Embed Size (px)
Citation preview

wine Systems 11 ( 1997) 133- 148
a Dbpartement de bioiogre. Unirersltt? iarai, Q&bec, QuG. CIK 7P4, Canada b Soka Un;L crsity, Laborato~ of biological Oceanography, Department of Bioengineering, Faculty of Engineering, 1-236 Tangi-cho, Tokyo
192, Jupun
Received 9 November 1994; accepted 14 September 1995
Abstract
Seasonal variations in under-ice microal,al sedimentation and plankton dynamics in Saroma-ko, a shallow seasonally ice-covered lagoon (Sea of Okhotsk, Hokkaido, Japan), were followed during a 4-week period at the end of winter. At 3-4 day intenals, sediment traps were deployed at three depths from the undersurface of the ice and water column samples were collected. Sampled variables included chlorophyll a (chl a) and pheopigments, particulate organic carbon and nitrogen (POC, PON), cell identification and enumeration, biogenic silica and dissolved inorganic nutrients. POC/PON, POC/chl a and Si/chl a ratios for suspended biomass as well as cell counts showed the presence of a diversified phytoplankton assemblage with a high mi:roheterotrophic biomass. A major peak in clgal sedimentation occurred at the end of the sampling season (chl u tlux LB. 5 mg m - ’ d- ’ ); the sed’ lmented algae included both ice algae and phytoplankton species. Ice algae did not remain suspended in the plankton biomass, but sedimented rapidly upon release from the ice matrix. Results show that Saroma-ko had a rather special food web structure at the end of winter, when both high microalgal export and water column recycling simultaneously occurred under the ice cover.
Keywords: ice; algae; phytoplankton; sedimentation: recycling
1. Introduction followed by massive sedimentation of intact algal
Spring diatom blooms are characteristic of tem- perate and high latitude marine ecosystems. They generally occur upon stabilization of the nutrient-rich surface mixed Iayer, at th ,.R time of increasing irradi- ante, according to the critical depth model of Riley (1942) and Sverdrup ( 1953). Such blooms are often
cells (Smetacek, 1980, 1985) or sinking of fecal pellets from herbivorous zooplankton (Von Bodun- gen et al., 1988; Laws et al., 1988), which lead to a large export of biogenic carbon from the euphotic zone (Legendre, 1990). In contrast, oceanic systems dominated by small phytoplankton (Li et al., 1983; Platt et al., 1983) are thought to sustain a microbial
* Corresponding author. Phone: 418 656 5917. Fax: 418 656 2339. E-mail: [email protected].
web, where recycling of organic matter is high and from which there is little export (Azam et al., 1983; Michaels and Silver, 1988). However, at high lati-
0924-?963/97/$17.00 Copyright 0 1997 Elsevier Science B.V. Ail rights reserved. Pll SO924-7963(96)00034-6
134 C. Mkhel er al. / Jownai cjkrr’tle Systems I I (1997) 133- 148
tudes, low water temperature might significantly re- duce microbial activity (Pomeroy and Deibei, 1986; Pomeroy and Wiebe, 1988).
In ice-covered regions, dense microalgal assem- blages are observed either within the sea ice (e.g., Apoiionio, 1961, 1965; Meguro et al., 1967), at the ice-water interface (e.g., Gosseiin et al., 1985; Trembiay et al., 1989; Johnsen and Hegseth, 1991), or at retreating ice edges (Smith and Nelson, 1985, 1986). These assemblages are generally dominated by diatoms (see Homer, 1985; Homer et al., 1992). In some cases, th!z release of ice algae into the water column is thought to initiate the spring development of phytoplankton (Schandeimeier and Alexander, 1981: Smith and Nelson, 1986; Garrison et al., 1987, 1993). Moreover, ice algae constitute a food source for pelagic copepods, which are capable of grazing near the undersurface of the ice (Runge and Ingram, 1988, 1991; Stretch et al., 19881, so that the down- ward flux of fecal material can represent an impor- tant export pathway for the ice algal biomass (Dunbar, 1984; Von Bodungen et al., 1988; Niithig and Von Bodungen, 1989; Trembiay et al., 1989; Gonzalez et al., 1994). It is thus expected that export of primary production in polar waters would mainly occur through sinking of intact ceils and herbivore fecal pellets. Indeed, the rapid sinking of ice algae during the period of ice melt or at retreating ice edges (Apoiionio, 1965; Alexander, 198U, Leventer and Dunbar, 1987; Bathmann et al,, 1991) can result in a large input of fresh organic material to the benthos (Sasaki and Hoshiai, 1986; Carey, 1987; Fukuchi et al., 1988).
The present study was conducted in Saroma-ko Lagoon, which is the southernmost area having sea- sonal sea ice in the Northern Hemisphere. It has supported a highly productive farming of scallops and oysters for more than 25 years &rata et al., 1991). Given the high benthic production in Saroma- ko, it may be hypothesized, on the one hand, that ice algae are a significant food source for the benthos at the time of ice melt. 0~ the other hand, Saroma-ko Lagoon exhibits less extreme temperature conditions (period of freezing generally < 3 months; Shira- sawa, 1993) than other ice-covered regions of the Northern Hemisphere, which could make it a favor- able area for the development of an under-ice micro- bial community.
The study was designed to assess the relative importance of recycling versus export processes un- der the sea ice of Saroma-ko during the period of ice melt. We concurrently investigated the fate of the ice algal biomass and the structure of the planktonic web at the end of the winter, during the period of growth and decline of the bottom ice-algal community.
2. Material and methods
The suspended and sedimenting alga! biomasses under the first -year ice cover of Saroma-ko Lagoon (Sea of Okhotsk, Hokkaido, Japan; Fig. 1) were
Fig. 1. Location of the sampling station in Saroma-‘ko (Sea of Okhotsk, Japan).

135
the ice at 0.5, 2.5 and 7.5 of the ice. These were
a dense stable layer at the bottom of the trap;
total volume of the trap was gently mixed and transferred to a dark plastic bottle for later analysis in a shore laboratory. The traps were redeploye within 30 min. Transportation of samples between the sampling site and the laboratory, by snowmobile, took about 10 min. Prior to analysis, the trap con- tents were prescreened through a 330 pm mesh to remove large swimmers. This procedure did not appear to remove a significant proportion of algal or fecal material from the samples since there was no significant difference in either chlorophyll a (chl a) or pheopigment concentrations when compared to unscreened samples (paired t-test, p > 0.05). Water samples (Niskin bottles) were taken at 0.5, 2.5, 5.0 and 7.5 m below the undersurface of the ice the day after the traps were retrieved.
Analyses for both suspended and sedimenting biomass included chl a and pheopigments, particu- late organic carbon and nitrogen (POC, PON), bio- genie silica. and cell enumeration. Concentrations of dissolved nutrients (NO,, NO,, PO,, SiO,) were determined on watp* -_w. samples filtered through 0.45 brn acetate filters, after rinsing the sample with sampled water (using a sterile disposable syringe equipped with a filter holder). Ammonium concen- trations were not determined. Samples were kept frozen at - 20°C until analyses on a Bran and Lubbe model TRACCS 800 autoanalyzer according to the methods provided by Strickland and Parsons (1972).
Chlorophyll a and pheopigments were determined fluorometrically with a Turner Designs fluorometer (Helm-Hansen et al., 1965) on subsamples filtered onto Whatman GF/F glass-fiber filters after extrac- tion in 5 ml DMF (n,n-Dimethylformamide) at 4°C in the dark. In the present paper, pheopigment con- centrations are given in chl a equivalents (cf. Conover et al., 1986). Other subsamples were fil- tered onto pre-combusted (24 h at 450°C) Whatman
ore filters using a plas
tion and enumeration were performed under the in- verted microscope (Lun et al., 1958); cell volumes were computed using th f0 for each species t measurements. h total of > 450 cells was measured, i.e., 30 cells per species for the most abundant species and only 10 for the least abundant. For each sampling date, calculation of the total cell volume was based on > 95% of the cell numbers.
3. I. Hydrographic conditions
A general description of Saroma-ko Lagoon is given in Shirasawa (1993) and Shirasawa et al. (1993). The lagoon is shallow ( 14.5 m average depth) and is connected to the Sea of Okhotsk by two channels, the largest one accounting for ca. 90% of the total exchange with the sea. The lagoon is also influenced by freshwater discharge from rivers, which keeps the salinity at < 32. In winter, there is forma- tion of first-year ice, but year to year variability in the duration and the extent of the ice cover is high. In 1992, Saroma-ko Lagoon was completely ice- covered from 5 February through 4 April; ice thick- ness varied between 33 and 39 cm during our sam- pling period (Shirasawa et al., 1993).
Time-series of water level, current speed and direction, temperature, and salinity at the sampling station during February and March 1992 are given in Shirasawa et al. (1993). A two-week record of water levels shows that the tidal cycle seemed to vary from a typically semi-diurnal cycle to a completely diurnal tide over that period. Current speed was low (Z = 4.2 cm s-r ) with little variability. However, sudden changes in current direction at times of diurnal high tides combined with a slow anticlockwise rotation
136 C. Michel et al. / Jomml qf Marine System I I f 1997) 133- 148
suggest that the general circulation pattern was vari- able during the sampling period (Shirasawa et al., 1993). Under-ice water temperature varied little be- fore mid-March, after which it increased slightly to values > - 1.O”C (Shirasawa et al., 1993).
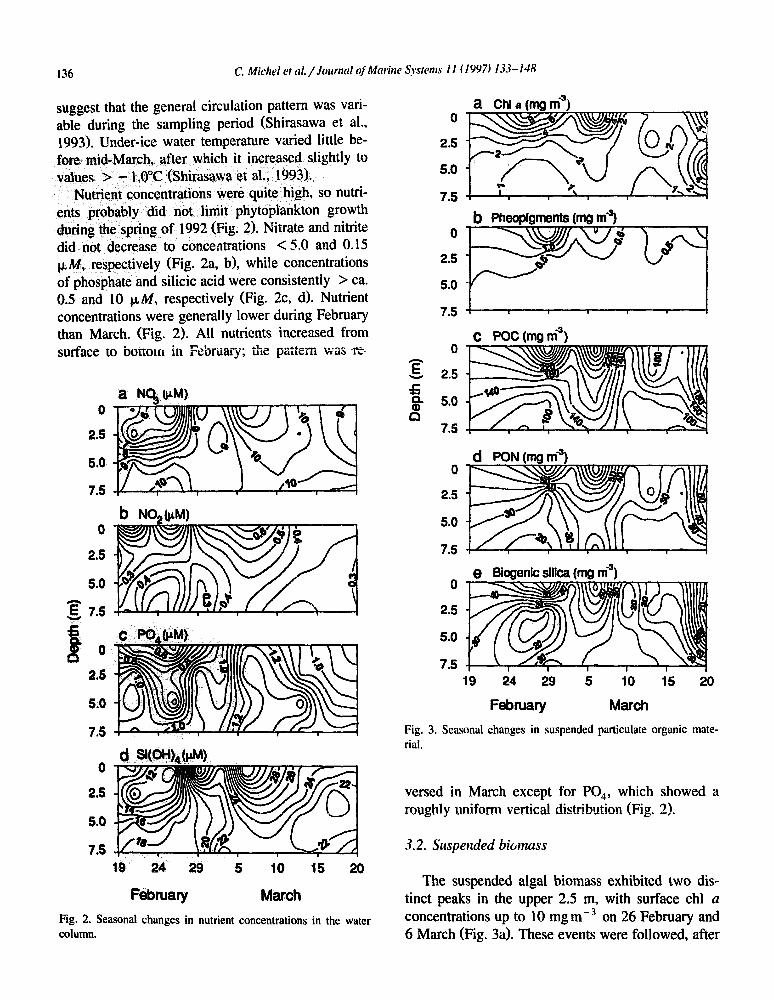
Nutrient concentrations were quite high, so nutri- ents probably did not limit phytoplankton growth during the spring of 1992 (Fig. 2). Nitrate and nitrite did not decrease to concentrations < 5.0 and 0.15 PM, respectively (Fig. 2a, b), while concentrations of phosphate and silicic acid were consistently > ca. 0.5 and 10 p&f, respectively (Fig. 2c, d). Nutrient concentrations were generally lower during February than March. (Fig. 2). All nutrients increased from surface to bottom in Febmary; the pattern ir;as -re--
0
2.5
5.0
7.5
0
2.5
5.0
g 7.5 6
80 2.5
5.0
7.5
0
2.5
5.0
7.5 19 24 29 5 10 15 20
February March
Fig. 2. Seasonal changes in nutrient concentrations in the water column.
0
2.5
5.0
7.5
0
2.5
5.0
7.5
0
E 2.5 5 8 5.0 0
7.5
0
2.5
5.0
7.5
0
2.5
5.0
7.5
a chi 8 (mg m”>
b Pheopigmmts (mg m9)
d PON (mg ni3)
e Biogenic silica (mg ti’)
19
Fig. 3. Seasonal Gal.
24 29 5 10 15 20
February March
changes in suspended particulate organic mate-
versed in March except for PO,, which showed a roughly uniform vertical distribution (Fig. 2).
3.2. Suspended bibmass
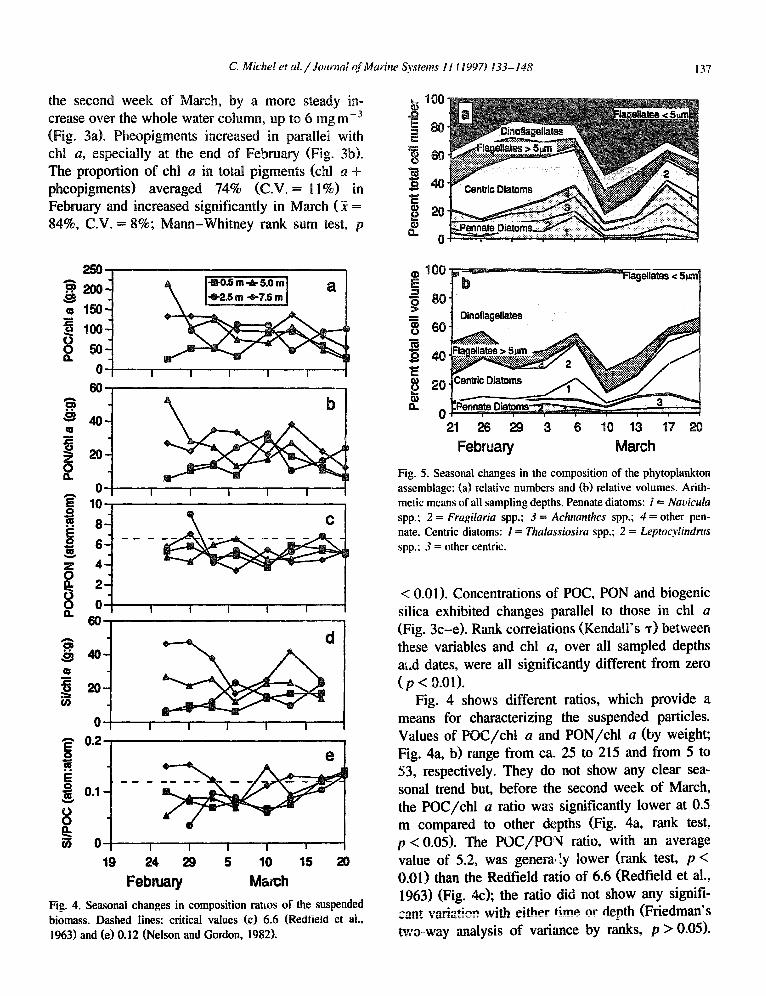
The suspended algal biomass exhibited two dis- tinct peaks in the upper 2.5 m, with surface chl a concentrations up to 10 mg mW3 on 26 February and 6 March (Fig. 3a). These events were followed, after
February and increased significant hitney rank sum test, p
6
6 -- -
4
2
0-T
00 19 24 29 5 10 15 20
February arch Fig. 4. Seasonal changes in composition rataos of the suspended - . biomass. Dashed lines: critical values (c) 6.6 (Redtield et ai., 1963) and (e) 0.12 (Nelson and Gordon, 1982).
Fig. 5. Seasonal changes in the composition of the phytoplankton assemblage: (a) relative numbers and (b) relative volumes. Arith- metic means of all sampling depths. Pennate diatoms: I = Nmiculu spp.; 2 = Fragilaria spp.; 3 = Achnanthes spp.; 4 = other pen- nate. Centric diatoms: I = Thalussiosira spp.; 2 = Leptocylindrus spp.; 3 = other centric.
< 0.01). Concentrations of POC, PON and biogenic silica exhibited changes parallel to those in chl a (Fig. 3c-e). Rank correlations (Kendall’s T) between these variables and chl a, over all sampled depths a;.d dates, were all significantly different from zero ( p < 0.01).
Fig. 4 shows different ratios, which provide a means for characterizing the suspended particles. Values of POC/chl a and PON/chl a (by weight; Fig. 4a, b) range from ca. 25 to 215 and from 5 to 53, respectively. They do not show any clear sea- sonal trend but, before the second week of March, the POC/chl a ratio was significantly lower at 0.5 m compared to other depths (Fig. 4a, rank test, p < 0.05). The POC/POY ratio, with an average value of 5.2, was general :y lower (rank test, p < 0.01) than the Redfield ratio of 6.6 (Redfield et al., 1963) (Fig. 4c); the ratio did not show any signifi- a~-* -rndq+;nn -rith ~ith~r time 0~ &+h (Friedman’s *_=a&; f a$=&‘-li_*ig ** ii*, ____=_ _____ _ two-way analysis of variance by ranks, p > 0.05).
138 C. Micltel et al. / Jmtrnal of Amine Systems I I (I 997) 133-148
spp. and Achnanthes spp., accounted for ca. 20 to 30% of cell numbers and showed highest proportions on 6 and 17 March. Naked flagellates (mostly < 20 pm) and dinoflagellates together comprised ca. 25% (median 23%; range 1 l-29%) of cell numbers (Fig. 5a). On a volume basis, the distribution pattern of
February March
Fig. 6. Seasonal changes in particulate organic matter flux in sediment traps at 0.5, 2.5 and 7.5 m: (a) absolute (histograms) and percent (chl u/total pigments) fluxes of chl n and (b) flux of particulate organic carbon (POC). No data at 0.5 m from 16 to 19 March.
The biogenic silica/chl a ratio ranged between ca. 5 and 50 and increased with depth, indicating loss of pigments relative to silica over the water column (Fig. 4d; Friedman’s, p < 0.05). Biogenic silica/POC values were often lower than 0.12 (Fig. 4e), which is a typical value for diatom assemblages (Nelson and Gordon, 1982). The ratio was slightly higher at 7.5 m than at all other depths (Fig. 4e), but the difference was not statistically significant (Fried- man’s, p > 0.05).
The suspended phytoplankton biomass was a mixed assemblage of pennate and centric diatoms together with naked flagellates and dinoflagellates. Cells < 10 pm in diameter accounted for ca. 70% of total cell numbers. Apart from flagellates < 5 pm, which made up a large proportion of cell num- bers (ca. 40%, Fig. 5a) but represented negligible cell volumes (ca. l%, Fig. 5b), the assemblage was dominated by ikptocylindrus spp., a centric diatom that made up ca. 25% of the cell numbers before the second week of March (Fig. 5a). Other centric di- atoms were almost absent before the second week of March, after which Thalassiosira spp. made up a smali fraction of ceil numbers (< IO%, Fig. 5a). Pennate diatoms, mostly Naoicula spp., Fragilaria
0 ; I I ‘I I I I
60
40
20
0 E 0.5 g 0.4- e
g 0.3- g OS- 8 O.l-* k in 0 I I I I I
19 24 29 5 10 15 20 February March
Fig. 7. Seasonal changes in composition ratios of the sedimented biomass. Dashed lines: critical values (c) 6.6 (Redfield et al., i863) and (e) 0.12 (Nelson and Gordon, 1982). No data at 0.5 m from 16 to 19 March.

ichel et al. /.hunlal qf wirte SyUerns I I f 1997) IS..?- 148 139
The contribution
the end of lLlarch (Fig. 5b).
3.3 Sedimer- fed biomass
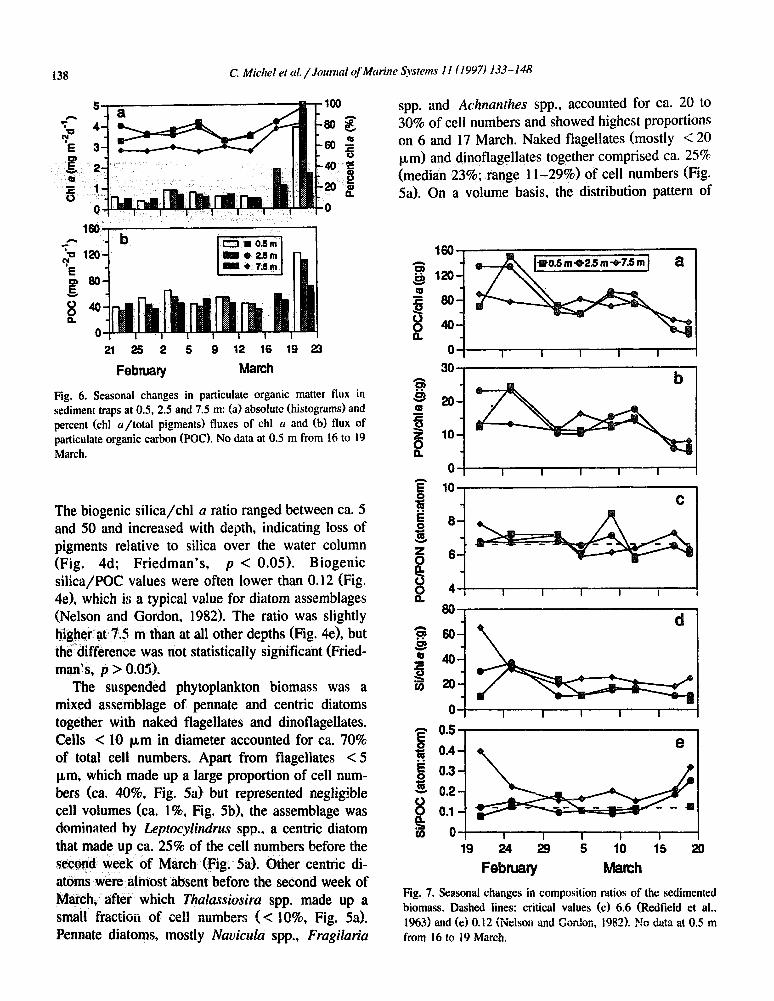
A major change in sedimentation occurred after 16 March, when the chl a flux increased fro mgme2 d-’ to a m aximum value of ca. 5 mg m- 2 d- ’ (Fig. 6a). Concomitantly, the proportion of chl a in total pigments (chl u + pheopigments) increased from an average value of 67% (C.V. = 13%; three depths, six sampli;lg dates) to 86% (C.V. = 11%; three depths, two samp!ing dates) (Fig. 6a). The contribution of chl a to trapped pigments increased at all depths, but it was significantly lower in the 7.5 m trap than at other depths during the entire sampling period (Fig. 6a; Friedman’s, p < 0.01; multiple comparison test: 7.5 m < 2.5 m; 7.5 m < 0.5 m).
The increased flux of chl c1 was accompanied by an increase in the flux of POC, from ca. 45 mgm-’ d-’ to a maximum value of 120 mgm-’ d-’ (Fig. 6b). However, the increase in POC sedimenta- tion (ca. three-fold) was small compared to that of
with depth (Friedman’s, p > 0.05). The POC/PON ratio did not show any significant change with either time or depth (Friedman’s, p > 0.05), and it was equivalent to the Redfield value of 6.6 (X = 6.6, C.V. = 8.5%) (Fig. 7~). Ratios of biocenic Si/chl a and biogenic Si,/POC were higher 8; X5 m than at
er depths (Fig. 7d, e), which is similar to what was observed for the same ratios in the suspended biomass (Fig. 4d, e). The difference was statistically significant for only the Si/POC ratio (Friedman’s, p < 0.05).
Table 1 shows the relative export of suspended chl a, POC and PON as well as their sinking rates. On a day-to-day basis, the biomass collected in sediment traps always represented < 8% of the SUS-
pended material. The percentoqe of suspended POC that sank out of the water column was always higher
Table 1 ’ Areal suspended biomass, proportion lost through sinking, and sedimentation rates. The sampling period is for each sediment trap mooring, the suspended biomass was sampled within each mooring period
Sampling period Area1 biomass, from 0 to 7.5 m Daily proportion of areal biomass Sedimentation rate at 7.5 m (mgm-*) a intrapsat7.5m(%d-‘)b (md-‘)’
Chl a POC PON Chl a POC PON Chl a POC PON
21-25 Feb. 25 Feb.-2 March 2-5 March S-9 March 9- 12 March ‘I 2- 16 March II 6-?O March 20-.I3 March
17.90 22.38 16.59 28.58
8.44 15.08 13.84
2.80 0.21 707.4 160.3 2.15 5.25 3.96 0.16 0.39 0.30 204.0 290.4 4.00 3.74 2.54 0.30 0.28 0.19 709.4 382.3 2.25 3.05 2.71 0.17 0.23 0.20 737.5 201.9 7.91 6.21 4.34 0.59 0.47 0.32 058.4 241.8 3.13 3.37 2.72 0.23 0.25 0.20 907.2 185.1 7.71 5.43 4.30 0.58 0.41 0.32
39.05 1551.4 357.9 4.28 4.57 3.65 0.32 0.34 0.27
a Trapezoidal integration from 0 to 7.5 m. b (Flux T.5 =/area1 biomass,_7.f; ,) X 100. c Flux 7.s &area1 biomass,_,,, J7.5 m).
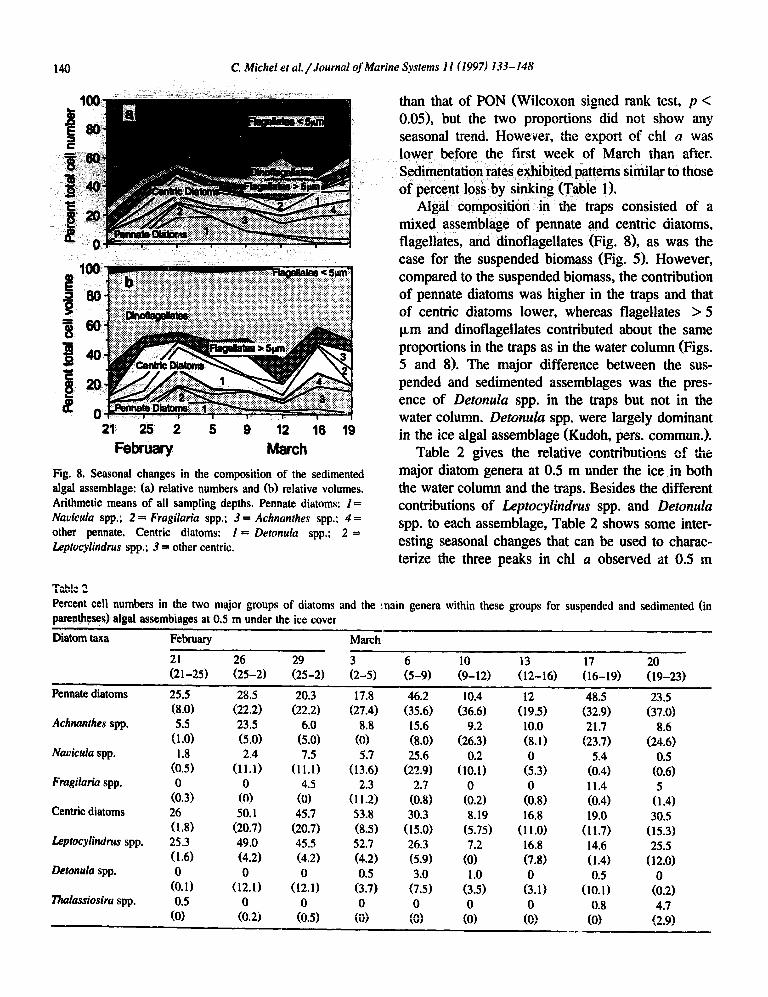
140 C. Niche1 et al. /Journal of Marine Systems 11 (1997) 133- I48
21 25 2 5 9 12 16 19 February March
Fig. 8. Seasonal changes in the composition of the sedimented algal assemblage: (a) relative numbers and (b) relative volumes. Arithmetic means of all sampling depths. Pennate diatoms: I = Navicula spp.; 2 = Fragilaria spp.; 3 = Achnanthes spp,; 4 = other pennate. Centric diatoms: I = Detonula spp,; 2 = Leptocylindrus spp.; 3 = other centric.
than that of PON (Wilcoxon signed rank test, p < 0.05), but the two proportions did not show any seasonal trend. However, the export of chl 611 was lower before the first week of March than after. Sedimentation rates exhibited patterns similar to those of percent loss by sinking (Table 1).
Algal composition in the traps consisted of a mixed assemblage of pennate and centric diatoms, flagellates, and dinoflagellates (Fig. 81, as was the case for the suspended biomass (Fig. 5). However, compared to the suspended biomass, the contribution of pennate diatoms was higher in the traps and that of centric diatoms lower, whereas flagellates > 5 p,rn and dinoflagellates contributed about the same proportions in the traps as in the water column (Figs. 5 and 8). The major difference between the sus- pended and sedimented assemblages was the pres- ence of Detonula spp. in the traps but not in the water column. Detonuka spp. were largely dominant in the ice algal assemblage (Kudoh, pers. commun.).
Table 2 gives the relative contributions of the major diatom genera at 0.5 m under the ice ,in both the water column and the traps. Besides the different contributions of LeptocyZindrus spp. and Detonula spp. to each assemblage, Table 2 shows some inter- esting seasonal changes that can be used to charac- terize the three peaks in chl a observed at 0.5 m
Tsb!e 2 Percent cell numbers in the two major groups of diatoms and the :nain genera within these groups for suspended and sedimented (in parentheses) algal assembiages at 0.5 m under the ice cover
Diatom taxa February March1
z-251 &*I ?k2) :2-5) (65_9, tso_lu ::2-16) 17 (16-19) E-*3)
Pennate diatoms 25.5 28.5 20.3 17.8 46.2 10.4 12 48.5 23.5
Achnanthes spp.
Navicula spp.
Fragilaria spp.
Centric diatoms
Leptocylindrus spp.
Detonula spp,
Thalassiosira spp.
03.0) (22.2) (22.2) (27.4) (35.6) (36.6) (19.5) (32.9) (37.0) (:::, 23.5 15.6 10.0 21.7
(5.0) (:::I (Y
(8.0) c2:::,
(8.1) (23.7) (2%
$, (1::S UT::, &::I 25.6
(22.9) (1::) z.3, (:::I E,
(i.31 (L 5011
zi5 (1:::) (G, (i.2) &I 11.4 (0.4) (7.4)
26 45.7 53.8 30.3 8.19 16.8 19.0 30.5 (1.8) (20.7) (20.7) (8.5) (15.0) (5.75) (11.0) (11.7) (15.3) 25.3 49.0 45.5 52.7 26.3 16.8 14.6 25.5 (1.6) (4.2) (4.2) (4.2) (5.9) G2 (7.8) (1.4) (12.0)
&I &I &) $1 (::$ & &I (1% (i.2) 0.5 0 0 0 0 0 0 4.7
(0) (0.2) (0.51 (0) (cj (91 (0) (2.9) _r_

sponded to a large increase i
Instead, high proportions of Nakula spp. and Detonulu spp. were found in the trapped assemblage. Tine second peak, on 6 Marcl,, ~~~~n-z--+~n~ +q h;k LuI kGz3ptiilubu rb 111gjll
proportions of Navicula spp. in both the water col- umn and trapped assemblages. Finally, the increase in suspended chl a observed after mid-March dif- fered from the previous ones, with significant contri- butions of Fragilaria and Thalassiosira species. trapped assemblage at 0.5 m was dominated by Achnanthes spp., with Leptocylindrus, Detonula and Thalassiosira also present (Table 2).
iscussion
4.1z Water column
Maximum nutrient concentrations in the under-ice water column (Fig. 2) were similar to those reported by Tada et al. (1993) for March 1987, i.e., lo- 12 @4 for NO, + NO, and 1.3 p M for PO,. Accord- ing to these authors, their values were similar to those recorded earlier (Kikuchi, 1979). Maximum chl a concentrations (ca. 10 mg m-“; Fig. 3a) are consistent with those reported in Saroma-ko just after ice melt (in April), i.e., 6.7 and 19.1 mg m -’ in 1987 and 1988, respectively (Tada et al., 1993). During the ice-covered period (in 1987) low sus- pended chl a concentrations ( < 1 mg mV3), reported by Fukuchi et al. (1989) and Satoh et al. (1989), were followed by a bloom-like increase with concen- trations reaching 3-5 mgmV3 (Fukuchi et al., 1989). These last values are similar to those we measured in 1992 at the end of March (up to ca. 6 mgms3; Fig. 3a). At that time, the steady increase in chl a concentrations over the water column likely reflected the onset of a phytoplankton bloom under the ice cover because of the relatively high occurrence cf Thalassiosiru (Fig. 5), which has been reported to dominate the water column assemblage in Saroma-ko (Satoh et al., 1983).
At all times during the sampling period. the phy- toplankton area1 standing stock (integrated values
coverage in Saroma-ko was 61 days 100 days in 1987 (Shirasawa et al., and constant proportions of Lep
equent decrease (Fig. 5), suggest that this species actively growing prior to the start of sampling,
during a period when Saroma-ko was still not com- pletely ice covered. In ad ition, the steady presence of dinoflagellates (Fig. 5) which were shown to be abundant after ice melt in Saroma-ko (Watanabe et al., 1993), suggests that a phytoplankton assemblage did persist during the 1992 winter due to the short ice-covered period and consequent increased under- water irradiance.
POC/chl a ratios (range: 25-214 by weight; Pig. 4a) are within the range of published values for phytoplankton assemblages in the Weddell Sea (31- 5 13; Nelson et al., 1989; Leynaert et al., 1991; Smetacek et al., 1992) and in the Ross Sea (138; Smith and Nelson, 1985) as well as for ice algae in the Canadian Arctic (30- 180; Gosselin et al., 1990). High POC/chl a ratios have been reported for phy- toplankton cultures under nutrient limitation (Caperon and Meyer, 1972; Harrison et al., 1977), yet the abundance of nutrients in Saroma-ko rules out nutri- ent limitation. The observed high POC/chl a ratios may reflect the presence of heterotrophic contribu- tors to the suspended biomass. Indeed, cell counts showed the presence of significant numbers of choanoflagellates (5 = 1.4 X lo4 ind. l- ‘) and cili- ates (E = 1.9 X lo3 ind. l- ‘) in the water column. The observed concentrations of choanoflagellates fall within the range of values found in the ice edge zone of the Weddell Sea, i.e., 1000 to 2 X lo6 ind. 1-l (Silver et al., 1980; Buck and Garrison. 1983, 1988). Numbers of ciliates similar to those in Saroma-ko have been reported in the Antarctic, i.e., 1 X IO3 to 9 x IO3 ind. l- ’ (Buck and Garrison, 1983; Leaky et al., 1994). It thus appears that an active microbial rood web coexisted witll phytopiankton in the water

142 C. Michel et at. / Jownai of Marine Systems I I f 1997) 133- 148
column during the sampling period. This agrees with previous findings of Takahashi ( 198 11, who observed a rich autottophic and heterotrophic community un- der the ice of Saroma-ko. The relatively low POC/PON for suspended biomass (Fig. 4c) is also consistent with the presence of high heterotrophic biomass. Indeed, in the absence of nutrient limita- tion, dinoflagellates have been shown to exhibit lower POC/PON ratios than phytoplankton (Cullen and Horrigan, 1981). Bacteria are also known to have low POC/PON ratios (ca. 4; Lee and Fuhrman, 1987) and, in a culture of bacterivorous flagellates with bacteria, Borsheim and Bratbak (1987) mea- sured POC/PON of ca. 5. For different tintinnid species, Verity and Langdon (1984) measured POC/PON ratios ranging from 4.6 to 5.5, and Klep- pel et al. (1991) reported that ciliates contain more proteins and lipids than phytoplankton. Finally, the slightly lower Si/POC ratio (Fig. 4e) compared to that of diatom-dominated assemblages (0.12; Nelson and Gordon, 1982) also reflects a diversified sus- pended biomass, with some contributors lacking siliceous structures.
4.2, Sedimentation
A typical feature of springtime sedimentation in ice-covered marine waters is a transient increase in the flux of pigments. This has been reported for both the Antarctic (Von Bodungen et al., 1986; Sasaki and Hoshiai, 1986; Matsuda et al., 1987; Fukuchi et al., 1988; Bathmann et al., 1991) and the Arctic (Tremblay et al., 1989; Wassmann et al., 1991 and references therein). The spring cycle of sedimenta- tion in Saroma-ko also exhibited an increase in the flux of pigments and POC (Fig. 6). In 1992, the flux of POC was within the range of previous estimates for Saroma-ko (Sasaki and Fukuchi, 1993), and the fluxes of both POC and chl a were comparable to those reported for some other ice-covered regions (Fukuchi and Sasaki, 198 1; Sasaki and Hoshiai, 1986; Fukuchi et al., 1~88). The proportionally stronger increase in the flux of chl a compared to that of POC implies that algae contributed most of the exported biomass during the period of high sedimen- tation, as previously observed in the Antarctic (Matsuda et al,, 1987). Using a POC/Chl a ratio of 20, which was the average value for ice algae and
phytoplankton in Saroma-ko for 1992 (Kudoh, 1993) algae would comprise ca. 60% of the sinking POC after mid-March (30% before that period; see Fig. 6) However, at the same time, chl a represented ca. 90% of the total pigments in the traps (Fig. 6a), so pheopigments (ca. 10% of the trapped pigments, presumably from copepod feces) can hardly explain the remaining 40% of sinking POC. Downs and Lorenzen (1985) reported that POC/pheopigment ratios of copepod fecal pellets range from 1 l- 18 for herbivorous animals to 25-29 for omnivores or car- nivores. Using a POC/pheopigment conversion fac- tor of 30 (fecal pellets of copepods with a broad diet; Downs and Lorenzen, 1985) to convert pheopig- ments into sinking POC results in only ca. 25% of non-algal POC being explained by sinking fecal pellets during the whole sampling period (23 and 27% during periods of high and low fluxes, respec- tively). This represents a minimum estimate, since the calculation accounts neither for the loss of pig- ments during digestion (Conover et al., 1986; Lopez et al., 1988; Head, 1992) nor for the formation of colorless residues (Kiorboe and Tiselius, 1987), both of which could have occurred during copepod feed- ing. In addition, the presence of dinoflagellates and flagellates in both the water column and sediment traps (Fig. 5 and Fig. 8) suggests that they con- tributed to the exported organic matter by direct sinking and fecal pellet production. Heterotrophic flagellates and dinoflagellates can degrade chl a into various types and proportions of pigments (Goldman and Caron, 1985; Strom, 1993) and also into color- less residues (Klein et al., 1986; Barlow et al,, 1988) which could make up as much as 95% of the in- gested pigments (Klein et al., 1986). Given the sig- nificant numbers of microheterotrophs, their feces could account for part of the unexplained sinking POC. Flagellates and dinoflagellates were not sys- tematically identified, but the dominant dinoflagel- lates were Gymnodinium spp., which are known to have a broad diet and produce feces (Btick et al., 1990). Finally, the higher POC/PON ratios in sedi- ment traps (Fig. 7c) compared to the water column (Fig. 4c) are indicative of accumulation of feces in the sinking material, since these ratios are often reported to be higher in feces than in the food source as a consequence of preferential nitrogen assimila- tion by animals (Morales, 1987).

tion is the incre
vide the season into three perio
half of March (ca. 0.7 mgm-* d- ‘), and was high at the end of March (ca. 3 mg m-* d- I). During the sampling period, the ice algal assemblage was domi- nated by Detonula spp. and Navicula F pers. commun.), so these two taxa can tracers for ice algae released from the ice. Similarly, Achnanthes spp. and Leptocylindrus spp. can be used to trace phytoplankton, since they were scarce or absent in the ice algal assemblage (Kudoh, pers. commun.). It is interesting to note that, during the whole season, ice algal taxa (mainly Detonula and Navicula) contributed to the trapped material (Table 2; Fig. 8) but did not make up a significant part of the suspended assemblage, except on one occasion as a transient contribution of Navicula (on 6 March; Table 2). This contribution of ice algae (Navicula) to the suspended biomass corresponds to a transient increase in chl a close to the undersurface of the ice (Fig. 3a). The reverse did not occur for phytoplank- ton, i.e., genera Achnanthes and Leptocylindrus were commonly found in sediment traps in various proportions; however, during the first part of the sampling season, their abundances in the trapped assemblage were quite low compared to those in the water column irable 2). At the end of the season, during the period of increased flux, both phytoplank- ton and ice algal taxa were abundant in sediment traps.
The above observations concerning sedimenting algae lead to two conclusions. First, except on one occasion, ice algae did not contribute to the phyto- plankton assemblage in Saroma-ko. This may be due to the shallowness of Saroma-ko Lagoon, which would prevent sinking algae from staying in the water column as part of the suspended biomass. The algal biomass still present in the ice at the beginning of ice melt thus rapidly reaches the bottom, where it provides a large quantity of food for the benthos. The rapid transit from the ice to the lagoon floor favors a supply of “high quality” food, as there is
phytoplankton durin ally so low that its
pellets from herbivorous zooplankton often make up a large part of the downward flux in ice-covered
1986; Carey, 1987; 1199 1) and, on the short term,
massive sinking of ice algae may occur Hoshiai, 1986). Saroma-ko is the southe sonally ice-covered area in the Northern so the ice is not very t!Cck (ca. 30 cm). This, combined with the fact that the ice-covered period was short in 1992, allowed relatively high phyto- plankton production under the ice cover. As a conse- quence, the area1 suspended biomass (chl a ranging from 8 to 39 mgm -* ; Table 1) was of the same order as the biomass of ice algae (from 5 to 38 mgm-*; Robineau, pers. commun.). It follows that phytoplankton significantly contributed to the pig- ment flux, as shown by species composition in the trapped material (Table 2).
4.3. Tropitic pathways
Both water column and sediment trap data indi- cate that the under-ice alga! cnsmt~nity in Saroma-ko _-__ __ --- probably supported an active microbial food web at the end of winter in 1992. Numbers of micro- heterotrophs in the water column were higher than in the ice (Sime-Ngando et al., 1997-this issue) and some recycling of algal material seemed to occur in the water column. This is shown by the increasing Si/chl a and Si/POC ratios with d water column and sediment traps 7d,e), and by the lower contribution of chl a to the total pigments in the 7.5 m sediment trap (Fig. 6a). Table 1 indicates that the daily proportion of ex- pofied algal binmass through sinking did not exceed

ca. 8% of the suspended biomass. The same table shows that the relative expoq of PON was consis- tently lower than that of POC, which led to lower sinking rates for PON than for POC. Again, this agrees with the view that some utilization and recy- cling of organic matter took place in the water column, since relatively more PON than POC re- mained suspended. The higher POC/PON ratio in sediment traps compared to that of suspended matter could reflect recycling of PON and/or the presence of fecal matter in the traps (see Section 4.2). The fact that this ratio did not change with depth (Fig. 7c) indicates that the material reaching the floor of the lagoon was still of high nutritional value (Findlay, 1982).
Sedimentation rates computed from water column and trap data were low (Table 1) and fall within the range of non-nutrient limited cells (e.g., Bienfang et al., 1982) actively growing cells (Eppley et al., 1967) and/or small cells (Smayda, 1970, 197 1). This is consistent with the facts that phytoplankton did not seem to experience nutrient limitation and that ca. 70% of the suspended algae (by numbers) were in the < 10 p,rn size class. Considering that algae released from the ice probably had higher sinking rates than phytoplankton (i.e., the former did not remain long in suspension), sedimentation rates derived from trap and water column data would represent maximum rates for phytoplankton algae. Saroma-ko would thus be a mixed environment, with high suspended biomass of small cells and, simulta- neously, high export of larger ice algae. This leads to both an active microbial web in the water column and a large export towards the bottom. In the Wed- dell Sea ice edge zone, Lancelot et al. (1993) ob- sef\lPd 0 84nminanfia nf nnnnnhxrtnnlnnlfltdx- -nil- A-t v -- Y YVLI~L~~UL~VV v* ukau~pa~ bupauunbuafi bbub LLICLC
they attributed to the presence of overwintering krill, whose grazing reduced diatom seeding in the water column. In Saroma-ko, the presence of overwintering zooplankton could also have reduced the abundance of large cells in the suspended assemblage. However, it appears that zooplankton grazing pressure was low during the sampling period (Saito and Hattori, 1997- this issue). It is thus suggested that the main factor preventing the establishment of large cells in the suspended assemblage was their rapid sedimentation to the bottom. Because of the shallowness of the lagoon, fast-sinking cells rapidly reach the bottom,
where strong grazing pressure by the cultured scal- lops removes them from the prinary production compartment. The role of benthos in shallow Saroma-ko would thus be similar to that of krill in the ice-edge zone, where the grazing pressure of krill on ice algae, especially at the time of ice melt, may control the initiation and magnitude of phytoplank- ton blooms dominated by diatoms (Marschall, 1988; Smetacek et al., 1990) and favor the development of nanophytoplankton (Lancelot et al., 1993). In Saroma-ko, the short herbivorous web (from ice algae to benthos), which results from rapid sinking of ice-algal cells, leaves way for a microbial web in the water column. Contrary to the idea that microbial activity may be very low in low-temperature waters (Pomeroy and Deibel, 1986; Pomeroy and Wiebe, 1988) data from Saroma-ko are consistent with the view that the microbial compartment may be an important component of some high-latitude ecosys- tems (Garrison and Buck, 1989; Azam et al., 1991; Garrison et al., 1993).
Saroma-ko showed coexistence of water column microheterotrophic production and rapid export of ice algae to the benthos during the period of ice melt. In the continuum of trophic pathways ranging from the microbial loop to the herbivorous web, proposed by Legendre and Rassoulzadegan (1995) Saroma-ko would represent the special case of a system where a pelagic microbial food web superimposes itself on a herbivorous web, leading to simultaneous high ex- port form the ice to the bottom and high recycling in the under-ice water column. The particular condi- tions favoring the transient co-occurrence of the two trophic pathways at the time of ice melt suggest that coupling between the water column and the benthos
^____ _^ -
Acknowledgements
The main source of funding for Canadian partici- pation in the SARES project was a grant from the Japan Science and Technology Fund (Department of External Affairs and Trade of Canada). C.M. re- ceived a post-graduate scholarship from the Natural Sciences and Engineering Council of Canada (NSERC). A research grant to L.L. from NSERC and a team grant from the Fonds FCAR of Quebec

thanks are due to t
ockanqr+ +i@es dl4 gg&?fbec grants from NSERC and the
and C. Manger and P. Peltola for their help in the field. This paper is a contribution to the programme of GIROQ and to the Hokkaido National Fisheries Institute.
eferences
Alexander, V., 1980. Interrelationships between the seasonal sea ice and biological regimes. Cold Reg. Sci. Tech. Rep., 2: 157-178.
Apollonio, S., 1961. The chlorophyll content of Arctic sea ice. Arctic, 14: 196-200.
Apollonio, S., 1965. Chlorophyll in the Arctic sea ice. Arctic, 18: 118-122.
Azam, F., Fenchel, T., Field, J.G., Gray, J.S., Meyer-Reil, L.A. and Thingstad, F., 1983. The ecological role of water column mtcrobes in the sea. Mar. Ecol. Prog. Ser., LO: 257-263.
Azam, F., Smith, D.C. and Hollibaugh, J.T., 1991. The role of the microbial loop in Antarctic pelagic ecosystems. In: E. Sak- shaug, C.C.E. Hopkins and N.A. 0ritsland (Editors), Proc. Pro Mare Symposium on Polar Marine Ecology. Polar Research, Trondheim, 12-16 May 1990, pp. 239-243.
Barlow, R.G., Burkill, P.H. and Mantoura, R.F.C., 1988. Grazing and degradation of algal pigments by the marine protozoan 0xyrrhis marina. J. Exp. Mar. Biol. Ecol., 119: 119-129.
Bathmann, U., Fisher, G., Mhller, P.J. and Gerdes, D., 1991. Short-term variations in particulate matter sedimentation off Kapp Norvegia, Weddell Sea, Antarctica: hielation to water
. . . lnass advectton, tee cover, plaukton biomass and feedmg activity. Polar Biol., 11: 185-195.
Bienfang, P.K., Harrison, P.J. and Quarmby, L.M., 1982. Sinking
uck, K.R. and Garrison, D.L., 1988. Distribution and abundance of choanoflagellates (Acanth ae) across ‘rhe ice-edge zone in
ddell Sea, Antarctica. ar. Biol., 98: 263-269.
fecal pellet production by an athecate dinoflagellate in Antarc- tic sea ice. Mar. Ecol. Prog. Ser., 60: 75-84.
Caperon, 9. and Meyer, J., 1972. Nitrogen-limited growth of marine phytopiankton. I. Changes in population characteristics with steady-state growth rate. Deep-Sea Res., 19: 601-618
Carey, A.G., 1987. Particle flux beneath fast ice in the shallow southwestern Beaufort Sea. Arctic Ocean. Mar. Ecol. Prog. Ser., 40: 247-257.
Conover, R.J., Durvasula, R., Roy, S. and Wang, R., 1986. Probable loss of chlorophyll-derived pigments during passage through the gut of zooplankton, and some of the conse- quences. Limnol. Oceanogr., 31: 878-887.
Cullen, J.J. and Horrigan, S.G., 1981. Effects of nitrate on the diurnal vertical migration, carbon to nitrogen ratio, and the photosynthetic capacity of the dinoflagellate Gymnodium splendens. Mar. Biol., 62: 81-89.
Downs, J.N. and Lorenzen, C.J., 1985. Carbon:pheopigment ratios of zooplankton fecal peliets as an index of herbivorous feed- ing. Limnol. Oceanogr., 30: 1024-1036.
Dunbar, R.B., 1984. Sediment trap experiments on the antarctic continental margin. Antarct. J. U.S., 19: 70-71.
Eppley, R.W., Holmes, R.W. and Strickland, J.D.H., 1967. Sink- ing rate of marine phytoplankton measured with a fluorometer. J. Exp. Mar. Biol. Ecol., 1: 191-208.
Findlay, S.E.G., 1982. Effect of detrital nutritional quality on population dynamics of a marine nematode (Dipfofuimellu chhvoodi). Mar. Eiol., 68: 223-227.
Fukuchi, M. and Sasaki, H., 1981. Phytoplankton and zooplankton standing stocks and downward flux of particulate material around fast ice edge of Liitzow-Holm Bay, Antarctica. Mem. ?&ii. inst. Poiar Res. Ser. E, 34: i3-36.
Fukuchi, M., Hattori, H., Sasaki, H. and Hoshiai, T., 1988. A phytoplankton bloom and associated processes observed with a long-term moored system in antarctic waters. Mar. Ec01. Prog. Ser., 45: 279-288.
Fukuchi, M., Watanabe, K., Tanimura, A., Hoshiai, T., Sasaki, H., Satoh, H. and Yamaguchi, Y., 1989. A phytoplankton bloom under sea ice recorded with a moored system in Lagoon Saroma Ko, Hokkaido, Japan. Proc. NIPR Symp. Polar Biol., 2: 9-15.
Garrison, D.L. and Buck, K.R., 1989. Protozooplankton in the eddell Sea, A~ta~ctica~ a ance a
edge zone. Polar Biol., 9: 341-35 1. Garrison, D.L ,, Buck, K.R. and Fryxell, G.A., 1987. Algal assem-
blages in antarctic pack ice and in ice-edge plankton. J. Phycol., 23: 564-572.

146 C. Michel et al, / Jourrwl of Marine Systems 1 I i 1997) 133- 148
Garrison, D.L., Buck, K.R. and Gowing, M.M., 1993. Winter plankton assemblage in the ice edge zone of the Weddell and Scotia Seas: composition, biomass and spatial distribution. Deep-Sea Res., 40: 3 1 l-338.
Goldman, J.C. and Caron, D.A., 1985. Experimental studies on an omnivorous microflagellate: implications for grazing and nu- trient regeneration in the marine microbial food chain. Deep- Sea Res., 32: 899-915.
Gonz6lez, H.E., Kurbjeweit, F. and Bathmann, U.V., 1994. OC- currence of cyclopoid copepods and faecal material in the Halley Bay region, Antarctica, during January-February 199 1. Polar Biol., 14: 33 l-342.
Gosselin, M., Legendre, L., Demers, S. and Ingram, R.G., 1985. Responses of sea-ice microalgae to climatic and fortnightly tidal energy inputs (Manitounuk Sound, Hudson Bay). Can. J. Fish. Aquat. Sci., 42: 999-1006.
Gosselin, M., Legendre, L., Therriault, J.-C. and Demers, S.. 1990. Light and nutrient limitation of sea-ice microalgae (Hudson Bay, Canadian Arctic). J. Phycol., 26: 220-232.
Harrison, P.J., Conway, H.L., Holmes, R.W. and Davis, C.O., 1977. Marine diatoms grown in chemostats under silicate or ammonium limitation. III. Cellular chemical composition and morphology of Chaetoceros debilis, Skeletonemu costatwn, and Thalassiosira gravida. Mar. Biol., 43: 19-31.
Head, E.J.H., 1992. Gut pigment accumulation and destruction by arctic copepods in vitro and in situ. Mar. Biol., ! 12: 583-592.
Helm-Hansen, 0.. Lorenzen, C.J., Holmes, R.W. and Strickland, J.D.H., 1965. Fluorometric determination of chlorophyll. J. Cons. Perm. Int. Explor. Mer, 30: 3-15.
Homer, R.A., 1985. Ecology of sea-ice microalgae. In: R.A. Homer (Editor), Sea Ice Biota. CRC Press, Boca Raton, FL, pp. 147- 157.
Homer, R., Ackley, S.F., Dicckmann, G.S., Gulliksen, B., Hoshiai, T., Melnikov, LA., Reeburgh, W.S., Splinder, M. and Sulli- van, CW., 1992. Ecology of sea ice biota. I. Habitat and terminology. Polar Biol., 12: 4 17-427.
Jahnsen, G. and Hegseth, E,N,, 1991. Photoadaptation of sea-ice microalgae in the Barents Sea, Polar. Biol., 11: 179-184.
Kikuchi, K., 1979. Water quality of Lake Saroma during the period from 1967 to 1977. J. Hokkaido Fish. Exp. Stn., 36: 233-254 (in Japanese).
Kis;rboe, T. and Tiselius, P.T., 1987. Gut clearance and pigment destruction in a herbivorous copepd, Acartia tonsa, and the determination of in situ grazing rates, J. Plankton Res., 9: 525-534.
Klein, B., Giekes, W.W.C. and Kraay, G.G., 1986. Digestion of chlorophylls and carotenoids by the marine protozoan Oxyrrhis marina studied by h.p.1.c. analysis of algal pigments. J. Plank- ton Res., 8: 827-836.
ueppel, G.S., Holliday, D.V. and Pieper, R.E., 1991. Trophic interactions between copepods and microplankton: a question about the role of dia!oms. Limnol. Oceanogr., 36: 172-1X
Kudoh, S,, 1993. Respor,ses of microalgal primary productivity to the reduction of sea ice coverage in Lake Saroma during winter. Bull. Plankton Sot. Jpn., 39: 155-156.
Kurata, M-, Hoshikawa, H. and Nishihama, Y., 1991. Feeding rates of Japanese scallop Patiopecten yessoensis in suspended
cages in Lagoon Saroma-ko. Sci. Rep. Hokkaido Fish. Exp. Stn., 37: 37-57 (in Japanese).
Lancelot, C., Mathot, S., Becquevort, S., Dandois, J.-M. and Billen, G., iy93. Carbon and nitrogen cycling through the microbial network of the marginal ice zone of the Southern Ocean with particular emphasis on the northwestern Weddell Sea. In: S. Caschetto (Editor), Belgian Scientific Research Programme on the Antarctic. Scientific Results of Phase Two (10/1988-05/1992). Vol. I. Plankton Ecoliogy and Marine Biogeochemistry. Belgian Sci. Policy Office, Brussels, pp. I-10.
Laws, E.A., Bienfang, P.K., Ziemann, D.A. and Conquest, L.. 1988. Phytoplankton population dynamics and the fate of production during the spring bloom in Agke Bay, Alaska. Limnol. Oceanogr., 33: 57-65.
Leaky, R.J.G., Fenton, N. and Clarke, A., 1993. The annual cycle of planktonic ciliates in nearshore waters at Signy Island, Antarctica. J. Plankton Res., 16: 841-856.
Lee, S. and Fuhrman, J.A., 1987. Relationships between biovol- ume and biomass of naturally Jerived marine bacterioplank- ton. Appl. Environ. Microbial., 53: 1298-1303.
Legendre, L., 1990. The significance of microalgal blooms for fisheries and for the export of particulate organic carbon in oceans. J. Plankton Res., 12: 68 l-699.
Legendre, L. and Rassoulzadegan, F., 1995. Planiclon and nlrtrient dynamics in marine waters. Ophelia, 4 1: 153- 172.
Leventer, A. and Dunbar, R.B., 1987. Diatom flux in McMurdo Sound, Antarctica. Mar. Micropaleontol., 12: 49-64.
Leynaert, A., Triguer, P., QuCguigner, B. and Morvan, J., 199 1, The distribution of biogenic silica and the composition of particulate organic matter in the Weddell-Scotia Sea during spring 1988. Mar. Chem., 35: 435-447.
Li, W.K.W., Subba Rao, D.V., Harrison, W.G., Smith. J.C., Cullen. J.J., Irwin, B. and Platt, T., 1983. Autotrophic pi- coplankton in the tropical ocean. Science, 219: 292-95.
Lopez, M.D.G.. Huntley, M.E. and Sykes, P.F., 1988. Pigmellt destruction by Culunus pucijkus: impact on the estimation of water column fluxes. J. Plankton Res., 10: 715-734.
Lund, J.W.G., Kipling, C. and Le Cren, E.D., 1958. The inverted microscope method of estimating algal cell numbers and the statistical basis of estimations by counting. Hydrobiology, 11: 143-170.
Marschall, H.P., 1988. The overwintering strategy of Antarctic krill under the pack-ice of the Weddell Sea. Polar Biol., 9: 129-135.
Matsuda, O., Ishikawa, S. and Kawaguchi, K., 1987. Seasonal variation of downward flux of particulate organic matter under the antarctic fast ice. Proc. NIPR Symp. Polar Biol., 1: 23-34.
Meguro, H., Ito, K. and Fukushima, H., 1967. Ice flora (bottom type): a mechanism of primary production in the polar seas and the growth of diatoms in sea ice. Arctic, 20: 114-133.
Michaels, A.F. and Silver, M.W., 1988. Primary production, sinking fluxes and the microbial food web. Deep-Sea Res., 35: 473-490.
Morales, C.E., 1987. Carbon and nitrogen content of copepod faecal pellets: effects of food concentration and feeding behav- ior. Mar. Ecol. Prog. Ser., 36: 107- 114.

and Gordon, L.I., 1982. reduction and pelagic of biogenic silica in the So
Cosmochim. Acta. 46: 49 l-50 1. , Gordon, L.I., Sulh-
ice-edge articulate matter and
nutrient distributions in the zone of the Weddell Sea: relationship to hydrography during late summer. Deep-Sea Res., 36: 191-209.
Niithig, E.-M. and Von Bodungen, B., 1989. Occurrence and vertical flux of faecal pellets of probably protozoan origin in the southeastern Weddell Sea (Antarctica). Mar. Ecol. Prog. Ser., 56: 281-289.
Paasche, E., 1980. Silicon content of five marine plankton diatom species measured with a rapid filter method. Limnol. Oceanogr., 25: 474-480.
Parsons, T.R., Maita, Y. and Lalli, C.M., 1984. A Manual of Chemical and Biological Methods for Seawater Analysis. Pergamon Press, Toronto, 173 pp.
Platt, T., Subba Rao, D.V. and Irwin, B., 1983. Photosynthesis of picoplankton in the oligotrophic ocean. Nature, 301: 702-704.
Pomeroy, L.R. and Deibel, D., !986. Temperature regulation of bacterial activity during the spring bloom in Newfoundland coastal waters. Science, 233: 359-36 1.
Pomeroy, L.R. and Wiebe, W.J., 19ti8. Energetics of microbial food webs. Hydrobiology, 159: 7- 18.
Redfield, AC., Ketchum, B.H. and Richards, F.A., 1963. The influence of organisms on the composition of sea-water. In: M.N. Hill (Editor), The Sea, Vol. 2. Interscience, New York, NY, pp. 26-77.
Riley, GA., 1942. The relationship of vertical turbulence and spring diatom flowerings. J. Mar. Res., 5: 67-87.
Runge, J.A. and Iugram, KG., 1988. Underice grazing by plank- tonic, calanoid copepods in relation to a bloom of ice microal- gae in southeastern Hudson Bay. Limnol. Oceanogr.. 33: 280-286.
Runge, J.A. and Ingram, R.G., 1991. Under ice feeding and die1 migration by the planktonic copepods Calanus glacialis and Pseudoculunus rninutus in relation to the ice algal production cycle in southeastern Hudson Bay. Mar. Biol., 108: 217-225.
Saito, H. and Hattori, H., 1997. Die1 vertical migration and feeding rhythm of copepods under sea ice at Saroma-ko Lagoon. J. Mar. Syst., 11: 191-204.
Sasaki, H. and Fukuchi, M., 1993. Production and sedimentation processes in the ice-covered Lake Saroma. Bull. Plankton Sot. Jpn., 39: 170-171.
Sasaki, H. and Hoshiai, T., 1986. Sedimentation of microalgae under the antarctic fast ice in summer. Mem. Natl. Inst. Polar Res. Spec. Issue, 40: 45-55.
Satoh, H., Yarnaguchi, Y., Watanabe, K., Tanimura, A., Fukuchi, M. and Aruga, Y., 1989. Photosynthetic nature of ice algae and their contribution to the primary production fq Lagoon Saroma Ko, Hokkaido, Japan. Proc. NIPR Symp. Polar Biol., 2: i-8.
Schandelmeier, L. and Alexander, V., 1981. An analysis of the influence of ice on spring phytoplankton population structure in the southeastern Bering Sea. Limnol. Qceanogr.. 26: 935- 943.
1993. Variations in atmos~~e~c conditions and ice formations in Lake Saroma. ~a~kto~ Sot. Jpn.. 39: 156-158.
Shi
Jshikawa, N., 1993. Atmospheric and port for Saroma-ko lagoon of the SA Studies) project. Low Temp. Sci. Ser.
Silver, M.W., Mitchell, J.G. and Ringo nannoplankton. IL Newly discovere choanoflagellates from the Weddell Sea, Antarctica. Mar. Biol., 58: 211-217.
Sime-Ngando, T., Juniper, SK. and Demers, S., 1997. Ice-brine and planktonic microheterotrophs from Saroma-ko Lagoon, Hokkaido (Japan): quantitative importance and trophodynam- its. J. Mar. Syst., 11: 149- 161.
king and floating of phytoplankton in Biol. Annu. Rev., 8: 353-414.
Smayda, T.J., 1971. al and accelerated sinking of phyto- plankton in the sea. Mar. Geol., 11: 105-122.
Smetacek. V., 1980. Armual cycle of sedimentation in relation to plankton ecology in Western Kiel Bight. Ophelia, Suppl. 1: 65-76.
Smetacek, V., 1985. Role of sinking in diatom life-history cycles: ecological, evolutionary and geological significance. Mar. Biol., 84: 239-251.
Smetacek, V., Scharek, R. and Niithig, E.-M., 1990. Seasonal and regional variations in the pelagial and its relationship to the life history cycle of krill. in: R. Kemy and G. Hempel (Editors), Antarctic Ecosystems: Ecological Change and Con- servations. Springer, Berlin, pp. 103- 114.
Smetacek, V., Scharek, R., Gordo , L.I., Eicken, H., Fahrbach, E., Rohardt, G. and Moore, S., 1992. Early spring phytoplankton blooms in ice platelet layers of the southern Weddell Sea, Antarctica. Deep-Sea Rcs., 39: 153- 168.
Smith, W.O. and Nelson, D.M., 1985. Phytoplankton bloom pro- duced by a receding ice edge in the Ross Sea: Spatial coher- ence with the density field. Science, 227: 163 -166.
Smith, W.O. and Nelson, D.M., 1986. importance of ice edge phytoplankton production in the Southern Ocean. Bioscience, 36: 25 1-257.
Stretch, J.J., Hamnsr, P.P., Hamner, WM., Michef, W.C., Cook, J. and Sullivan, C.W., 1988. Foraging behavior of antarctic krilll Euphuusia superbu on sea ice microalgae. Mar. Ecol. Prog. Ser., 44: 131-139.
Strickland, J.D.H. and Parsons, T.R., 1972. A practical handbook of seawater analysis. Fish. Res. Bd. Can., Bull. 167, 310 pp.
Strom, S.L., 1993. Production of pheopigments by marine prom- zoa: results of laboratory experiments analysed by HPLC. Deep-Sea Res .40: 57-80.
Sverdrup, H.U., 1953. On conditions for the vernal blooming of phytoplankton. J. Cons. Perm. Int. Explor. Mer, 18: 287-295.
Tada, K., Kurata, M. and Nishihama, Y., 1993. Seasonal changes of chlorophyll a and nutrients in Lake Saroma. Bull. Plankton Sot. Jpn., 39: 163-165.
T&&as& E., 1981. moru&c study of ice a@ from tk ~2 ice

148 C. Michel et al. / Journal of Marine Systenls I I f 1997) 133-148
of a lagoon, Lake Saroma, Hokkaido. Mem. Natl. Inst. Polar Res. Ser. E, 34: 49-56.
Tremblay, C., Runge, J.A. and Legendre, L., 1989. Grazing and sedimentation of ice a!gac d~uring and mumediately after a bloom at the ice-water interface. Mar. Ecol. Prog. Ser., 56: 291-300.
Verity, P.G. and Langdon, C., 1984. Relationships between Iorica volume, carbon, nitrogen, and ATP content of tintinnids in Narragansett Bay. J. Plankton Res., 6: 859-868.
Von Bodungen, B., Smetaceck, VS., Tilzer, M.M. and Zeitzschel, B., 1986. Primary production and sedimentation during spring in the Antarctic Peninsula region. Deep-Sea Res., 33: 177-194.
Von Bodungen, B., Niithig, E.-M. and Sui, Q., 1988. New produc-
tion of phytoplankton and sedimentation during summer 1985 in the southeastern Weddeli Sea. Comp. B&hem. Physiol., 9OB: 425-487.
Wassmann, P., Peinert, R. and Smctacek, V., 1991. Patterns of production and sedimentation in the boreal and polar North- east Atlantic. In: E. Sakshaug, C.C.E. Hopkins and N.A. @ritsland (Editors), Proc. Pro Mare Symposium on Polar Marine Ecology. Polar Research, Trondheim, 12-16 May 1990, pp. 209-228.
Watanabe, K., Satoh, H. and Yamaguchi, Y., 1993. Phytoplankton and ice algae in Lake Saroma. Bull. Plankton Sot. Jpn., 39: 16% 166.





























