minor extent, to ATP ionotropic responses in granulecell neurons. Although appreciable at the messenger level,no specific P2X6 immunolabeling was observed, and forP2X4, it seemed restricted to a minor population of cells.When these studies were carried out in slices from maturecerebellum, however, specific and intense labeling forP2X4 was seen in granule cell somas (Rubio and Soto,2001), emphasizing the difficulty in comparing evolutionof dissociated cells in culture with intact mature tissue.The restricted expression of P2X6 subunits in Purkinjeneurons was also reported (Rubio and Soto, 2001). Inagreement with that, studies we carried out in individualPurkinje cells of neonatal rat have demonstrated that phar-macologic responses to nucleotides were typical of aP2X2/6 heterodimer (Mateo et al., 1998; Garcıa-Lecea etal., 1999), which indicates a different pattern of expressionand functionality of P2X subunits in Purkinje cells com-pared to that in granule neurons. In addition, this expres-sion pattern could change during cerebellum developmentand be replaced by a P2X 4/6 heterodimer in adult animals,in accordance with a quantitative study that shows that, inthe adult whole cerebellum, P2X2 subunit expression de-creases and P2X4 expression becomes significantly in-creased (Garcıa-Lecea, 2001).
A final discussion point is the remarkable codistribu-tion of P2X3 and P2X7 subunits with synaptophysin, aprotein of synaptic vesicle membranes, which suggests apresynaptic location of these subunits in cerebellar syn-apses. Due to the extreme thinness of granule cell axo-dendritic prolongations, however, it is difficult to accu-rately measure Ca2� increases when challenging withATP. Thus, P2X3 and P2X7 function in this subcellularlocation of cultured granule cells remains to be demon-strated. Interestingly, although P2X3 subunits were com-monly thought to be restricted to peripheral sensory neu-
396 Hervas et al.

rons, where important actions have been described, thishypothesis recently has begun to change, because P2X3subunits have been detected in sympathetic thoracolumbarneurons, and neurons from hypothalamic supraoptic andsolitary tractus nuclei in the CNS (Norenberg and Illes,2000). The restricted expression in some brain areas, andits clear presynaptic locations have probably been amongthe reasons for previous failure to detect this subunit inCNS using classic techniques, immunohistochemistry orin situ hybridization. In addition, the specialization ofP2X3 at the presynapse together with P2X7 opens manyfuture possibilities for study of the role of these subunits inmodulation of synaptic secretion. In this respect, P2X3 andP2X7 are the most abundant subunits in presynaptic ter-minals at the CNS, and their function has been demon-strated by microfluorimetry in individual isolated synap-tosomes (Dıaz-Hernandez et al., 2000; Gomez-Villafuertes et al., 2001). Previous studies in cerebellarsynaptosomes showed a high percentage responding toATP and other nucleotides by Ca2� influx (Hervas et al.,2002). In midbrain synaptic terminals, the abundant pres-ence of P2X3 and P2X7 has been described, as has theirability to induce exocytotic release of neurotransmittersstored in secretory vesicles, such as GABA, acetylcholine,or glutamate (Dıaz-Hernandez et al., 2000; Gomez-Villafuertes et al., 2001). Activation of presynaptic P2Xreceptors has been demonstrated to facilitate glutamaterelease in hippocampal neurons (Inoue et al., 1992) and inthe nucleus tractus solitarii (Kato and Shigetomi, 2001);P2X7 is also present at presynaptic locations (Deuchars etal., 2001) and is involved in depression of mossyfiber-CA3 synaptic transmission (Amstrong et al., 2002).In addition, P2X presynaptic receptors seem to be underthe control of metabotropic receptors, most notablyGABAB and adenosine A1 and A2A (Dıaz-Hernandez etal., 2002; Gomez-Villafuertes et al., 2003).
Presynaptic ionotropic receptors and the controlof transmitter release are expanding issues regardingneurosecretion and neuromodulation. Colocalization ofionotropic receptors for different neurotransmitters atthe same presynaptic area has now been reported, as isthe case for nicotinic and nucleotidic P2X receptors(MacDermott et al., 1999; Dıaz-Hernandez et al., 2000;Khakh and Henderson, 2000). Due to the abundance ofNMDA and GABAA receptors, among others, in cul-tured and mature granule neurons, questions arise re-garding their reciprocal interaction with P2X3 andP2X7 nucleotidic receptor-containing subunits. Thecolocalization of synaptophysin with P2X3 and P2X7subunits in granular cell axodendritic terminals high-lights the relevance of this neural model in understand-ing presynaptic organization and functionality. In con-clusion, our results demonstrate the heterogeneity andabundance of nucleotide responses in cultured granuleneurons. Additional work will be required to approachin vivo studies with native granule neurons to fullyunderstand the physiological role of nucleotidic recep-tors. In this respect, a recent in vivo study has reported
significant upregulation of nucleotidic P2X1 and P2X2receptors after cerebellar lesion (Florenzano et al.,2002), evidencing a key role of nucleotide receptors inthe recovery of damaged neurons of the CNS.
ACKNOWLEDGMENTSWe thank S. Pons and I. Torres-Aleman (Instituto
Cajal, Madrid) for assistance in the preparation of granulecell cultures. We also thank F. Soto (Max-Planck Institutefor Experimental Medicine, Gottingen, Germany) and M.Voight (St. Louis University School of Medicine, St.Louis, Missouri) for their kind gift of P2X antibodies. C.Hervas holds a predoctoral fellowship from ComunidadAutonoma de Madrid.
REFERENCESAbbracchio M, Burnstock G. 1994. Purinoceptors: are there families of
P2X and P2Y purinoceptors? Pharmacol Ther 64:445– 475.Amadio S, Dambrossi N, Cavaliere F, Murra B, Sancesario G, Bernardi
G, Burnstock G, Volonte C. 2002. P2 receptor modulation andcytotoxic function in cultured CNS neurons. Neuropharmacology42:489 – 450.
Amstrong JN, Brust TB, Lewis RG, MacVicar BA. 2002. Activation ofpresynaptic P2X7-like receptors depresses mossy fiber-CA3 synaptictransmission through p38 mitogen-activated protein kinase. J Neurosci22:5938–5945.
Benham CD, Tsien RW. 1987. A novel receptor-operated Ca2�-permeable channel activated by ATP in smooth muscle. Nature 328:275–278.
Brandoli C, Sanna A, De Benardi MA, Follesa P, Broker G, Mocchetti I.1998. Brain-derived neurotrophic factor and basic fibroblast growth factordownregulate NMDA receptor function in cerebellar granule cells.J Neurosci 18:7953–7961.
Castro E, Mateo J, Tome AR, Barbosa RM, Miras-Portugal MT, RosarioLM. 1995. Cell specific puzinergic receptors coupled Ca2� entry andCa2� release from internal stores in adrenal chromaffin cells. J Biol Chem270:5098–5106.
Collo G, Neidhart S, Kawashima E, Kosco-Vilbois M, North RA, Buell G.1997. Tissue distribution of the P2X7 receptor. Neuropharmacology36:1277–1283.
Collo G, North RA, Kawashima E, Merlo-Pich E, Neidhart S, SurprenantA, Buell G. 1996. Cloning of P2X5 and P2X6 receptors and the distri-bution and properties of an extended family of ATP-gated ion channels.J Neurosci 16:2495–2507.
Communi D, Gonzalez NS, Detheux M, Brezillon S, Lannoy V, Parmen-tier M, Boeynaems JM. 2001. Identification of a novel human ADPreceptor coupled to G(i). J Biol Chem 276:41479–41485.
Courtney MJ, Lambert JJ, Nicholls DG. 1990. The interactions betweenplasma membrane depolarization and glutamate receptor activation in theregulation of cytoplasmic free calcium in cultured cerebellar granule cells.J Neurosci 10:3873033 –3879.
Deuchars SA, Atkinson L, Brooke RE, Musa H, Milligan CJ, Batten TFC,Buckley NJ, Parson SH, Deuchars J. 2001. Neuronal P2X7 receptors aretargeted to presynaptic terminals in the central and peripheral nervoussystem. J Neurosci 21:7143033 –7152.
Dıaz-Hernandez M, Pintor J, Castro E, Miras-Portugal MT. 2001. Inde-pendent receptors for diadenosine pentaphospate and ATP in rat midbrainsingle synaptic terminals. Eur J Neurosci 14:918–926.
Dıaz-Hernandez M, Pintor J, Castro E, Miras-Portugal MT. 2002.Co-localisation of functional nicotinic and ionotropic nucleotidicreceptors in isolated cholinergic synaptic terminals. Neuropharmacol-ogy 42:20 –33.
P2X and P2Y Receptors in Granule Neurons 397

Dıaz-Hernandez M, Pintor J, Miras-Portugal MT. 2000. Modulation of thedinucleotide receptor present in rat midbrain synaptosomes by adenosineand ATP. Br J Pharmacol 130:434–440.
Edwards FA, Gibb AJ, Colquhoun D. 1992. ATP receptor-mediated syn-aptic currents in the central nervous system. Nature 359:144–147.
Florenzano F, Viscomi MT, Cavaliere F, Volonte C, Molinari, M. 2002.Cerebellar lesion up-regulates P2X1 and P2X2 purinergic receptors inprecerebellar nuclei. Neuroscience 115:425–434.
Gallo V, Kingsbury A, Balazs R, Jorgensen OS. 1987. The role of depo-larization in the survival and differentiation of cerebellar granule cells inculture. J Neurosci 7:2203–2213.
Garcıa-Lecea M, Delicado EG, Miras-Portugal MT, Castro E. 1999. P2X2characteristics of the ATP receptor coupled to Ca2� increase in culturedPurkinje neurons from neonatal rat cerebellum. Neuropharmacology38:699–706.
Garcıa-Lecea M, Sen RP, Soto F, Miras-Portugal MT, Castro E. 2001. P2receptors in cerebellar neurons: molecular diversity of ionotropic ATPreceptors in Purkinje cells. Drug Dev Res 52:104–113.
Garcıa-Lecea M. 2001. Receptors P2X en neuronas de purkinje de cer-ebelo de rata neonatal. Tesis Doctoral. Universidad Complutense deMadrid. Spain.
Gomez-Villafuertes R, Gualix J, Miras-Portugal MT. 2001. SingleGABAergic synaptic terminals from rat midbrain exhibit functional P2Xand dinucleotide receptor, able to induce GABA secretion. J Neurochem77:84–93.
Gomez-Villafuertes R, Pintor J, Gualix J, Miras-Portugal MT. 2003.GABAB receptor-mediated presynaptic potentiation of ATP ionotropicreceptors in rat midbrain synaptosomes. Neuropharmacology 44:311–323.
Grynkyewicz G, Poenie M, Tsien RY. 1985. A new generation of Ca2�indicators with greatly improved fluorescence properties. J Biol Chem260:3440–3450.
Hervas C, Dıaz-Hernandez M, Gomez-Villafuertes R, Miras-Portugal MT.2002. Presencia de receptores de ATP y Ap5A en sinaptosomas decerebelo de rata. XXV Congreso de la SEBBM.
Herzog E, Bellenchi GC, Gras C, Bernard V, Ravassard P, Bedet C,Gasnier B, Giros B, El Mestikawy S. 2001. The existence of a secondvesicular glutamate transporter specifies subpopulations of glutamatergicneurons. J Neurosci 21:1–6.
Hevers W, Luddens. 2002. Pharmacological heterogeneity of �-aminobutyricacid receptors during development suggests distinct classes of rat cerebellargranule cells in situ. Neuropharmacology 42:34–47.
Hollopeter G, Jantzen HM, Vincent D, Li G, England L, Ramakrishnan V,Yang RB, Nurden P, Nurden A, Julius D, Conley PB. 2001. Identifica-tion of the platelet ADP receptor targeted by antithrombotic drugs.Nature 409:202–207.
Huston E, Cullen GP, Burley JR, Dolphin AC. 1995. The involvement ofmultiple calcium channel sub-types in glutamate release from cerebellargranule cells and its modulation by GABAB receptor activation. Neuro-science 68:465–478.
Inoue K, Nakazawa K, Fujimori K, Watano T, Takanaka A. 1992. Extra-cellular adenosine 5�-triphosphate-evoked glutamate release in culturedhippocampal neurons. Neurosci Lett 134:215–218.
Jimenez AI, Castro E, Communi D, Boeynaems JM, Delicado EG, Miras-Portugal MT. 2000. Coexpression of several types of metabotropic nu-cleotide receptors in single cerebellar astrocytes. J Neurochem 71:2071–2079.
Jimenez AI, Castro E, Delicado EG, Miras-Portugal MT. 2002. Specificdiadenosine pentaphosphate receptor coupled to extracellular regulatedkinases in cerebellar astrocytes. J Neurochem 83:299–308.
Jimenez AI, Castro E, Maribet M, Franco R, Delicado EG, Miras-PortugalMT. 1999. Potentiation of ATP calcium responses by A2b receptorstimulation and other signals coupled to Gs proteins in type 1 cerebellarastrocytes. Glia 26:119–128.
Jin J, Tomlinson W, Kirk IP, Kim YB, Humphries RG, Kunapuli SP. 2001.The C6-2B glioma cell P2Y(AC) receptor is pharmacologically andmolecularly identical to the platelet P2Y(12) receptor. Br J Pharmacol133:521–528.
Kanjhan R, Housley GD, Thorne PR, Christie DL, Palmer DJ, Luo L,Ryan AF. 1996. Localization of ATP-gated ion channels in cerebellumusing P2X2R subunit-specific antisera. Neuroreport 7:2665–2669.
Kato F, Shigetomi E. 2001. Distinct modulation of evoked and spontaneousEPSCs by purinoceptors in the nucleus tractus solitarii of the rat. J Physiol530:469–86.
Khakh BS, Henderson G. 2000. Modulation of fast synaptic transmission byligand-gated ion channels. J Auton Nerv Syst 81:110–121.
Kirischuk S, Voitenko N, Kostyuk P, Verkhratsky A. 1996. Calciumsignalling in granule neurones studied in cerebellar slices. Cell Calcium19:59–71.
Loesch A, Burnstock G. 1998. Electron-immunocytochemical localizationof P2X1 receptor in the rat cerebellum. Cell Tissue Res 294:253–260.
Luo X, Zheng W, Yan M, Lee MG, Muallem S. 1999. Multiple functionalP2X and P2Y receptors in the luminal and basolateral membranes ofpancreatic duct cells. Am J Physiol 277:205–215.
Lundy PM, Hamilton MG, Mi L, Gong W, Vair C, Sawyer TW, Frew R.2002. Stimulation of Ca(2�) influx through ATP receptors on rat brainsynaptosomes: identification of functional P2X(7) receptor subtypes. Br JPharmacol 135:1616–1626.
MacDermott AB, Role LW, Siegelbaum SA. 1999. Presynaptic ionotropicreceptors and the control of transmitter release. Annu Rev Neurosci22:443–485.
Mateo J, Garcıa-Lecea M, Miras-Portugal MT, Castro E. 1998. Ca2�signals mediated by P2X-type purinoceptors in cultured cerebellar Pur-kinje cells. J Neurosci 18:1704–1712.
Meyer-Franke A, Kaplan MR, Pfrieger FW, Barres BA. 1995. Character-ization of the signaling interactions that promote the survival and growthof developing retinal ganglion cells in culture. Neuron 15:805–819.
Moore D, Chambers J, Waldvogel H, Faull R, Emson P. 2000. Regionaland cellular distribution of the P2Y(1) purinergic receptor in the humanbrain: striking neuronal localisation. J Comp Neurol 421:374–384.
Moran-Jimenez MJ and Matute C. 2000. Immunohistochemical localiza-tion of the P2Y1 purinergic receptor in neurons and glial cells of thecentral nervous system. Brain Res Mol Brain Res 78:50–58.
Norenberg W, Illes P. 2000. Neuronal P2X receptors: localisation andfunctional properties. Naunyn Schmiedebergs Arch Pharmacol 362:324–339.
North A. 2002. Molecular physiology of P2X receptors. Physiol Rev82:1013–1067.
Pankratov Y, Castro E, Miras-Portugal MT, Krishtal O. 1998. A purinergiccomponent of the excitatory postsynaptic current mediated by P2Xreceptors in the CA1 neurons of the rat hippocampus. Eur J Neurosci10:3898–3902.
Pintor J, Miras-Portugal MT. 1995. A novel receptor for diadenosinepolyphosphates coupled to calcium increase in rat midbrain synaptosomes.Br J Pharmacol 115:895–902.
Pons S, Trejo JL, Martınez-Morales JR, Martı E. 2001. Vitronectin regu-lates Sonic hedgehog activity during cerebellum development throughCREB phosphorylation. Development 128:1481–1492.
Ralevic V, Burnstock G. 1998. Receptors for purines and pyrimidines.Pharmacol Rev 50:413–492.
Rieff HI, Raetzman LT, Sapp DW, Yeh HH, Siegel RE, Corfas G. 1999.Neuregulin induces GABA(A) receptor subunit expression and neuriteoutgrowth in cerebellar granule cells. J Neurosci 19:10757–10766.
Rubio ME, Soto F. 2001. Distinct localization of P2X receptors at exci-tatory postsynaptic specializations. J Neurosci 21:641–653.
Sanz JM, Vendite D, Fernandez M, Andres A, Ros M. 1996. Adenosine A1receptors in cultured cerebellar granule cells: role of endogenous adeno-sine. J Neurochem 67:1469–1477.
398 Hervas et al.

Soto F, Garcıa-Guzman M, Gomez-Hernandez JM, Hollmann M, KarschinC, Stuhmer W. 1996. P2X4: an ATP-activated ionotropic receptorcloned from rat brain. Proc Natl Acad Sci USA 93:3684–3688.
Stewart RR, Bossu J-L, Muzet M, Dupont J-L, Feltz A. 1995. Voltage-activated ionic currents in differentiating rat cerebellar granule neuronscultured from the external germinal layer. J Neurobiol 28:419–432.
Thompson CL, Pollard S, Stephenson FA. 1996. Developmental regulationof expression of GABAA receptor alpha1 and alpha6 subunits in culturedrat cerebellar granule cells. Neuropharmacology 35:1337–1346.
Thompson CL, Stephenson FA. 1994. GABAA receptor subtypes expressedin cerebellar granule cells: A developmental study. J Neurochem 62:2037–2044.
Torres-Aleman I, Pons S, Arevalo MA. 1994. The insulin-like growth
factor I system in the rat cerebellum: developmental regulation and role inneuronal survival and differentiation. J Neurosci Res 39:117–126.
Uterberger T, Moskvina E, Scholze T, Freissmuth M, Boehm S. 2002.Inhibition of adenylate cyclase by neuronal P2Y receptors. Br J Pharmacol135:673–684.
Vulchanova L, Arvidsson U, Riedl M, Wang J, Buell G, Surprenant A,North RA, Elde R. 1996. Differential distribution of two ATP-gated ionchannels (P2X receptors) determined by immunocytochemistry. ProcNatl Acad Sci USA 93:8063–8067.
Vulchanova L, Riedl M, Shuster SJ, Buell G, Surprenant A, North RA,Elde R. 1997. Immunohistochemical study of the P2X2 and P2X3receptor subunits in rat and monkey sensory neurons and their centralterminals. Neuropharmacology 36:1229–1242.
P2X and P2Y Receptors in Granule Neurons 399





























