Embed Size (px)
Citation preview

93 (2007) 51–57www.elsevier.com/locate/schres
Schizophrenia Research
CNS 14-3-3ζ: Changes with sex but not psychiatric diagnoses orpsychotropic drug treatment
Brian Dean a,c,d,e,⁎, Simone A. Boer a,b, Andrew Mackinnon f, Michael Berk a,b
a The Rebecca L. Cooper Research Laboratories, The Mental Health Research Institute, Parkville, Australiab The University of Melbourne Department of Clinical and Biomedical Sciences, Geelong, Australia
c The University of Melbourne Department of Psychiatry, Parkville, Australiad The University of Melbourne Department of Pathology, Parkville, Australia
e The Department of Psychological Medicine, Monash University, Clayton, Victoria, Australiaf Centre for Mental Health Research, Australian National University, Canberra, ACT, Australia
Received 2 October 2006; received in revised form 18 February 2007; accepted 27 February 2007Available online 30 March 2007
Abstract
mRNA for 14-3-3ζ, an abundant signalling protein in human CNS, is reported as decreased or unchanged in cortex fromsubjects with schizophrenia. Addressing this dichotomy, using Western blot analyses, we measured levels of 14-3-3ζ proteins incortex and caudate nucleus from subjects with schizophrenia, bipolar disorder, age/sex matched controls and in analogous CNSregions from rats treated with psychotropic drugs. Anti-14-3-3ζ antibody bound to three proteins (molecular weights: 27, 54 and70 kDa), in all CNS tissue. Levels of all proteins did not vary with diagnoses (27 kDa: F2,42.0=0.35, p=0.71; 54 kDa:F2,42.1=0.62, p=0.54; 70 kDa: F2,41.0=2.43, p=0.10). By contrast, independent of diagnoses, there were significant increases inthe levels of the 27 kDa protein (+32%; pb0.001) and 54 kDa protein (51%; p=0.001) in the caudate nucleus from malescompared to females. In addition, there was a trend (−25%; p=0.06) to decreased levels of the 70 kDa protein in BA 9 in malescompared to females. Treating with haloperidol, olanzapine, lithium or a combination thereof did not alter 14-3-3ζ levels in ratcortex or striatum. Therefore, this study suggests that 14-3-3ζ proteins are not altered in the cortex or caudate nucleus inschizophrenia, bipolar disorder or in analogous regions in psychotropic drug treated rats. By contrast, our study suggests that levelsof 14-3-3ζ in some regions of the human CNS may be modulated by some sex-specific mechanism.© 2007 Elsevier B.V. All rights reserved.
Keywords: Schizophrenia; Bipolar disorder; Sex; Antipsychotic drugs; Lithium; 14-3-3ζ
1. Introduction
Amicroarray study first suggested that the expressionof the 14-3-3 protein family was altered in the CNS from
⁎ Corresponding author. NHMRC Senior Research Fellow andHead, The Rebecca L. Cooper Research Laboratories, The MentalHealth Research Institute, Locked Bag 11, Parkville, Victoria 3052,Australia. Tel.: +61 3 9389 2940; fax: +61 3 9387 5061.
E-mail address: [email protected] (B. Dean).
0920-9964/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.schres.2007.02.022
subjects with schizophrenia (Middleton et al., 2005). The14-3-3 family of proteins are encoded by separate genesthat were named using Greek letters (β, ε, γ, η, σ, τ, ζ)according to elution order following reverse phasechromatography (Fu et al., 2000), with the α and δmembers being shown to be phosphorylated forms of βand ζ, respectively. 14-3-3 proteins are involved withintracellular signalling, cell division, cell differentiation,apoptosis, ion channel functioning and neurotransmis-sion (Berg et al., 2003). Therefore changes in any of the

52 B. Dean et al. / Schizophrenia Research 93 (2007) 51–57
14-3-3 proteins could have profound effects on CNSfunction and contribute to the onset of the symptoms ofpsychiatric diseases such as schizophrenia and bipolardisorder. The notion that changes in that 14-3-3 proteinscan result in diseases of the human CNS is supported byevidence implicating these proteins in the pathologies ofCreutzfeldt–Jacob disease, Alzheimer's disease, neuro-degenerative disorders with Lewy bodies and Polyglu-tamine disease (Berg et al., 2003).
In the microarray study in schizophrenia, the mostrobust change in 14-3-3 mRNAwas a decrease in levelsof mRNA for 14-3-3ζ in the dorsolateral prefrontalcortex (DFLPC) in subjects with schizophrenia (Mid-dleton et al., 2005). However, a second study usingquantitative PCR failed to detect changes in levels of 14-3-3ζ mRNA in the DFLPC from subjects withschizophrenia or bipolar disorder (Wong et al., 2005).These discrepant findings could be due to regional-specific changes in the DFLPC from subjects withschizophrenia, as the microarray study was in Brod-mann's area (BA) 9 (Middleton et al., 2005) while theqPCR study was in BA 46 (Wong et al., 2005). It is alsoof note that studies on 14-3-3ζ in schizophrenia werecompleted using tissue from subjects treated withantipsychotic drugs. It is therefore significant that astudy has shown increased levels of 14-3-3ζ protein inCSF from subjects with mixed dementias after treatmentwith either typical or atypical antipsychotic drugs(Zanusso et al., 2005). Contrasting these findings is astudy that failed to show any change in levels of cortical14-3-3ζ mRNA after treating primates with antipsy-chotic drugs (Middleton et al., 2005). It is thereforeunclear if treatment with antipsychotic drugs could bethe cause of changes in levels of 14-3-3ζ levels insubjects with schizophrenia.
To try and resolve discrepancies raised by earlierstudies, we measured levels of 14-3-3ζ protein in theDLPFC (BA 9), frontal pole (BA 10) and caudatenucleus from subjects with schizophrenia, bipolardisorder and age and sexed matched control subjects.In addition, levels of 14-3-3ζ proteins were measured inthe cortex and striatum of rats treated with antipsychoticdrugs, the mood stabiliser lithium, as well as combinedtreatment with antipsychotic drugs and lithium.
2. Experimental materials and methods
2.1. Human tissue collection, clinical follow up anddiagnosis
Ethical approval for these studies was obtained fromthe Ethics Committee of the Victorian Institute of
Forensic Medicine and the North Western MentalHealth Program Behavioural and Psychiatric Researchand Ethics Committee.
Psychiatric diagnoses were made according to DSM-IV criteria (American Psychiatric Association, 1994), byconsensus between two senior psychiatrists and apsychologist, following extensive case history review(using a structured diagnostic instrument, the DiagnosticInstrument for Brain Studies (DIBS) (Hill et al., 1996)).Subsequently, duration of illness (DOI) was calculatedas the time from first contact with a psychiatric serviceto death. The final recorded dose of antipsychotic drug(FRADD) was converted to chlorpromazine equiva-lents. In cases where death was witnessed, the timebetween death and autopsy was taken as the postmorteminterval (PMI). Where death was not witnessed, tissuewas only taken from individuals who had been seenalive up to 5 h before being found dead, in these casesPMI was taken as the interval half way between thedonor being found dead and last being seen alive. In allcases, cadavers were refrigerated within 5 h of beingfound and tissue was rapidly frozen to −70 °C within30 min of autopsy. The pH of the CNS tissue wasmeasured as described previously (Kingsbury et al.,1995) because such a measure provides a betterindication of tissue integrity than PMI (Torrey et al.,2000).
2.2. Rat treatments and tissue collection
After obtaining authorization from the University ofMelbourne Animal Experimentation and Ethics Com-mittee, six week old male Sprague–Dawley rats weredivided into 6 treatment groups with 10 rats in eachgroup. Rats receiving plain chowwere either treated withvehicle (1% ethanol, 0.9% sodium chloride), haloperidol(0.1 mg/kg/day dissolved in ethanol) or olanzapine(1 mg/kg/day dissolved in ethanol) in drinking water or aspecial chow containing lithium (1.7 mg/kg/day). Twogroups received lithium in rat chow, and eitherolanzapine or haloperidol in drinking water. All drugswere delivered for 4 weeks while the animals werehoused in normal animal house conditions. Aftertreatment, rats were decapitated and the brains wereremoved and rapidly frozen in isopentane on dry ice.
2.3. Human tissue preparation
Tissue was obtained from the left hemisphere of 20subjects with a DSM IV diagnosis of schizophrenia,8 subjects with a diagnosis of bipolar 1 disorder and 20control subjects (Supplementary material online). There

53B. Dean et al. / Schizophrenia Research 93 (2007) 51–57
was no significant variation in age (F2,45=0.76, p=0.47),sex (χ1
2=0.07, p=0.97), PMI (F2,45=0.03, p=0.97) orCNS pH (F2,45=0.39, p=0.68) across the diagnosticcohorts. A Mann–Whitney U test showed that levels ofantipsychotic drugs prescribed close to death, whenexpressed as chlorpromazine equivalents, did not differbetween the subjects with schizophrenia and those withbipolar disorder (U=17.50, p=0.37).
Cortical tissue dissection was completed using grosslandmarks to define cytoarchitectonic regions; henceBA9 was taken as the region of the CNS on the lateralsurface of the frontal lobe and includes the middlefrontal gyrus superior to the inferior frontal sulcus.BA10 was taken as the most rostral portions of thesuperior frontal gyrus and middle frontal gyrus,bounded ventrally by the superior rostral sulcus andcaudate nucleus. Tissue was taken from a defined area ofthe caudate nucleus rostral to the anterior commissure.
2.4. Rat tissue preparation
Tissue was obtained from the right hemisphere of ratCNS. Tissue was taken from both the cortex andstriatum, approximately 1.6 mm anterior to the Bregma.
2.5. Western blot analysis
Human and rat tissue was homogenised in a glassTeflon homogeniser (5×vol. tissue g wht) for 20 strokesinto 10 mM Tris containing 1% sodium dodecylsulphate and 1mM sodium orthovanidate (pH 7.4).The protein concentration in each homogenate was thenmeasured using the modified BioRad Detergent Com-patible assay (Lowry method). An aliquot of eachhomogenate was diluted in reducing buffer (0.5M Tris–HCl, pH 6.8, 20% glycerol, 10% SDS, 10% 2-β-mercaptoethanol, and 0.05% Bromophenol Blue) to10 μg protein, heated for 4 min at 80 °C and thenresolved in duplicate on a 12% SDS gel. Each gel alsocontaining a BioRad low range standard (MW markers:2 lanes) and internal control (2 lanes: see below) at aconstant voltage of 150 V.
The proteins in each gel were transferred ontonitrocellulose membrane (Hybond-ECL) at a constant40 mA, overnight. Each nitrocellulose membrane wasstained with Ponceau S to ensure proteins hadtransferred. After destaining, the nitrocellulose mem-branes were washed in 0.1% tween-20 in Tris-bufferedsaline (TTBS) for 1 h at r/t, incubated with TTBScontaining 1:500 dilution of a rabbit anti-human 14-3-3ζantibody (IBL, Japan: cat. # 18644) in TTBS o/n at 4 °C,washed 4×5 min in TTBS, and then incubated with a
DAKO goat anti-rabbit IgG antibody (cat. #: P-0448)diluted 1:2000 in TTBS for 2 h at r/t. After washing for4×5 min in TTBS, the nitrocellulose membranes wereexposed to Pierce ECL for 5 min at r/t and then imagedusing a Kodak 440CF image station. Two images werecaptured (light mode to show the location of the MWstandards and chemiluminescent mode to show theimmunogenic bands) and merged to allow the molecularweights of each antigenic band to be estimated. Thedensities of the immunogenic bands were measuredusing the chemiluminescent image and then expressedas a ratio of the corresponding immunogenic bands inthe internal control to accommodate gel-to-gel variation(see below).
Prior to measuring 14-3-3ζ in the samples, sufficientprotein homogenate (Internal Control: IC) was preparedfrom the frontal cortex of a subject with no history ofpsychiatric or neurological illness. Initially, this homog-enate was run in each of 12 wells on 2 gels (24 samplesover all), and the OD of each immunopositive 14-3-3ζband was measured as described above. These experi-ments revealed that the anti-human 14-3-3ζ antibodybound to three proteins of molecular weights 27, 54 and70 kDa, respectively (see below). Using data from theIC, the intra-gel variation was shown to be 8.7% for the27 kDa protein, 9.3% for the 54 kDa protein and 11%for the 70 kDa protein. The inter-gel variation was 10%for the 27 kDa protein, 25% for the 54 kDa protein and19% for the 70 kDa protein. A sample of IC wasincluded in two lanes of each subsequent gel on whichprotein from samples were separated and the OD of eachsample was expressed as a ratio of the IC to control forgel to gel variation in our analyses.
To measure the relative contribution of each immu-nogenic protein to the overall measurement of 14-3-3ζ,the sum intensity of each band was totalled and thepercent contribution of each individual band to the totalsum intensity was calculated.
3. Statistics
Each data set was tested for Gaussian distributionusing the D'Agostino and Pearson omnibus normalitytest. Where data conformed to an approximate Gaussiandistribution, the following statistical strategies wereadopted. Variation in demographic and tissue collectiondata across diagnosis was assessed using a one-wayANOVA. Comparisons of pharmacological data betweenpsychiatric diagnoses, the impact of suicide, “on” and“off” drug status and sex on experimental measures wereachieved using t-tests. Relationships between experimen-tal parameters and potential confounding factors were
54 B. Dean et al. / Schizophrenia Research 93 (2007) 51–57
analysed using Pearson product moment correlations witha Bonferroni correction for multiple analyses. In caseswhere data did not approximate a Gaussian distribution,equivalent non-parametric tests were utilised. All theseanalyses were completed using GraphPad Prism Version4.0. Variation in means between diagnosis and CNSregion was assessed using mixed model repeatedmeasures ANOVAwith post hoc analysis used to identifythe source of the variation. These analyses included sex asa factor in order to assess its impact on protein expressionwithin regions and diagnoses. This technique is anadvance on conventional ANOVA as it recognizes thatcorrelations between brain regions may not be uniform(Gueorguieva and Krystal, 2004). These analyses werecompleted using SPSS v14 MIXED procedure.
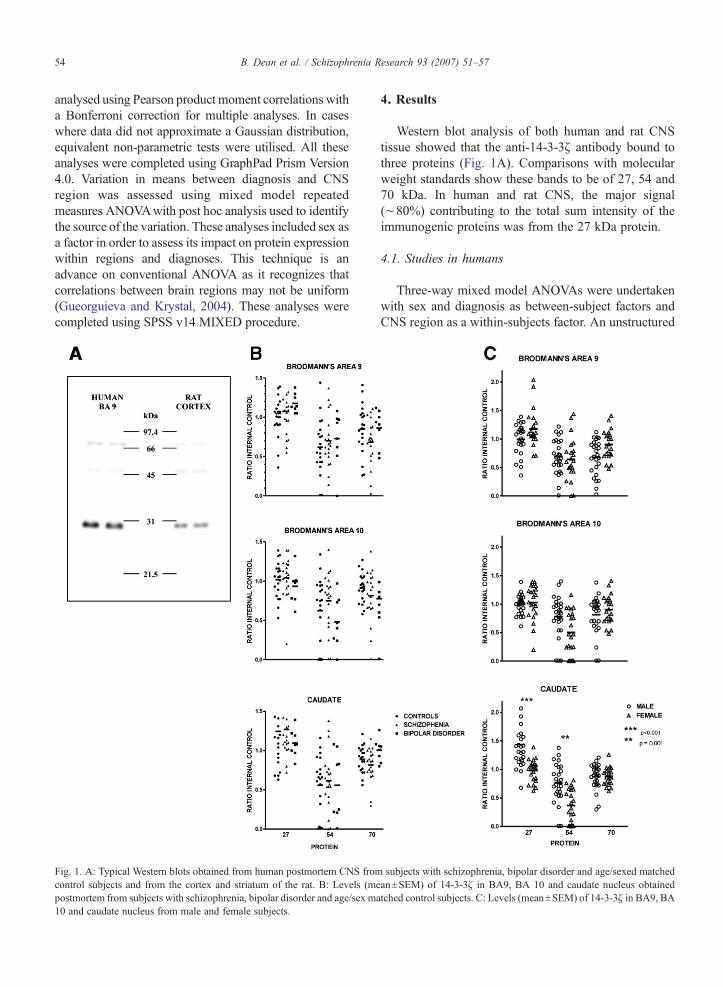
Fig. 1. A: Typical Western blots obtained from human postmortem CNS fromcontrol subjects and from the cortex and striatum of the rat. B: Levels (mepostmortem from subjects with schizophrenia, bipolar disorder and age/sex ma10 and caudate nucleus from male and female subjects.
4. Results
Western blot analysis of both human and rat CNStissue showed that the anti-14-3-3ζ antibody bound tothree proteins (Fig. 1A). Comparisons with molecularweight standards show these bands to be of 27, 54 and70 kDa. In human and rat CNS, the major signal(∼80%) contributing to the total sum intensity of theimmunogenic proteins was from the 27 kDa protein.
4.1. Studies in humans
Three-way mixed model ANOVAs were undertakenwith sex and diagnosis as between-subject factors andCNS region as a within-subjects factor. An unstructured
subjects with schizophrenia, bipolar disorder and age/sexed matchedan±SEM) of 14-3-3ζ in BA9, BA 10 and caudate nucleus obtainedtched control subjects. C: Levels (mean±SEM) of 14-3-3ζ in BA9, BA
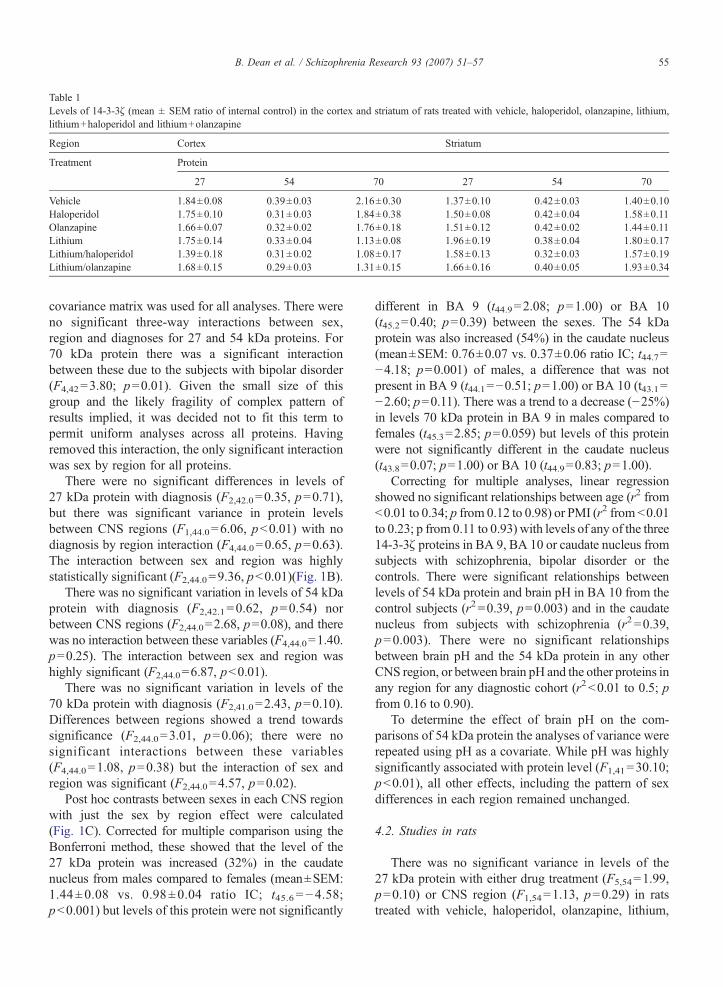
Table 1Levels of 14-3-3ζ (mean ± SEM ratio of internal control) in the cortex and striatum of rats treated with vehicle, haloperidol, olanzapine, lithium,lithium+haloperidol and lithium+olanzapine
Region Cortex Striatum
Treatment Protein
27 54 70 27 54 70
Vehicle 1.84±0.08 0.39±0.03 2.16±0.30 1.37±0.10 0.42±0.03 1.40±0.10Haloperidol 1.75±0.10 0.31±0.03 1.84±0.38 1.50±0.08 0.42±0.04 1.58±0.11Olanzapine 1.66±0.07 0.32±0.02 1.76±0.18 1.51±0.12 0.42±0.02 1.44±0.11Lithium 1.75±0.14 0.33±0.04 1.13±0.08 1.96±0.19 0.38±0.04 1.80±0.17Lithium/haloperidol 1.39±0.18 0.31±0.02 1.08±0.17 1.58±0.13 0.32±0.03 1.57±0.19Lithium/olanzapine 1.68±0.15 0.29±0.03 1.31±0.15 1.66±0.16 0.40±0.05 1.93±0.34
55B. Dean et al. / Schizophrenia Research 93 (2007) 51–57
covariance matrix was used for all analyses. There wereno significant three-way interactions between sex,region and diagnoses for 27 and 54 kDa proteins. For70 kDa protein there was a significant interactionbetween these due to the subjects with bipolar disorder(F4,42=3.80; p=0.01). Given the small size of thisgroup and the likely fragility of complex pattern ofresults implied, it was decided not to fit this term topermit uniform analyses across all proteins. Havingremoved this interaction, the only significant interactionwas sex by region for all proteins.
There were no significant differences in levels of27 kDa protein with diagnosis (F2,42.0=0.35, p=0.71),but there was significant variance in protein levelsbetween CNS regions (F1,44.0=6.06, pb0.01) with nodiagnosis by region interaction (F4,44.0=0.65, p=0.63).The interaction between sex and region was highlystatistically significant (F2,44.0=9.36, pb0.01)(Fig. 1B).
There was no significant variation in levels of 54 kDaprotein with diagnosis (F2,42.1=0.62, p=0.54) norbetween CNS regions (F2,44.0=2.68, p=0.08), and therewas no interaction between these variables (F4,44.0=1.40.p=0.25). The interaction between sex and region washighly significant (F2,44.0=6.87, pb0.01).
There was no significant variation in levels of the70 kDa protein with diagnosis (F2,41.0=2.43, p=0.10).Differences between regions showed a trend towardssignificance (F2,44.0=3.01, p=0.06); there were nosignificant interactions between these variables(F4,44.0=1.08, p=0.38) but the interaction of sex andregion was significant (F2,44.0=4.57, p=0.02).
Post hoc contrasts between sexes in each CNS regionwith just the sex by region effect were calculated(Fig. 1C). Corrected for multiple comparison using theBonferroni method, these showed that the level of the27 kDa protein was increased (32%) in the caudatenucleus from males compared to females (mean±SEM:1.44±0.08 vs. 0.98±0.04 ratio IC; t45.6 =−4.58;pb0.001) but levels of this protein were not significantly
different in BA 9 (t44.9=2.08; p=1.00) or BA 10(t45.2=0.40; p=0.39) between the sexes. The 54 kDaprotein was also increased (54%) in the caudate nucleus(mean±SEM: 0.76±0.07 vs. 0.37±0.06 ratio IC; t44.7=−4.18; p=0.001) of males, a difference that was notpresent in BA 9 (t44.1=−0.51; p=1.00) or BA 10 (t43.1=−2.60; p=0.11). There was a trend to a decrease (−25%)in levels 70 kDa protein in BA 9 in males compared tofemales (t45.3=2.85; p=0.059) but levels of this proteinwere not significantly different in the caudate nucleus(t43.8=0.07; p=1.00) or BA 10 (t44.9=0.83; p=1.00).
Correcting for multiple analyses, linear regressionshowed no significant relationships between age (r2 fromb0.01 to 0.34; p from 0.12 to 0.98) or PMI (r2 from b0.01to 0.23; p from 0.11 to 0.93) with levels of any of the three14-3-3ζ proteins in BA 9, BA 10 or caudate nucleus fromsubjects with schizophrenia, bipolar disorder or thecontrols. There were significant relationships betweenlevels of 54 kDa protein and brain pH in BA 10 from thecontrol subjects (r2=0.39, p=0.003) and in the caudatenucleus from subjects with schizophrenia (r2 =0.39,p=0.003). There were no significant relationshipsbetween brain pH and the 54 kDa protein in any otherCNS region, or between brain pH and the other proteins inany region for any diagnostic cohort (r2b0.01 to 0.5; pfrom 0.16 to 0.90).
To determine the effect of brain pH on the com-parisons of 54 kDa protein the analyses of variance wererepeated using pH as a covariate. While pH was highlysignificantly associated with protein level (F1,41=30.10;pb0.01), all other effects, including the pattern of sexdifferences in each region remained unchanged.
4.2. Studies in rats
There was no significant variance in levels of the27 kDa protein with either drug treatment (F5,54=1.99,p=0.10) or CNS region (F1,54=1.13, p=0.29) in ratstreated with vehicle, haloperidol, olanzapine, lithium,

56 B. Dean et al. / Schizophrenia Research 93 (2007) 51–57
lithium+haloperidol or lithium+olanzapine (Table 1)and there was no significant interaction between thevariables (F5,54=2.21, p=0.07). There was significantvariation in levels of 54 kDa between CNS regions(F1,54=12.90, pb0.001) but not with drug treatment(F5,54=1.70, p=0.15); there was no interaction betweenthese two variables (F5,54=0.68, p=0.64). Finally, therewas no significant variation in levels of 70 kDa proteinbetween CNS regions (F1,54=0.50, p=0.48) or withdrug treatments (F5,54=1.10, p=p=0.37) but there wasa significant interaction between these two variables(F5,54=3.6, pb0.01).
5. Discussion
This study has shown that an anti-human 14-3-3ζbinds to three proteins of molecular weights of 27, 54and 70 kDa, in both human and rat CNS. Themonomeric molecular mass of 14-3-3ζ is 27.7 kDa(Fu et al., 2000) and therefore the lower molecular massprotein identified in our studies must be monomeric 14-3-3ζ. 14-3-3ζ predominantly exist as monomers anddimers in the CNS (Fu et al., 2000) and therefore the54 kDa protein measured in this study likely representsdimers of 14-3-3ζ. The identity of the 70 kDa protein isnot clear but it is known that dimers of 14-3-3ζ bind toother target molecules (Coblitz et al., 2006) andtherefore the 70 kDa may be 14-3-3ζ bound to a targetprotein. Significantly, the levels of none of these threeproteins differed significantly in three CNS regions fromsubjects with schizophrenia, bipolar 1 disorder orcontrols or in analogous CNS regions from rats treatedwith antipsychotic drugs, the mood stabiliser lithium ora combination of those drugs.
Two previous studies on 14-3-3ζ in postmortem CNSreported an 18.2% decrease in the levels of 14-3-3æmRNA in BA 9 from subjects with schizophrenia(Middleton et al., 2005) and no difference in levels of14-3-3ζ mRNA in BA 46 from subjects with schizo-phrenia and bipolar 1 disorder (Wong et al., 2005),respectively. Our study adds to data from the latter studyto suggest that 14-3-3ζ proteins are not altered in theCNS of subjects with schizophrenia or bipolar 1 disorder.However, it does remain possible that altered levels of14-3-3ζ mRNA in BA9 from subjects with schizophre-nia have not translated into changed levels of 14-3-3ζprotein. Moreover, it is also possible that signallingthrough 14-3-3ζ protein could vary in the CNS ofsubjects with schizophrenia or bipolar 1 disorder withouta change in levels of these proteins per se.
One potential confounding factor in a study of humanpostmortem CNS is the impact of PMI on protein
integrity. Significantly, a 2 dimensional electrophoresisstudy has reported that levels of 14-3-3ε and 14-3-3γ aresignificantly increased in rat brain left at roomtemperature for more than 48 h (Fountoulakis et al.,2001). As 14-3-3ζ was not reported to be affected byPMI in this model of postmortem delay, this study lendssupport to our finding that PMI does not impact onlevels of 14-3-3ζ in brain tissue.
This study suggests there may be complex sex-related changes in levels of 14-3-3ζ in different CNSregions. In particular, we have shown significantincreases in levels of what are likely to be monomersand dimers of 14-3-3ζ in the caudate-nucleus in malescompared to females that were independent of psychi-atric diagnoses. A more comprehensive study of 14-3-3ζis required to fully elucidate the extent of sex-relatedchanges in 14-3-3 proteins in human CNS. Unfortu-nately, neither of the earlier studies on 14-3-3 proteins inpsychiatric illnesses has reported an analyses of 14-3-3ζdata by sex (Middleton et al., 2005; Wong et al., 2005).Moreover, a recent comprehensive review of 14-3-3proteins in the nervous system (Berg et al., 2003) makesno mention of any data showing variation in levels ofCNS 14-3-3ζwith sex. Therefore our finding may be thefirst to highlight this phenomenon. One explanation forour findings on 14-3-3ζ protein and sex comes fromresearch in cancer showing that 14-3-3 proteins aredownstream targets for estrogen receptors (Ikeda andInoue, 2004). These data could therefore indicate thatthe sex differences we observed in levels of 14-3-3ζcould be related to differences in sex hormone activity.
Data in this study has also shown that treatment withtypical and atypical antipsychotic drugs, a moodstabiliser or a combination of these two classes ofdrugs does not affect levels of 14-3-3ζ in the rat cortexor striatum. These findings are in agreement with a studyin cynomolgous monkeys (Middleton et al., 2005),which showed that treatment with haloperidol did notaffect levels of cortical 14-3-3ζ.
In conclusion, this study has failed to show changes inlevels of 14-3-3ζ proteins in a number of CNS regionsfrom subjects with schizophrenia or bipolar disorder.However, this study suggests that levels of 14-3-3ζproteins may differ in males and females. In addition,levels of cortical and striatal 14-3-3ζ were not alteredfollowing treatment with either antipsychotic drugs orlithium alone, or combined treatment with antipsychoticdrugs and lithium. Therefore, overall, our data does notsuggest changes in the levels of 14-3-3ζ contribute tochanges in themolecular cytoarchitecture in schizophreniaor bipolar 1 disorder or to the mechanisms of action ofantipsychotic drugs or mood stabilisers.

57B. Dean et al. / Schizophrenia Research 93 (2007) 51–57
6. Role of Funding Source
Funding for this study was provided by the NHMRC,The Australian Rotary Health Research Fund and TheRebecca L. Cooper Medical Research Foundation. Noneof the funding bodies had any further role in the studydesign; in the collection, analysis and interpretation ofdata, in the writing of the report or in the decision tosubmit the paper for publication.
7. Contributors
Author Brian Dean was involved in the design of thestudy, undertook data analysis and wrote the manuscript.Author Simone Boer was involved in the design of thestudy, completed the experimental protocols andassisted by editing each draft of the manuscript. AuthorAndrew Mackinnon undertook or reviewed statisticalanalyses of the data and edited drafts of the manuscript.Author Michael Berk was involved in the design of thestudy and in editing all drafts of the manuscript. Allauthors have contributed and approved the finalmanuscript.
Acknowledgement
BD is an NHMRC Senior Research Fellowship(# 400016) and SB is the recipient of an Ian ScottScholarship from The Australian Rotary Health Re-search Fund. The study was also supported by NHMRCProject Grant # 114253, NIMH Grant # MH069691-01A1 and The Rebecca L. Cooper Medical ResearchFoundation.
Appendix A. Supplementary data
Supplementary data associated with this article canbe found, in the online version, at doi:10.1016/j.schres.2007.02.022.
References
American Psychiatric Association, 1994. Diagnostic and StatisticalManual of Mental Disorders, 4 edn. American PsychiatricAssociation, Washington, D.C.
Berg, D., Holzmann, C., Riess, O., 2003. 14-3-3 proteins in thenervous system. Nat. Rev., Neurosci. 4, 752–762.
Coblitz, B., Wu, M., Shikano, S., Li, M., 2006. C-terminal binding: anexpanded repertoire and function of 14-3-3 proteins. FEBS Lett.580, 1531–1535.
Fountoulakis, M., Hardmeier, R., Hoger, H., Lubec, G., 2001.Postmortem changes in the level of brain proteins. Exp. Neurol.167, 86–94.
Fu, H., Subramanian, R.R., Masters, S.C., 2000. 14-3-3 proteins:structure, function, and regulation. Annu. Rev. Pharmacol.Toxicol. 40, 617–647.
Gueorguieva, R., Krystal, J.H., 2004. Move over ANOVA: progress inanalyzing repeated-measures data and its reflection in paperspublished in the Archives of General Psychiatry. Arch. Gen.Psychiatry 61, 310–317.
Hill, C., Keks, N., Roberts, S., Opeskin, K., Dean, B., Mackinnon, A.,Copolov, D., 1996. Problem of diagnosis in postmortem brainstudies of schizophrenia. Am. J. Psychiatry 153, 533–537.
Ikeda, K., Inoue, S., 2004. Estrogen receptors and their downstreamtargets in cancer. Arch. Histol. Cytol. 67, 435–442.
Kingsbury, A.E., Foster, O.J., Nisbet, A.P., Cairns, N., Bray, L., Eve,D.J., Lees, A.J., Marsden, C.D., 1995. Tissue pH as an Indicator ofmRNA Preservation in Human.
Middleton, F.A., Peng, L., Lewis, D.A., Levitt, P., Mirnics, K., 2005.Altered expression of 14-3-3 genes in the prefrontal cortex ofsubjects with schizophrenia. Neuropsychopharmacology 30,974–983.
Torrey, E.F., Webster, M., Knable, M., Johnston, N., Yolken, R.H.,2000. The Stanley Foundation brain collection and neuropathologyconsortium. Schizophr. Res. 44, 151–155.
Wong, A.H., Likhodi, O., Trakalo, J., Yusuf, M., Sinha, A., Pato, C.N.,Pato, M.T., Van Tol, H.H., Kennedy, J.L., 2005. Genetic and post-mortem mRNA analysis of the 14-3-3 genes that encodephosphoserine/threonine-binding regulatory proteins in schizo-phrenia and bipolar disorder. Schizophr. Res. 78, 137–146.
Zanusso, G., Fiorini, M., Farinazzo, A., Gelati, M., Benedetti, M.D.,Ferrari, S., Dalla, L.A., Capaldi, S., Monaco, H.L., Rizzuto, N.,Monaco, S., 2005. Phosphorylated 14-3-3zeta protein in the CSFof neuroleptic-treated patients. Neurology 64, 1618–1620.





























