Embed Size (px)
Citation preview

FEMS Microbiology Letters 43 (1987) 173-176 173 Published by Elsevier
FEM02840
Clostridium thermoaceticum forms methanol from carbon monoxide in the presence of viologen dyes
Hil t rud White, H e r b e r t Leber tz , Io rdanes Th an o s and H e l m u t S imon
Lehrstuhl fur Organische Chemie und Biochemie, Technische Universitiit MiJnchen, D-8046 Garching, F R. G.
Received 10 March 1987 Revision received 30 March 1987
Accepted 31 March 1987
Key words: Clostridium thermoaceticum; Carbon monoxide; Methanol; Viologen dye
1. SUMMARY
Whole cells of Clostridium thermoaceticum, crude extracts of such cells as well as the super- natant of 100000 x g centrifugations catalyse the reduction of carbon monoxide to methanol in the presence of viologens or cobalt sepulchrate. Without such a mediator methanol could not be detected. The reaction shows a marked optimum at pH 5. The incubation of [5-14C]methyltetrahy- drofolate led only to the formation of 14C-labeled ethanol; the radioactivity in methanol was negligi- ble. The reaction seems to be catalysed by carbon monoxide dehydrogenase.
2. I NTR ODUC TION
During our studies [1] on the use of carbon monoxide as an electron source for the reduction of different unsaturated compounds catalyzed by C. thermoaceticum, we observed the reduction of
very different carboxylates to the corresponding alcohols together with small amounts of methanol if the resting cells were shaken under an atmo- sphere of CO in the presence of 1-2 mM methyl- viologen or other viologens. Without reducible substrates, and under otherwise identical condi- tions the amount of methanol produced was much higher. In the absence of viologens or other medi- ators methanol was not formed. Surprisingly, in this methanol formation methyltetrahydrofolate does not seem to be an intermediate. The capabil- ity of methanol production is also shown by crude extracts of C. thermoaceticum and their mem- brane-free supernatant. Also a purified prepara- tion of carbon monoxide dehydrogenase from such C. thermoaceticum cells is able to form methanol in the presence of mediators and CO. To our knowledge such a methanol formation has not been published.
3. MATERIALS AND METHODS
Correspondence to: H. Simon, Lehrstuhl fiir Organische Chemie und Biochemie, Technische Universit~it Miinchen, D-8046 Garching, F.R.G.
Enzymes were obtained from Boehringer, Mannheim. [5-14C]methyltetrahydrofolate was purchased from Amersham. As stated by Amersham, their product is a mixture of the two
0378-1097/87/$03.50 © 1987 Federation of European Microbiological Societies

174
diastereoisomers at the 6-position. Only 50% of the material seems to be biologically active. The unlabeled methyltetrahydrofolate, methylviologen and cobalt sepulchrate were products from Sigma. Carbamoylmethylviologen was synthesized as de- scribed by us [2]. C. thermoaceticum DSM521 was obtained from Deutsche Sammlung von Mikroorganismen. The cells were grown on a 200-1 scale according to [3] with slight modifications, harvested and stored at -15 ° C.
In a standard experiment for methanol synthe- sis 400 mg wet packed cells were shaken in 3.0 ml 0.3 M phosphate buffer pH 5.5 containing 1 mM methylviologen or another mediator at 40 °C un- der CO atmosphere. Experiments with 0.3 M sodium formate as electron donor were conducted under an N atmosphere.
Crude extracts of cells were obtained by treat- ing 4.0 g wet packed cells in 26 ml phosphate buffer containing 4 mg lysozyme and 0.4 mg DNase for 30 rain at 25 o C followed by sonication for 100 s. The membrane-free supernatant was prepared by centrifugation at 100 000 x g for 1 h.
Carbon monoxide dehydrogenase was de- termined and purified according to [4,5]. Methanol and ethanol formation was measured by gas-liquid chromatography using packed columns filled with polyethylene glycol-400 adsorbed on Chromosorb G/AW-DMCS 80-100 mesh.
Methanol formation in the presence of 5114C]methyltetrahydrofolate was conducted by an incubation composed and treated as described above, however, in the presence of 1.0 /LCi [14C]methyltetrahydrofolate together with 0.5 ~mol unlabeled material. After 90 and 180 rain aliquots of 800 ffl were withdrawn and after ad- ding 100/~1 3 N sodium hydroxide, quantitatively lyophilized in a small vacuum device chilled with liquid nitrogen. The dry residue as well as the distillate were analyzed for radioactivity. In a control experiment a cell suspension boiled for several rain was used. To 300 ffl of the volatile fraction of the 14C-labeled material 25/L1 ethanol were added and then 20 ml 5N chromic acid and 5 ml concentrated sulphuric acid. This mixture was heated in a closed vial for 90 min at 120°C. The content of the vial was distilled in a Wiesenberger apparatus [6]. The acetic acid was titrated to pH
7.5, the solution concentrated and measured for radioactivity.
4. RESULTS AND DISCUSSION
Table 1 shows the formation of methanol and ethanol under the conditions described in MATERI- ALS AND METHODS. Depending on the mediator between 10 to 136 ffmol methanol were formed under otherwise identical conditions. Whole cells, crude extracts, the 100 000 x g supernatant as well as a combination of such a supernatant and the corresponding sediment showed roughly the same amount of methanol. In the presence of methyl- viologen the time course of methanol formation showed a lag phase of about 1 h. In this time period the amount of formed ethanol and methanol was about the same. About 100 ffmol methanol and 5 ~mol ethanol could be detected after 6 h. In this 5-h period methanol was synthesized at a constant rate of roughly 20 ffmol per h and 400 mg of cells and remained then constant for an additional 12 h (not shown). During an 18-h time period with formate as electron donor under an atmosphere of nitrogen led only to the formation of 10 ffmol methanol. However, the amount of ethanol increased to about 20 ffmol. In the ab- sence of a mediator no methanol could be de- tected. The same was true if boiled cells were used.
The formation of ethanol can be explained by the reduction of acetate to ethanol [1]. Acetate was synthesized by cell-free extracts of the same material of C. thermoaceticum in the absence of a mediator. Recently, we showed that the methyl group of methyltetrahydrofolate is converted via overall retention into acetate [7].
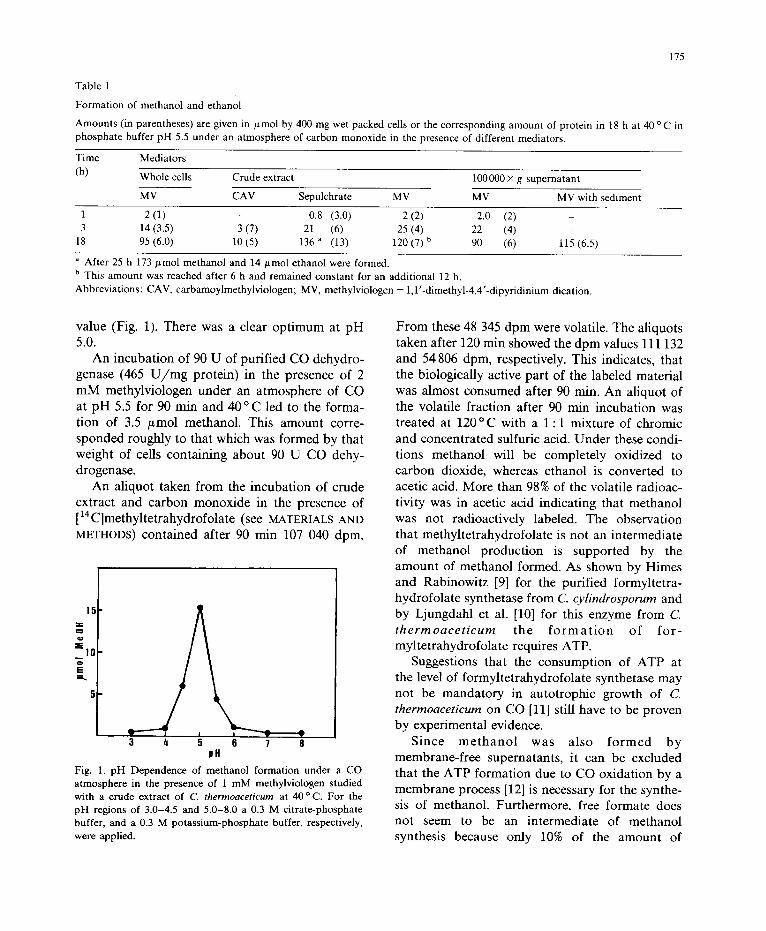
The various viologens behaved differently (Ta- ble 1). The more negative the viologen the more methanol was formed. The redox potential of methylviologen (E o -440 mV) is 145 mV more negative than that of carbamoylmethylviologen ( -295 mV). The redox potential of cobalt sep- ulchrate depends heavily on the pH and ionic strength and may very between -300 to -500 mV. (H. Gi]nther and H.S., unpublished). The methanol synthesis markedly depended on the pH
175
Table 1
Formation of methanol and ethanol
Amounts (in parentheses) are given in ~tmol by 400 mg wet packed cells or the corresponding amount of protein in 18 h at 40 ° C in phosphate buffer pH 5.5 under an atmosphere of carbon monoxide in the presence of different mediators.
Time Mediators
(h) Whole cells Crude extract 100 000 × g supernatant
MV CAV Sepulchrate MV MV MV with sediment
1 2 (1) - 0.8 (3.0) 2 (2) 2 .0 (2) -
3 14 (3.5) 3 (7) 21 (6) 25 (4) 22 (4) - 18 95 (6.0) 10 (5) 136 a (13) 120 (7) b 90 (6) 115 (6.5)
a After 25 h 173/~mol methanol and 14/Lmol ethanol were formed. b This amount was reached after 6 h and remained constant for an additional 12 h. Abbreviations: CAV, carbamoylmethylviologen; MV, methylviologen = 1,1'-dimethyl-4,4'-dipyridinium dication.
value (Fig. 1). There was a clear optimum at pH 5.0.
An incubation of 90 U of purified CO dehydro- genase (465 U / r a g protein) in the presence of 2 mM methylviologen under an atmosphere of CO at pH 5.5 for 90 min and 40 °C led to the forma- tion of 3.5 tzmol methanol. This amount corre- sponded roughly to that which was formed by that weight of cells containing about 90 U CO dehy- drogenase.
An aliquot taken from the incubation of crude extract and carbon monoxide in the presence of [14C]methyltetrahydrofolate (see MATERIALS AND METHODS) contained after 90 min 107 040 dpm.
15
==
o E
3 4 5 6 7 8 pH
Fig. 1. pH Dependence of methanol formation under a CO atmosphere in the presence of 1 m M methylviologen studied with a crude extract of C. thermoaceticum at 40 o C. For the pH regions of 3.0-4.5 and 5.0-8.0 a 0.3 M citrate-phosphate buffer, and a 0.3 M potass ium-phosphate buffer, respectively, were applied.
From these 48 345 dpm were volatile. The aliquots taken after 120 min showed the dpm values 111 132 and 54 806 dpm, respectively. This indicates, that the biologically active part of the labeled material was almost consumed after 90 rain. An aliquot of the volatile fraction after 90 min incubation was treated at 120 °C with a 1 : 1 mixture of chromic and concentrated sulfuric acid. Under these condi- tions methanol will be completely oxidized to carbon dioxide, whereas ethanol is converted to acetic acid. More than 98% of the volatile radioac- tivity was in acetic acid indicating that methanol was not radioactively labeled. The observation that methyltetrahydrofolate is not an intermediate of methanol production is supported by the amount of methanol formed. As shown by Himes and Rabinowitz [9] for the purified formyltetra- hydrofolate synthetase from C. cylindrosporurn and by Ljungdahl et al. [10] for this enzyme from C. thermoacet icum the f o r m a t i o n of fo r - myltetrahydrofolate requires ATP.
Suggestions that the consumption of ATP at the level of formyltetrahydrofolate synthetase may not be mandatory in autotrophic growth of C. thermoaceticum on CO [11] still have to be proven by experimental evidence.
Since m e t h a n o l was also fo rmed by membrane-free supernatants, it can be excluded that the ATP formation due to CO oxidation by a membrane process [12] is necessary for the synthe- sis of methanol. Furthermore, free formate does not seem to be an intermediate of methanol synthesis because only 10% of the amount of

176
methanol was formed in the presence of 0.3 M formate under an N atmosphere and otherwise identical conditions. With - 16.5 kcal/mol the overall reaction from CO to methanol is suffi- ciently exergonic. (Values of free energies of for- mation are taken from [13].) The increased rate of methanol formation after 1 h as well as the fact that the methanol formation stopped after 6 h may be due to the pH shift observed in such incubations. It dropped in 3 h from pH 5.5 to 4.9, and in 18 h to pH 4.2. The pH drop was probably caused by the rather large amount of formic acid (100 pmol in 18 h) which was formed most likely from carbon dioxide by reduction, catalyzed by formate dehydrogenase also present in this organism.
ACKNOWLEDGEMENTS
This work was supported by Deutsche For- schungsgemeinschaft (SFB 145) and Fonds der Chemischen Industrie. We thank Prof. H.G. Floss and Dr. G. Diekert for invaluable discussions and Mr. H. Leichmann for skilled technical assistance.
REFERENCES
[ll
121
[31
[41
[51
[61
[71
[81
[91
[lOI
[Ill
U21
1131
Simon, H., White, H., Lebertz, H. and Thanos I. (1987)
Angew. Chem. accepted.
Gunther, H., Neumann, S. and Simon, H. (1987) J. Bio-
technol. 5, 53-65.
Ljungdahl, L.G. and Andreesen, J.R. (1978) Methods
Enzymol. 53, 360-372.
Drake, H.L., Hu, S.-I. and Wood, H.G. (1980) J. Biol.
Chem. 255, 7174-7180.
Diekert, G. and Ritter, M. (1983) FEBS Lett. 151, 41-44. Wiesenberger, E. (1947) Mikrochem. Verein. Mikrochim.
Acta 33, 51-69. Lebertz, H., Simon, H. Courtney, L.F., Benkovic, S.J.,
Zydowsky, L.D., Lee, K. and Floss, H.G. (1987) J. Am.
Chem. Sot. accepted.
Simon, H. and Floss, H.G. (1967) Anwendung van Iso-
topen in der organ&hen Chemie und Biochemie, Bd. I,
pp. 12-14. Springer Verlag, Berlin.
Himes, R.H. and Rabinowitz, J.C. (1962) J. Biol. Chem.
237, 2915-2925.
Ljungdahl, L., Brewer, J.M. Neece, S.H. and Fairwell, T.
(1970) J. Biol. Chem. 245, 4791-4797.
Kellum, R. and Drake, H.L. (1984) J. Bacterial. 160,
466-469.
Diekert, G., Schrader, E. and Harder, W. (1986) Arch.
Microbial. 144, 386-392.
Barin, I. and Knacke, 0. (1973) Thermochemical Proper-
ties of Inorganic Substances. Springer Verlag, Berlin.





























